
Abstract
Abundant shark and rare actinopterygian teeth are reported from a locality within the early Eocene (Ypresian) lower part of the Crescent Formation exposed in the Hamma Hamma River valley on the eastern Olympic Peninsula, Washington State, USA. This part of the Crescent Formation is predominantly submarine volcanic basalt with some sedimentary interbeds deposited in deep water. The teeth are derived from sediments that appear to directly overlay and in places interfinger with the margins of an anomalous lenticular barite deposit; one tooth was found in the barite. Genera represented include deep-water taxa (aff. Chlamydoselachus, Mitsukurina, Notorynchus, Odontaspis) and open marine, epipelagic sharks (Alopias, Brachycarcharias, Jaekelotodus, Macrorhizodus, Otodus, Striatolamia). The only other fossils found were two fragmentary shark vertebrae, numerous shark dermal ossicles, a single teleost tooth (Egertonia) and abundant, minute valves of a discinid brachiopod. This is the first report of macrofossils from the lower part of the Crescent Formation and the only early Eocene shark assemblage described from the North Pacific Basin. The shark assemblage also corroborates paleodepositional interpretations of the lower Crescent Formation as being in part ancient volcanic seamounts during early Eocene time.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Early Eocene fishes are well known from Europe, Africa, and eastern North America (e.g., Kent 1999; Adnet and Cappetta 2008; Adnet et al. 2008; Iserbyt and De Schutter 2012; Padilla et al. 2014; Marramà et al. 2018, 2019; Popov and Lopyrev 2023, and references therein), however, there are surprisingly few reports of fishes from early Eocene strata of the Pacific Basin. They are limited to only a few incidental records and preliminary reports from the Chatham Islands and Australia in the south (Keyes 1987; Kemp 1991) and western North America in the north (e.g., Squires and Goedert 1994; Gonzalez-Barba 2004). Therefore, any additional data on early Eocene fishes from the North Pacific Basin is of significance in understanding distributions of various taxa.
On the Olympic Peninsula in western Washington State, early and middle Eocene molluscan fossils have been recorded from several localities in the uppermost part of the Crescent Formation (e.g., Arnold 1906; Weaver and Palmer 1922; Squires et al. 1992; Squires and Goedert 1994). However, except for foraminiferans (Pardee 1921; Rau 1964), fossils have not been previously reported from the lower part of the Crescent Formation. There are few previous reports of fossil shark teeth from the Crescent Formation. The earliest that we are aware of is Park (1946: p. 308) who stated shark teeth had been found at “three widely separated localities” associated with volcanic rocks, although he did not disclose what part of the Crescent Formation these localities were in, nor did he mention a repository for the specimens. Two shark teeth were recorded by Squires and Goedert (1994) from the upper part of the Crescent Formation on the south side of the Olympic Mountains. Kočí et al. (2022) mentioned shark teeth from early Eocene rocks of the Crescent Formation in the Black Hills near Olympia, and an unidentified shark tooth was reported from the middle Eocene ‘Crescent/McIntosh transition zone’ in the Doty Hills (Squires and Goedert 1995) of southwestern Washington.
The purpose of this paper is to document a previously unknown low diversity fish assemblage based on teeth and dermal ossicles found in the early Eocene lower unit of the Crescent Formation, in marine deep-water sediments associated with a small, anomalous barite deposit.
Geological setting
In western Washington State, the basement strata are the Crescent Formation (Arnold 1906), part of Siletzia, a terrane comprised of submarine volcanic rocks accreted to North America in Eocene time (e.g., Wells et al. 2014; Eddy et al. 2017; Anderson et al. 2024). The Crescent Formation is predominantly submarine tholeiitic basalt, and just to the north of the study area its thickness is as much as 15 km (Cady et al. 1972a), making it one of the thickest exposed sections of submarine volcanic rocks anywhere in the world. The formation has been informally divided into an upper and lower unit (Cady et al. 1972a; Cady 1975; Tabor and Cady 1978); the lower unit is pillow and massive basalt and associated deep water sediments, and the upper unit is pillow basalt with subaerial basalt flows in its upper part. Sediments of the upper unit were deposited in a warm, upper bathyal to upper neritic marine environment based on foraminiferans and mollusks (e.g., Rau 1966; Squires et al. 1992) and preserve wood fragments (Kiel 2008).
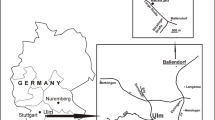
The study area (Fig. 1) is in the Hamma Hamma River valley in the eastern Olympic Peninsula. An anomalous barite deposit (Hughes 1939) is deep within the lower unit of the Crescent Formation and has only been briefly mentioned in subsequent reports, listed as the ‘Maple Creek prospect’ (Park 1942; Green 1945; Moen 1964). It is shown as a manganese prospect on geologic maps (Cady et al. 1972a; Tabor and Cady 1978), even though manganese is apparently present only in trace amounts (Hughes 1939). Although Hughes (1939) refers to the barite deposit as a ‘vein’, it appears to be a lens-like body approximately 4.5 m thick and 46 m long (truncated by faulting and/or erosion at the western end, concealed at the eastern end) within sedimentary and volcanic deposits. The genesis of the barite deposit, speculated by Nisbet and Ambrose (1986) to be an ancient ‘white-smoker’ hydrothermal vent deposit (e.g., sensu Torres et al. 2003) is under investigation (J. Peckmann, pers. comm. 2021). Dips in the study area are steep, in places more than 80 degrees, generally toward the east or southeast (Cady et al. 1972a, b; Tabor and Cady 1978). At the northeastern end of the outcrop, the upper surface of the barite deposit is exposed on a cliff face as smooth spheroidal masses up to 1 m across. This uneven surface is capped by reddish colored mudstone with fish teeth, isolated matrix-supported and well-rounded pebbles, and pebbly sandstone approximately 15 cm thick, with another more indurated 13 cm of pebbly sandstone above that, continuous with sandstone, pebbly sandstone, and argillite with a combined thickness estimated to be 150 m (Hughes 1939). In places these sediments appear to interfinger with parts of the barite deposit. Most of the pebbles in the sandstone are 2–3 mm in their largest dimension although a few rare pebbles were found up to 15 mm across, and one was approximately 25 by 35 mm in diameter. The pebbles are angular to well-rounded. There are also small patchy layers of red mudstone, crystals and pebbles of barite, chlorite, and pyrite. Hematite is abundant in parts of the barite deposit as well as the associated sediments. The sediments directly above the barite preserve several fining-upward cycles and appear to be turbidites, with the coarsest material toward the barite, indicating the base is in the direction of the barite deposit. Exposed on the southwestern end of the outcrop and underlying the barite deposit are an undetermined thickness of basalt capped by manganese-stained, pebbly sediments less than 1 m thick, with some jasper-like material near the contact with barite. No fish teeth were observed in the sediments underlying the barite.

Map of the study area, Olympic Peninsula, Washington State, USA, and generalized stratigraphic position of the Hamma Hamma barite within the nearly 15 km thick Crescent Formation. BMU (Blue Mountain Unit) is comprised of younger rocks thrust below the Crescent Formation (Eddy et al. 2017). Geology modified from Babcock et al. (1992), age data compiled from Squires et al. (1992), Babcock et al. (1994), Wells et al. (2014) and Eddy et al. (2017)
In the process of obtaining samples from the barite deposit, we unexpectedly found fossil fish teeth in sediments directly associated with and stratigraphically above the barite. Contra Nisbet and Ambrose (1986), no fossils have been found in the barite except one poorly preserved shark tooth (Fig. 2). Other than fish teeth, two shark vertebrae and numerous dermal ossicles, the only fossils found so far in the associated sediments are minute valves of a discinid brachiopod. Most of the teeth were found in the basal part of the pebbly sandstone bed in the eastern part of the outcrop. These sediments are obscured by talus and vegetation in the southwestern part of the outcrop. The teeth are mostly small (less than 15 mm height); the largest tooth is approximately 70 mm high. Preservation in the hard, barite-rich matrix is generally poor to good, and while some teeth preserve delicate tips and cusplets, many teeth appear to lack their roots. The roots of the teeth appear in some cases to be replaced by barite making it difficult to distinguish them from the barite-rich matrix. Some of the teeth have chipped or worn apices, but because some teeth preserve delicate cusplets they were likely not transported far, if at all, despite their occurrence in a possible turbidite deposit. Most rock samples containing teeth also yield minute dermal ossicles.

Weathered shark tooth (UWBMVP 124420) found in barite from the Hamma Hamma deposit. Scale bar is 3 mm
The paleoenvironment of the lower unit of the Crescent Formation has been interpreted as open sea, based on planktic foraminiferans, with water depths of probably more than 400 m (Rau 1964, 1966). This is supported by the fish taxa described herein and the presence of minute discinid brachiopods. The lack of wood fragments and other detritus from terrestrial plants supports a depositional environment in deep water, far away from any land mass, however, clasts within the turbidites in the lower Crescent Formation are apparently of continental origin (Cady 1975; Babcock et al. 1992; McCrory and Wilson 2013). Cady (1975) estimated the continental margin to have been approximately 900 km east of its present location during deposition of the Crescent Formation. Phillips et al. (2017) and Ciborowski et al. (2020) hypothesized that Siletzia represents an ancient oceanic plateau, and McCrory and Wilson (2013) suggested the present-day Cocos and Carnegie oceanic ridges as possibly analogous to the paleodepositional setting for the lower Crescent Formation. Some parts of the lower Crescent Formation may be as old as 56 Ma (Babcock et al. 1994) however more recent age estimates for the lower member indicate that these strata are Ypresian, early Eocene, approximately 53–48 Ma (e.g., Wells et al. 2014; Eddy et al. 2017; Ciborowski et al. 2020).
Material and methods
The locality is very difficult to access because the terrain is forested and very steep; Moen (1964) stated that he was unable to find the outcrop. Although only approximately 350 m north of the nearby road, the locality is also 300 m in elevation above the road. Rock samples were broken at the outcrop and those that contained teeth were brought back to the lab. Preparation of individual teeth in the hard matrix was done with a small pneumatic scribe and binocular microscope. A circular saw with a diamond blade was used to trim samples, along with tile nippers and similar hand tools. Some rock samples contain several teeth, and some of these were purposely not trimmed to preserve the original context. Once in the lab, all surfaces of the samples were examined with a binocular microscope and the best results were achieved at 30× magnification to find small teeth and brachiopod fossils. Approximately 210 teeth and numerous placoid scales were collected. Most of the fossils recovered are curated at the Burke Museum of Natural History and Culture, University of Washington, Seattle, Washington 98195 (UWBM—IP and VP are abbreviations for Invertebrate and Vertebrate Paleontology); three microscopic brachiopod fossils and several teeth and dermal ossicles found in processing of bulk rock samples are curated in the Swedish Museum of Natural History, Department of Palaeobiology, 10405 Stockholm, Sweden (NRM PAL). Precise locality details cannot be published herein due to United States rules protecting paleontological resources on federal land, however, this information is available and on file at the UWBM and NRM.
Systematic paleontology
Remarks. We follow here mainly the systematic arrangement of Cappetta (2012) using Neoselachii sensu Compagno (1970) for modern sharks, rays, and skates instead of Elasmobranchii sensu Maisey (2012).
Stone and Shimada (2019) re-examined the phylogeny of extant odontaspidids and demonstrated that this group is not monophyletic and consequently resurrected the family Carchariidae. These authors suggested to restrict Odontaspididae to Odontaspis, while transferring Carcharias to Carchariidae. However, they limited these two families to extant taxa only not providing any information about the exact familial placements of the many fossil odontaspidids. Consequently, we use the systematic framework of Cappetta (2012) for Odontaspididae well knowing that this is rather artificial. A revision of fossil odontaspidids is beyond the scope of this study. We have cited references to taxonomic synonymies that have been recently published and are easily available. Lengthy synonymies for other taxa are outside the scope of this paper.
Class Chondrichthyes Huxley, 1880
Subclass Elasmobranchii Bonaparte, 1838
Cohort Euselachii Hay, 1902
Subcohort Neoselachii Compagno, 1970
Superorder Squalomorphii Compagno, 1973
Order Hexanchiformes de Buen, 1926
Family Chlamydoselachidae Garman, 1884
Genus Chlamydoselachus Garman, 1884
Type species. Chlamydoselachus anguineus Garman 1884, Recent.
Stratigraphic range. Campanian (Late Cretaceous) to Recent.
aff. Chlamydoselachus sp.
Figure 3a–c

Teeth of Hexanchiformes, Squaliformes, and Heterodontiformes from the Hamma Hamma locality, lower unit of the Crescent Formation, early Eocene: a aff. Chlamydoselachus sp., UWBMVP 124381, complete tooth, labial view, with cusplet on the right side partially concealed by overlying tooth; b aff. Chlamydoselachus sp., UWBMVP 124382, isolated cusp with slight posteriorly directed bend near base; c aff. Chlamydoselachus sp., same specimen as b lateral view; d Notorynchus sp., UWBMVP 124383, lower lateral tooth, posterior half in lingual view; e Notorynchus sp., UWBMVP 124383, counterpart of d anterior half of tooth, labial view, with serrate cutting edge visible on right side; f Isistius cf. trituratus, UWBMVP 124384; g Isistius cf. trituratus, UWBMVP 124385; h Isistius cf. trituratus, UWBMVP 124386; i Heterodontus cf. pineti, NRM PAL P19790, lateral tooth. Scale bars equal 2 mm
Material. One tooth, UWBMVP 124381, and one isolated cusp, UWBMVP 124382.
Locality. “Maple Creek prospect”, lower unit of the Crescent Formation, Hamma Hamma River, Mason County, Washington (UWBM VP loc. C3331; UWBM IP loc. B9564).
Description. The incomplete tooth (Fig. 3a) is embedded in hard matrix and somewhat overlain by two other elasmobranch tooth crowns so that only the labial face is partly exposed. The central cusp and both lateral cusplets are preserved, one cusplet partly concealed by an appressed lamniform tooth. The main cusp is rather narrow with almost parallel cutting edges on the exposed portion. The labial face is rather flat transversely and completely smooth. Basally, it is rather stout and seemingly depressed rather than being circular. The lateral cutting edges reach the base of the cusp and extend at least for a short distance on the lateral heel between cusp and cusplets.
The lateral cusplets are well separated and divergent from the main cusp, and sigmoidal in labial view giving the crown a lyre-like appearance. The cutting edges are visible. The root is not preserved.
The second specimen tentatively assigned to aff. Chlamydoselachus sp. (Fig. 3b, c), an isolated cusp, has a smooth lingual face. The cusp is extremely long, thin, only slightly sigmoidal, and slightly bent posteriorly near the base.
