
Abstract
The collection of fossil Rodentia from the Miocene Tagay fauna comprises Aplodontidae, Mylagaulidae, Sciuridae, Gliridae, Castoridae, Eomyidae and Cricetodontinae. Represented by three taxa the Cricetodontinae dominate the rodent assemblage of the Tagay fauna. The fossils were collected by wet-screening of test samples along the section Tagay-1 at Tagay Bay of Olkhon Island. Here, we report on the medium-sized Democricetodon cf. lindsayi, the very small Democricetodon sp. and the large Gobicricetodon filippovi. The primitive dental pattern of D. cf. lindsayi and G. filippovi suggest archaic members of the genera, and an age around the Early/Middle Miocene transition.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
The Tagay site is located at the north-western part of Olkhon Island in the Tagay Bay of Lake Baikal, where Neogene sediments of the Tagay Formation (Mats et al. 2011) became exposed by coastal erosion. During the field campaign in 2014, an integrated geoscientific research programme was carried out by a team of specialists from Russia, Mongolia and Austria. The coastal Tagay-1 section was extensively studied along the sedimentary sequence of 12-13 m thickness. From bottom (coastline of Lake Baikal) to the top, samples were collected for palaeomagnetic, geochemical, sedimentological and palaeontological investigations. Palaeontological test samples along the Tagay-1 section were wet-screened in order to identify the fossil content of the individual sediment layers. Eight out of seventeen sediment layers yielded small vertebrate and gastropod fossils. The vertebrates comprise fish teeth, different bone fragments of lizards, turtles, and small mammal teeth of eleven families/subfamilies: Erinaceidae, Talpidae, Soricidae, Palaeolagidae, Sciuridae, Aplodontidae, Mylagaulidae, Gliridae, Castoridae, Eomyidae and Cricetodontinae (Daxner-Höck et al. 2022a, 2022b; Erbajeva et al. 2022; Mörs et al. 2022, Voyta et al. 2022; all in this issue). Lower vertebrate remains and Gastropoda are not considered in the taxonomic studies. The present paper provides descriptions of the Cricetodontinae rodent species, that were collected from layers 11, 10, 9, 7 and 5 of the Tagay-1 section during our field campaigne 2014.
Material and methods
The sampling methods along the Tagay-1 section and the wet-screening process at the Baikal Lake in 2014 were documented by Daxner-Höck et al. (2022a and 2022b, this issue). After publication, the mammal fossils will be stored in the collection of the Zoological Institute of the Russian Academy of Sciences (ZIN), St. Petersburg. Some additional teeth, collected by A. Kossler in the 1990s from the same locality, are housed in the collection of the NHMW in Vienna.
SEM images were made by Daniela Gruber [Core Facility Cell Imaging and Ultrastructure Research, University of Vienna - member of the Vienna Life-Science Instruments (VLSI)] by using the Scanning Electron Microscope (SEM JEOL IT 300 LV LaB6) and Sputter coater (JEOL JFC-2300HR). For classification above genus level we follow McKenna and Bell (1998).
To facilitate comparisons, right-side teeth are figured as mirror images (as if they were from the left side), and their figure letters are underlined (e.g. Fig. 2a shows the M1 from the right side). All measurements are given in mm.
Abbreviations
Systematic Palaeontology
Family Muridae Illiger, 1811
Subfamily Cricetodontinae Stehlin and Schaub, 1951
Genus Democricetodon Fahlbusch, 1964
Democricetodon cf. lindsayi Qiu, 1996

Democricetodon cf. lindsayi Qiu, 1996 of the Tagay-1 section (beds 7 and 5; Olkhon Island, Baikal Region, Siberia). All teeth are in occlusal view. a Right M1 (ZIN 106859), layer 7. b Right M1 (ZIN 106860), layer 7. c Right m2 (ZIN 106862), layer 5. d Right maxillary fragment (ZIN 106861), layer 7. (underlined = right, not underlined = left)
Locality, Stratigraphy: Tagay, layers 5 and 7 of the Tagay-1 section, Olkhon Island, Baikal Region, Siberia; Tagay Formation (Mats et al. 2011); Early/Middle Miocene transition.
Material: Three teeth (ZIN 106859, 106860, 106862; Fig. 1a, 1b, 1c) of layers 5 and 7 and part of a maxilla without teeth (ZIN 106861; Fig. 1d) from layer 7 of the Tagay-1 section (collected 2014).
Measurements: see Table 1.
Description: The fragment of a right maxilla (ZIN 106861; Fig. 1d) shows three alveoli of M1. Three more partial alveoli suggest three roots of M2. The crowns of the three isolated molars are extremely low, and so are the cones and crests.
