
Abstract
The first fossil harvestmen (Arachnida: Opiliones) from the Messel Pit in Hesse, Germany, are described as ?Leiobunum messelense sp. nov. and ?Leiobunum schaali sp. nov. This is the first formal description of fossil harvestmen from the Eocene (ca. 47 Ma) of Messel, and one of the few non-amber records of harvestmen in the Cenozoic. Remarkably, these new specimens show iridescent, metallic colouration. This was previously unknown in the arachnid fossil record, but is seen often in insect fossils from Messel. The new harvestmen exhibit the typical ‘daddy long legs’ morphotype, with small, rounded bodies and long, slender legs. In detail, most of the dorsal opisthosomal tergites appear to be fused together into a single large scute, or scutum parvum, which strongly suggests the modern family Sclerosomatidae, especially in combination with a pectinate pedipalp claw. The iridescent colouration further implies that these fossils can be referred to either the subfamilies Gagrellinae or Leiobuninae, some of which also have a characteristic metallic lustre. Iridescence of the body is more typical for tropical sclerosomatids and is not known from extant European sclerosomatids, with the exception of a single, probably invasive, Leiobunum species. The warmer temperatures during the Eocene might have allowed exchange of early iridescent sclerosomatids between Eurasia and North America, which led to their wide present day distribution and diversity. A few not very well preserved harvestman fossils from Messel are recorded and/or figured for completeness as Leiobuninae / Gagrellinae indet. and Opiliones indet.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Harvestmen (Arachnida: Opiliones) are one of the more diverse lineages of arachnids, with more than 6,700 living species (Kury et al. 2023). From the fossil record, about sixty species are known, the oldest of which goes back more than 410 million years to the Early Devonian (Dunlop et al. 2003). Most of the known fossils come from the Cenozoic, especially the Baltic, Bitterfeld and Rovno ambers of Europe (e.g. Dunlop and Mitov 2009; Elsaka et al. 2019; Mitov et al. 2021). These deposits probably have all an Eocene age (Priabonian: ca. 34-37 Ma), although debate about the age of the Bitterfeld amber remains. There are also a few species of laniatorean harvestmen known from the Miocene Dominican Republic amber (e.g. Cokendolpher and Poinar 1998). By contrast, non-amber records of Cenozoic harvestmen are rare and not always convincing. Based on published figures, the putative harvestmen from the Eocene of Geiseltal in Germany (Haupt 1956), the Oligocene of Piatra Neamț in Romania (Ciobanu 1977) and the Oligocene of Aix-en-Provence in France (Gourret 1887) are probably misidentifications. The only unequivocal Cenozoic harvestmen preserved in shales are therefore those from the Eocene (Priabonian: ca. 34 Ma) Florissant Formation in Colorado, USA, which were revised by Cokendolpher and Cokendolpher (1982), and a species from the Miocene (Burdigalian: ca. 15–20 Ma) Rubielos de Mora in north-eastern Spain (Palencia et al. 2019). Non-amber records from the Mesozoic are similarly sparse, but two specimens belonging to the family Sclerosomatidae have been described from the Jurassic of China (Huang et al. 2009; Giribet et al. 2012). Mesobunus dunlopi Giribet et al., 2012 is the oldest unequivocal member of the Gagrellinae / Leiobuninae group based on the preserved sacular winglets of the male genitalia, a character still found in extant specimens. The second species from the same deposit and genus, Mesobunus martensi Huang et al., 2009, also showed potential Gagrellinae-like affinities in form of pseudoarticulation on the leg femora.
The Messel Pit near Darmstadt in Hesse, Germany is a Fossillagerstätte recognised as an UNESCO world heritage site. It is best known for yielding a range of, often spectacular, vertebrate fossils from upper lower to middle Eocene (ca. 48-47 Ma) subtropical to tropical forest environment. For an overview see, e.g. Smith et al. (2018). Perhaps less well known, but ecologically probably even more important, is the invertebrate fauna of Messel. Insects have been documented in some detail (e.g. Wedmann 2018; Wedmann et al. 2021a, b; Abels and Wedmann 2022) and many different orders are present. Arachnids are also known from Messel, but only in very few numbers compared to the insects. Spiders have been reported by Wunderlich (1986) and by Selden and Wappler (2019, and references therein). Selden and Wappler (2019) formally described a putative member of the family Cybaeidae, but noted that many of the spider fossils are not particularly well-preserved which may make assigning them to families challenging.
Here, we augment the arachnid fauna by describing the first fossil harvestmen from the Messel Pit. Harvestmen were for the first time reported from Messel by Wedmann (2018), but were not described formally. The small-bodied, long-legged fossils are modern in appearance and can be assigned with some confidence to the living family Sclerosomatidae based on the presence of fused opisthosomal tergites and pectinate pedipalpal claws. These Messel fossils represent the oldest harvestmen from Germany. Of particular interest is the fact that several of them show evidence of metallic iridescence in the cuticle: the first occurrence of this phenomenon in a fossil arachnid. The rather robust opisthosomal sclerotisation and the often shiny, metallic appearance are strongly reminiscent of some modern sclerosomatids in the largely tropical subfamily Gagrellinae and the primarily Holarctic subfamily Leiobuninae (Fig. 1).

Examples of extant iridescent sclerosomatid harvestmen. a Pseudogagrella sp. (Photo courtesy of Nicky Bay; nickybay.com) b Leiobunum viridorsum (Photo courtesy of Marco Zozaya; inaturalist.org). c Leiobunum sp. with prey (Photo courtesy of Hay Wijnhoven). d Pseudogagrella amamiana (Photo courtesy of Ryosuke Kuwahara; inaturalist.org)
Material and Methods
Nine specimens of harvestmen were available for study, all originating from the Messel Pit, near Darmstadt in Hesse, Germany. They are deposited in the collection of Messel fossils of the Senckenberg Forschungsinstitut und Naturmuseum Frankfurt/M, Germany (SF), division Messel Research and Mammalogy. The collection of the invertebrate fossils from Messel is specifically located at the Senckenberg Forschungsstation Grube Messel. The examined specimens have the following repository numbers, and also further stratigraphic information is given:
SF-MeI 00878, found in the year 1982 at digging site 3 (grid square D11); SF-MeI 03415, found in 1990 in grid square E11, in layers 4.5m to 5.16m below local marker horizon alpha; SF-MeI 04143, found in 1992 in grid square D10, in layers 5.31m to 5.7m below local marker horizon alpha; SF-MeI 10391, found in 1997 in grid square G8, in layers 0.3m to 0.7m above local marker horizon alpha; SF-MeI 10994 AB, found in 2002 in grid square G8/9, in layers 0.3m to 0.9m above local marker horizon alpha; SF-MeI 10995, found in 2002 in grid square G8/9, in layers 0.3m to 0.9m above local marker horizon alpha; SF-MeI 18990 AB, found in 2019 in grid square F9-Q13, in layers 1.97m to 2.33m above local marker horizon alpha; SF-MeI 19539, found in 2020 in grid square F9-Q8, in layers 1.97m to 2.32m above local marker horizon alpha; SF-MeI 20021, found in 2021 in grid square F9-Q10, in layers 1.85m to 2.27m above local marker horizon alpha (Schaal and Rabenstein 2012).