Remarks. The dentition of both extant species, Chlamydoselachus anguineus and C. africana (Ebert and Compagno, 2009), is characterized by very conspicuous teeth, which are arranged in distinct files forming a clutching-type dentition sensu Cappetta (2012). The morphological differences in teeth of both extant species are not well established but are seemingly very minor. In the fossil record, conversely, teeth generally assigned to Chlamydoselachus show high degrees of morphological variation. This led Cappetta et al. (2021) to recognize three distinct morphotypes representing three different taxa: Chlamydoselachus, Rolfodon Cappetta et al. 2021, and Dykeius Cappetta et al. 2021. Teeth of late Cretaceous to late Miocene species currently assigned to Rolfodon (Cappetta et al. 2021) most conspicuously differ from those of Chlamydoselachus and Dykeius in the morphology of the root, i.e., a massive root that is longer than broad and has a pair of labio-lingual crests.
Teeth of Dykeius garethi Cappetta et al. 2021, from late Cretaceous rocks of Hornby Island, Canada, are significantly larger than those of any other chlamydoselachid with gracile and high, completely smooth cusps and cusplets, but no accessory cusplets. The root is much broader than long and has two very well-developed, high and labio-lingually oriented lingual crests.
Unfortunately, the two fragmentary teeth from the Crescent Formation lack the root preventing unequivocal assignment to any of the three chlamydoselachid genera. Nevertheless, the combination of slender and completely smooth cusp and cusplets, very divergent and in labial view sigmoidal lateral cusplets suggest a species of Chlamydoselachus rather than Rolfodon or Dykeius. More material is necessary to confirm the genus, and the fossils are too incomplete to assign to any species.
The extant species, Chlamydoselachus anguineus, is very wide-ranging with a patchy distribution ranging from 20 to 1500 m depths, whereas C. africana is only known from the western coast of Africa from depths of 300 to 1400 m (e.g., Ebert and Compagno 2009). Most fossil species of Chlamydoselachus described so far occur in deep-water sediments (e.g., Cigala-Fulgosi et al. 2009; Cappetta et al. 2021; but see Consoli (2008), for a different interpretation). The occurrence of chlamydoselachid teeth in deep-water sediments of the Crescent Formation thus agrees well with the general assumption of chlamydoselachids being benthopelagic deep-sea sharks. Although Chlamydoselachus has been reported from late Eocene rocks of Karaginsky Island in the Bering Sea (Nazarkin and Malyshkina 2020), the tooth reported herein is tentatively the northernmost Paleogene record for the eastern North Pacific Basin.
Family Hexanchidae Gray, 1851
Genus Notorynchus Ayres, 1855
Type species. Notorynchus maculatus Ayres, 1855, Recent.
Stratigraphic range. Ypresian (Eocene) to Recent.
Remarks. Generally, the fossil range of Notorynchus is considered to extend back to the early Cretaceous (Hauterivian) (e.g., Cappetta 2012). However, Underwood et al. (2011a) consider most Cretaceous teeth assigned to Notorynchus to belong either to Gladioserratus Underwood et al. 2011a, or another distinct hexanchiform, a view supported by Argyriou et al. (2022).
Notorynchus sp.
Figure 3d, e
Material. One tooth, broken through the central section and preserved on two pieces of rock, UWBMVP 124383.
Locality. “Maple Creek prospect”, lower unit of the Crescent Formation, Hamma Hamma River, Mason County, Washington (UWBM VP loc. C3331; UWBM IP loc. B9564).
Description. The specimen is a lower lateral tooth broken through its central section and now preserved in two pieces of rock. The principal cusp (acrocone) is preserved (Fig. 3e) in one piece of rock, with two following distal cusplets (conules) which decrease in size and posteriorly discernable as impressions of the cusps or cusplets preserved in the other piece of rock. The principal cusp and distal cusplets are rather massive with rounded apices, which are directed distally. The mesial cutting edge of the principal cusp bears marked serrations basally with recurved apices. Remnants of the root are not identifiable. The apices are blunt with the mesial cutting edge being longer than the distal one. One of the preserved cusps is basally broken and exhibits an oval to roundish cross section.
Remarks. Extant hexanchids include three genera (Notorynchus, Heptranchias Rafinesque, 1810, and Hexanchus Rafinesque, 1810) whose interrelationships are still not completely resolved (e.g., Naylor et al. 2012). While Hexanchus is characterized, inter alia, by six gills slits, Heptranchias and Notorynchus have seven gill slits. All three have typical, mesio-distally elongated, labio-lingually compressed and comb-like lower teeth with successively arranged cusps.
The Hamma Hamma specimen, although somewhat incomplete, can be unambiguously identified as hexanchid due to the mesio-distally, consecutively arranged cusps. The distally decreasing size of cusps identify it as either Hexanchus or Notorynchus rather than Heptranchias. The presence of marked, rather strong and seemingly apically recurved basal mesial serrations indicates that this tooth belongs to Notorynchus rather than Hexanchus. Teeth of Notorynchus are less mesio-distally elongated than teeth of Hexanchus, which is, nevertheless, not possible to establish for the the Hamma Hamma specimen. Teeth of early Paleogene species of Notidanodon Cappetta 1975, possess very enlarged mesial cusplets and teeth of the Eocene hexanchid Weltonia Ward, 1979, differ in the very enlarged anterior-most principal cusp.
Teeth of the Eocene hexanchid, Notorynchus serratissimus (Agassiz, 1843) differ from typical teeth of extant Notorynchus because they have a low and mesially rounded root outline in lateral view and very massive cusps with evenly mesial serrations, and Adolfssen and Ward (2015) accordingly suggested to include this species in Gladioserratus, a hexanchid that is very abundant in Cretaceous rocks. Additionally, the principal cusp and first distal cusplet are of equal size in lower lateral teeth of N. serratissimus and differ in this from the Hamma Hamma specimen.
The Hamma Hamma Notorynchus most closely resemble upper Eocene teeth of N. kempi Ward, 1979, from England and Miocene N. primigenius (Agassiz, 1835) from Switzerland in having rather coarse mesial serrations and distally inclined principal cusp and first distal cusplets with a clear size difference between both. Although it is most parsimonious to assume that the specimen from the Hamma Hamma locality represents N. kempi, we refrain from assigning it to any species because it is incomplete.
While teeth of Notorynchus are quite common in Oligocene and Miocene strata of North America (Cappetta 2012), they are very rare in Eocene deposits of North America and mainly known from the Chesapeake region (N. serratissiumus, Ypresian; Kent 1994) and South Carolina (N. cf. kempi, Priabonian; Cicimurri and Knight 2019). Notorynchus borealus (Jordan and Hannibal 1923; originally described as Notidanion boreale) was based on teeth from the late Eocene Quimper Formation in western Washington, however its taxonomic status is somewhat questionable because few specimens are known (Ward 1979). Unidentified species of Notorynchus have also been reported from latest Eocene to Oligocene strata in Oregon and Washington (e.g., Welton 1972; Moore 1984). The presence of Notorynchus in the Hamma Hamma locality is the earliest record from the Pacific Coast of North America.
Order Squaliformes Goodrich, 1909
Family Dalatiidae Gill, 1893
Remarks. Although Gray (1851) is generally considered the author of Dalatiidae, he actually used ‘Dalatiana’, which was modified by Gill (1893) into Dalatiina, placing it together with his Somniosina into the new family Dalatiidae.
Genus Isistius Gill, 1864
Type species. Scymnus brasiliensis Quoy and Gaimard, 1824, Recent.
Stratigraphic range. Thanetian (late Paleocene) to Recent.
Isistius cf. trituratus (Winkler, 1874b)
Figure 3f–h
Material. Nine isolated tooth crowns; figured specimens UWBMVP 124384, 124385, 124386, and six referred specimens, UWBMVP 124387–124392.
Locality. “Maple Creek prospect”, lower unit of the Crescent Formation, Hamma Hamma River, Mason County, Washington (UWBM VP loc. C3331; UWBM IP loc. B9564).
Description. This species is represented by isolated and slightly damaged tooth crowns, all without any visible remnants of the root, embedded in hard matrix. All are strongly compressed labio-lingually and the shape of the crown is that of an isosceles triangle with smooth cutting edges. The exposed crown face is devoid of any ornamentation and is approximately as wide as high; ranging in size from 1 mm, with the largest being 6 mm. The very triangular shape of the crowns is most similar to that of lower teeth of the dalatiid cookie cutter shark Isistius.
Remarks. Extant species of Isistius are characterized by a strongly heterodont dentition with very small, upper grasping teeth and relatively large, triangular lower cutting-type teeth. The fossil record of Isistius is dominated by lower teeth, probably because upper teeth are more difficult to identify. Up to now, only a single upper tooth of Isistius has been reported, from Eocene deposits of Europe (Adnet, 2006). All fossil lower teeth are generally assigned to two distinct species with different stratigraphic ranges: I. trituratus from the Paleocene to Eocene of Europe, North America and North Africa and I. triangulus (Probst, 1879) from the Miocene to Pliocene of Europe and South America (see Cappetta 2012, and references therein). Morphological differences between both species are feeble, but the teeth of I. trituratus can easily be differentiated from those of I. triangulus by having smooth cutting edges. So far, Isistius has not been reported from Oligocene or Pleistocene deposits (Kriwet and Klug 2009; Cappetta 2012). Although I. trituratus might have been common in eastern North America during the Eocene (e.g., Ward and Wiest 1990; Kent 1999), its exact distribution remains somewhat ambiguous. Welton (1974) reported I. cf. trituratus from Paleocene rocks in California, however the specimens from the Crescent Formation are the first Eocene occurrence of Isistius from the North Pacific Basin. Nevertheless, more and better-preserved material is needed to unambiguously confirm the identity.
Extant and extinct squaliform (dogfish) sharks are a highly diverse group with many taxa occurring in coastal and oceanic, cool temperate, and deep tropical waters worldwide and include some of the largest predators in the deep sea (Gartner et al. 1997; Compagno 1999; Musick et al. 2004). Most squaliforms are benthonic, but mesopelagic forms such as Isistius undertake nightly vertical migrations towards the sea surface in search of food (Martin and Treberg 2002; Adnet et al. 2021). The two extant species, I. brasiliensis (Quoy and Gaimard, 1824) and I. plutodus (Garrick and Springer, 1964), however, display very distinct depth distributions. Isistius brasiliensis inhabits depths of more than 3500 m, while I. plutodus generally occurs between 60 and 200 m water depth but was also reported from depths greater than 6000 m (e.g., Kiraly et al. 2003; Compagno et al. 2005). The occurrence of I. cf. trituratus thus agrees well with the inferred paleodepth of more than 400 m for deposition of the lower unit of the Crescent Formation.
Superorder Galeomorphii Compagno, 1973
Order Heterodontiformes Berg, 1937
Family Heterodontidae Gray, 1851
Genus Heterodontus Blainville, 1816
Type species. Squalus philippi Schneider, 1801, original designation, in Bloch and Schneider (1801) = Heterodontus portusjacksoni (Meyer, 1793). Recent.
Stratigraphic range. Valanginian (Early Cretaceous) to Recent.
Heterodontus cf. pineti Case, 1981
Figure 3i
Material. A single, isolated tooth crown, NRM PAL P19790.
Locality. “Maple Creek prospect”, lower unit of the Crescent Formation, Hamma Hamma River, Mason County, Washington (UWBM VP loc. C3331; UWBM IP loc. B9564).
Description. The specimen is a lateral tooth embedded in rock so that only the crown is exposed. It is poorly preserved with most of its lingual margin missing, while the preserved part is slightly domed and displays faintly rugose ornamentation. Both labial and lingual crown faces are separated by a blunt, mesio-distally directed elevation, which is devoid of any distinct crest or cutting edge. This, however, might relate to the incompleteness of the crown. The apical elevation is displaced labially giving the tooth crown an asymmetric and bulbous appearance.
The lingual crown face is narrower and steeper, forming almost a transversally running depression. It possesses a distinct shallow pit-like ornamentation with pits that are smaller, more circular and densely arranged along the labial margin, whereas they become more sparsely arranged and labio-lingually elongated towards the apical elevation. The lingual ornamentation, as far as preserved, also comprises shallow, but more labio-lingually elongated and more parallel arranged pits.
Remarks. Two distinct dental morphological types can be distinguished in Heterodontus (e.g., Herman et al. 1993; Cappetta 2012; Hovestadt 2018). According to Hovestadt (2018), distinction between the species within a morphogroup is difficult, however, all fossil species that he included in Heterodontus can also be confidently assigned to one of these two groups.
Up to now, seven species have been described and figured from Eocene deposits that are considered valid (Hovestadt 2018). According to Hovestadt (2018), the species, H. eocenicus Tate, 1894, H. pineti Case, 1981, H. wardenensis Casier, 1966, and H. woodwardi Casier, 1946, display dental traits that are characteristic for the Morphotype 1 group, whereas H. elongatus Case and Borodin, 2000, H. sowasheense Case, 1994, and H. vincenti Leriche, 1905, should be assigned to Morphotype 2 group. The shape and ornamentation pattern of the single preserved lateral tooth crown described here associate it with extant members of the Heterodontus portusjacksoni group (Morphotype 1 group). The rather incomplete nature of the specimen, unfortunately, renders a definite identification difficult, but the bulbous crown and distinct ornamentation pattern ("conchoidal" ornamentation pattern of Case (1981)) resemble most closely what is seen in H. pineti. Lateral teeth of H. eocenicus and H. woodwardi are, conversely, flatter and the ornamentation patterns include reticulated costules (the lateral teeth of H. wardenensis are unknown). Similar lateral teeth of Morphotype 2 group likewise are flatter and display a different ornamentation pattern. Nevertheless, more and better-preserved specimens are necessary for an unambiguous taxonomic identification.
Case (1981) erected the species H. pineti for two teeth (one anterior, one lateral) from Priabonian strata in Georgia (USA) that are generally like those of H. lerichei Casier, 1943. While Parmley et al. (2003:158) judged the material to be ambiguous and Müller (1999) assumed that H. pineti might be a junior synonym of H. janefirdae Case, 1980 (most likely from Oligocene rather than Miocene sediments according to Müller 1999), Hovestadt (2018) deemed the morphological differences to be sufficient for identifying a distinct species, H. pineti. We follow this opinion here.
So far, H. pineti was only known from upper Eocene strata of Georgia and Louisiana in southeastern North America (Case 1981; Manning and Standhardt 1986). The new record reported here would expand the stratigraphic range to the early Eocene and the geographic range to northwestern North America.
Extant bullhead sharks (family Heterodontidae) are strictly bottom-dwellers on continental shelves and slopes up to ca. 300 m depth, but mostly associated with reefs and nearshore, shallow-water environments (Compagno 2001; Ebert et al. 2013). The record of the single, damaged tooth in the lower unit of the Crescent Formation likely represents an allochthonous rather than an autochthonous faunal element.