M1 (ZIN 106859-860; Fig. 1a, 1b): The anterocone is wide and bilobed (in occlusal view), however, its anterior surface is not divided by a sulcus nor by a groove. The anterolophule is connected with the lingual part of the anterocone. The labial and lingual anterolophs reach the base of the paracone and protocone, and close the anterosinus and protosinus, respectively. Additionally, a small conule terminates the labial anteroloph of one M1 (ZIN 106860; Fig. 1b). The paracone and metacone are located slightly posterior to the protocone and hypocone. The protolophule is directed backwards. It connects with the entoloph halfway between the protocone and mesoloph. The metalophule is transverse and connects with the posterior part of the hypocone (not with the posteroloph). The mesoloph is transverse and long, dividing the mesosinus into two almost equal parts. The pronounced posteroloph labially connects to the base of the metacone, and closes the narrow posterosinus. The sinus is symmetrical. There are three roots of M1, a wide lingual and two rounded labial roots.
m2 (ZIN 106862; Fig. 1c): The lingual metaconid and entoconid are placed anterior to the labial protoconid and hypoconid. The lingual anterolophid is short, the labial one is longer and pronounced. Accordingly, the anterosinusid is much smaller than the protosinusid. The mesolophid is of medium length. The metalophulid and hypolophulid are anteriorly directed. So is the sinusid. The m2 has two roots, an anterior and a posterior one.
Discussion: The medium sized Democricetodon teeth from Tagay differ by larger tooth sizes and the bilobed anterocone of M1from two very small Asian Democicetodon species, i.e. Democricetodon tongi Qiu, 1996 and Democricetodon sui Maridet et al., 2011. Democricetodon tongi is known from the Early and Middle Miocene; Shanwangian and Tunggurian of China, and from the Early Miocene; biozone D1 of Mongolia. Democricetodon sui is known from the Early Miocene; Xiejian and Shanwangian LMS/A of China, and from the Early Miocene; Xiejian LMS/A; biozone D of Mongolia).
The medium sized Democricetodon-teeth from Tagay are closely comparable with the Asian species Democricetodon lindsayi in size and molar pattern. The shared morphological characters are: the bilobed anteroloph of M1, the closed anterosinus and mesosinus, the long mesoloph, the backwards directed protolophule and the symmetrical sinus.
The type locality of D. lindsayi is Moergen II from the Middle Miocene of Nei Mongol. The stratigraphic range of D. lindsayi is Early to Late Miocene; Shanwangian to Baodean LMS/A of China (Qiu and Li 2016: 614). In the course of the long stratigraphic range some molar characteristics changed with time: e.g. 1. increasing elevation of cusps, 2. the metalophule of M1 is originally transverse and connected to the hypocone, later it turns backwards, connects with the posteroloph, and the posterosius diminishes, and 3. gradual development of protolophule I of M1 and M2 (Qiu and Li 2016: 614).
The primitive characters of M1 from Tagay are: very low crowns, missing protolophule 1, transverse metalophule and narrow distinct posterosinus. These characters suggest an early rather than an advanced evolutionary stage of the Tagay specimens. Presently, we classify the few teeth from Tagay as D. cf. lindsayi.
Democricetodon sp.

Democricetodon sp. of the Tagay-1 section (layer 7) and sample Kossler (Ko), Olkhon Island, Baikal Region, Siberia). All teeth are in occlusal view. a Right M1 (ZIN 106474), layer 7. b Left M2 (NHMW2009/0073/0001), Ko. c Left m1 (NHMW2009/0073/0003), Ko. d Left m3 (NHMW2009/0073/0002), Ko. (underlined = right, not underlined = left)
2013 Democricetodon sp. – Daxner-Höck, Böhme and Kossler: 511–512, Plate 22.1, figs 17–19.
Locality, Stratigraphy: Tagay, layer 7 of the Tagay-1 section, Olkhon Island, Baikal Region, Siberia; Tagay Formation; Early/Middle Miocene transition.
Material: Left M2 (NHMW2009/0073/0001), left m3 (NHMW2009/0073/0002) and left m1 (NHMW2009/0073/0003) from test sample Kossler (collected 1990ies; Kossler 2003; Daxner-Höck, Böhme and Kossler 2013:511–512, Plate 22.1, Figs 17–19) and 1 right M1 (ZIN 106474) from the Tagay-1 section, layer 7 (collected 2014).
Measurements: see Table 2.
Description: In all documented tooth positions the molar crowns are extremely low, and so are cones(ids) and crests.