All specimens except SF-MeI19539, which is poorly preserved, were photographed using a Leica MZ12.5 stereomicroscope and an Nikon D300 digital camera and compared to the literature (e.g. Roewer 1923 in the first instance). Pencil drawings were made with a Leica M165 C stereomicroscope with a camera lucida attachment and then digitally redrawn following the methods of Coleman (2003) using Adobe Illustrator 2022 and a Wacom Intuos graphic tablet. All measurements in mm. Average values are marked with an asterisk.
Geological and climatic setting of the Messel Pit
The fossils are embedded in organic-rich laminated claystone sediments (commonly called oil shale) which were deposited in a meromictic lake (Goth 1990). Former Lake Messel was a maar lake which was created by phreatomagmatic eruptions (Schulz et al. 2002; Felder and Harms 2004). These eruptions are currently estimated to have occurred 48.06 million years ago (Kaboth-Bahr et al. 2024). Local marker horizon alpha, around which all harvestmen specimens have been found, was deposited about 47.357 million years ago, following the calculations of Kaboth-Bahr et al. (2024).
Detailed reconstructions of the Messel palaeoclimate were inferred by Grein et al. (2011) based on 55 plant families from Messel. The analysed flora suggests a humid subtropical–tropical climate with a mean annual temperature of approximately 22 °C (up to 24 °C) and mean annual precipitation rates up to 2540 mm. Comparable climate conditions can be found today in Mexico, Brazil, Paraguay, Madagascar, Laos and Vietnam a. o. (Grein et al. 2011).
Systematic palaeontology
Order Opiliones Sundevall, 1833
Suborder Eupnoi Hansen and Sørensen, 1904
Family Sclerosomatidae Simon, 1879
Subfamily Gagrellinae Thorell, 1889 or Leiobuninae Banks, 1893
Remarks: Four suborders of harvestmen are conventionally recognised. Based on the habitus of the fossils we can exclude Cyphophthalmi which have short legs, Laniatores which have large, raptorial pedipalps and two claws on legs III/IV, and Dyspnoi in which the pedipalp lacks a claw. This leaves the suborder Eupnoi which comprises five families. Groups like Caddidae, which are tiny and have proportionally massive eyes, are easily excluded, just like Neopilionidae, which are known for their enormous chelicerae, at least in some males. Key features of the new fossils are most of the tergites covering the dorsal side of the opisthosoma being fused into a single well-defined plate (the scutum parvum) and the pedipalp claw (where preserved) being pectinate. These characters strongly support referral of the Messel fossils to the family Sclerosomatidae (cf. Tourinho 2007). Other eupnoid families, such as Phalangiidae, tend to have a softer and more leathery dorsal opisthosoma.
Following Kury (2000), four subfamilies of modern sclerosomatids can be recognised, Leiobuninae, Sclerosomatinae, Gyinae and Gagrellinae, although Hedin et al. (2012) subsequently transferred Gyinae to the family Phalangiidae. Explicit diagnostic characters for these sclerosomatid subgroups are hard to find; see e.g. comments in Tourinho (2007) and Taylor (2009). We can probably exclude the Messel fossils from Sclerosomatinae based on the absence of strongly armed bodies (often with spines) and short legs. Nevertheless, at times these subfamilies seem to grade into one another. This is particularly true for the two remaining subfamilies, Leiobuninae and Gagrellinae, both of which could potentially encompass the Messel fossils. For example, Martens (1973, p. 211) mentioned Asian gagrellines with genitalia reminiscent of leiobunines. The problematic relationships between these subfamilies was also discussed by Giribet et al. (2012) and Hedin et al. (2012). In particular, the phylogenetic study of Hedin et al. (2012) recovered most subfamilies of sclerosomatids as poly- or paraphyletic. Genital characters are, unfortunately, not available in the Messel fossils and so we are forced to rely on somatic characters for any further systematic referrals. As noted by Taylor (2009), early studies by Roewer (1910, 1923) proposed that Gagrellinae could be diagnosed on the presence of pseudoarticulations called nodules on the femora of the legs, and by an opisthosoma with a strongly sclerotised scutum parvum. This scute is clearly present in the new fossils. These characters were also accepted by Crawford (1992), although their reliability as unique autapomorphies of Gagrellinae remains questionable. Femoral nodules are also known from some species currently placed in Leiobuninae (Tourinho 2007).
The shiny bodies of some of the Messel fossils are reminiscent of several modern harvestman species belonging to Gagrellinae (e.g. Fig. 1a, d). This subfamily is currently represented by more than 1000 living species in 127 genera (Taylor 2009). The modern fauna is distributed across the Indo-Malayan region, down into Papua New Guinea and northern Australia, as well as into the Neotropics. They are apparently absent today from Africa and Europe. We should note that many modern gagrellines have a prominent spine (or spines) on the dorsal opisthosomal scute, but we see no evidence for spines in the Messel fossils. Roewer (1954) suggested that at least the Asian gagrellines could be divided into two subgroups: Gagrelleae (with spines) and Zalepteae (without spines) but, as noted by Taylor (2009), this distinction may be unreliable. For example, Martens (1987) described a genus which could be well established based on genital characters, but in which species both with and without spines exist.
Iridescence of the body is, however, not unique to Gagrellinae and has also been observed in some living members of Leiobuninae (Goodnight and Goodnight 1942; Nakatsuji 1942); see also Discussion. This character seems to be more typical for species from warmer regions of the world, but it is also known from Central Europe for a probably invasive Leiobunum species. This unidentified species was documented in detail by Wijnhoven et al. (2007), it is characterised by a „metallic green shimmer” which is reminiscent of the Messel fossils.
In summary, the combination of a sclerotised scutum parvum, the presence of iridescent colouration and the absence of spination, pedipalp apophyses (where preserved) and pseudoarticles on the legs in the Messel fossils tend to favour placement in the subfamily Leiobuninae. However, affinities to Gagrellinae cannot be completely excluded, due to the character combinations which overlap between these two subfamilies. If our tentative assignment to Leiobuninae is correct, we can further consider whether the fossils match any of the extant genera. From the European genera Amilenoides Martens and Wijnhoven, 2022, Leiobunum C. L. Koch, 1839, Nelima Roewer, 1910, Leiolima Prieto and Wijnhoven, 2020, Micronelima Schenkel, 1938 and Cosmobunus Simon, 1879 we can exclude Amilenoides, Micronelima and Nelima, due to their smooth leg coxae, whereas at least SF-MeI 00878, SF-MeI 04143 and SF-MeI 10995 bear tuberculated leg coxae. Cosmobunus can also be excluded, as these usually have a smooth pedipalp claw and transverse rows of granules on the opisthosomal sternites, not present in any of the here described Messel fossils. Leiolima specimens have somewhat shorter legs, where femur I is less than two times longer than the prosomal width (measured between coxae II and III). All described Messel fossils surpass this ratio.