Order Lamniformes Berg, 1937
Remarks. Lamniform sharks represent the most abundant and diverse group of elasmobranchs in the Hamma Hamma assemblage including bathydemersal and epipelagic taxa which are characteristic of an open marine environment with considerable water depths. Most common are isolated tooth crowns that either represent members of carchariid or odontaspidid sandtiger sharks. The material, however, is too fragmentary and still embedded in hard matrix so that distinguishable characters in many cases cannot be identified conclusively for appropriate taxonomic assignments.
Family Mitsukurinidae Jordan, 1898
Genus Mitsukurina Jordan, 1898
Type species. Mitsukurina owstoni Jordan, 1898, Recent.
Stratigraphic range. Ypresian (early Eocene) to Recent.
Mitsukurina cf. maslinensis (Pledge, 1967).
Figure 4a–d

Teeth of Lamniformes (Mitsukurina, Striatolamia, Odontaspis, Palaeohypotodus), from the Hamma Hamma locality, lower unit of the Crescent Formation, early Eocene: a Mitsukurina cf. maslinensis, UWBMVP 124393, tooth crown in lingual view; b M. cf. maslinensis, UWBMVP 124394, tooth crown in labial view, part of tooth broken away showing mold with folds on lingual surface; c M. cf. maslinensis, UWBMVP 124395, base of tooth crown in lingual view; d M. cf. maslinensis, UWBMVP 124396, tooth crown in lingual view, partly concealing tooth of aff. Chlamydoselachus sp. in Fig. 3a; e Striatolamia macrota, UWBMVP 124401, tooth crown, lingual view; f Odontaspis cf. winkleri, UWBMVP 124402, lingual view; g O. cf. winkleri, NRM PAL P19791, lingual view; h O. cf. winkleri, UWBMVP 124403, labial view; i O. cf. winkleri, NRM PAL P19792, lingual view; j Palaeohypotodus sp., UWBMVP 124409, lingual view; k same as j apical view. Scale bars equal 2 mm
Material. Eight incomplete tooth crowns; four figured specimens, UWBMVP 124393–124396, and four referred specimens, UWBMVP 124397–124400.
Locality. “Maple Creek prospect”, lower unit of the Crescent Formation, Hamma Hamma River, Mason County, Washington (UWBM VP loc. C3331; UWBM IP loc. B9564).
Description. Tooth crowns without any remnants of the root or additional lateral cusplets are very small, very slender and slightly sigmoidal in lateral view. The labial face is convex mesio-distally, and devoid of any ornamentation. The lingual face, as far as can be ascertained, except for a thin band on either side next to the cutting edge is covered by dense enameloid folds that originate at the base, where they are approximately parallel and extend for 75 to 80 percent of cusp height. The crown of the most complete specimen (Fig. 4a; UWBMVP 124393) has its apex slightly twisted axially with respect to the base. A few other teeth also possess this slight axial twist toward the apex which makes it possible to identify them even when just the smooth labial face is exposed. The lateral cutting edges are weak and seemingly do not reach the base of the tooth.
Remarks. The extant goblin shark, Mitsukurina owstoni, has a wide distribution in both Atlantic and Pacific oceans, but is apparently absent in the Mediterranean Sea as well as polar regions (Ebert et al. 2021). The fossil record of Mitsukurina includes only two species, M. maslinensis (Pledge, 1967) and M. lineata (Probst, 1879) from the Paleogene of Australia and Neogene of Europe, respectively. Paleogene records are very rare in comparison to Neogene occurrences (e.g., Cappetta 2012, and references therein).
Teeth of M. maslinensis differ from those of M. lineata predominantly in the smaller size and the occasional presence of diminutive lateral cusplets in anterior teeth. The teeth from the Crescent Formation are very incomplete and do not allow observation of taxonomically important characters such as the possible presence of vestigial lateral cusplets. Nevertheless, we identify them as M. cf. maslinensis, representing anterior teeth, pending the discovery of more complete teeth. They differ from teeth of Anomotodon Arambourg, 1952, in lacking the medio-basal crest on the labial face (e.g., Carlsen and Cuny 2014) and stronger striations. More and better-preserved material is necessary to unambiguously verify the taxonomic assignment of the teeth from the Crescent Formation. These are, to our knowledge, the first record of Mitsukurina in Paleogene rocks of the North Pacific Basin.
Family Odontaspididae Müller and Henle, 1839
Genus Striatolamia Glikman, 1964
Type species. Otodus macrotus Agassiz, 1838, Eocene, Paris Basin (without more precise information).
Stratigraphic range. Danian (early Paleocene) to Priabonian (late Eocene).
Remarks. The systematic position of Striatolamia is still debated because of dental similarities to living and extinct carchariid and odontaspidid sand tiger sharks from which they nevertheless can be easily distinguished; Kriwet et al. 2016, provided a review of differentiating characters. Long (1992) and Purdy (1998) for example, dismissed Striatolamia and assumed it to be congeneric with Carcharias Rafinesque, 1810. Siversson (1995), Cappetta and Nolf (2005), and Cappetta (2012) conversely, regarded Striatolamia as a member of Mitsukurinidae, because of similarities to teeth of Anomotodon hermani Siversson 1992. Striatolamia has also been classified as a mitsukurinid recently by others, although mostly without further comment (e.g., Ebersole et al. 2019; Kolvalchuk et al. 2023; Popov and Lopyrev 2023), and still by some as an odontaspidid (e.g., Adnet 2006; Malyshkina 2021; Rodriguez et al. 2023; Verma 2023). We follow the more traditional view here and consider Striatolamia to be a valid taxon within Odontaspididae for reasons provided by Kriwet et al. (2016). A complete review and analysis of the exact taxonomic relationship of Striatolamia is beyond the scope of this paper.
Striatolamia macrota (Agassiz, 1838).
Figure 4e
Material. One fragmentary tooth crown, UWBMVP 124401.
Locality. “Maple Creek prospect”, lower unit of the Crescent Formation, Hamma Hamma River, Mason County, Washington (UWBM VP loc. C3331; UWBM IP loc. B9564).
Description. Small very fragmentary main cusp like those assigned to Mitsukurina cf. maslinensis above but differs in that it is broader and more triangular with lingual enameloid folds that do not reach as far toward the apex. Additionally, the lingual crown face is not regularly convex mesio-distally but is medially flattened. The cutting edges are very prominent.
Remarks. We follow Ebersole et al. (2019) in considering Striatolamia striata (Winkler, 1874a) to be a junior synonym of S. macrota (Agassiz, 1838). Kovalchuk et al. (2023) and Verma (2023) have provided recent, lengthy, and comprehensive synonymies for S. macrota. The presence of short lingual apico-basally oriented folds that do not extend to the apex and the medially flattening of the lingual cusp face allow assignment of this fragmentary tooth to the extinct sandtiger shark Striatolamia macrota.
The dental similarities of Striatolamia with Carcharias, but also with Mitsukurina suggest that Striatolamia probably occupied similar ecological niches. Striatolamia macrota has been reported from early to late Eocene rocks in Washington, Oregon, California, Baja California Sur (Mexico) and Chiapas (Mexico) on the Pacific Coast of North America (e.g., Applegate 1968; Squires et al. 1992; Ferrusquía-Villafranca et al. 1999; Squires 2001). Striatolamia is a common element of pelagic elasmobranch faunas in the Eocene worldwide including very high latitudes (e.g., Welton and Zinsmeister 1980; Padilla et al. 2014) indicating that this shark genus was very successful. The reasons for its disappearance at the end of the Eocene, however, remain unknown.
Genus Odontaspis Agassiz, 1838
Type species. Squalus ferox Risso, 1810, Recent.
Stratigraphic range. Ypresian (early Eocene) to Recent.
Odontaspis cf. winkleri Leriche, 1905
Figure 4f–i
Material. Nine incomplete teeth, some with remnants of the root; four figured specimens, NRM PAL P19791, 19792, UWBMVP 124402, 124403, and five referred specimens, UWBMVP 124404–124408.
Locality. “Maple Creek prospect”, lower unit of the Crescent Formation, Hamma Hamma River, Mason County, Washington (UWBM VP loc. C3331; UWBM IP loc. B9564).
Description. In addition to numerous isolated tooth cusps that most likely represent either carchariids or odontaspidids, some teeth are better preserved with lateral cusplets and remnants of the root. In anterior teeth, the main cusp is rather high and narrow, but flares basally. The preserved lateral cusplets are needle-like and either slightly divergent (in anterior teeth) or upright (in lateral teeth), decreasing in size mesially and distally. The lingual crown face is smooth, devoid of ornamentation. Anterior teeth additionally possess an awl-like, slender main cusp. In some specimens, a labio-basal short vertical ridge is present. The labial face is flat while the lingual face is slightly more convex mesio-distally. The cutting edges do not reach the base of the main cusp. Lateral teeth differ in having more slender and upright main cusps. The root lobes, as far as they are preserved display the typical morphology and shape for odonatspidids.
Remarks. Kovalchuk et al. (2023) provided a lengthy and comprehensive synonymy of this species. The preserved characters do not allow an unambiguous assignment to any species. However, they resemble teeth of Odontaspis winkleri, which was originally described from Lutetian strata of Belgium (Leriche 1905) but also occurs in Paleocene strata (e.g., Ward and Wiest 1990; Reinecke and Engelhard 1997) to some extent. However, typical teeth of O. winkleri differ in having more delicate lateral cusplets with more flared bases, which are much shorter compared to the height of the main cusp (compare Hovestadt and Steurbaut 2023). Odontaspis winkleri is very common in rocks deposited in pelagic environments of the Northern Hemisphere during the Ypresian, including North America, but also is known from the Southern Hemisphere (e.g., Smith et al. 1999; Kriwet 2005a; Rodriguez et al. 2023). Ward and Wiest (1990) reported O. winkleri from Danian and Thanetian (Paleocene) and Ypresian (Eocene) strata of Maryland and Virginia.
Case and Borodin (2000) introduced a new species, O. carolinensis from the Lutetian (middle Eocene) of North Carolina based on four mostly fragmentary teeth that very much resemble those of O. winkleri. Burdon (2009: figs. 16, 17), nevertheless, argued that the morphology of these teeth is different from typical teeth of O. winkleri from Europe and therefore considers O. carolinensis valid. The teeth figured by Burdon (2009) differ from those described here in displaying a more lanceolate main cusp; we agree that O. carolinensis appears to be a valid taxon.
Recently, Pollerspöck and Straube (2017) introduced a new Miocene deep-water shark, Pseudoapristurus, from Germany, characterized by very high and slender, pointed lateral cusplets that are well-separated from the main cusp, which also is high and very slender. However, the teeth of Pseudoapristurus differ from those of O. winkleri in possessing a very characteristic baso-lingual, reticulated ornamentation with a main cusp having a very circular cross section basally.
The teeth of O. winkleri resemble those of the extant sandtiger shark, O. ferox (Risso, 1810) and O. winkleri probably had similar feeding and environmental adaptations. We therefore hypothesize that O. winkleri was most likely a benthopelagic shark which we also infer for the species from the Crescent Formation described here because of similar dental morphologies.
Genus Palaeohypotodus Glikman, 1964
Type species. Odontaspis rutoti Winkler, 1874a; middle Paleocene (Selandian), Belgium.
Stratigraphic range. Danian (early Paleocene) to Priabonian (late Eocene).
Palaeohypotodus sp.
Figure 4j, k
Material. A single, very fragmentary tooth, UWBMVP 124409.
Locality. “Maple Creek prospect”, lower unit of the Crescent Formation, Hamma Hamma River, Mason County, Washington (UWBM VP loc. C3331; UWBM IP loc. B9564).
Description. A single, very fragmentary tooth displays a basally broad main cusp, of which the upper part, probably two-thirds, is missing. The main cusp is flanked by broad and low lateral cusplets that are basally divergent but with apices that are curved towards the main cusp. The base of the main cusp and cusplets bear strong, flexuous basal folds. Along the basal margin of the crown, additional, fine vertical wrinkles occur. The cutting edge of the main cusp as well as those of the lateral cusplets are very prominent and reach the corresponding basis. The labial face of the main cusp is rather flat while the lingual face is strongly convex mesio-distally.
Remarks. The few traits that are preserved and accessible for examination unambiguously allows referral to genus Palaeohypotodus. Ebersole et al. (2024) recently reviewed the taxonomy of Palaeohypotodus concluding that the species P. rutoti as currently understood represents a wastebasket taxon and this species should probably best restricted to occurrences from the Selandian. The distinct morphology of the single specimen described here indicates it may represent a hitherto unknown species of Palaeohypotodus. However, more and better-preserved material is necessary to identify this species beyond any doubt. Generally, species of Palaeohypotodus are considered to represent sharks adapted to cool waters (e.g., Kriwet et al. 2016). Palaeohypotodus ranges stratigraphically from the early Paleocene to the Ypresian and occurs in Antarctica (Long 1992; Kriwet et al. 2016), Europe (Leriche 1951; Gurr 1962; Casier 1967; Herman 1972; Ward 1980; Rayner et al. 2009; Iserbyt and De Schutter 2012; Cappetta 2012), Greenland (Bendix-Almgreen 1969), Africa (Herman 1973; Cappetta 2012), central Asia (Glikman 1964), South America (Otero and Soto-Acuna 2015; Rodriguez et al. 2023) and North America (Ward and Wiest 1990; Ebersole et al. 2024). The only previous record of Palaeohypotodus from the Pacific Coast of North America was a single tooth from the middle-early Eocene uppermost part of the Crescent Formation on the south side of the Olympic Peninsula (Squires and Goedert 1994). The stratigraphically youngest occurrences are from late Eocene rocks in the Southern Hemisphere suggesting that high latitude environments might have served as refugium before the extinction of Palaeohypotodus at the end of the Eocene.
Genus Brachycarcharias Cappetta and Nolf, 2005
Type species. Lamna lerichei Casier, 1946; lower Eocene, Ypresian, Forest-lez-Bruxelles, Belgium.
Stratigraphic range. Danian (early Paleocene) to Lutetian (middle Eocene).
Brachycarcharias cf. lerichei (Casier, 1946).
Figure 5a–d

Teeth of Lamniformes (Brachycarcharias, Macrorhizodus, Jaekelotodus), from the Hamma Hamma locality, lower unit of the Crescent Formation, early Eocene: a Brachycarcharias cf. lerichei, UWBMVP 124410, labial view; b B. cf. lerichei, NRM PAL P19793, labial view; c B. cf. lerichei, UWBMVP 124411, labial view; d B. cf. lerichei, UWBMVP 124412, labial view; e Macrorhizodus praecursor, UWBMVP 124415, lingual view; f M. praecursor, same specimen as e, lateral view; g M. praecursor, UWBMVP 124416, labial view; h Jaekelotodus sp., UWBMVP 124417, labial view; i Jaekelotodus sp., UWBMVP 124418, labial view; j Jaekelotodus sp., UWBMVP 124419, lingual view. Scale bars equal 2 mm
Material. Three incomplete anterior and three incomplete lateral teeth; four figured specimens NRM PAL P19793, UWBMVP 124410, 124411, 124412, and two referred specimens UWBMVP 124413, 124414.