M1 (Fig. 2a): The anterocone is wide and undivided, but in asymmetrical position labial of the median line. The anterolophule is connected to the lingual part of the anterocone immediately at the connection of the anterocone and the lingual anteroloph. The labial and lingual anterolophs reach towards the base of the paracone and protocone, and close the anterosinus and protosinus, respectively. The paracone and metacone are located more posteriorly than the protocone and hypocone. The protolophule is very weak and double. Its connection with the entoloph is opposite of the protocone. The metalophule turns backwards and connects to the lingual part of the posteroloph. The transverse mesoloph is long, but strongly thinning towards its labial end. The pronounced posteroloph connects to the posterior base of the metacone, and closes the small posterosinus. The entoloph is curved. The sinus is symmetrical and lingually closed by a short lingual cingulum. There are three roots of M1, a wide lingual and two rounded labial roots.
M2 (Fig. 2b): The paracone and metacone are located more posteriorly than the protocone and hypocone. The longer labial and shorter lingual anteroloph extend to the base of the paracone and protocone, and close the anterosinus and protosinus, respectively. The protolophule is double, but the posterior arm is very weak. The connection of protolophule and entoloph is opposite of the protocone. The metalophule turns forwards and connects to the entoloph halfway between mesoloph and hypocone. The mesoloph is of medium length. A small mesostyle is present. The pronounced posteroloph connects to the posterior base of the metacone, and closes the large posterosinus. The entoloph is curved. The sinus is slightly directed backwards. It is lingually closed by a weak cingulum. The M2 has three roots.
m1 (Fig. 2c): The anteroconid is small and simple. The lingual anterolophid is shorter than the labial one. Accordingly, the anterosinusid is much smaller than the protosinusid. The metaconid and entoconid are located slightly anteriorly of the protoconid and hypoconid. The connection of anterolophulid and metalophulid is weak. The hypolophulid is directed forwards. It connects to the ectolophid halfway between mesolophid and hypoconid. The mesolophid is long and very low. The posterolophid attaches to the posterior base of the entoconid and closes the posterosinusid. The entolophid is curved, the sinusid directed forward. M1 has two wide roots, one anterior and one posterior.
m3 (Fig. 2d): The tooth narrows posteriorly. It has three main conids, the metaconid, protoconid and hypoconid. The entoconid is very small and continuous with the hypolophulid. The lingual anterolophid is short, the labial one is longer and more pronounced. The metalophulid is forwards directed and connected to the lingual anterolophid. A short posterior arm of the protoconid is visible. The hypolophulid is short and connected to the ectolophid close to the mesolophid. The mesolophid is transverse, ending lingually in a small mesostylid. The sinusid is directed backwards. No roots of the m3 are preserved.
Discussion: The very small Democricetodon from Tagay was described as Democricetodon sp. (Daxner-Höck et.al. 2013: 511-512, Plate 22.1, figs 17-19) on the basis of three teeth from test sample Kossler (NHMW 2009/0073/0001-0003). A fourth specimen was found in the course of the field campaign in 2014. The Siberian teeth are slightly larger, but do not differ substantially in molar morphology from D. sui and D. tongi. These two species are very rare elements of only a few Chinese and Mongolian assemblages. They are very similar to each other in dental features and size, and it is impossible to classify the Tagay specimens as either D. sui or D. tongi. More material is needed for a definitive determination, but with current data at hand we classify the small specimens as Democricetodon sp.
Genus Gobicricetodon Qiu, 1996
Gobicricetodon filippovi Sen and Erbajeva, 2011

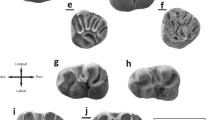
Gobicricetodon filippovi Sen and Erbajeva, 2011 of the Tagay-1 section (layers 10, 9 and 7), Olkhon Island, Baikal Region, Siberia). All teeth except d are in occlusal view. a Right M1 (ZIN 106864), layer 9. b Right M1 (ZIN 106868), layer 7. c Left M1 (ZIN 106870), layer 9. d Right M1, lingual view (ZIN 106863), layer 9. e Left M2 (ZIN 106874), layer 10. f Left M3 (ZIN 106875), layer 9. g Right M3 (ZIN 106877), layer 9. h Left M3 (ZIN 106876), layer 7. (underlined = right, not underlined = left)

Gobicricetodon filippovi Sen and Erbajeva, 2011 of the Tagay-1 section (layers 11, 9 and 7), Olkhon Island, Baikal Region, Siberia). All teeth, except d, are in occlusal view. a Left m1 (ZIN 106885), layer 9. b Right m1 (ZIN 106882), layer 9. c Right m1 (ZIN 106881), layer 7. d Left m1, labial view (ZIN 106884), layer 7. e Left m2 (ZIN 106886), layer 9. f Right m2 (ZIN 106887), layer 9. g Left m3 (ZIN 106891), layer 11. h Left m3 (ZIN 106890), layer 7. (underlined = right, not underlined = left)
1964Cricetodon cf. sansaniensis – Logatchev, N.A., Lomonosova, T.K. and Klimanova, V.M., p. 41.