Additional genera within Leiobuninae from North America and Asia like Dilophiocara Redikortzev, 1931, Eumesosoma Cokendolpher, 1980, Hadrobunus Banks, 1900, Microliobunum Roewer, 1912, Leuronychus Banks, 1900, Eusclera Roewer, 1910, Paranelima Caporiacco, 1938, Togwoteeus Roewer, 1952 and Schenkeliobunum Starega, 1964 can be ruled out based on the absence of a raised area in front of the ocularium (Dilophiocara, Eumesosoma and Togwoteeus), short legs (Hadrobunus and Microliobunum), a smooth pedipalp claw (Leuronychus), a smooth ocularium (Eusclera), a smooth leg coxa (Paranelima) and rows of four cone-shaped tubercles on the dorsal tergites (Schenkeliobunum) respectively. We can also exclude affinities to the extinct Eocene genus Amauropilio Mello-Leitão, 1937, as both known fossils are much larger with relatively short legs and a smooth pedipalp claw (Cokendolpher and Cokendolpher 1982).
Based on the preserved characters, at least two of the Messel fossils best match the extant genus Leiobunum. In particular the long legs, rows of tubercles on the coxae, the pectinate pedipalp claw and the iridescence of the dorsal body. Nevertheless, it should be noted that most extant European Leiobunum specimens are less sclerotized with a smooth ocularium according to Martens (1978), although more sclerotized specimens can also be found within the genus such as the invasive Leiobunum sp. (Wijnhoven et al. 2007). Most of the fossils characters seem to be more consistent with the genus Leiobunum and thus two of the fossils are provisionally placed as ?Leiobunum Koch, 1839, due to the incomplete preservation and missing genitalia. Further remarks and comparisons to fossil and extant specimens are given below.
Genus ?Leiobunum C. L. Koch, 1839
Type species: Leiobunum rotundum Latreille, 1798
Diagnosis: See Martens (1978) for a diagnosis of the genus.
Remarks: The Messel fossils are best compared with Leiobunum longipes Menge, 1854 described from late Eocene Baltic (Dunlop 2006) and Eocene–Oligocene Bitterfeld amber (Dunlop and Mitov 2009). However, the amber species has been described as having a „slightly granular ornament" on the opisthosoma and a smooth ocularium (Roewer 1939; Dunlop and Mitov 2009), which clearly distinguishes it from the Messel fossils. Additionally, the amber species does not show any signs of iridescence, but this character may be unreliable when used in comparison to amber fossils as fossilised resin seems to not preserve the original colouration.
From the living Leiobunum species the invasive Leiobunum sp. looks quite similar to the Messel fossils, in particular due to its greenish-golden iridescence of the dorsal body, but the ornament seems to be stronger developed in the fossils (Wijnhoven et al. 2007). Since SF-MeI 00878 and SF-MeI 03415 can easily be distinguished from the known sclerosomatid fossils, two new fossil species are described.
Species ?Leiobunum messelense sp. nov.
urn:lsid:zoobank.org:act:60D28B29-BD95-4509-A38E-64BA1A64BD6A
(Fig. 2a–d)

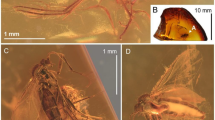
?Leiobunum messelense sp. nov., holotype: SF-MeI 00878. a Photographic overview of the complete specimen, body marked with arrow. b Photographic close-up of the body in lateral view, iridescent parts marked with arrows. c Camera lucida drawing of the body and basal parts of the legs. d Photo of specimen in lateral view, inset shows claw of the left pedipalp. Abbreviations: ap apophysis; cl claw; oc ocularium; pp pedipalp; ta tarsus; legs numbered from I–IV. Scale bars equal 10 mm (a), 1 mm (b, c, d) and 0.1 mm (d inset)
Holotype: SF-MeI 00878, unknown gender. Lateral aspect of body with mostly complete legs.
Type locality: Messel Pit (latitude 49°55’N, longitude 8°45’E) near Darmstadt, Hesse, Germany. The holotype specimen was found in grid square D11 as given in the map of the Messel Pit fossil site by Schaal and Rabenstein (2012). Precise sediment layer not known.
Derivation of name: Named after the Messel Pit.
Diagnosis: Body oval and dorsally ornamented with numerous small tubercles. At least opisthosoma with golden iridescence. Ocularium armed with few small tubercles. Pedipalp claw pectinate with reduced number of teeth. Leg femur I longer than body. Coxae with marginal rows of tubercles.
Can be distinguished from Leiobunum longipes Menge, 1854 from Baltic and Bitterfeld amber based on the stronger developed ornament on the body and ocularium.
Description: SF-MeI 00878. Body oval in lateral view. Dorsal segments fused into scutum parvum. Second prosomal tergite present between anterior carapace and opisthosoma. Total body length 3.04, maximum preserved height 0.95. Dorsal surface generally (both pro- and opisthosoma) ornamented with numerous small tubercles and opisthosomal scute with golden iridescence (Fig. 2b). Ocularium (L 0.32, H 0.26) somewhat removed from the anterior border (by ca. the length of the ocularium), of moderate size and with a few small tubercles (Fig. 2b, c). Eye lens, L 0.14, H 0.14. Chelicerae equivocal. Pedipalps presumably very long but incomplete. Pedipalp tarsus elongated and with a few sensory setae on its tip. Patella possibly with inner club-shaped apophysis, structure could alternatively represent the outline of the proximal cheliceral segment. Pedipalp claw pectinate with row of four very small teeth (Fig. 2d inset). Pedipalp length: fe ?, pa 0.30, ti 0.34, ta 0.82. Legs extremely long (leg II longest). Femur I longer than body. Femur II and III somewhat granulated. Leg coxae and trochanter covered with smaller tubercles (Fig. 2b, c). Tuberculate condition on coxa I equivocal. Coxa II and IV with a short posterior row of tubercles. Coxa III with an anterior and posterior? row of tubercles. Leg length: Leg I cx 0.48, tr 0.30, fe 4.60, pa 0.89, ti 3.89, mt+ta 13.00, total (cx–ta) 23.16; Leg II cx 0.58, tr 0.28, fe 9.78, pa 0.89, ti+mt+ta 43.11, total (cx–ta) 54.64; Leg III cx 0.72, tr 0.24, fe 5.33, pa 0.78, ti 3.78, mt+ta 11.34, total (cx–ta) 22.19; Leg IV cx 0.92, tr 0.18, fe 6.56, pa 1.33, ti 5.67, mt+ta 17.33, total (cx–ta) 31.99.