Locality. “Maple Creek prospect”, lower unit of the Crescent Formation, Hamma Hamma River, Mason County, Washington (UWBM VP loc. C3331; UWBM IP loc. B9564).
Description. Three incomplete teeth preserve an upright to slightly distally inclined main cusp with a single pair of lateral cusplets in labial view. Unfortunately, the teeth lack roots. The labial face of the main cusp is flat and completely devoid of any ornamentation, but bears a short, vertical ridge basally. The cutting edges of the main cusp reach the base. The lateral cusplets are broadly triangular, rather low and somewhat divergent. The mesial cusplet is more slender with almost straight cutting edges, while the distal cusplet is broader with a concave mesial cutting edge. Both also lack any ornamentation.
Lateral teeth display a more triangular, comparably lower main cusp. Unfortunately, these lack any remnants of the root and are exposed in labial view only. The preserved lateral cusplets are low, triangular and not well separated from the main cusp. Both main cusp and lateral cusplets are devoid of ornamentation.
Remarks. Based on the preserved characters, the teeth are assigned to the extinct sandtiger shark Brachycarcharias and are most similar to those of the type species, B. lerichei which is known from Ypresian deposits of Europe, North America, Africa, and Antarctica (e.g., Cappetta 2006). Unfortunately, the teeth from the Hamma Hamma locality are too incomplete to unambiguously assign them to any species. The specimen in Fig. 5d displays rather high lateral cusplets and its assignment to B. lerichei is probably questionable as it could represent B. atlasi (J.A. Ebersole, pers. comm. 2024). However, it would be necessary to also study the lingual crown face to unambiguously assign it to B. atlasi, which currently is not possible. Consequently, we refer this tooth tentatively to B. lerichei pending more and better-preserved material. Brachycarcharias was most likely an epipelagic shark, which was adapted to warmer waters and its disappearance (like that of Striatolamia) at the end of the Eocene might be related to the gradual cooling of sea-water temperatures and the establishment of vast polar ice shields at the Eocene–Oligocene transition (e.g., Marramà et al. 2018).
Genus Jaekelotodus Menner, 1928
Type species. Hypotodus trigonalis Jaeckel, 1895; from the lower Oligocene, Ukraine.
Stratigraphic range. Danian (early Palaeocene) to Rupelian (early Oligocene).
Jaekelotodus sp.
Figure 5h–j
Material. Seven incomplete teeth; three figured specimens, UWBMVP 124417–124419, and four referred specimens: UWBMVP 124423, 124424, 124425, and 124427. Two additional specimens, 124421 and 124422 are not well exposed but are tentatively referred here as well.
Locality. “Maple Creek prospect”, lower unit of the Crescent Formation, Hamma Hamma River, Mason County, Washington (UWBM VP loc. C3331; UWBM IP loc. B9564).
Description. Only isolated tooth crowns, lacking the root and lateral cusplets, are preserved. They display either the labial (Fig. 5h, i) or lingual crown face (Fig. 5j). All specimens display a rather distally hooked crown. The labial face is flat without any ornamentation but may display a short basal vertical groove. The lingual face is rather convex transversely and also completely devoid of any ornamentation. The cutting edges reach towards the base of the crown in all specimens.
Remarks. Unfortunately, these teeth are too incomplete for any definite taxonomic assignment. The distally hooked cusp with seemingly complete cutting edges and completely smooth labial and lingual crown faces allow identification of them as upper lateral teeth of Jaekelotodus, probably of J. trigonalis (see Cappetta and Nolf 2005). However, any of the other 14 species assigned to Jaekelotodus cannot be ruled out. In North America, reported records of Jaekelotodus are rare up to now. All North American material was either assigned to J. trigonalis from the Lutetian of the Chesapeak area (Ward and Wiest 1990) or J. robustus from the Thanetian–early Eocene of Mississippi (Case 1994) and Lutetian–Bartonian of Alabama (Ebersole et al. 2019).
The new record reported here from the Ypresian fall well within the known stratigraphic range of this odonatspidid in North America. Although very poorly preserved they are important because they extend the geographic distribution of this epipelagic shark into the North Pacific Basin and represent the northernmost Paleogene record on the North American continent.
Family Lamnidae Müller and Henle, 1838
Genus Macrorhizodus Glikman, 1964
Type species. Oxyrhina falcata Rogovich, 1861; from the upper Priabonian (Upper Eocene) of Ukraine.
Stratigraphic range. Ypresian (early Eocene) to Rupelian (early Oligocene).
Macrorhizodus praecursor (Leriche, 1905).
Material. Five incomplete teeth; three figured specimens, UWBMVP 124415–124416, 124420; and two referred specimens, UWBMVP 124426 and 124428.
Locality. “Maple Creek prospect”, lower unit of the Crescent Formation, Hamma Hamma River, Mason County, Washington (UWBM VP loc. C3331; UWBM IP loc. B9564).
Description. Lamnid sharks are only represented by rare, incomplete teeth of Macrorhizodus, all still embedded in matrix. The crowns of antero-lateral teeth are high (the largest is approximately 26 mm long), rather narrow and faintly inclined distally with a very mesio-distally convex lingual face. The tooth crown of the specimen in Fig. 2, which also is accessible in lingual view, is embedded in barite and very weathered and crushed but displays the outline of an asymmetrical root with an elongated and quite narrow mesial root lobe and a shorter, broader distal root lobe with a rounded extremity. This specimen is an anterolateral tooth, as is the isolated tooth crown in Fig. 5g.
Remarks. Kovalchuk et al. (2023) provided a lengthy and comprehensive synonymy for this species. Teeth of Macrorhizodus differ from teeth of Isurus Rafinesque 1810, most significantly in having a massive cusp that is lingually inclined with rather slender and elongated root lobes in anterior teeth, a narrow lingual neck and a very prominent lingual root protuberance.
Five species of Macrorhizodus each with successive stratigraphic occurrences are currently recognized, which represent an evolutionary lineage according to Zhelezko and Kozlov (1999): M. nolfi Zhelezko and Kozlov, 1999, from the Ypresian, M. praecursor (Leriche, 1905) (= M. americanus (Leriche, 1942); Ebersole et al. 2019) from the Ypresian and Bartonian, M. falcatus (Rogovich, 1861) from the Priabonian, and M. flandricus (Leriche 1910) from the Rupelian. The two Eocene species, M. nolfi and M. praecursor, display distinct paleogeographic distributions with M. nolfi occurring in England, Kazahkstan and Denmark, whereas M. praecursor has a worldwide distribution during the middle to late Eocene (e.g., Adnet et al. 2021, and references therein). The main characters distinguishing both species are the presence of small and vestigial lateral cusplets or hump-like structures on the lateral heels in M. nolfi, which also is characterized by smaller overall size of the teeth. Although the specimens from the Hamma Hamma locality are incomplete, they seem to lack such vestigial lateral cusplets and therefore are assigned here to M. praecursor.
Family Otodontidae Glikman, 1964
Remarks. According to Cappetta (2012), Otodontidae is comprised of two genera of iconic macrophageous sharks, Otodus and Parotodus. He further separated the genus Otodus into three subgenera based on dental characters: Otodus (Otodus) Agassiz, 1838, Otodus (Carcharocles) (Jordan and Hannibal, 1923), and Otodus (Megaselachus) (Glikman, 1964). Recently there have been arguments in favor of suppressing the use of otodontid subgenera (e.g., Ehret and Ebersole 2014; Shimada et al. 2017; Boessenecker et al. 2019), whereas others continue to use the subgeneric assignments (e.g., Ebersole et al. 2019; Kovalchuck et al. 2023), and some (e.g., Perez et al. 2019) use them as genera; there still seems to be no consensus. We follow Cappetta (2012) herein. As noted by Brignon (2021), Carcharocles Jordan and Hannibal, introduced by them as a new genus in Jordan (1923), a classification of fishes, is often misattributed to the separate paper on fossil sharks (Jordan and Hannibal 1923) published in the same year.
Genus Otodus (Carcharocles) Jordan and Hannibal, in Jordan, 1923
Type species. Squalus auriculatus Blainville, 1818, from the Lutetian (middle Eocene) of Belgium.
Stratigraphic range. Ypresian (early Eocene) to Burdigalian (early Miocene).
Otodus (Carcharocles) auriculatus (Blainville, 1818).
Figure 6a–e

Teeth of Lamniformes (Otodus) from the Hamma Hamma locality, lower unit of the Crescent Formation, early Eocene: a Otodus (Carcharocles) auriculatus, UWBMVP 124429, tooth fragment showing irregular serrations; b O. (C.) auriculatus, UWBMVP 124430, left lateral cusplet, labial view; c same as b lingual view; d same specimen as in b main cusp, labial view; e same as d lingual view. Scale bars for a–c equal 2 mm; for d, e equal 10 mm
Material. A single incomplete and fragmentary tooth with one associated lateral cusplet, UWBMVP 124430, and a second tooth represented by a small fragment, UWBMVP 124429.
Locality. “Maple Creek prospect”, lower unit of the Crescent Formation, Hamma Hamma River, Mason County, Washington (UWBM VP loc. C3331; UWBM IP loc. B9564).
Description. This otodontid is represented by an incomplete and fragmentary tooth crown (Fig. 6d, e), which would have had a height exceeding 70 mm if complete, the largest tooth from the Hamma Hamma assemblage, as well as its now isolated lateral cusplet (Fig. 6b, c). The cusplet was originally part of the single tooth when found, but weathering had reduced most of that portion of the tooth to powder. The tooth crown is slender, high and distally curved with continuous, somewhat irregular serrated cutting edges. The small, serrated lateral cusplet possesses very irregular serrae. The tooth fragment (Fig. 6a) also displays irregular serrations.
Remarks. The tooth crown and lateral cusplet from the Hamma Hamma locality are very similar to those assigned to Otodus (Otodus) and Otodus (Megaselachus), but differ from species of the first subgenus by having serrated cutting edges, and from species of the second subgenus by the more irregular serrations along the tooth crown and on the lateral cusplet, as well as a much more developed lateral cusplet, which are much smaller in Otodus (Megaselachus), if present at all. Therefore, the Hamma Hamma tooth crown and lateral cusplet are assigned to Otodus (Carcharocles).
Identification of species within Otodus (Carcharocles) is generally based on serration patterns of the central cusp and the form and serration of lateral cusplets (Kriwet et al. 2016). Following Cappetta (2012), species generally assigned to Otodus (Carcharocles) are O. (C.) aksuaticus (Menner, 1928), O. (C.) angustidens (Agassiz, 1835), O. (C.) auriculatus, O. (C.) poseidoni Zhelezko in Zhelezko and Kozlov, 1999 (including at least three different “subspecies”), O. (C.) sokolovi (Jaekel, 1895), and O. (C.) subserratus (Agassiz, 1838).
The fossils from the Hamma Hamma locality are best assigned to O. (C.) auriculatus based on the slender crown with slightly irregular serrae, which diminish in size apically, and the ragged lateral cusplets. In O. (C.) sokolovi, conversely, the serration of the lateral cusplets is finer and more regular. Otodus (C.) auriculatus was almost globally distributed during most of the Eocene, and especially in many Atlantic and Tethyan open marine settings (e.g., Ward and Wiest 1990; Long 1992; Cappetta 2012; Carlsen and Cuny 2014; Ehret and Ebersole 2014). There is an early Eocene record of O. (C.) auriculatus from Chatham Island (Keyes 1987) in the southwestern Pacific Basin. From the North Pacific Basin the only previous records of O. (C.) auriculatus are from Japan (Yabumoto and Uyeno 1994) in the “Kattachi Formation”, now included in the Miike Formation of middle Eocene age (Iguchi 2016), the Namigata Formation (Tanaka et al. 2006) shown to be latest Eocene (Matsubara 2013), and middle Eocene rocks in southern Mexico (Ferrusquía-Villafranca et al. 1999). This is the first report of O. (C.) auriculatus from the northeastern Pacific Ocean.
Family Alopiidae Bonaparte, 1838
Genus Alopias Rafinesque, 1810
Type species. Alopias macrourus Rafinesque, 1810, Recent.
Stratigraphic range. Ypresian (early Eocene) to Recent.
Alopias sp.
Figure 7a–c

Teeth of Lamniformes (Alopias, Carchariidae and Odontaspididae incertae sedis), Carcharhiniformes, and Osteichthyes (Egertonia), from the Hamma Hamma locality, lower unit of the Crescent Formation, early Eocene: a Alopias sp., NRM PAL P19795, labial view; b Alopias sp., UWBMVP 124431, labial view; c Alopias sp., NRM PAL P19796, labial view; d Scyliorhinidae indeterminate, UWBMVP 124433, lingual view; e Carcharhiniformes indeterminate, UWBMVP 124434, labial(?) view; f Egertonia cf. isodonta, NRM PAL P19797, basal view; g same specimen as f, apical view; h–j, unidentified carchariids or odontaspidids, h UWBMVP 124435, labial view; i UWBMVP 124436, labial view; j NRM PAL P19794, labial view. Scale bars for f, g equal 0.2 mm, all others 2 mm
Material. Four teeth: a single nearly complete tooth with parts of the root preserved, UWBMVP 124431, and three isolated tooth crowns; figured specimens NRM PAL P19795, 19796, and referred specimen UWBMVP 124432.
Locality. “Maple Creek prospect”, lower unit of the Crescent Formation, Hamma Hamma River, Mason County, Washington (UWBM VP loc. C3331; UWBM IP loc. B9564).
Description. The nearly complete tooth assigned to Alopias (Fig. 7b; UWMBVP 124431) is embedded in matrix and only exposed in labial view. The cusp is rather slender, triangular, completely devoid of any ornamentation, and slightly inclined distally with an almost straight mesial cutting edge and a basally convex distal cutting edge, which straightens apically. The cutting edges extend to the base forming a very short distal heel-like structure. Lateral cusplets are absent. The basal margin of the crown is damaged so that its true form (straight or curved) remains ambiguous. The root is very incomplete and only the distal root lobe is discernable, which has a convexly curved outer margin. The root lobe does not extend far distally below the crown. The three isolated tooth crowns lack any remnants of the root and are fragmentary and very triangular with a flat and unornamented labial cusp face. The cutting edges extend to the base of the cusp and continue distally to form low heels.