1997Cricetodon cf. sansaniensis – Erbajeva M.A. and Alexeeva, N.V., p. 242.
2011Cricetodon filippovi – Sen, S. and Erbajeva, M.A. p. 259–270, figs, 2-4.
Locality, Stratigraphy: Tagay, layers 11, 10, 9 and 7 of the Tagay-1 section, Olkhon Island, Baikal Region, Siberia; Tagay Formation; Early/Middle Miocene transition.
Material: 1 left maxillary fragment with M1 (ZIN 106871), 8 M1 right (ZIN 106863–106869; partly fragmentary), 3 left M1 (ZIN 106870,106872–106873; partly fragmentary), 1 left M2 (ZIN 106874), 2 left M3 (ZIN 106875–106876), 4 right M3 (ZIN 106877–106880; partly fragmentary), 3 right m1 (ZIN 106881–106883), 2 left m1 (ZIN 106884–106885), 1 left m2 (ZIN 106886), 2 right m2 (ZIN 106887–106888), a fragment of right m2 (ZIN 106889) and 3 left m3 (ZIN 106890–106892).
Measurements : see Table 3.
Description: The left maxillary fragment shows M1and part of the alveoli of M2. The M1 has three roots, of which the very wide lingual root has a sulcus and split tips.
M1 (Figs 3a, 3b, 3c, 3d): The anterocone is rather narrow, has a labial position and is bilobed. A shallow sulcus or groove at the anterior surface of M1 marks this division, which quickly disappears with wear. The labial anteroloph is a strong crest, that extends from the tip of the labial anterocone in labial direction. At the labial tooth margin, it is continuous with a cingulum that borders the anterosinus and ends at the antero-labial base of the paracone. The lingual anteroloph is shorter, borders the small protosinus and ends at the antero-lingual base of the protocone. Unworn teeth have four high slender cones of equal hight, higher than the anterocone cusps. The position of the labial cones (paracone and metacone) is slightly posterior to the lingual main cones (protocone and hypocone). The protocone has a posterior spur ranging from top to base, that deforms the otherwise symmetrical sinus. Connecting lophs of unworn M1 (anterolophule, entoloph, posteroloph) are lower than the cones. The longitudinally directed anterolophule connects the protocone to the lingual part of the anterocone. The transverse or posteriorly directed protolophule II, present in all M1, extends from the tip of the paracone downwards to meet the entoloph in a right angle, halfway between the protocone and the remnant of the mesoloph. The metalophule is directed posteriorly and extends from the tip of the metacone downwards to connect with the posteroloph. The posteroloph is strong, extends to the base of the metacone and borders the posterosinus. The posterior spur of the paracone is absent in the majority of the M1, only one fragmentary M1 (ZIN 106873) shows a short spur. The mesoloph is absent or very short, in three M1 it is short and connects with the base of the metacone. There is no ectoloph. The entoloph is curved, it becomes very thin or is almost constricted at its contact with the protocone. The narrow sinus turns forwards, it is bordered by a short lingual cingulum in 50% of the specimens. Three out of eleven M1 (partly fragmentary) are completely unworn juvenile teeth without roots. Two M1 have three roots, of which the wide lingual root shows a sulcus, and is split into two tips. The roots of all other specimens are missing or broken.
M2 (Fig. 3e): Only one slightly worn M2 is available. Similar to the M1, the labial anteroloph of M2 is a strong antero-labial crest, that borders the anterosinus and ends at the antero-labial base of the paracone. The lingual anteroloph is very weak and ends at the antero-lingual base of the protocone. No protosinus is visible. The positions of the labial cones are slightly posterior to the lingual ones. Among the four main cones, the metacone is smallest. M2 has a prominent paracone spur and a mesoloph of medium length. The mesoloph turns backwards and meets the metacone almost at its base. The protolophule II and the metalophule are slightly directed backwards. Similar to the M1, the protolophule II starts at the tip of the paracone and meets the entoloph halfway between its connections with the protocone and mesoloph, respectively. The metalophule extends from the tip of the metacone, it is directed posteriorly and connected to the posteroloph. The posteroloph is strong, extends to the tip of the metacone and borders the tiny posterosinus. The sinus is directed forwards. M2 has three roots, two in labial and one in lingual position. The lingual root has a deep sulcus and is split into two tips.