Genital operculum mostly obscured. Sternites IX–XII present and with rather straight borders. Anal plate equivocal.
Species ?Leiobunum schaali sp. nov.
urn:lsid:zoobank.org:act:0BE8D435-0FE8-40F4-A166-DB1CD788308B
(Fig. 3a–d)

?Leiobunum schaali sp. nov., holotype: SF-MeI 03415. a Photographic overview of the complete specimen. b Photographic close-up on iridescent parts behind the ocularium. c Camera lucida drawing of d. d Photo of specimen in dorsolateral view. Abbreviations: oc ocularium; pp pedipalp; legs numbered from I–IV. Scale bars equal 10 mm (a), 0.5 mm (b) and 1 mm (c, d)
Holotype: SF-MeI 03415, unknown gender. Dorsolateral aspect of body visible, legs and pedipalps incompletely preserved.
Type locality: Messel Pit (latitude 49°55’N, longitude 8°45’E) near Darmstadt, Hesse, Germany. The holotype specimen was found in grid square E11 as given in the map of the Messel Pit fossil site by Schaal and Rabenstein (2012), in a sediment horizon 4.5m to 5.16m below local marker horizon alpha.
Derivation of name: Named after Stephan Schaal, former head of the department Messelforschung and Mammalogie of the Senckenberg Forschungsinstitut und Naturmuseum Frankfurt.
Diagnosis: Large ocularium and eye lenses, larger than in other fossil and most living leiobunine species of similar size (Fig. 3c, d). Dorsal ornament with a greenish iridescence and interconnected tubercles that appear to form irregular ripples (Fig. 3b).
Description: SF-MeI 03415. Body oval in dorsolateral view. Total body length 3.66, maximum width 2.14. Dorsal surface completely ornamented with small tubercles and interconnected tubercles forming irregular ripples (Fig. 3b). Dorsal body with green iridescence. Ocularium large, L 0.54, W 0.60, and covered with small tubercles and granules (Fig. 3b–d). Eye lens also large, L 0.37, H 0.23. Chelicerae and pedipalps equivocal. Legs very long but fragmentary (Fig. 3a). Leg length: Leg I cx ?, tr 0.51, fe ,7.00 pa 1.00, ti 5.44, mt+ta 15.33, total (tr–ta) 29.28; Leg II cx ?, tr 0.34, fe 10.56, pa 0.89, ti+mt 14.22, ta ?, total (tr–mt) 26.01; Leg III cx ?, tr 0.37, fe 6.11, pa 1.11, ti 5.56, mt+ta 14.78, total (tr–ta) 27.93; Leg IV cx ?, tr 0.23, fe 8.56, pa 1.33, ti 4.67, mt+ta 11.78, total (tr–ta) 26.57.
Ventral details not preserved.
Remarks: SF-MeI 03415 represents a unique specimen, despite its incomplete preservation with missing pedipalps, chelicerae and leg coxae. First of all its ocularium and eye lens are much larger compared to SF-MeI 00878 and SF-MeI 04143 and also to most living leiobunines of similar size (Fig. 3c, d). Additionally, its dorsal ornament differs significantly from SF-MeI 00878 and SF-MeI 04143 in which it not only has a more greenish iridescence but also interconnected tubercles that appear to form irregular ripples (Fig. 3b). We can exclude sexual dimorphism here as in extant leiobunines differences are mostly restricted to colouration and body size (Tourinho 2007). Thus, SF-MeI 03415 is placed in a separate new species.
Leiobuninae Banks, 1893 or Gagrellinae Thorell, 1889 indet.

Leiobuninae or Gagrellinae indet., SF-MeI 04143. a Photographic overview of the complete specimen. b Photographic close-up on the upper body and ocularium, iridescent part arrowed. c Camera lucida drawing of d. d Photo of specimen in dorsolateral view. Abbreviations: ch chelicerae; oc ocularium; pp pedipalp; legs numbered from I–IV. Scale bars equal 10 mm (a), 0.5 mm (b) and 1 mm (c, d)

Leiobuninae or Gagrellinae indet., SF-MeI 10995. a Overview of the body in ventral view. b Camera lucida drawing of a. Abbreviations: ap anal plate; ch chelicerae; go genital opening; pp pedipalp; legs numbered from I–IV. Scale bars equal 1 mm
Material: SF-MeI 04143 (Fig. 4), unknown gender. Lateral aspect of body. Legs almost complete. Pedipalps preserved from tarsus to patella; SF-MeI 10995 (Fig. 5), unknown gender. Ventral aspect of body. Legs mostly complete, except tarsus tips. Pedipalps and chelicerae incomplete.
Remarks: SF-MeI 04143 appears to be somewhat similar to SF-MeI 00878, due to the comparable golden iridescence and dorsal ornament. Nevertheless, the dorsal tubercles are more densely packed in this fossil, than in SF-MeI 00878 and the general habitus also differs in being more circular (Fig. 4b, d). This could be related to a different state of development or sexual dimorphism. For that reason we prefer to place this fossil as Leiobuninae or Gagrellinae indet. for the time being. The same applies to SF-MeI 10995 where only the ventral side is preserved as we cannot unequivocally assign this fossil to one of the other specimens.
Description: SF-MeI 04143. Body circular in lateral view (Fig. 4). Dorsal segments fused into scutum parvum, with three free tergites. Total body length 2.98, maximum width 1.62. Dorsal body almost completely covered with small, densely packed tubercles (Fig. 4b). At least first and second free tergite also ornamented with numerous small tubercles. Surface between free tergites with a golden iridescence (Fig. 4b, d). Ocularium relatively small and removed from the anterior border, L 0.33, W 0.26 (Fig. 4c). Dorsal surface of ocularium with a few small tubercles arranged in rows. Eye lens present, L 0.089, W 0.045. Chelicerae equivocal. Pedipalps rather long but fragmentary. Pedipalp length: fe ?, pa 0.35, ti 0.62, ta 0.93?. Legs very long (leg II longest) but incomplete with missing tarsi. Femur I shorter than body. Coxa I and II with numerous small tubercles. Coxa III and IV with a posterior row of tubercles (near the marginal borders). Coxa IV deformed. Leg length: Leg I cx 0.80, tr 0.33, fe 2.89, pa 0.77, ti 2.56, mt 5.00, ta ?, total (cx–mt) 12.35; Leg II cx 0.77, tr 0.35, fe 5.89, pa 0.88, ti 7.78, mt+ta 11.33, total (cx–ta) 27.00; Leg III cx 0.86, tr ?, fe 3.00, pa 0.61, ti 2.67, mt+ta 6.78, total (cx–ta, without tr) 13.92; Leg IV cx 1.11, tr 0.40, fe 3.89, pa 0.77, ti 3.00, mt 2.22, ta ?, total (cx–mt) 11.39.