Remarks. The tooth and the two tooth crowns from the Hamma Hamma locality resemble most closely teeth of the Alopias superciliosus (Lowe, 1841) group. In the Eocene, Alopias is predominantly known from tropical Tethyan realms of Europe and North Africa (Case and Cappetta 1990; Underwood et al. 2011b; Cappetta 2012; Adnet et al. 2021). Three species occur in the Eocene of Europe: Alopias crochardi Ward, 1978, from the Ypresian (early Eocene) of England, A. cf. denticulatus Cappetta, 1981, from the Ypresian (early Eocene) of France (Adnet 2006), which was originally described from Ypresian (early Eocene) rocks of Morocco. Alopias leensis Ward, 1978, from the middle-late Eocene boundary in England, was reassigned to Usakias Zhelezko and Kozlov 1999, by Kozlov (2000). Conversely, in North America, Alopias is very rare in Eocene strata. White (1956) ascribed some Eocene teeth from Alabama to a new species, A. latidens alabamensis. This species, however, is somewhat dubious and needs a detailed reanalysis to confirm its taxonomic status. For example, Ebersole et al. (2019) placed this species into synonymy with Negaprion gilmorei (White, 1956). Burdon (2009) presented some Middle Eocene (Lutetian) teeth assigned to Alopias latidens Leriche, 1909, from North Carolina, which slightly resemble the tooth described here. Nevertheless, we do not assign the tooth from the North Pacific Basin to any species until more material is available. This is the first Eocene record of Alopias from the North Pacific Basin.
Order Carcharhiniformes Compagno, 1973
Family Scyliorhinidae Gill, 1862
Genus and species indeterminate
Figure 7d
Material. A single incomplete tooth, UWBMVP 124433.
Locality. “Maple Creek prospect”, lower unit of the Crescent Formation, Hamma Hamma River, Mason County, Washington (UWBM VP loc. C3331; UWBM IP loc. B9564).
Description. The single small and incomplete tooth is exposed in lingual view based on the mesio-distally very convex cusp face. The cusp is cone-shaped with a rather blunt apex. The cutting edges seem weak extending to the base of the cusp. Lateral cusplets are not preserved, which, however, is more likely due to damage during or after deposition. There are well-developed short and flexuous ridges in the basal part of the cusp that extend apically for a short distance and basally across a narrow rim-like shelf. Some of the folds bifurcate basally. The basal margin of the crown is almost vertical in lateral view and the upper edge of the rim-like shelf appears very crenulated due to the bent ridges. The root is not preserved.
Remarks. The sturdy shape of the cusp and strong ornamentation suggests that this tooth belongs to a member of carcharhiniform sharks, most likely a scyliorhinid. However, more and better material showing both labial and lingual faces as well as potential lateral cusplets is necessary for any more definite taxonomic assignment.
Order Carcharhiniformes Compagno, 1973
Family, genus and species indeterminate
Figure 7e
Material. A single, very incomplete tooth, UWBMVP 124434.
Locality. “Maple Creek prospect”, lower unit of the Crescent Formation, Hamma Hamma River, Mason County, Washington (UWBM VP loc. C3331; UWBM IP loc. B9564).
Description. This very incomplete tooth displays a rather broad, basally flaring, upright main cusp in labial view, which is approximately symmetrical. The labial face is flat and devoid of any ornamentation. The upper part of the cusp is missing. The cutting edge extends to the base of the main cusp and is continuous with the one preserved lateral cusplet. This lateral cusplet is very small with a very distinct ornamentation pattern consisting of a ridge that follows the outline of the lateral cusplet. An additional ridge occurs connecting the main ridge with the cutting edge close to the transition of the cusp-cusplet cutting edge. Shorter ridges also are present. The base of the crown is heavily damaged and the root is missing as is the lateral cusplet of one side.
Remarks. It is not possible to assign this very fragmentary tooth to any taxon although the ornamentation pattern of the single preserved lateral cusplet is very distinct. Based on this the tooth might represent a scyliorhinid, but more and better-preserved material is necessary for any conclusive identification.
Class Osteichthyes Huxley, 1880
Subclass Actinopterygii sensu Goodrich, 1930 (see Schwarzhans et al., 2020).
Division Teleosteomorpha Arratia et al., 2004
Subdivision Teleostei Müller, 1845
Supercohort Teleocephala de Pinna, 1996
Cohort Elopomorpha Greenwood et al., 1966
Order Elopiformes Sauvage, 1875
Family Phyllodontidae Dartevelle and Casier, 1943
Subfamily Phyllodontinae Dartevelle and Casier, 1949
Genus Egertonia Cocchi, 1864
Type species. Egertonia isodonta Cocchi, 1864, Ypresian (early Eocene), Isle of Sheppey, United Kingdom.
Stratigraphic range. Maastrichtian (late Cretaceous) to Bartonian (middle Eocene).
Egertonia cf. isodonta Cocchi, 1864
Figure 7f, g
Material. A single, isolated tooth, NRM PAL P19797.
Locality. “Maple Creek prospect”, lower unit of the Crescent Formation, Hamma Hamma River, Mason County, Washington (UWBM VP loc. C3331; UWBM IP loc. B9564).
Description. The single tooth is circular in outline with a very low, slightly convex crown. The crown surface and its base are completely smooth without any traces of ornamentation. The basal edge of the crown is broad. A very shallow and wide pulp cavity, which is not surrounded by a distinct band of dentine, is discernable in basal view.
Remarks. Several actinopterygian fishes with sub-circular to circular, hemispherical crushing teeth such as the pycnodontid Pycnodus Agassiz, 1835, the phyllodontids Egertoni Cocchi, 1864, and Phyllodus Agassiz, 1837 (non 1835; see Brown 1890), and the albulid Albula Gronow, 1763, co-occurred in Eocene marine environments. Complete dentitions of these taxa are easy to discriminate, but it is more problematic to separate them based on isolated teeth. Nevertheless, all taxa display characters that help in categorizing isolated teeth with a fair degree of confidence. Accordingly, teeth of Pycnodus, which is the last survivor of a previously very successful stem teleostean lineage (Kriwet 2005b), are not circular but rather semi-circular to oval in outline and display an ornamented occlusal crown surface when not abraded. Whereas teeth of Paralbula Blake, 1940, are very similar to the one presented here, especially in having unornamented, convex crowns, they differ in that they are more domed and have a central pulp cavity that is surrounded by a distinct band of dentine. Teeth of Phyllodus are circular (in lateral files) to oval (in median files) and thus might resemble that from Hamma Hamma locality in some cases. These teeth, nevertheless, can be distinguished by a less domed crown and presence of ornamentation patterns. Teeth of Albula can be readily differentiated from the tooth described here by a nearly flat to slightly convex occlusal surface and having a base with either straight and parallel, biconvex or tapering edges. Consequently, it is most plausible to assign the isolated tooth to the genus Egertonia.
This taxon currently is considered monospecific (Ebersole et al. 2019), although some other species previously were described: E. gaultina Cornuel, 1877, from the upper Gault (Lower Cretaceous) of France, which has been considered invalid and was synonymized with Casierius heckelii Estes, 1969a; E. gosseleti Leriche, 1900, from the lower Eocene of France, which represents a junior synonym of E. isodonta; E. insignis Cocchi, 1864, from the Paleocene of Belgium, which has to be considered invalid (nomen nudum); and E. stromeri Weiler, 1929, from the Eocene of Egypt, which was transferred to Paralbula by Estes (1969b).
While Egertonia is rather common in Paleogene, especially Eocene strata; Cretaceous records are less common and known unambiguously only from Maastrichtian strata in Madagascar (Ostrowski 2021) and India (Halliday et al. 2016). North American records from Campanian and Maastrichtian rocks (Wheeler 1966; Glaser 1979) need to be re-evaluated. Nonetheless, it is evident that this phyllodont teleost had a rather wide distribution already in the late Cretaceous that included the Southern Hemisphere, while this fish only occurred in the Northern Hemisphere during the Paleogene according to our current knowledge. During the Eocene, Egertonia was widespread across North America, having been found in Alabama, Georgia, Mississippi, North Dakota, South Carolina, and Virginia (Ebersole et al. 2019). The new record presented here expands the range of Egertonia into the northeastern Pacific Ocean during early Eocene time.
Egertonia was predominantly a marine, near-coastal fish that nevertheless also entered estuaries and even continental environments (e.g., Ebersole et al. 2019). The occurrence of the single tooth described here from the deep-water deposits of the Crescent Formation most likely represents an allochthonous element.
Conclusions
The Hamma Hamma fish fauna includes at least 14 taxa and is the first early Eocene shark assemblage to be reported from western North America. It includes the first early Eocene records of aff. Chlamydoselachus, Notorynchus, Isistius, Mitsukurina, Jaekelotodus, Otodus (C.) auriculatus, Alopias and the teleost Egertonia from the North Pacific Basin. Most common are robust, indeterminate teeth (Fig. 7h–j, and 25 additional specimens, lot UWBMVP 124437) of carchariids and odontaspidids (sensu Stone and Shimada 2019), while remains of typical deep-sea sharks such as squaliforms, with the exception of one probably mesopelagic squaliform taxon, as well as batomorphs (rays and skates) have not yet been discovered. Elasmobranch teeth are common at the Hamma Hamma site, but unfortunately taxonomic identification is rendered difficult or even impossible by their incomplete and fragmentary nature. Many seem to lack the root, or the root cannot be readily differentiated from the enclosing matrix, whereas in some specimens remnants of the upper root portions are visible. Moreover, all specimens are embedded in extremely hard matrix, which does not allow extraction of the teeth without damaging them further. Importantly, the hard matrix does not allow for disaggregation and screening; that a tooth is discovered at all is partly serendipitous, depending on how each piece of rock is broken. In spite of this, very small teeth may actually be rare in this deposit because very small dermal denticles were often found, so it is unlikely that small teeth were missed because of collecting bias. However, this report on the Hamma Hamma assemblage should be considered preliminary because more detailed collecting over a longer time would undoubtedly reveal more taxa.
The extant goblin shark, Mitsukurina owstoni and frilled shark, Chlamydoselachus anguineus have a depth range of ca. 100 to more than 1000 m, but seem to regularly frequent deep-water seamounts (e.g., Compagno 2001; Kukuev and Pavlov 2008; Delgado et al. 2017), which is in good accordance with the depositional setting for the submarine basalts of the lower Crescent Formation interpreted as being, in part, ancient seamounts. The extant species, Notorynchus cepedianus (Péron, 1807) has a depth range of 0 to ca. 600 m (Cox and Francis 1997) but is seemingly very common inshore up to ca. 80 m depth (e.g., Compagno et al. 1989). Therefore, Notorynchus is considered a demersal shallow water hexanchiform that also thrives into the mesopelagic zone, but not deeper.
The clear dominance of lamniform sharks, especially of odontaspidids, is striking but most likely represents a collection or preservational artifact. This interpretation is supported by the complete lack of fossils of batoids (rays and skates). The two unidentified small carcharhiniforms may represent hitherto unknown Ypresian taxa. The taxonomic composition and dominance of epipelagic lamniform sharks indicate that this is a taxonomically highly uneven association with many typical deep-water taxa such as typical squaliforms, and rajiforms still to be discovered. So far, only one type of bony fish (Egertonia) has been recovered, which may be allochthonous. The Hamma Hamma assemblage nevertheless provides novel information about early Eocene fishes of North America and especially of the North Pacific Basin. It is a mixed fauna comprised of benthopelagic deep-water taxa (aff. Chlamydoselachus) and bathydemersal deep-water taxa (Mitsukurina, Odontaspis), open marine, epipelagic sharks (e.g., Alopias, Brachycarcharias, Jaekelotodus, Macrorhizodus, Otodus, Striatolamia), and at least one demersal, shallow marine shark (Heterodontus). Together, this assemblage corroborates paleoenvironmental interpretations for deposition of the lower Crescent Formation that were previously based only on microfossils and geology.
References
Adnet, S. 2006. Nouvelles faunes de sélaciens (Elasmobranchii, Neoselachii) de l’Éocène des Landes (Sud-Ouest, France). Implication dans les connaissances des communautés d’eaux profondes. Palaeo Ichthyologica 10: 1–128.
Adnet, S., and H. Cappetta. 2008. New fossil triakid sharks from the early Eocene of Prémontré, France, and comments on fossil record of the family. Acta Palaeontologica Polonica 53: 433–448.
Adnet, S., H. Cappetta, and J. Reynders. 2008. Contribution of Eocene sharks and rays from southern France to the history of deep-sea selachians. Acta Geologica Polonica 58: 261–264.
Adnet, S., I. Feichtinger, M. Harzhauser, and J. Pollerspöck. 2021. A mesopelagic selachian fauna from the middle Eocene of St. Pankraz (Austria) reveals homogeneity in deep-marine environments during the warm period in Europe. Neues Jahrbuch Für Geologie Und Paläontologie, Abhandlungen 301: 25–63.
Adolfssen, J.S., and D.J. Ward. 2015. Neoselachians from the Danian (early Paleocene) of Denmark. Acta Palaeontologica Polonica 60: 313–338.
Agassiz, L. 1835. Recherches sur les poissons fossiles, 5th livraison. Neuchâtel, Switzerland, Petitpierre et Prince and H. Nicolet, Tome 2 (livr. 5): 85–200.
Agassiz, L. 1837. Recherches sur les poissons fossiles, 8th livraison. Neuchâtel, Switzerland, Petitpierre et Prince and H. Nicolet, Tome 2 (livr. 8): 225–241.
Agassiz, L. 1838. Recherches sur les poissons fossiles, 11th livraison. Neuchâtel, Switzerland, Petitpierre et Prince and H. Nicolet, Tome III (livr. 11): 73–140.
Agassiz, L. 1843. Recherches sur les poissons fossiles, 15th et 16th livraisons. Neuchâtel, Switzerland, Petitpierre et Prince and H. Nicolet, Tome III (livr. XV, XVI): 157–390.
Anderson, M.L., R.J. Blakely, R.E. Wells, and J.D. Dragovich. 2024. Deep structure of Siletzia in the Puget Lowland: imaging an obducted plateau and accretionary thrust belt with potential fields. Tectonics 43: e2022TC007720. https://doi.org/10.1029/2022TC00720.
Applegate, S.P. 1968. A large fossil sand shark of the genus Odontaspis from Oregon. State of Oregon, Department of Geology and Mineral Industries, The Ore Bin 30: 32–36.
Arambourg, C. 1952. Les vertebres fossiles des gisements de phosphates (Maroc-Algerie-Tunisie). Notes et Mémoires du Service Géologique du Maroc 92: 1–372.
Argyriou, T., A. Alexopoulos, J.D. Carrillo-Briceño, and L. Cavin. 2022. A fossil assemblage from the mid–late Maastrichtian of Gavdos Island, Greece, provides insights into the pre-extinction pelagic ichthyofaunas of the Tethys. PLoS ONE 17: e0265780.
Arnold, R. 1906. Geological reconnaissance of the coast of the Olympic Peninsula, Washington. Geological Society of America Bulletin 17: 451–468.
Arratia, G., R. Scasso, and W. Kiessling. 2004. Late Jurassic fishes from Longing Gap, Antarctic Peninsula. Journal of Vertebrate Paleontology 24: 41–55.