M3 (Figs 3f, 3g, 3h): The third molar resembles M2 in many respects, but differs in the following characteristics: The lingual anteroloph is very weak or absent. The paracone spur is weak or absent. In all specimens, except for one, the mesoloph is long. The metalophule is directed forwards or absent. If present, it is connected to the entoloph anterior to the hypocone. The M3 narrows in its posterior part, and the metacone is either reduced or fused with the metalophule. The hypocone is also strongly reduced. The M3 has three roots, except for one specimen with a split lingual root.
m1 (Figs 4a, 4b, 4c, 4d): The first lower molar is the longest of all lower teeth. It is slender, specifically in its anterior part. The single anteroconid is in line with the longitudinal axis of the tooth. It is lower than the lingual conids, but almost as high as the labial ones. It is accompanied by a lingual and a labial anterolophid. The labial anterolophid extends to the base of the protoconid and borders the protosinusid. The lingual anterolophid is also long, but does not completely close the anterosinusid in all specimens. The four main conids of unworn teeth are very slender and slightly inclined anteriorly. The anterolophulid is straight, connecting the protoconid and the labial part of the anteroconid. The metalophulid is double. Metalophulid I and metalophulid II start at the tip of the metaconid. The metalophulid I is long and connected to the anteroconid in two out of five specimens. In the remaining three, the metalophulid I is shorter and forms a sharp crest at the anterior side of the metaconid but does not reach the anteroconid. The metalophulid II is weak in all five specimens. It is a short crest arising from the tip of the metaconid and pointing posteriorly. In only two specimens it extends as far as the protoconid as a thin crest, or almost so. The hypolophulid is anteriorly directed, connecting to the ectolophid almost opposite to the rudimentary ectomesolophid. The mesolophid is strongly reduced. The ectolophid-hypoconid connection is weak.
The hypoconid has an anterior spur ranging from top to base, constricting the middle part of the sinusid. The sinusid is closed by a labial cingulum. The strong posterolophid joins the base of the entoconid and closes the posterosinusid. There is a sulcus at the posterior side of the tooth, arising from the hypoconid-posterolophid connection. The m1 has two roots.
m2 (Figs 4e, 4f): The description is based on three m2 and the anterior part of a fourth m2. The m2 is shorter, but wider than m1. It is almost equal in width at the anterior and posterior part. The four main conids of unworn teeth are very slender and slightly inclined anteriorly. The lingual anterolophulid is absent, or very weak. The anterosinusid is absent. The labial anterolophid extends to the base of the protoconid and borders the protosinusid. The short metalophulid is directed anteriorly, the hypolophulid is transverse and connected to the ectolophid halfway between the mesolophid and hypoconid. Close to to the hypoconid-connection, the ectolophid is constricted. The mesolophid is short (2) or of medium length (2). The strong posterolophid joins the base of the entoconid and closes the posterosinusid. The sulcus at the posterior side of the tooth is very weak. The sinusid is closed by a short labial cingulum. The m2 has 2 roots.
m3 (Figs 4g, 4h): The third molar is almost equal in length with m2, but narrows posteriorly. The morphology is similar to m2, except for a few differing characteristics: lingual anterolophulid present but short, mesolophid of medium length or long. The m3 has 2 roots.
Discussion: The genus Gobicricetodon with the type species G. flynni Qiu, 1996 was first described from the locality Moergen II in Nei Mongol, China. Six species are known, i.e. G. flynni Qiu, 1996 from Tairum Nur in Nei Mongol (Middle Miocene; Tunggurian LMS/A; MN6) and Moergen II (Middle Miocene; Tunggurian LMS/A; MN7/8), G. aff. flynni from Amuwusu and Balunhalagen in Nei Mongol (Late Miocene; Bahean LMS/A; MN9), G. robustus Qiu, 1996 from Moergen V, Amuwusu, Balunhalagen and Bilutu in Nei Mongol (Middle–Late Miocene; Tunggurian–Baodean LMS/A; MN7/8–MN12), G. arshanensis Qiu and Li, 2016 from Amuwusu, Huitenghe, Bilutu in Nei Mongol (Late Miocene; Bahean–Baodean LMS/A; MN9–MN12), G. natalia Tyutkova, 2008 from Ashut in Kazakhstan (Middle Miocene; MN7/8) and G. filippovi Sen and Erbajeva, 2011 from Aya Cave in Siberia (Middle Miocene; MN6) and Tagay in Siberia (Early/Middle Miocene transition; MN5). Hitherto, the stratigraphic range of Gobicricetodon was Middle to Late Miocene. The new finds of G. filippovi from Tagay suggest an Early Miocene origin of the genus Gobicricetodon (Fig. 5).

The cladogram of Sen and Erbajeva (2011: 267, fig. 5) illustrates the relationships of different Cricetodontinae genera and species of Asia and Europe. There, Gobicricetodon is grouped within Clade 2 (Node 46) together with Plesiodipus Young, 1927, Mixocricetodon Rummel, 1997, Tsaganocricetus Topachevsky and Skorik, 1988 and three species of Cricetodon (C. sansaniensis, C. jotae and C. volkeri).