Sternites IX–XIV somewhat granulated and with mostly straight borders.
Description: SF-MeI 10995. Body oval, preserved in ventral view (Fig. 5). Body length 2.68, width 1.87. Chelicerae of moderate size. Cheliceral proximal segment L ?, distal segment L 0.36, movable finger L 0.14, fixed finger L 0.18. Pedipalps very long (fe–ta 1.67), without armature and apophyses (Fig 5b). Pedipalp tarsus elongated and longer than tibia. Tarsus claw not preserved. Pedipalp length: fe 0.45, pa 0.34, ti 0.29, ta 0.59. Legs extremely long (leg II longest), approximately up to 10 times the body length. Femora sometimes with small granules. Femur I longer than body. Coxa I and II with anterior and posterior rows of larger tubercles (up to 0.04) and additional cover of irregular tubercles (Fig. 5a, b). Coxa III with a short posterior row of somewhat smaller tubercles. Coxa IV largest, covered mostly with irregular tubercles and with intimated anterior and posterior rows. Leg length: Leg I cx 0.57, tr 0.30, fe 4.22, pa 0.66, ti 3.00, mt+ta 10.22, total (cx–ta) 18.97; Leg II cx 0.66, tr 0.30, fe 8.89, pa 0.66, ti+mt+ta 31.67, total (cx–ta) 42.18; Leg III cx 0.71, tr 0.30, fe 4.00, pa 0.88, ti 4.44, mt+ta 8.89, total (cx–ta) 19.22; Leg IV cx 0.84, tr 0.29, fe 5.67, pa 0.55, ti 5.11, mt 4.56, ta 9.33, total (cx–ta) 26.35.
Genital operculum subrectangular, L 0.84, W 0.48. Sternites IX–XIV with mostly straight borders, except Sternite IX, which is somewhat curved. Anal plate rounded L 0.16, W 0.33.
Opiliones Sundevall, 1833 indet.
(Fig. 6a–d)

Photos of unidentified harvestmen fossils from the Messel Pit. a Opiliones indet., SF-MeI 10994. b Opiliones indet., SF-MeI 20021. c Opiliones indet., SF-MeI 10391. d Opiliones indet. (body marked with arrow), SF-MeI 18990. Scale bars equal 5 mm (a), 1 mm (b, c), 10 mm (d)
Material: SF-MeI 10391 (Fig. 6c); SF-MeI 10994 (Fig. 6a); SF-MeI 18990 (Fig. 6d); SF-MeI 19539 (not figured); SF-MeI 20021(Fig. 6b).
Remarks: Further fossils, which likely represent Opiliones, are known from the Messel Pit. Unfortunately, these cannot be reliably identified due to their preservation. The better preserved specimens are illustrated here for completeness.
Discussion
Perhaps the most interesting aspect of the new Messel harvestmen is their iridescent metallic colouration (Fig. 2b, 3b, 4b). To the best of our knowledge this has not been observed in any other fossil harvestmen (or other arachnid), but has an obvious parallel with the preservation of structural colours in the fossils of insects from Messel. Structural colours are preserved in fossil insects from Messel quite regularly, as has been already noted by Lutz (1990). Structural colours are often documented in diverse groups of Messel beetles, but they have also been found in true bugs, hymenopterans, flies, grasshoppers and dragonflies, i.e. in insect groups where today structural colours also occur (Wedmann 2018). Proof for the originality of the structural colours are records of similar coloured conspecific jewel beetles (Buprestidae) from both Messel and from the fossil site Geiseltal which have very different fossilisation conditions (Hörnschemeyer and Wedmann 1994). Parker and McKensie (2003) found that the structural colours of beetles from Messel are based on multilayer systems, and both beetles and moths were further investigated in detail by McNamara et al. (2011a, b). Especially in the moths, very complex cuticular nanostructures scatter light to produce the characteristic lustre, and here the multilayer reflector is modified to reduce iridescence, and to make the colour visible over a broad range of angles. The yellow-greenish colouration of the moths would have provided an excellent match for a diffuse leafy background, suggesting a primarily defensive, cryptic function while at rest. For the zygaenid moths, a possible additional warning function was suggested during feeding on flowers, which is consistent with the biochemistry of modern zygaenids, which are highly toxic (McNamara et al. 2011a). Similar mechanisms and defensive functions presumably underlie the metallic colouration observed in some harvestmen (including now these Messel fossils), but these phenomena have not been rigorously investigated in modern species, and also not yet in the Messel fossils.
As noted above, metallic colouration has been described in several living sclerosomatids, particularly in Gagrellinae from south-east Asia (e.g. Roewer 1929; Nakatsuji 1942; Suzuki 1967, 1979) and Leiobuninae from Mexico and Europe (Goodnight and Goodnight 1942; Wijnhoven et al. 2007). Numerous internet images of quite striking iridescent harvestmen from these geographical regions are also available online (see e.g. inaturalist.org), albeit sometimes without species-level identification. Of particular interest here is the invasive Leiobunum sp. which has rapidly conquered large parts of Western and Central Europe over the past 20 years and which shows comparable iridescence to the Messel fossils (Wijnhoven et al. 2007; Nedvěd et al. 2011; Wijnhoven 2011b). The origin of this iridescent species probably lies in North Africa or the Middle East and its European expansion apparently started in the Netherlands, where it may have been first introduced via shipping in the early 2000s (Wjinhoven 2011a). Leiobunum sp. mostly occurs in large aggregations of hundreds or even thousands of individuals, rendering them a threat to the local arthropod fauna, especially in combination with their omnivorous diet, short reproduction cycle and their ability of parthenogenesis (Nedvěd et al. 2011; Wjinhoven 2011b). It is interesting to speculate whether the Messel harvestmen also aggregated, and in this context it is worth noting that SF-MeI 10994 and SF-MeI 10995 were discovered close together during excavation, which is very unusual for such a rare group of animals in the Messel Pit.
Biogeography
The discovery of harvestmen at Messel offers a valuable opportunity to draw comparisons with the European amber harvestmen. The forests yielding the amber fauna were probably slightly younger. Baltic amber was traditionally dated to ca. 44–49 Ma, but recent publications have begun to favour a slightly younger date of 34–37 Ma (see e.g. Iakovleva 2017; Sadowski et al. 2017, 2022). The Baltic amber forest has been interpreted as having a warm-temperate and humid palaeoclimate, including a mosaic of habitats, and was thus perhaps less overtly subtropical to tropical than the forests which harboured the Messel fauna (Grein et al. 2011; Sadowski et al. 2017). Baltic and Bitterfeld amber have yielded the sclerosomatid harvestman Leiobunum longipes Menge in Koch and Berendt, 1854. Leiobunum is a Holarctic genus with more than 100 species found today across Europe and North America. The only other amber sclerosomatid is Eumesosoma abdelmawlai Elsaka et al., 2019. This genus occurs in North America today, and it is one of several interesting examples of North American harvestmen being found in European ambers which implies a formerly Holarctic distribution.