Ayres, W. 1855. Descriptions of new species of California fishes. Proceedings of the California Academy of Natural Science 1: 23–77.
Babcock, R.S., R.F. Burmester, D.C. Engebretson, A. Warnock, and K.P. Clark. 1992. A rifted margin origin for the Crescent basalts and related rocks in the northern Coast Range volcanic province, Washington and British Columbia. Journal of Geophysical Research 97: 6799–6821.
Babcock, R.S., C.A. Suczek, and D.C. Engebretson. 1994. The Crescent “Terrane”, Olympic Peninsula and southern Vancouver Island. Washington Division of Geology and Earth Resources, Bulletin 80: 141–157.
Bendix-Almgreen, S.E. 1969. Notes on the Upper Cretaceous and lower Tertiary fish faunas of northern West Greenland. Meddelelser Fra Dansk Geologisk Forening 19: 204–217.
Berg, L.S. 1937. A classification of fish-like vertebrates. Izvestiya Akademii Nauk USSR, Seriya Biologischeskaya 4: 1277–1280 (In English with Russian Summary).
Blainville, H.M.D. 1816. Prodrome d’une distribution systematique du regne animal. Bulletin Des Sciences Pars La Société Philomathique De Paris 8: 105–124.
Blainville, H.M.D. 1818. Sur les ichthyolites ou les poissons fossils. Nouveau Dictionnaire D’histoire Naturelle 27: 310–391.
Blake, S.F. 1940. Paralbula: A new fossil fish based on dental plates from the Eocene and Miocene of Maryland. Journal of the Washington Academy of Sciences 30: 205–209.
Bloch, M.E., and J.G. Schneider. 1801. Systema Ichthyologiae iconibus ex illustratum. Post obitum auctoris opus inchoatum absolvit, correxit, interpolavit. Saxo: J.G. Schneider.
Boessenecker, R.W., D.J. Ehret, D.J. Long, M. Churchill, E. Martin, and S.J. Boessenecker. 2019. The Early Pliocene extinction of the mega-toothed shark Otodus megalodon: A view from the eastern North Pacific. PeerJ 7: e6088. https://doi.org/10.7717/peerj.6088.
Bonaparte, C.L. 1838. Selachorum tabula analytica. Nuovi Annali Delle Scienze Naturali 1 (2): 195–214.
Brignon, A. 2021. Historical and nomenclatural remarks on some megatoothed shark teeth (Elasmobranchii, Otodontidae) from the Cenozoic of New Jersey (U.S.A.). Revista Italiana Di Paleontologia e Stratigrafia 127: 595–625.
Brown, W.H. 1890. Dates of publication of Recherches sur les Poissons fossiles, par L. Agassiz. P. XXV-XXIX. In A Catalogue of British Fossil Vertebrata, ed. A.S. Woodward and C.D. Sherborn, I–XXXV, 1–396. London: Dulau and Co.
Burdon, J. 2009. The life and times of long dead sharks – Genera – Paleogene – Odontaspis. http://www.elasmo.com/frameMe.html?file=home.html&menu=bin/menu_home-alt.html. Accessed 25 Apr 2024.
Cady, W.M. 1975. Tectonic setting of the Tertiary volcanic rocks of the Olympic Peninsula, Washington. U.S. Geological Survey Journal of Research 3: 573–582.
Cady, W.M., M.L. Sorensen, and N.S. MacLeod. 1972a. Geologic map of the Brothers Quadrangle, Jefferson, Mason, and Kitsap counties, Washington. U.S. Geological Survey Map GQ-969. Scale 1:62,500, 1 sheet.
Cady, W.M., R.W. Tabor, N.S. MacLeod, and M.L. Sorensen. 1972b. Geologic map of the Tyler Peak quadrangle, Clallam and Jefferson counties, Washington. U.S. Geological Survey Map GQ-970. Scale 1:62,500, 1 sheet.
Cappetta, H. 1975. Sélaciens et holocéphale du Gargasien del la région de Gargas (Vaucluse). Géologie Mediterranéenne 2: 115–134.
Cappetta, H. 1981. Additions à la faune de sélaciens fossiles du Maroc. 1: Sur la présence des genres Heptranchias, Alopias et Odontorhytis dans l’Yprésien des Ouled Abdoun. Geobios 14: 563–575.
Cappetta, H. 2006. Elasmobranchii Post-Triadici (Index specierum et generum). Leiden: Backhuys Publishers.
Cappetta, H. 2012. Handbook of Paleoichthyology, Vol. 3E: Chondrichthyes - Mesozoic and Cenozoic Elasmobranchii: Teeth. Munich: Dr. Friedrich Pfeil.
Cappetta, H., and D. Nolf. 2005. Révision de quelques Odontaspididae (Neoselachii: Lamniformes) du Paléocène et de l’Eocène du Bassin de la Mer du Nord. Bulletin De L’institut Royal Des Sciences Naturelles De Belgique, Science De La Terre 75: 237–266.
Cappetta, H., K. Morrison, and S. Adnet. 2021. A shark fauna from the Campanian of Hornby Island, British Columbia, Canada: An insight into the diversity of Cretaceous deep-water assemblages. Historical Biology 33: 1121–1182. https://doi.org/10.1080/08912963.2019.1681421.
Carlsen, A.W., and G. Cuny. 2014. A study of the sharks and rays from the Lillebælt Clay (early–middle Eocene) of Denmark, and their palaeoecology. Bulletin of the Geological Survey of Denmark 62: 39–88.
Case, G.R. 1980. A selachian fauna from the Trent Formation, Lower Miocene (Aquitanian) of eastern North Carolina. Palaeontographica, Abt. A 171: 75–103.
Case, G.R. 1981. Late Eocene selachians from south-central Georgia. Palaeontographica, Abt. A 176: 52–79.
Case, G.R. 1994. Fossil fish remains from the late Paleocene Tuscahoma and early Eocene Bashi Formations of Meridian, Lauderdale County, Mississippi. Part I. Selachians. Palaeontographica, Abt. A 230: 97–138.
Case, G.R., and H. Cappetta. 1990. The Eocene selachian fauna from the Fayum Depression in Egypt. Palaeontographica, Abt. A 212: 1–30.
Case, G.R., and P.D. Borodin. 2000. A middle Eocene selachian fauna from the Castle Hayne Limestone Formation of Duplin County, North Carolina. Münchner Geowissenschaftliche Abhandlungen 39: 17–32.
Casier, E. 1946. La faune ichthyologique de l’Yprésien de la Belgique. Mémoires Du Musée Royal D’histoire Naturelle De Belgique 104: 1–267.
Casier, E. 1966. Faune ichthyologique du London Clay. London: Trustees of the British Museum.
Casier, E. 1967. Le Landénien de Dormaal (Brabant) et sa faune ichthyologique. Mémoires de l'Institut Royal des Sciences Naturelles de Belgique 156: 1–66.
Ciborowski, T.J.R., B.A. Phillips, A.C. Kerr, D.N. Barfod, and D.F. Mark. 2020. Petrogenesis of Siletzia: The world’s youngest oceanic plateau. Results in Geochemistry 1: e100004. https://doi.org/10.1016/j.ringeo.2020.100004.
Cicimurri, D.J., and J.L. Knight. 2019. Late Eocene (Priabonian) elasmobranchs from the Dry Branch Formation (Barnwell Group) of Aiken County, South Carolina, USA. PaleoBios 36: 1–31.
Cigala-Fulgosi, F., S. Casati, A. Orlandini, and D. Persico. 2009. A small fossil fish fauna, rich in Chlamydoselachus teeth, from the Late Pliocene of Tuscany (Siena, central Italy). Cainozoic Research 6: 3–23.
Cocchi, I. 1864. Monografia dei Pharyngodopilidae. Nuova famiglia di pesci Labroidi: Studi paleontologici. Firenze: Cocchi.
Compagno, L.J.V. 1970. Systematics of the genus Hemitriakis (Selachii: Carcharinidae), and related genera. Proceedings of the California Academy of Sciences (Series 4) 38: 63–98.
Compagno, L.J.V. 1973. Interrelationships of living elasmobranchs. Zoological Journal of the Linnean Society 53 (Supplement 1): 15–61.
Compagno, L.J.V. 1999. Checklist of living elasmobranchs. In Sharks, Skates, and Rays: The Biology of Elasmobranch Fishes, ed. W.C. Hamlett, 471–498. Baltimore: Johns Hopkins University Press.
Compagno, L.J.V. 2001. Sharks of the world. An annotated and illustrated catalogue of shark species known to date. Bullhead, mackerel and carpet sharks (Heterodontiformes, Lamniformes and Orectolobiformes). FAO Species Catalogue for Fishery Purposes, 1, Vol. 2. Rome: FAO.
Compagno, L.J.V., D.A. Ebert, and M.J. Smale. 1989. Guide to Sharks and Rays of Southern Africa. London: New Holland (Publ.) Ltd.
Compagno, L.J.V., M. Dando, and S. Fowler. 2005. A Field Guide to the Sharks of the World. London: Harper Collins Publishers Ltd.
Consoli, C.P. 2008. A Rare Danian (early Paleocene) Chlamydoselachus (Chondrichthyes: Elasmobranchii) from the Takatika Grit, Chatham Islands, New Zealand. Journal of Vertebrate Paleontology 28: 285–290.
Cornuel, M.J. 1877. Description de débris de Poissons fossiles provenant principalement du calcaire néocomien du département de la Haute-Marne. Bulletin De La Société Géologique De France, Série 3 (5): 604–626.
Cox, G., and M. Francis. 1997. Sharks and Rays of New Zealand. Canterbury (Australia): Canterbury University Press.
Dartevelle, E., and E. Casier. 1943. Les poissons fossiles du Bas-Congo et des régions voisines, première partie. Annales de Musèe du Congo Belge, Tervuren (Belgique), Série A: Géologie, Géographie Physique, Minéralogie, Paléontologie 2(1): 1–200.
Dartevelle, E., and E. Casier. 1949. Les Poissons fossiles du Bas-Congo et des régions voisines (Deuxième partie). Annales Du Musée Du Congo Belge, Série a: Géologie, Géographie Physique, Minéralogie, Et Paléontologie 2 (2): 201–256.
de Buen, F. 1926. Catalogo ictiologico del Mediterraneo Español y de Marruecos recopilando 10 publicado sobrepeces de las costas mediterraneas y proximas del Atlantico (Mar de España). Resultado De Las Ampafias Campañas Realizadas Por Acuerdos Internacionales, Instituto Español De Oceanografia 2: 1–221.
de Pinna, M.C.C. 1996. Teleostean monophyly. In Interrelationships of Fishes, ed. M.L.J. Stiassny, L.D. Parenti, and G.D. Johnson, 147–162. San Diego: Academic.
Delgado, J., D. Carvalho, M. Freitas, M. Biscoito, and E.P. Gouveia. 2017. Records of some rare deep-sea fishes caught in the Lion, Susan, and Unicorn seamounts, off the archipelago of Madeira (east-central Atlantic). Acta Ichthyologica Et Piscatoria 47: 91–96.
Ebersole, J.A., D.J. Cicimurri, and G.L. Stringer. 2019. Taxonomy and biostratigraphy of the elasmobranchs and bony fishes (Chondrichthyes and Osteichthyes) of the lower-to-middle Eocene (Ypresian to Bartonian) Claiborne Group in Alabama, USA, including an analysis of otoliths. European Journal of Taxonomy 585: 1–274.
Ebersole, J.A., D.J. Cicimurri, and T.L. Harrell Jr. 2024. A new species of Palaeohypotodus Glückman, 1964 (Chondrichthyes, Lamniformes) from the lower Paleocene (Danian) Porters Creek Formation, Wilcox County, Alabama, USA. Fossil Record 27: 111–134.
Ebert, D.A., and L.J.V. Compagno. 2009. Chlamydoselachus africana, a new species of frilled shark from southern Africa (Chondrichthyes, Hexanchiformes, Chlamydoselachidae). Zootaxa 2173: 1–18.
Ebert, D.A., S. Fowler, and L.J.V. Compagno. 2013. Sharks of the World – A fully illustrated guide, 528. San Francisco: Wild Nature Press.
Ebert, D.A., S.P. Wintner, and P.M. Kyne. 2021. An annotated checklist of the chondrichthyans of South Africa. Zootaxa 4947: 1–127.
Eddy, M.P., K.P. Clark, and M. Polenz. 2017. Age and volcanic stratigraphy of the Eocene Siletzia oceanic plateau in Washington and on Vancouver Island. Lithosphere 9: 652–664.
Ehret, D.J., and J. Ebersole. 2014. Occurrence of the megatoothed sharks (Lamniformes: Otodontidae) in Alabama, USA. PeerJ 2: e625. https://doi.org/10.7717/peerj.625.
Estes, R. 1969a. Studies on fossil phyllodont fishes: Casierius, a new genus of albulid from the Cretaceous of Europe and North America. Eclogae Geologicae Helvetiae 62: 751–755.
Estes, R. 1969b. Studies on fossil phyllodont fishes: Interrelationships and evolution in the Phyllodontidae. Copeia 2: 317–331.
Ferrusquía-Villafranca, I., S.P. Applegate, and L. Espinosa-Arrubarrena. 1999. First Paleogene selachifauna of the middle American-Caribbean-Antillean region, La Mesa de Copoya, west-central Chiapas, Mexico—systematics and paleontological significance. Revista Méxicana De Ciencias Geológicas 16: 155–174.
Garman, S. 1884. An extraordinary shark. Bulletin of the Essex Institute 16: 47–55.
Garrick, J.A.F., and S. Springer. 1964. Isistius plutodus, a new squaloid shark from the Gulf of Mexico. Copeia 1964: 678–682.
Gartner, J.V., Jr., R.E. Crabtree, and K.J. Sulak. 1997. Feeding at depth. In Deep-sea Fishes, ed. D.J. Randall and A.P. Farrell, 115–194. San Diego: Academic Press.
Gill, T. 1862. Squalorum generum novorum descriptiones diagnosticae. Annals of the Lyceum of Natural History of New York 7: 409–413.
Gill, T. 1864. Second contribution to the Selachology of California. Proceedings of the Academy of Natural Sciences of Philadelphia 16: 147–151.
Gill, T. 1893. Families and subfamilies of fishes. Memoirs of the National Academy of Sciences, Washington 6: 125–138.
Glaser, J.D. 1979. Collecting fossils in Maryland. Maryland Geological Survey Educational Series 4: 1–83.
Glikman, L.S. 1964. Class Chondrichthyes, Subclass Elasmobranchii (in Russian). In Fundamentals of Paleontology, Vol. 11, ed. D.V. Obruchev, 196–237. Moscow–Leningrad: Nauka SSSR.
Gonzalez-Barba, G. 2004. Ypresian sharks, batoids, and teleost fishes from the Tepetate and Bateque formations, Baja California Sur, Mexico. Journal of Vertebrate Paleontology 24, supplement 3 (abstracts of papers): 65A.