These taxa more or less share the main characteristic, the “medial diagonal ridge” (Qiu and Li 2016), i.e. the oblique connections of paracone-hypocone and protoconid-entoconid in upper and lower molars, respectively. The cladistic analysis also supports the opinion that Gobicricetodon was derived from an Early Miocene Cricetodon-like ancestral stock, as suggested before by Qiu (1996). A recent revision of “Mixocricetodon” (Europe) confirms the close relationship with the Asian Gobicricetodon, and it shows that “Mixocricetodon” is synonymous with the European genus Lartetomys, known to range from the Early to Middle Miocene (Prieto et al. 2022).
The Gobicricetodon from Tagay shares the general morphology, size, height of molars and root numbers with G. filippovi from Aya Cave. However, the “medial diagonal ridge” of the Tagay specimens – specifically of upper molars – is not well developed. Moreover, the protolophule II of M1 from Tagay connects with the entoloph in a right-angle, the anterocone of M1 is narrower, it has distinct labial and lingual anterolophs, a shorter or absent mesoloph. M2 has a more distinct posterior spur of the paracone and the mesoloph contacts the metacone. The M3 is longer, has a narrowed posterior part and a forwards directed sinus. The labial and lingual anterolophids of m1 are pronounced and the anterosinusid and protosinusid are present, and the metalophulid I reaches the anteroconid only in two out of five m1. These latter characters of G. filippovi from Tagay bear resemblence to the Early Miocene Cricetodon species C. tobieni De Bruijn et al., 1993 from Horlak 1a in Turkey and C. sonidensis Qiu and Li, 2016 from Aoerban U in China (both from the Shanwangian LMS/A, Early Miocene), giving it a primitive appearance. The morphological differences between specimens from Tagay and Aya Cave are interpreted as intraspecific variations rather than species differences.
Hitherto, G. filippovi from Aya Cave (Middle Miocene; MN6) was thought to be oldest and most primitive of all Gobicricetodon species, concerning root numbers, crown hight, size and molar morphology (Qiu and Li 2016; Sen and Erbajeva 2011). However, the obviously more primitive tooth pattern of the Tagay specimens suggests Tagay to be older than Aya Cave (Fig. 5).
Conclusions
Cricetodontinae rodents of the Tagay-1 section (layers 5, 7, 9, 10, 11) and of the collection Kossler (Ko) are the small- to medium-sized Democricetodon cf. lindsayi Qiu, 1996 and Democricetodon sp. and the large-sized Gobicricetodon filippovi Sen and Erbajeva, 2011. Democricetodon cf. lindsayi is represented by three teeth and a maxilla without teeth (layers 5 and 7), Democricetodon sp. by four teeth (layer 7 and coll. Ko) and G. filippovi by 29 teeth and a maxilla without teeth (layers 7, 9, 10, 11).
The medium-sized Democricetodon cf. lindsayi of the Tagay fauna is in good morphological agreement with an early evolutionary stage of D. lindsayi of China. The species is known to range from the Early to the Late Miocene. The small-sized Democricetodon sp. cannot be differentiated from other European and Asian species of its size range, because of the scanty material. The third and large hamster Gobicricetodon filippovi is well represented in Tagay-1. It shares the general molar morphology of the type material from Aya Cave, a Middle Miocene fauna of the Baikal region, however, primitive dental features similar to early Cricetodon species suggest a slightly older age.
Data Availability
All data generated or analysed during this study are included in this published article.
References
Bruijn, H. de, Fahlbusch, V., Sarac, G., & Unay, E. (1993). Early Miocene rodent faunas from the eastern Mediterranean area. Part III. The genera Deperetomys and Cricetodon with a discussion of the evolutionary history of the Cricetodontini. Proceedings of the Koninklijke Nederlandse Akademie van Wetenschappen, 96 (2), 151–216.
Daxner-Höck, G., Böhme, M., & Kossler, A. (2013). 22. New Data on the Miocene Biostratigraphy and Paleoclimatology of Olkhon Island (Lake Baikal, Siberia). In X.-M Wang, L. J. Flynn,. & M. Fortelius (Eds.), Fossil Mammals of Asia. Neogene Biostratigraphy and Chronology (pp. 508–517). New York: Columbia University Press.