The Messel sclerosomatid(s) could also belong to a different subfamily, Gagrellinae, not least by virtue of their robust and iridescent cuticle (see below). Living Gagrellinae are represented by more than a thousand species found in the Indomalayan, Neotropical and Nearctic regions. In this context it is interesting to compare several possible modern equivalents for the Messel environment indicated by Grein et al. (2011, fig. 9) to the distribution of extant harvestmen. Gagrelline harvestmen would not be found in the East African and Madagascan habitats, but the comparative tropical forests in Florida, Central America, Brazil, south-east Asia and north-east Australia would be broadly consistent with the distribution of gagrelline harvestmen today. If the systematic referral to Leiobuninae or Gagrellinae is correct, we may have evidence that an essentially tropical group of harvestmen was present in Central Europe during the Eocene (47 Ma), not long after the Paleocene–Eocene thermal maximum (ca. 55.5 Ma). This would also be expected taking the Boreotropic concept into account, which allowed exchange of early gagrellines and leiobunines like ?Leiobunum messelense and ?Leiobunum schaali between Eurasia and North America through Beringian and North Atlantic terrestrial corridors (Hedin et al. 2012 and references therein). According to Hedin et al. (2012) the warmer early Tertiary temperatures led to multiple radiation events, particularly in the newly developing temperate forests, where leiobunines represent the subtropical / temperate descendants of the tropical gagrellines. This hypothesis would also explain the close relationship of both groups. However, gagrellines do not appear to have survived into the amber-producing forests of north–central Europe a few million years later which have, so far, only yielded Leiobuninae. Declining temperatures possibly led to their subsequent extinction, or forced them to shift further south, perhaps as early as the mid Eocene. Nevertheless, we are aware of multiple undescribed sclerosomatids in Baltic amber, which might reveal additional extinct species in future studies.
Data availability
The material is deposited in the collection of Messel fossils of the Senckenberg Forschungsinstitut und Naturmuseum Frankfurt, Germany (SF), division Messel Research and Mammalogy. The collection of the invertebrate fossils from Messel is specifically located at the Senckenberg Forschungsstation Grube Messel.
References
Abels, J., & Wedmann, S. (2022). A fossil wasp (Hymenoptera: Vespidae: Vespula? hassiaca) from the Eocene of Messel. Palaeontographica Abteilung A, 105–117. https://doi.org/10.1127/pala/2021/0122
Banks, N. (1893). The Phalanginae of the United States. Canadian Entomologist, 25, 205–21.
Banks, N. (1900). New genera and species of American Phalangida. Journal of the New York Entomological Society, 8(3), 199–201.
Caporiacco, L. di. (1938). Aracnidi del Messico, di Guatemala e Honduras Britannico. Atti della Società Italiana di Scienze Naturali, 77(3), 251–282.
Ciobanu, M. (1977). Fauna fosilă din Oligocenul de la Piatra Neamț. Bucuresti: Academiei Republicii socialiste Romania.
Cokendolpher, J. C. (1980). Replacement name for Mesosoma Weed, 1892, with a revision of the genus (Opiliones, Phalangiidae, Leiobuninae). Occasional Papers of The Museum, Texas Tech University, 66, 1–19. https://doi.org/10.5962/bhl.title.156534
Cokendolpher, J. C., & Cokendolpher, J. E. (1982). Reexamination of the Tertiary harvestmen from the Florissant Formation, Colorado (Arachnida: Opiliones: Palpatores). Journal of Paleontology, 56, 1213–1217.
Cokendolpher, J. C., & Poinar Jr., G. O. (1998). A new fossil harvestman from Dominican Republic amber (Opiliones, Samoidae, Hummelinckiolus). Journal of Arachnology, 26, 9–13.
Coleman, C. O. (2003). “Digital inking”: How to make perfect line drawings on computers. Organism, Diversity and Evolution, Electronic Supplement, 14, 1–14. https://doi.org/10.1078/1439-6092-00081
Crawford, R. L. (1992). Catalogue of the genera and type species of the harvestman superfamily Phalangioidea (Arachnida). Burke Museum Contributions in Anthropology and Natural History, 8, 1–60.
Dunlop, J. A. (2006). Baltic amber harvestmen types (Arachnida: Opiliones: Eupnoi and Dyspnoi). Fossil Record, 9(2), 167–182. https://doi.org/10.1002/mmng.200600006
Dunlop, J. A., & Mitov, P. G. (2009). Fossil harvestmen (Arachnida, Opiliones) from Bitterfeld amber. ZooKeys, 16, 347–375. https://doi.org/10.3897/zookeys.16.224
Dunlop, J. A., Anderson, L. I., Kerp, H., & Hass, H. (2003). Preserved organs of Devonian harvestmen. Nature, 425, 916. https://doi.org/10.1038/425916a
Elsaka, M., Mitov, P. G., & Dunlop, J. A. (2019). New fossil harvestmen (Arachnida: Opiliones) in the Hoffeins amber collection. Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen 292(2), 155–169. https://doi.org/10.1127/njgpa/2019/0815
Felder, M., & Harms, F.-J. (2004). Lithologie und genetische Interpretation der vulkano-sedimentären Ablagerungen aus der Grube Messel an Hand der Forschungsbohrung Messel 2001 und weiterer Bohrungen. Courier Forschungsinstitut Senckenberg, 252, 151–203.
Giribet, G., Tourinho, A. L., Shih, C., & Ren, D. (2012). An exquisitely preserved harvestman (Arthropoda, Arachnida, Opiliones) from the Middle Jurassic of China. Organisms Diversity & Evolution, 12, 51–56. https://doi.org/10.1007/s13127-011-0067-x
Goodnight, C. J., & Goodnight, M. L. (1942). New and little known Phalangida from Mexico. American Museum Novitates New York, 1163, 1–16. https://doi.org/10.2307/2421154
Goth, K. (1990). Der Messeler Ölschiefer – ein Algenlaminit. Courier Forschungsinstitut Senckenberg, 131, 1–143.
Gourret, P. (1887). Recherches sur les Arachnides tertiaires d’Aix en Provence. Recueil Zoologique Suisse, 4, 431–496.
Grein, M., Utescher, T., Wilde, V., & Roth-Nebelsick, A. (2011). Reconstruction of the middle Eocene climate of Messel using palaeobotanical data. Neues Jahrbuch für Geologie und Paläontologie-Abhandlungen, 260(3), 305–318. https://doi.org/10.1127/0077-7749/2011/0139
Haupt, H. (1956). Beitrag zu Kenntnis der eozänen Arthropodenfauna des Geiselthals. Nova Acta Leopoldina n.s., 128, 1–90.