Goodrich, E.S. 1909. Vertebrata Craniata (First fascicle: Cyclostomes and Fishes). In A treatise on Zoology, ed. R. Lankester, 1–514. London: Adam and Charles Black.
Goodrich, E.S. 1930. Studies on the Structure and Development of Vertebrates. London: Macmillan and Co.
Gray, J. 1851. List of the Specimens of Fish in the Collection of the British Museum. Part I -Chondropterygii. London: Printed by the Order of the Trustees.
Green, S.H. 1945. Manganese deposits of the Olympic Peninsula, Washington. State of Washington, Division of Mines and Mining, Report of Investigations 7: 1–45, map in pocket.
Greenwood, P.H., D.E. Rosen, S.H. Weitzman, and G.S. Myers. 1966. Phyletic studies of teleostean fishes, with a provisional classification of living forms. Bulletin of the American Museum of Natural History 131: 339–456.
Gronow, L.T. 1763. Zoophylacii Gronoviani fasciculus primus exhibens animalia quadrupeda, amphibia atque pisces, quae in museo suo adservat, rite examinavit, systematice disposuit, descripsit atque illustravit Laur. Theod. Gronovius, J.U.D. Lugduni Batavorum (Haak and Luchtmans), 136 p.
Gurr, P.R. 1962. A new fish fauna from the Woolwich Bottom Beds (Sparnacian) of Herne Bay, Kent. Proceedings of the Geologists’ Association 73: 419–447.
Halliday, T.J.D., A.R. Cuff, G.V.R. Prasad, M.S. Thanglemmoi, and A. Goswami. 2016. New record of Egertonia (Elopiformes, Phyllodontidae) from the Late Cretaceous of South India. Papers in Palaeontology 2: 287–294.
Hay, O.P. 1902. Bibliography and catalogue of the fossil Vertebrata of North America. Bulletin of the United States Geological and Geographical Survey of the Territories 179: 1–868.
Herman, J. 1972. Les vertébrés du Landénien Inférieur (LIa ou Heersien) de Maret (Hameau d’Orp-le-Grand). Bulletin De La Société Belge De Géologie, De Paléontologie Et D’hydrologie 81: 191–207.
Herman, J. 1973. Contribution à la connaissance de la faune ichthyologique des phosphates du Maroc. Annales De La Société Géologique De Belgique 95: 271–284.
Herman, J., Hovestadt-Euler, M., & Hovestadt, D.C. 1993. Contributions to the study of the comparative morphology of teeth and other relevant ichthyodorulites in living superaspecific taxa of Chondrichthyan fishes. Addendum to Part A, No. 1b: Hexanchiformes - Family: Chlamidoselachidae; No. 5: Order: Heterodontiformes - Family: Heterodontidae; No. 6: Order: Lamniformes - Families: Cetorhinidae, Megachasmidae; Addendum 1 to No. 3: Order: Squaliformes; Addendum 1 to No. 4: Order Orectolobiformes; General Glossary; Summary Part A. Bulletin de l'Institut Royal des Sciences Naturelles de Belgique, Biologie 63: 185–256.
Hovestadt, D.C. 2018. Reassessment and revision of the fossil Heterodontidae (Chondrichthyes: Neoselachii) based on tooth morphology of extant taxa. Palaeontos 30: 1–73.
Hovestadt, D., and E. Steurbaut. 2023. Annotated iconography of the type specimens of fossil chondrichthyan fishes in the collection of the Royal Belgian Institute of Natural Sciences, Brussels. Royal Belgian Institute of Natural Sciences, Brussels: Monographs in Natural Sciences 1: 1–122.
Hughes, G.E. 1939. Geology of a barite prospect on the Hamma Hamma River and treatment of the ore [B.S. thesis]. Seattle: University of Washington.
Huxley, T.H. 1880. On the application of the laws of evolution to the arrangement of the Vertebrata, and more particularly of the Mammalia. Proceedings of the Zoological Society London 43: 649–662.
Iguchi, T. 2016. Geology of the Paleogene Miike Coalfield in the Ariake Sea area and its litho-biostratigraphic correlation with Paleogene coalfields in northwestern Kyushu, Japan. Journal of Geography (chigaku Zasshi) 125: 325–351.
Iserbyt, A., and P.J. De Schutter. 2012. Quantitative analysis of elasmobranch assemblages from two successive Ypresian (early Eocene) facies at Marke, western Belgium. Geologica Belgica 15: 146–153.
Jaekel, O. 1895. Unter-tertiäre Selachier aus Südrussland. Mémoirs du Comité geologique de St. Petersburg 9: 19–35, 2 pl.
Jordan, D.S. 1898. Description of a species of fish (Mitsukurina owstoni) from Japan, the type of a distinct family of lamnoid sharks. Proceedings of the California Academy of Sciences, (Series 3) Zoology 1: 199–204.
Jordan, D.S. 1923. A classification of fishes, including families and genera as far as known. Stanford University Publications, Biological Sciences 3: 79–243.
Jordan, D.S., and H. Hannibal. 1923. Fossil sharks and rays of the Pacific Slope of North America. Bulletin of the Southern California Academy of Sciences 22: 27–68.
Kemp, N.R. 1991. Chondrichthyans in the Cretaceous and Tertiary of Australia. In Vertebrate Palaeontology of Australasia, ed. P. Vickers-Rich, J.M. Monaghan, R.F. Baird, and T.H. Rich, 497–568. Lilydale: Pioneer Design Studio.
Kent, B. 1994. Fossil sharks of the Chesapeake Bay Region. Baltimore: Egan, Rees and Boyer Publishers.
Kent, B.W. 1999. Sharks from the Fisher/Sullivan site. Part 2. Virginia Division of Mineral Resources Publication 152: 11–37.
Keyes, I.W. 1987. Eocene elasmobranchs from Te Whanga Lagoon, Chatham Island. Geological Society of New Zealand Newsletter 77: 31–35.
Kiel, S. 2008. Fossil evidence for micro- and macrofaunal utilization of large nekton-falls: Examples from early Cenozoic deep-water sediments in Washington State, USA. Palaeogeography, Palaeoclimatology, Palaeoecology 267: 161–174.
Kiraly, S.J., J.A. Moore, and P.H. Jasinski. 2003. Deepwater and other sharks of the U.S. Atlantic Ocean Exclusive Economic Zone. Marine Fisheries Review 65: 1–63.
Kočí, T., J.L. Goedert, and J.S. Buckeridge. 2022. Eocene tube-dwelling annelids (Polychaeta: Sedentaria) from the Black Hills, western Washington State: The first record of Neodexiospira from North America. Paläontologische Zeitschrift 96: 631–653.
Kovalchuk, O., J. Kriwet, K. Shimada, T. Ryabokon, Z. Barkaszi, A. Dubikovska, G. Anfimova, and S. Davydenko. 2023. Middle Eocene cartilaginous fishes (Vertebrata: Chondrichthyes) of the Dnieper-Donets Basin, northern Ukraine. Paleontologia Electronica 26 (2): a32.
Kozlov, V.A. 2000. Systematics and evolution of sharks from family Alopiidae (based on the materials from Palaeogene deposits of Middle Asia and western Kazakhstan). Materialy Po Stratigrafii i Paleontologii Urala 4: 142–147 (in Russian).
Kriwet, J. 2005a. Additions to the Eocene selachian fauna of Antarctica with comments on Antarctic selachian diversity. Journal of Vertebrate Paleontology 25: 1–7.
Kriwet, J. 2005b. A comprehensive study of the skull and dentition of pycnodont fishes (Neopterygii, Pycnodontiformes). Zitteliana 45: 135–188.
Kriwet, J., and S. Klug. 2009. Fossil record and origin of squaliform sharks. In Biology and Management of Dogfish Sharks, ed. V.F. Gallucci, G.A. McFarlane, and G.G. Bargmann, 19–38. Bethesda: American Fisheries Society.
Kriwet, J., A. Engelbrecht, T. Mörs, M. Reguerocd, and C. Pfaff. 2016. Ultimate Eocene (Priabonian) chondrichthyans (Holocephali, Elasmobranchii) of Antarctica. Journal of Vertebrate Paleontology 36: e1160911.
Kukuev, E.I., and V.P. Pavlov. 2008. The first case of mass catch of a rare frill shark Chlamydoselachus anguineus over a seamount of the Mid-Atlantic Ridge. Journal of Ichthyology 48: 676–678.
Leriche, M. 1900. Faune ichthyologique des sables Unios et Térédines des environs d’Epernay (Marne). Annales De La Sociétè Géologique Du Nord 29: 173–196.
Leriche, M. 1905. Les poissons éocènes de la Belgique. Mémoires Du Musée Royal D’histoire Naturelle De Belgique 3: 49–228.
Leriche, M. 1909. Note prélininaire sur des poissons nouveaux de l’Oligocène Belge. Bulletin De La Société Belge De Géologie, De Paléontologie Et D’hydrologie 22: 378–384.
Leriche, M. 1910. Les poissons tertiaires de la Belgique. III. Les poissons oligocènes. Mémoires Du Musée Royal D’histoire Naturelle De Belgique 5: 229–363.
Leriche, M. 1942. Contribution à l’étude des faunes ichthyologiques marines des terrains tertiaires de la Plaine Côtière Atlantique et du centre des Etats-Unis. Les synchronismes des formations tertiaires des deux côtés de l’Atlantique. Mémoires De La Société Géologique De France 45: 1–110.
Leriche, M. 1951. Les poissons tertiaires de la Belgique (Supplément). Mémoires De L’institut Royal Des Sciences Naturelles De Belgique 118: 473–600.
Long, D.J. 1992. Sharks from the La Meseta Formation (Eocene), Seymour Island, Antarctica Peninsula. Journal of Vertebrate Paleontology 12: 11–32.
Lowe, R.T. 1841. A paper from the Rev. R.T. Lowe, M.A., describing certain new species of Madeiran fishes, and containing additional information relating to those already described. Proceedings of the Zoological Society of London 8: 36–39.
Maisey, J.G. 2012. What is an ‘elasmobranch’? The impact of palaeontology in understanding elasmobranch phylogeny and evolution. Journal of Fish Biology 80: 918–951.
Malyshkina, T.P. 2021. Striatolamia tchelkarnurensis Glikman (Elasmobranchii: Lamniformes), the youngest valid Striatolamia species. Paleontological Journal 55: 193–204.
Manning, E.M., and B.R. Standhardt. 1986. Late Eocene sharks and rays of Montgomery Landing, Louisiana. In Montgomery Landing Site, Marine Eocene (Jackson) of Central Louisiana, ed. J.A. Schiebout and W. van den Bold, 133–169. Baton Rouge: Gulf Coast Association of the Geological Societies.
Marramà, G., A. Engelbrecht, T. Mörs, M. Reguero, and J. Kriwet. 2018. The southernmost occurrence of Brachycarcharias (Lamniformes, Odontaspididae) from the Eocene of Antarctica provides new information about the paleobiogeography and paleobiology of Paleogene sand tiger sharks. Rivista Italiana Di Paleontologia e Stratigrafia 124: 283–297.
Marramà, G., A. Engelbrecht, G. Carnevale, and J. Kriwet. 2019. Eocene sand tiger sharks (Lamniformes, Odontaspididae) from the Bolca konservat-lagerstätte, Italy: Palaeobiology, palaeobiogeography and evolutionary significance. Historical Biology 31: 102–116.
Martin, R.A., and J. Treberg. 2002. Biology of Deep-Sea Sharks: a review [Abstract]. International Congress on the Biology of Fish, July 2002, Vancouver, Canada, and extended abstract published in Symposium Proceedings.
Matsubara, T. 2013. Molluscan fauna from the “Miocene” Namigata Formation in the Namigata area, Okayama Prefecture, southwest Japan. Journal of the Geological Society of Japan 119: 249–266.
McCrory, P.A., and D.S. Wilson. 2013. A kinematic model for the formation of the Siletz-Crescent forearc terrane by capture of the coherent fragments of the Farallon and Resurrection plates. Tectonics 32: 718–736.
Menner, V.V. 1928. Les Sélaciens du Paléogène de Manghyschlak, d’Emba et du versant oriental d’Oural. Bulletin De La Société Impériale Des Naturalistes De Moscou, Section Géologique 6: 292–338.
Meyer, F.A.A. 1793. Systematisch-summarische Uebersicht der neuesten zoologischen Entdeckungen in Neuholland und Afrika. Leipzig: Dykischen.
Moen, W.S. 1964. Barite in Washington. State of Washington, Division of Mines and Geology Bulletin 51: 1–112, pl. 2 and Appendix B in pocket.
Moore, E.J. 1984. Molluscan paleontology and biostratigraphy of the lower Miocene upper part of the Lincoln Creek Formation in southwestern Washington. Contributions in Science 351: 1–42.
Müller, A. 1999. Ichthyofaunen aus dem atlantischen Tertiär der USA. Leipziger Geowissenschaften 9–10: 1–360.
Müller, J. 1845. Über den Bau und die Grenzen der Ganoiden, und über das natürliche System der Fische. Archiv Für Naturgeschichte 11: 91–141.
Müller, J., and F.G.J. Henle. 1838. Systematische Beschreibung der Plagiostomen. Berlin, Germany: 1–28.
Müller, J., and F.G.J. Henle. 1839. Systematische Beschreibung der Plagiostomen. Berlin, Germany: 29–10.
Musick, J.A., M.M. Harbin, and L.J.V. Compagno. 2004. Historical zoogeography of the Selachii. In Biology of Sharks and their relatives, ed. J.C. Carrier, J.A. Musick, and M.R. Heithaus, 33–78. Boca Raton: CRC Press.
Naylor, G.J.P., J.N. Caira, K. Jensen, K.A.M. Rosana, N. Straube, and C. Lakner. 2012. Elasmobranch phylogeny: A mitochondrial estimate based on 595 species. In Biology of Sharks and their relatives, ed. J.C. Carrier, J.A. Musick, and M.R. Heithaus, 31–56. Boca Raton: CRC Press.
Nazarkin, M.V., and T.P. Malyshkina. 2020. Eocene ichthyofauna of Karaginsky Island (Bering Sea). Biogeography and Evolutionary Processes, Materials of the LXVI session of the Paleontological Society at the Russian Academy of Sciences, Saint-Petersburg, Kartfabrika VSEGEI: 261–263.
Nisbet, P.C., and A.V. Ambrose. 1986. The geology and geochemistry of exhalative ore deposits in Tertiary basic fore-arc volcanic terranes, Olympic Peninsula, Washington. Offshore Technology Conference, Houston, Texas, abstracts: 413–416.
Ostrowski, S.A. 2021. Late Cretaceous Elopomorpha (Actinopterygii: Teleostei) from the Mahajanga Basin of Madagascar and impacts on paleobiogeography. Palaeontologia Electronica. https://doi.org/10.26879/1151.