Daxner-Höck, G., Mörs, T., Filinov I.A., Shchetnikov A.A., & Erbajeva, M.A. (2022a). Geology and lithology of the Tagay-1 section at Olkhon Island (Lake Baikal, Eastern Siberia), and description of Aplodontidae, Mylagaulidae and Sciuridae (Rodentia, Mammalia). In G. Daxner-Höck, & A. A. Shchetnikov (Eds.), The Miocene Tagay locality of Olkhon Island (Lake Baikal, Eastern Siberia) – a multidisciplinary approach. Palaeobiodiversity and Palaeoenvironments. https://doi.org/10.1007/s12549-022-00548-w. [this issue]
Daxner-Höck, G., Mörs, T., Filinov I.A., Shchetnikov, A.A., Bayarmaa, B., Namzalova, O., & Erbajeva, M.A. (2022b). Gliridae and Eomyidae (Rodentia) of the Miocene Tagay fauna (Olkhon Island, Lake Baikal, Eastern Siberia). In G. Daxner-Höck, & A. A. Shchetnikov (Eds.), The Miocene Tagay locality of Olkhon Island (Lake Baikal, Eastern Siberia) – a multidisciplinary approach. Palaeobiodiversity and Palaeoenvironments. https://doi.org/10.1007/s12549-022-00551-1. [this issue]
Erbajeva, M.A., Daxner-Höck, G., & Mörs, T. (2022). Amphilagus plicadentis (Lagomorpha, Mammalia) from the Tagay locality (Olkhon Island, Baikal region, Eastern Siberia). In G. Daxner-Höck, & A. A. Shchetnikov (Eds.), The Miocene Tagay locality of Olkhon Island (Lake Baikal, Eastern Siberia) – a multidisciplinary approach. Palaeobiodiversity and Palaeoenvironments. https://doi.org/10.1007/s12549-022-00554-y. [this issue]
Erbajeva, M.A., & Alexeeva, N. V. (1997). Neogene Mammalian Sequence of the Eastern Siberia. In J. P. Aguilar, S. Legendre, & J. Michaux (Eds.), Biochronologie mammalienne du Cénozoique en Europe et domaines reliés. Mémoires et Travaux de l’Institut de Montpellier, E.P.H.E., 21, 241–248.
Illiger, J.K.W. (1811). Prodromus systematis mammalium et avium: additis terminis zoographicis utriusque classis, eorumque versione germanica (pp. 301). Berlin: Salfeld.
Kossler, A. (2003). Neogene sediments of Olkhon and Svyatoy Nos (Baikal Rift System, East Siberia): Suggestions about the development of Lake Baikal. Berliner Paläobiologische Abhandlungen, 4, 55–63.
Logachev, N.А., Lomonosova, Т.K., & Klimanova, В.М. (1964). Cenozoic deposits of the Irkutsk Amphitheatre (pp. 1–196). Moscow: Nauka. [in Russian]
Maridet, O., Wu, W.-Y., Ye, J., Bi, S.-D., Ni, X.-J., & Meng, J. (2011). Earliest occurrence of Democricetodon in China, in the early Miocene of the Junggar Basin (Xinjiang), and comparison with the genus Spanocricetodon. Vertebrata PalAsiatica, 49(4), 393–405. [in Chinese with English summary]
Mats, V.D., Shcherbakov, D.Y., & Efimova, I.M. (2011). Late Cretaceous–Cenozoic History of the Lake Baikal Depression and Formation of its Unique Biodiversity. Stratigraphy and Geological Correlation, 19/4, 404–423. Pleiades Publishing Ltd. ISSN 0869-5938.
McKenna, M.C., & Bell, S.K. (1998). Classification of Mammals above the Species Level (pp. 1–631). New York: Columbia University Press.
Mörs, T., Hägglund, S., Erbajeva, M.A., Alexeeva, N., Shchetnikov, A.A., & Daxner-Höck, G. (2022). The northernmost Eurasian Miocene beavers: Euroxenomys (Castoridae, Mammalia) from Olkhon Island, Lake Baikal (Eastern Siberia). In G. Daxner-Höck, & A. A. Shchetnikov (Eds.), The Miocene Tagay locality of Olkhon Island (Lake Baikal, Eastern Siberia) – a multidisciplinary approach. Palaeobiodiversity and Palaeoenvironments. https://doi.org/10.1007/s12549-022-00555-x. [this issue]
Prieto, J., Rummel, M., Scholtz, H., & Mein, P. (2022). A new middle Miocene lineage based on taxonomic revision of the large and rare cricetid-rodent genus Lartetomys. Palaeobiodiversity and Palaeoenvironments, 102(1), 223–236.
Qiu, Z.-D. (1996). Middle Miocene micromammalian fauna from Tunggur, Nei Mongol. (pp.1–216). Beijing. Science Press. [in Chinese with English summary]
Qiu, Z.-D., & Li, Q. (2016). Neogene Rodents from Central Nei Mongol, China. Palaeontologica Sinica, N.S. Series C, 198(30), 1–492. [in Chinese with English summary]
Qiu, Z.-X., Qiu, Z.-D., Deng, T, Li, C.-K., Zhang, Z.-Q., Wang, B.-Y., & Wang, X. (2013). 1. Neogene Land Mammal Stages/Ages of China. Toward the Goal to establish an Asian Land Mammal Stage/Age Scheme. In X. Wang, L. F. Flynn, & M. Fortelius (Eds.), Fossil Mammals of Asia. Neogene Biostratigraphy and Chronology (pp. 29–83). New York: Columbia University Press.