Hedin, M., Tsurusaki, N., Macías-Ordóñez, R., & Shultz, J. W. (2012). Molecular systematics of sclerosomatid harvestmen (Opiliones, Phalangioidea, Sclerosomatidae): Geography is better than taxonomy in predicting phylogeny. Molecular phylogenetics and Evolution, 62, 224–236. https://doi.org/10.1016/j.ympev.2011.09.017
Hörnschemeyer, T., & Wedmann, S. (1994). Fossile Prachtkäfer (Coleoptera: Buprestidae: Buprestinae) aus dem Mitteleozän der Grube Messel bei Darmstadt, Teil 1. Courier Forschungsinstitut Senckenberg, 170, 85–136.
Huang, D., Selden, P. A., & Dunlop, J. A. (2009). Harvestmen (Arachnida: Opiliones) from the Middle Jurassic of China. Naturwissenschaften, 96, 955–962. https://doi.org/10.1007/s00114-009-0556-3
Iakovleva, A.I. (2017). Detalization of Eocene dinocyst zonation for Eastern Peritethys. Bulletin of Moscow Society of Naturalists. Geological Series, 92(2), 32–48.
Kaboth-Bahr, S., Schmitt, C., Bauersachs, T., Zeeden, C., Wonik, T., Schandl, J., Lenz, O., Wedmann, S., Vasiliev, I., Mulch, A., Lourens, L., Pross, J., & Bahr, A. (2024). Improved chronostratigraphy for the Messel Formation (Hesse, Germany) provides insight into early to middle Eocene climate variability. Newsletters of Stratigraphy. https://doi.org/10.1127/nos/2024/0799
Koch, C. L. (1839). Übersicht des Arachnidensystems. Zweites Heft. C.H. Zeh, Nürnberg.
Kury, A. B. (2000, onwards). Classification of Opiliones. Retrieved October 23, 2023, from http://www.museunacional.ufrj.br/mndi/Aracnologia/opiliones.html.
Kury, A. B., Mendes, A. C., Cardoso, L., Kury, M. S., Granado, A. de A. Giribet, G., Cruz-López J. A., Longhorn S. J., Medrano, M., Kury, I. S., & Souza-Kury, M. A. (2023). World Catalogue of Opiliones. WCO-Lite version 2.6. Retrieved October 20, 2023, from https://wcolite.com/.
Lutz, H. (1990). Systematische und palökologische Untersuchungen an Insekten aus dem Mittel-Eozän der Grube Messel bei Darmstadt. Courier Forschungsinstitut Senckenberg, 124, 1–165.
Martens, J. (1973). Opiliones aus dem Nepal-Himalaya. II. Phalangiidae und Sclerosomatidae (Arachnida). Senckenbergiana biologica, 54, 181–217.
Martens, J. (1987). Opiliones aus dem Nepal-Himalaya. VI. Gagrellinae (Arachnida: Phalangiidae). Courier Forschungsinstitut Senckenberg, 93, 87–202.
Martens, J., & Wijnhoven, H. (2022). A new ancient leiobunine genus and species from the Russian Northwest Caucasus (Opiliones: Sclerosomatidae). Zoology in the Middle East, 68(4), 359–368. https://doi.org/10.1080/09397140.2022.2121084
McNamara, M. E., Briggs, D. E., Orr, P. J., Wedmann, S., Noh, H., & Cao, H. (2011a). Fossilized biophotonic nanostructures reveal the original colours of 47-million-year-old moths. PLoS Biology, 9, e1001200. https://doi.org/10.1371/journal.pbio.1001200
McNamara, M. E., Briggs, D. E., Orr, P. J., Noh, H., & Cao, H. (2011b). The original colours of fossil beetles. Proceedings of the Royal Society B, 279, 1114–1121. https://doi.org/10.1098/rspb.2011.1677
Mello-Leitão, C. de (1937). Distribution et Phylogénie des Faucheurs Sud-Américains. Proceedings of the 12th International Congress of Zoology, Lisbon, 2(5), 1217–1228.
Menge, A. (1854). Footnotes. In C. L. Koch, & G. C. Berendt (Eds.), Die im Bernstein befindlichen Crustaceen, Myriapoden, Arachniden und Apteren der Vorwelt (1st ed., pp. 102). Berlin: Nicholai.
Mitov, P. G., Perkovsky, E. E., & Dunlop, J. A. (2021). Harvestmen (Arachnida: Opiliones) in Eocene Rovno amber (Ukraine). Zootaxa, 4984, 43–72. https://doi.org/10.11646/zootaxa.4984.1.6
Nakatsuji, K. (1942). On a new species of Opiliones from the island of Amami-Ôsima. Zoological Magazine Tokyo, 54(12), 506–508.
Nedvěd, O., Pekár, S., Bezděčka, P., Líznarová, E., Řezáč, M., Schmitt, M., & Sentenská, L. (2011). Ecology of Arachnida alien to Europe. BioControl, 56, 539–550. https://doi.org/10.1007/s10526-011-9385-3
Palencia, L., Peñalver, E., Prieto, C. E., & Poyato-Ariza, F. J. (2019). First fossil harvestmen (Arachnida: Opiliones) from Spain and notes on the fossil record of Opiliones. Palaeontologia Electronica, 22, 1–18. https://doi.org/10.26879/855
Parker, A. R., & McKenzie, D. R. (2003). The cause of 50 million-year-old colour. Proceedings of the Royal Society of London. Series B: Biological Sciences, 270(suppl_2), S151-S153. https://doi.org/10.1098/rsbl.2003.0055
Prieto, C. E., & Wijnhoven, H. (2020). Leiolima iberica, a new harvestmen genus and species from the Iberian Peninsula (Arachnida, Opiliones, Sclerosomatidae). European Journal of Taxonomy, 609, 1–20. https://doi.org/10.5852/ejt.2020.609
Redikortzev, V. V. (1931). Ein neuer Weberknecht aus Buchara. Zoologischer Anzeiger, 97(1/2), 31–32.
Roewer, C.-F. (1910). Revision der Opiliones Plagiostethi (= Opiliones, Palpatores) I. Teil: Familie der Phalangiidae (Subfamilien: Gagrellini, Leiobunini, Leptobunini). Abhandlungen aus dem Gebiete der Naturwissenschaften, herausgegeben vom Naturwissenschaftlichen Verein in Hamburg, 19, 1–29.
Roewer, C.-F. (1912). Einige neue Gattungen und Arten der Opiliones Palpatores aus den Subfamilien der Gagrellinae und Liobuninae der Familie der Phalangiidae. Archiv für Naturgeschichte, Berlin, Abt. A, Original-Arbeiten, 78(1), 27–59.
Roewer, C.-F. (1923). Die Weberknechte der Erde. Jena: Gustav Fischer.
Roewer, C.-F. (1929). On a collection of Indian Palpatores (Phalangiidae) with a revision of the continental genera and species of the sub-family Gagrellinae Thorell. Records of the Zoological Survey of India, 31(2), 107–159.
