
Abstract
The Carboniferous outcrops from Adarouch (central Morocco) are composed of siliciclastic and carbonate rocks, and the latter have yielded abundant fossils. The upper part of the marine succession in the Idmarrach Formation and its laterally equivalent Tirhela Formation belong to the Bashkirian. More recent investigations of poorly preserved coral assemblages from the upper part of Idmarrach 2 section and the upper part of the Tirhela Formation are here described. The assemblage from the upper Idmarrach Formation comprises several typical Mississippian taxa, such as Aulophyllum fungites, Dibunophyllum bipartitum, Koninckophyllum interruptum, Siphonodendron scaleberense and Lithostrotion decipiens, and some highly evolved forms previously undescribed of Mississippian genera such as Haplolasma sp., Arachnolasma sp., Siphonodendron sp., Diphyphyllum sp., Solenodendron sp., Clisiophyllum sp. and a new species Corwenia tirhelensis sp. nov. In addition, an undetermined petalaxid has been also recorded. Similarly, the assemblage from near the top of the Tirhela Formation yielded some species regarded usually as Mississippian, such as Palaeosmilia murchisoni and Lithostrotion decipiens. We also identified the long-ranging tabulate Syringopora sp., Corwenia tirhelensis sp. nov., and a single species of the fasciculate rugosan, Siphonodendron tindoufense, previously described from Bashkirian rocks in the Tindouf Basin (southern Morocco). The presence of the latter taxon in both the Idmarrach and Tirhela sections implies a communication between the Adarouch region and the Saharan basins. The occurrence of S. tindoufense at the approximately same stratigraphic level in Tindouf, Taoudenni, Reggan-Ahnet and Adarouch, demonstrates its important regional stratigraphic value in North Africa.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
The Carboniferous stratigraphic successions from Adarouch, in the Moroccan Central Meseta (Fig. 1), are composed both of siliciclastic and carbonate rocks, with the latter rich in diverse fossil groups such as foraminifers, algae, brachiopods, conodonts, bryozoans, crinoids and corals. Termier (1936) first described the stratigraphy of the Palaeozoic from the Moroccan Meseta. Termier and Termier (1950) mentioned the Tizra outcrops at Adarouch and described Dibunophyllum vaughani from that locality and Dedok and Hollard (1980) identified a probable Westphalian age for the Migoumess Formation, based on the presence of the brachiopod Brachythyrina strangwaisi. This dating was confirmed with foraminifers and algae by Berkhli and Vachard (2002), who rejected other datings. Termier et al. (1975) regarded the Adarouch outcrops as ranging in age from the Viséan to Serpukhovian. The first detailed stratigraphical and palaeontological studies were undertaken by Berkhli (1999), who defined three depositional sequences in the Carboniferous of Adarouch. In that region he defined the Oued Amhares Formation at the base, the Tizra and Mouarhaz formations in the middle and the Akerchi Formation at the top (Fig. 1). In a thrust sheet farther to the east, are the laterally equivalent Idmarrach and Tirhela formations. The continental siliciclastic rocks of the Migoumess Formation crop out further east (Berkhli and Vachard 2001, 2002). The coral assemblages from the Tizra, Akerchi and Idmarrach formations were originally described by Said (2005) in an unpublished Ph.D. thesis and later reported by Said et al. (2007). The aulophyllids from those formations were described by Said et al. (2008). The taphonomical and palaeoecological features of those assemblages and their environmental implications were the aim of two later papers (Said et al. 2010, 2011).