Rummel, M. (1997). Mixocricetodon dehmi n. gen., n. sp., ein neuer Cricetide (Rodentia, Mammalia) aus dem Mittelmiozän von Süddeutschland. Mitteilungen der Bayerischen Staatssammlung für historische Geologie, 37, 75–93.
Sen, S., & Erbajeva, M.A. (2011). A new species of Gobicricetodon Qiu, 1996 (Mammalia, Rodentia, Cricetidae) from the middle Miocene Aya Cave, Lake Baikal. Vertebrata PalAsiatica, 49(3), 257–274. [in English with Chinese summary]
Stehlin, H.G., & Schaub, S. (1951). Die Trigonodontie der simplicidentaten Nager. Schweizerische Paläontologische Abhandlungen, 67, 1–385.
Steininger, F.F. (1999). 1. The Continental European Miocene. Chronostratigraphy, Geochronology and Biochronology of the Miocene „European Land Mammal Mega-Zones“ (ELMMZ) and the Miocene „Mammal-Zones (MN-Zones)“. In G. E. Rössner, & K. Heissig (Eds.), The Miocene Land Mammals of Europe. (pp. 9–24). München: Verlag Dr. Friedrich Pfeil.
Topachevsky, V.A., & Skorik, A.F. 1988. The vole-toothed Cricetodontidae (Rodentia, Cricetidae) from Vallesian of Eurasia and some questions of supergeneric systematics of the subfamily. Vestnik Zoologii, 5, 37–45. [in Russian with English summary]
Tyutkova, L. (2008). The Middle Miocene rodents of the Ashut locality (Turgay Depression). In S. G., Lucas, G. S. Morgan, J. A. Spielmann, et al. (Eds.), Neogene Mammals. New Mexico Museum of Natural History and Science, 44, 437–442.
Voyta, L.L, Mörs, T., & Erbajeva, M.A. (2022). Erinaceomorpha and Soricomorpha (Mammalia) of the Miocene Tagay fauna (Olkhon Island, Lake Baikal, Eastern Siberia): A preliminary report. In G. Daxner-Höck, & A. A. Shchetnikov (Eds.), The Miocene Tagay locality of Olkhon Island (Lake Baikal, Eastern Siberia) – a multidisciplinary approach. Palaeobiodiversity and Palaeoenvironments. https://doi.org/10.1007/s12549-022-00557-9. [this issue]
Young, C.C. (1927). Fossile Nagetiere aus Nord-China. Paleontologica Sinica, Serie C, 5 (3), 1–82.
Acknowledgements
B. Bayarmaa, O. Namzalova, S. Veshcheva, G. Matasova, I. Filinov, A. Shchetnikov, †G. Safronov and G. Sushpanova are acknowledged for their support at the Tagay-1 section, by wet-screening of samples and sorting fossils from residues. The SEM images were made by Daniela Gruber (University of Vienna). We thank the reviewers L. van den Hoek Ostende and S. Sen for careful reading and correcting the manuscript. We also thank the Editors P. Königshof and S. Weber for manifold information and help. All these persons and institutions are gratefully acknowledged.
The fieldwork was supported by grants of the following organisations: Russian Fund of Basic Research (RFBR) Project 12-04-00081_a and RFBR-Siberia Project 12-05-98000-p-Siberia_a to M. A. Erbajeva, and the Austrian Science Fund (FWF) Project P 23061-N19 to G. Daxner-Höck. The research was partly supported via grants frothe RFBR (№ 20-05-00247), the Vinogradov Institute of Geochemistry, SB RAS (project no. 0284-2021-0003), the RSF (project no. 19-17-00216, lithology, stratigraphy and geomorphology), the Government of the Russian Federation (project no. 075-15-2021-631) to A. A. Shchetnikov. We are grateful to the Directorate of the Institute of Paleontology and Geology of the Mongolian Academy of Sciences for the loan of the wet screening equipment.
Funding
Open access funding provided by Swedish Museum of Natural History.
Author information
Authors and Affiliations
Corresponding authors
Ethics declarations
Conflict of Interest
The authors declare that they have no conflict of interest.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
This article is a contribution to the special issue “The Miocene Tagay locality of Olkhon Island (Lake Baikal, Eastern Siberia) – a multidisciplinary approach”
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Daxner-Höck, G., Mörs, T., López-Guerrero, P. et al. Cricetodontinae (Rodentia, Mammalia) of the Miocene Tagay fauna (Olkhon Island, Lake Baikal, Eastern Siberia). Palaeobio Palaeoenv 102, 885–895 (2022). https://doi.org/10.1007/s12549-022-00553-z
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12549-022-00553-z