Otero, R.A., and S. Soto-Acuña. 2015. New chondrichthyans from Bartonian-Priabonian levels of Río de Las Minas and Sierra Dorotea, Magallanes Basin, Chilean Patagonia. Andean Geology 42: 268–283.
Padilla, A., J.J. Eberle, M.D. Gottfried, A.R. Sweet, and J.H. Hutchison. 2014. A sand tiger shark-dominated fauna from the Eocene Arctic greenhouse. Journal of Vertebrate Paleontology 34: 1307–1316.
Pardee, J.T. 1921. Deposits of manganese ore in Montana, Utah, Oregon, and Washington. U.S. Geological Survey Bulletin 725-C: 141–243.
Park, C.F., Jr. 1942. Manganese resources of the Olympic Peninsula, Washington, a preliminary report. U.S. Geological Survey Bulletin 931-R: 435–457, pls. 68–74.
Park, C.F., Jr. 1946. The spilite and manganese problems of the Olympic Peninsula, Washington. American Journal of Science 244: 305–323.
Parmley, D., D. Cicimurri, and R. Campbell. 2003. Late Eocene sharks of the Hardie Mine local fauna of Wilkinson County, Georgia. Georgia Journal of Science 61: 153–179.
Perez, V.J., S.J. Godfrey, B.W. Kent, R.E. Weems, and J.R. Nance. 2019. The transition between Carcharocles chubutensis and Carcharocles megalodon (Otodontidae, Chondrichthyes): lateral cuspet loss through time. Journal of Vertebrate Paleontology. https://doi.org/10.1080/02724634.2018.1546732.
Péron, F. 1807. Voyage de Découvertes aux Terres Australes, exécuté par ordre de sa majesté l’Empereur et Roi, sur les Corvettes la Géographe, la Naturaliste et la Goulette le Casuarina, pendant les années 1800, 1801, 1803 et 1804. Paris. Voyage De Découvertes Aux Terres Australes 1: 1–496.
Phillips, B.A., A.C. Kerr, E.K. Mullen, and D. Weis. 2017. Oceanic mafic magmatism in the Siletz terrane, NW North America: Fragments of an oceanic plateau? Lithos 274–275: 291–303.
Pledge, N.S. 1967. Fossil elasmobranch teeth of South Australia and their stratigraphic distribution. Transactions of the Royal Society of South Australia 91: 135–164.
Pollerspöck, J., and N. Straube. 2017. A new deep-sea elasmobranch fauna from the Central Paratethys (Neuhofener Beds, Mitterdorf, near Passau, Germany, Early Miocene, Middle Burdigalian). Zitteliana 90: 27–53.
Popov, E.V., and V.A. Lopyrev. 2023. Elasmobranchs from the lower Eocene Kalinino Formation of the Alai River, Saratov Region of Russia. Historical Biology. https://doi.org/10.1080/08912963.2023.2184691.
Probst, J. 1879. Beiträge zur Kenntnis der fossilen Fische aus der Molasse von Baltringen. Hayfische. (Schluss). Jahreshefte Des Vereins Für Vaterländische Naturkunde in Württemberg 35: 127–191.
Purdy, R.W. 1998. The early Miocene fish fauna from the Pollack Farm Site, Delaware. In Geology and paleontology of the lower Miocene Pollack Farm Site, Delaware, ed. R.N. Benson, 133–139. Newark: Delaware Geological Survey.
Quoy, J.R.C., and J.P. Gaimard. 1824. Description des poissons. Chapître IX. In, Voyage autour du monde, entrepris par ordre du roi. Exécuté sur les corvettes de S.M. l'Uranie et la Physicienne, pendant les années 1817, 1818, 1819 et 1820, ed. L. de Freycinet, 192–401. Paris: Chez Pillet aîné.
Rafinesque, C.S. 1810. Indice d'ittiologia siciliana ossia catalogo metodico dei nomi latini, italiani, e siciliani, e siciliani dei pesci, che si rinvengono in Sicilia disposti secondo un metodo naturale eseguito da un appendice che contiene la descrizione di alcuni nuovi pesci siciliani. Ill Opuscolo del signore C.S. Rafinesque Schmaltz, Messina, Italy, Giovanni del Nobolo, 70 p.
Rau, W.W. 1964. Foraminifera from the northern Olympic Peninsula, Washington. U.S. Geological Survey Professional Paper 374-G: 1–33, pls. 1–7.
Rau, W.W. 1966. Stratigraphy and Foraminifera of the Satsop River area, southern Olympic Peninsula, Washington. State of Washington, Division of Mines and Geology Bulletin 53: 1–66.
Rayner, D., T. Mitchell, and M. Rayner. 2009. London Clay Fossils of Kent and Essex. Rochester: Medway Fossil and Mineral Society.
Reinecke, T., and P. Engelhard. 1997. The selachian fauna from Geschiebe of the Lower Selandian basal conglomerate (Thanetian, Late Paleocene) in the Danish subbasin (Sealand, Scania, Western Baltic Sea). Erratica 2: 3–45.
Risso, A. 1810. Ichthyologie de Nice, ou, histoire naturelle des poissons du département des Alpes Maritimes. Paris: F. Schoell.
Rodríguez, D., D.J. Ward, and J.A. Quezada. 2023. Paleontology and stratigraphic implications of a late Paleocene elasmobranch assemblage in Talcahuano, southcentral Chile. Andean Geology 50: 217–247.
Rogovich, A.S. 1861. On fossil fishes of provinces of the Kiev Academic District. First issue. Placoid fishes. Placoidei Ag. and Ganoid fishes. Ganoidei Ag. Natural History of the Provinces of the Kiev Academic District: Paleontology. Systematic part. Kiev, 87 [in Russian]
Sauvage, H.E. 1875. Notes sur les poissons fossiles. I-VI. Bulletin De La Société Géologique De France (serie 3) 3: 631–642.
Schwarzhans, W., G. Stringer, and R. Fricke. 2020. On the authorship of Actinopteri and Actinopterygii. Cybium 44: 7–8.
Shimada, K., R.E. Chandler, O.L.T. Lam, T. Tanaka, and D.J. Ward. 2017. A new elusive otodontid shark (Lamniformes: Otodontidae) from the lower Miocene, and comments on the taxonomy of otodontid genera, including the ‘megatoothed’ clade. Historical Biology 29: 704–714.
Siversson, M. 1992. Biology, dental morphology and taxonomy of lamniform sharks from the Campanian of the Kristianstad Basin, Sweden. Palaeontology 35: 519–554.
Siversson, M. 1995. Revision of the Danian cow sharks, sand tiger sharks, and goblin sharks (Hexanchidae, Odontaspididae, and Mitsukurinidae) from southern Sweden. Journal of Vertebrate Paleontology 15: 1–12.
Smith, R., T.H. Smith, and E. Steurbaut. 1999. Les élasmobranches de la transition Paléocène-Eocène de Dormaal (Belgique): Implications biostratigraphiques et paléobiogéographiques. Bulletin De La Société Géologique De France 170: 327–334.
Squires, R.L. 2001. Additions to the Eocene megafossil fauna of the Llajas Formation, Simi Valley, southern California. Contributions in Science 489: 1–40.
Squires, R.L., and J.L. Goedert. 1994. Macropaleontology of the Eocene Crescent Formation in the Little River Area, Southern Olympic Peninsula, Washington. Contributions in Science 444: 1–32.
Squires, R.L., and J.L. Goedert. 1995. New species of middle Eocene gastropods from the northern Doty Hills, southwestern Washington. The Veliger 38: 254–269.
Squires, R.L., J.L. Goedert, and K.L. Kaler. 1992. Paleontology and stratigraphy of Eocene rocks at Pulali Point, Jefferson County, eastern Olympic Peninsula, Washington. Washington Division of Geology and Earth Resources, Report of Investigations 31: 1–27.
Stone, N.R., and K. Shimada. 2019. Skeletal anatomy of the Bigeye Sand Tiger shark, Odontaspis noronhai (Lamniformes: Odontaspididae), and its implications for lamniform phylogeny, taxonomy, and conservation biology. Copeia 107: 632–652.
Tabor, R.W., and W.M. Cady. 1978. Geologic map of the Olympic Peninsula, Washington. U.S. Geological Survey Miscellaneous Investigations Series Map I-994, scale 1:125,000, 2 sheets.
Tanaka, T., Y. Fujita, and S. Morinobu. 2006. Fossil shark teeth from the Namigata Formation in Ibara City, Okayama Prefecture, central Japan. Bulletin of the Mizunami Fossil Museum 33: 103–109.
Tate, R. 1894. Unrecorded genera of the older Tertiary fauna of Australia, including diagnoses of some new genera and species. Journal and Proceedings of the Royal Society of New South Wales 27: 167–197.
Torres, M.E., G. Bohrmann, and T.E. Dubé. 2003. Formation of modern and Paleozoic stratiform barite at cold methane seeps on continental margins. Geology 31: 897–900.
Underwood, C.J., A. Goswami, G.V.R. Prasad, O. Verma, and J.J. Flynn. 2011a. Marine vertebrates from the “middle” Cretaceous (early Cenomanian) of South India. Journal of Vertebrate Paleontology 31: 539–552.
Underwood, C.J., D.J. Ward, C. King, S.M. Antar, I.S. Zalmout, and P.D. Gingerich. 2011b. Shark and ray faunas in the middle and late Eocene of the Fayum Area, Egypt. Proceedings of the Geologist’s Association 122: 47–66.
Verma, S.K. 2023. A new Bartonian elasmobranch assemblage from the Kutch Basin, western India, and its significance in the context of paleoclimate change. Historical Biology. https://doi.org/10.1080/08912963.2023.2238736.
Ward, D.J. 1978. Additions to the fish fauna of the English Palaeogene. 1. Two new species of Alopias (Thresher Shark) from the English Eocene. Tertiary Research 2: 23–28.
Ward, D.J. 1979. Additions to the fish fauna of the English Palaeogene. 3. A review of the hexanchid sharks with a description of four new species. Tertiary Research 2: 111–129.
Ward, D.J. 1980. The distribution of sharks, rays and chimaeroids in the English Palaeogene. Tertiary Research 3: 13–19.
Ward, D.J., and R.L. Wiest Jr. 1990. A checklist of Palaeocene and Eocene sharks and rays (Chondrichthyes) from the Pamunkey Group, Maryland and Virginia, USA. Tertiary Research 12: 81–88.
Weaver, C.E., and K.V.W. Palmer. 1922. Fauna from the Eocene of Washington. University of Washington Publications in Geology 1: 1–56.
Weiler, W. 1929. Ergebnisse der Forschungsreisen Prof. E. Stromers in den Wüsten Ägyptens. V. Tertiäre Wirbeltiere. 3. Die Mittel-und obereocäne Fischfauna Ägyptens mit besonderer Berücksichtigung der Teleostomie. Abhandlungen Der Bayerischen Akademie Der Wissenschaften Mathematisch-Naturwissenschaftliche Abteilung, Neue Folge 1: 1–57.
Wells, R., D. Bukry, R. Friedman, D. Pyle, R. Duncan, P. Haeussler, and J. Wooden. 2014. Geologic history of Siletzia, a large igneous province in the Oregon and Washington Coast Range: Correlation to the geomagnetic polarity time scale and implications for a long-lived Yellowstone hotspot. Geosphere 10: 692–719. https://doi.org/10.1130/GES01018.1.
Welton, B.J. 1972. Fossil sharks in Oregon. State of Oregon, Department of Geology and Mineral Industries. The Ore Bin 34: 161–170.
Welton, B.J. 1974. Preliminary note on the Paleocene elasmobranchs of the Lodo Formation, Fresno County, California. The Paleocene of the Panoche Creek – Cantua Creek Area, Central California, Society for Sedimentary Geology (SEPM) Fall Field Trip Guidebook: 91–97.
Welton, B.J., and W.J. Zinsmeister. 1980. Eocene neoselachians from the La Meseta Formation, Seymour Island, Antarctic Peninsula. Contributions in Science 329: 1–10.
Wheeler, W.H. 1966. A mosasaur mandible from the Black Creek Formation, Cretaceous, of North Carolina. The Journal of the Elisha Mitchell Scientific Society 82: 92–93.
White, E.I. 1956. The Eocene fishes of Alabama. Bulletins of American Paleontology 36: 123–150.
Winkler, T.C. 1874a. Mémoire sur quelques restes de poissons du système heersien. Archives Du Musée Teyler 4: 1–15.
Winkler, T.C. 1874b. Deuxième mémoire sur des dents de poissons fossiles du terrain bruxellien. Archives Du Musée Teyler 4: 16–48.
Yabumoto, Y., and T. Uyeno. 1994. Late Mesozoic and Cenozoic fish faunas of Japan. The Island Arc 3: 255–269.
Zhelezko, V.I., and V.A. Kozlov. 1999. Elasmobranhii i biostratigraphia paleogena Zauralia i Srednei Asii. Materialy Po Stratigrafii i Paleontologii Urala 3: 1–324 (in Russian).
Acknowledgements
We thank S.R. Benham (Pacific Lutheran University, Tacoma, Washington) and G.H. Goedert (deceased) for assistance with fieldwork during early stages of this project, and A.S. Fidler (Phoenix, Arizona) for assistance with fieldwork in 2023. We are grateful to L.S. Guthrie (Lacey, Washington) for providing a pneumatic tool used for preparing the shark teeth. Specimens were curated and numbers provided by K.L. Anderson (Vertebrate Paleontology Collections Manager, UWBM) and R.C. Eng (Invertebrate Paleontology, UWBM). We thank C. Sidor and G. Wilson (both UWBM) and staff of the U.S. Department of Agriculture, Forest Service (USFS), especially B. Schumacher (Lakewood, Colorado) and A. Pfeffer (Olympia, Washington) for facilitating permit requests in connection with this study. Fossils used in this study were collected under USFS permit (R6-OLYMPIC-MGM-FY23-001). This research was funded in part by the Austrian Science Fund (FWF) [P 33820] to JK, and in part by the Swedish Research Council (Vetenskapsrådet; 2016-03920) to SK. We are extremely grateful to J. Ebersole (McWane Science Center, Birmingham, Alabama) and an anonymous reviewer for their detailed suggestions which greatly improved this paper.
Funding
Open access funding provided by Swedish Museum of Natural History. This article is funded by Vetenskapsrådet, 2016-03920, Steffen Kiel, Austrian Science Fund, P 33820, Jürgen Kriwet.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare no competing interests.
Additional information
Handling Editor: Walter Joyce.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Goedert, J.L., Kiel, S., Thomas, E.J. et al. An early Eocene fish assemblage associated with a barite deposit in the lower part of the Crescent Formation, Olympic Peninsula, Washington State, USA. PalZ (2024). https://doi.org/10.1007/s12542-024-00692-y
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s12542-024-00692-y