Roewer, C.-F. (1939). Opilioniden im Bernstein. Palaeobiologica, 7(1), 1–5.
Roewer, C.-F. (1952). Einige Phalangiiden aus dem Vereinigten Staaten von Nord-Amerika. Zoologischer Anzeiger, Jena, 149(11/12), 267–273.
Roewer, C.-F. (1954). Indoaustralische Gagrellinae (Opiliones, Arachnidae). (Weitere Weberknechte XVIII). 1. Teil. Senckenbergiana biologica, 35, 181–236.
Sadowski, E. -M., Seyfullah, L. J., Schmidt, A. R., & Kunzmann, L. (2017). Conifers of the ‘Baltic amber forest’ and their palaeoecological significance. Stapfia, 106, 1–73.
Sadowski, E. -M., Schmidt, A. R., & Kunzmann, L. (2022). The hyperdiverse conifer flora of the Baltic amber forest. Palaeontographica Abteilung B, 304(1–4), 1–148. https://doi.org/10.1127/palb/2022/0078
Schaal, S. F. K., & Rabenstein, R. (2012). Der Tagebau Messel in Linien und Zahlen. Senckenberg Natur Forschung Museum, 142, 376–377.
Schenkel, E. (1938). Spinnentiere von der iberischen Halbinsel, gesammelt von Prof. Dr. O. Lundblad, 1935. Arkiv för Zoologi, Stockholm, 30(24), 1–29.
Schulz, R., Harms, F.-J., & Felder, M. (2002). Die Forschungsbohrung Messel 2001: Ein Beitrag zur Entschlüsselung der Genese einer Ölschieferlagerstätte. Zeitschrift für Angewandte Geologie, 4, 9–17.
Selden, P., & Wappler, T. (2019). A new spider (Arachnida: Araneae) from the Middle Eocene Messel Maar, Germany. Palaeoentomology, 2, 596–601. https://doi.org/10.11646/palaeoentomology.2.6.10
Simon, E. (1879). Essai d'une classification des Opiliones Mecostethi. Remarques synonymiques et descriptions d'espèces nouvelles. Première partie. Annales de la Société Entomologique de Belgique, 22, 183–241.
Smith, K. T., Schaal, S. F., & Habersetzer, J. (2018). Messel - An ancient Greenhouse Ecosystem. Stuttgart: E. Schweizerbart‘sche Verlagsbuchhandlung.
Starega, W. (1964). Materialien zur Kenntnis der ostasiatischen Weberknechte (Opiliones). I-IV. Annales Zoologici, Warsawa, 22(17), 387–410.
Suzuki, S. (1967). Three species of Opiliones from Nepal. Annotationes Zoologicae Japonenses, 40(2), 115–122.
Suzuki, S. (1979). Metagagrella koyamai n. sp. (Opiliones, Gagrellidae). Annotationes Zoologicae Japonenses, 52, 142–145.
Taylor, C. K. (2009). Revision of the Australian Gagrellinae (Arachnida: Opiliones: Sclerosomatidae), with a description of a new species. Australian Journal of Entomology, 48, 217–222. https://doi.org/10.1111/j.1440-6055.2009.00707.x
Thorell, T. (1889). Viaggio di Leonardi Fea in Birrnania e regione vicine XXI. Aracnidi Artrogastri Birrnani. Annali del Museo civico di Storia naturale di Genova, 27(2), 521–729. https://doi.org/10.5962/bhl.title.10400
Tourinho, A. L. (2007). Sclerosomatidae Simon, 1879. In R. Pinto-da-Rocha, G. Machado, & G. Giribet (Eds.), Harvestmen. The Biology of Opiliones (pp. 128–131). Cambridge: Harvard University Press.
Wedmann, S. (2018). Jewels in the Oil Shale – Insects and Other Invertebrates. In K. T. Smith, S. F. K. Schaal, & J. Habersetzer (Eds.), Messel - An ancient Greenhouse Ecosystem (pp. 63–103). Stuttgart: E. Schweizerbart‘sche Verlagsbuchhandlung.
Wedmann, S., Hörnschemeyer, T., Engel, M. S., Zetter, R., & Grímsson, F. (2021a). The last meal of an Eocene pollen-feeding fly. Current Biology, 31(9), 2020–2026. https://doi.org/10.1016/j.cub.2021.02.025
Wedmann, S., Kment, P., Campos, L. A., & Hörnschemeyer, T. (2021b). Bizarre morphology in extinct Eocene bugs (Heteroptera: Pentatomidae). Royal Society Open Science, 8(12), 211466. https://doi.org/10.1098/rsos.211466
Wijnhoven, H. (2011a). De invasieve hooiwagen Leiobunum sp. A in Nederland (Arachnida: Opiliones). Entomologische Berichten, 71(5),123–129.
Wijnhoven, H. (2011b). Notes on the biology of the unidentified invasive harvestman Leiobunum sp. (Arachnida: Opiliones). Arachnologische Mitteilungen, 41, 17–30. https://doi.org/10.5431/aramit4103
Wijnhoven, H., Schönhofer, A. L., & Martens, J. (2007). An unidentified harvestman Leiobunum sp. alarmingly invading Europe (Arachnida: Opiliones). Arachnologische Mitteilungen, 34, 27–38. https://doi.org/10.5431/aramit3406
Wunderlich, J. (1986). Die ersten Spinnen aus dem Mittel-Eozän der Grube Messel. Senckenbergiana lethaea, 67, 171–176.
Acknowledgements
We thank Uta Kiel, Messel, for technical assistance and for the careful preparation of the invertebrate fossils from Messel. Ricardo Pinto-da-Rocha and Gonzalo Giribet are acknowledged for reviewing this manuscript and their helpful comments. Additionally, we want to thank Nicky Bay, Ryosuke Kuwahara, Marco Zozaya and Hay Wijnhoven for allowing the use of their pictures of extant harvestmen specimens in Figure 1. Also many thanks to the digging teams for their great work.
Funding
Open Access funding enabled and organized by Projekt DEAL.
Author information
Authors and Affiliations
Corresponding authors
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
This article is a contribution to the special issue "Pre-Quaternary maar/volcanogenic lakes as Konservat Lagerstätten—Messel and beyond”
The new taxa are registered in Zoobank under
urn:lsid:zoobank.org:act:60D28B29-BD95-4509-A38E-64BA1A64BD6A
urn:lsid:zoobank.org:act:0BE8D435-0FE8-40F4-A166-DB1CD788308B
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Bartel, C., Dunlop, J.A. & Wedmann, S. Iridescent harvestmen (Arachnida: Opiliones: Sclerosomatidae) from the Eocene of Messel, Germany. Palaeobio Palaeoenv (2024). https://doi.org/10.1007/s12549-024-00607-4
Received:
Revised:
Accepted:
Published:
DOI: https://doi.org/10.1007/s12549-024-00607-4