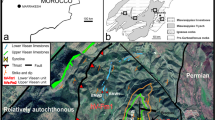
Modified from Cózar et al. (2011)
Location of the Adarouch area (encased box) in the Moroccan Meseta. The photogeological map shows the location of the studied stratigraphic sections.
Berkhli and Vachard (2001, 2002), based on foraminifers, assigned a Viséan age to the Oued Amhares, Tizra, Mouarhaz, lower Akerchi and Idmarrach formations, and a Serpukhovian age to the upper Akerchi Formation and the top of the Idmarrach Formation. However, Cózar et al. (2008, 2011) subsequently demonstrated that only the base of the Akerchi Formation is uppermost Viséan and that the Idmarrach Formation (and its equivalent, the Tirhela Formation) is almost entirely Serpukhovian, but reaching the Bashkirian in the upper beds. These revised age determinations were based on the analysis of foraminifers and conodonts and implies that the transition from a marine to continental environment in the Adarouch region happened later, in the Bashkirian, and not in the Serpukhovian, as previously inferred. The coral assemblages identified in the lower beds of both the Idmarrach and Tirhela formations comprise typical late Viséan and Serpukhovian species (Said 2005; Said et al. 2007; Rodríguez et al. 2015, 2016). However, the coral assemblages recorded at the top of the Idmarrach and Tirhela formations are poorly preserved and were not previously described. They come from a mixed siliciclastic calcareous facies that represents the transition to the Migoumess Formation that is composed of entirely siliciclastic continental rocks.
Although the corals in this present study are quite abundant and diverse, they are commonly dolomitized, silicified and in some cases partly decalcified. The importance of these assemblages is twofold: (1) they represent the last in situ marine fossils present in the Palaeozoic from the Adarouch region, and (2) being recorded in the lowermost Bashkirian, they may represent the transition from Mississippian to Pennsylvanian coral assemblages across the mid-Carboniferous boundary.
Coral assemblages
As stated above, the coral assemblages identified from the lower beds of the Idmarrach Formation were listed in Said et al. (2007). New sampling campaigns have been undertaken since then, and the description of corals collected from the Idmarrach and Tirhela formations was subsequently published (Rodríguez et al. 2015, 2016; Somerville et al. 2016). However, the assemblages recorded in the upper levels of the Idmarrach and Tirhela formations are very important from a biostratigraphic and palaeogeographic point of view and they are described herein (see Appendix Table 1 and Table 2 for list of specimens and locations). However, for those taxa identified previously from these sections, only brief comments are recorded to highlight diagnostic properties or unusual features. Comparisons have been made with rugose coral faunas documented from other Serpukhovian and younger locations in Morocco (Rodríguez et al. 2013) and Algeria (Semenoff-Tian-Chansky 1974, 1985; Aretz 2011; Atif et al. 2016).
Idmarrach formation
Corals are abundant in some levels of the Idmarrach Formation. Three combined logged sections (Idmarrach-1, -2, -3) represent the composite stratigraphy that was previously reported in Cózar et al. (2011). The section Idmarrach-1, and the lower levels of Idmarrach-2 yielded abundant corals that have been described by Said (2005) and listed by Said et al. (2007) and were regarded as Brigantian; the assemblages of section Idmarrach-3, and lower levels of Idmarrach-2 were described in more detail in Rodríguez et al. (2015, 2016), but the upper levels of Idmarrach 2 have been not described yet. The upper levels of Idmarrach-2 section in units 8–9 (Figs. 2, 3:1–2) show a marked and abrupt enrichment in siliciclastics and consequent generally poor preservation of the coral specimens. Nevertheless, the uppermost limestone beds of the formation (IDM2/8, 9; see Fig. 2) are quite rich in corals and yielded a mixed assemblage composed of solitary and colonial rugose corals. Although the diversity is quite high (12 genera), with 6 solitary genera and 6 colonial genera recorded, only a few of the 14 species are abundant (Appendix Table 1). The assemblage is paraautochthonous, because the corals are not in growth position, and many fasciculate colonies are fragmented, but most of the solitary corals show little abrasion (Fig. 3:3). As they have been recorded in a sandy and marly limestone that shows some extraclasts and local erosion surfaces, it is interpreted that the corals were living in the depositional area or nearby, but they were disturbed by periodic siliciclastic imputs. Most corals, mainly in unit, 9 show recrystallization and dolomitisation. The early Bashkirian age of units 8 and 9 is established by the presence of the conodont Declinognathodus noduliferus first recorded from the base of unit 8 (Cózar et al. 2011; Fig. 2).

adapted from Cózar et al. 2011)
Log of the upper part of the Idmarrach-2 Section (units 7–9), showing the coral distribution (

Idmarrach-2 section 1. General view of the section (units 3–8). 2. Upper part of the Idmarrach-2 section showing the boundary between units 8 and 9. 3. Detail of a bedding plane in unit 8 showing some solitary corals mainly Aulophyllum (black arrows)
The coral assemblage (Appendix Table 1) is composed of:
Aulophyllum fungites (Fleming, 1828) (Fig. 4:1–4)—It is quite common in units 7, 8 and 9, with 17 specimens collected. It has been also recorded in lower units of the Idmarrach-2 section. The specimens from Idmarrach show quite a high variability in size and thickening of skeletal structures. They show a smaller size than the typical representatives of the species and the dissepimentarium is also narrower. Those differences with older specimens may be explained, either, due to their evolutionary development or perhaps more likely to difficult (stressful) environmental conditions.

Solitary rugose corals of the upper beds from Idmarrach-2 Section exposing > 35 m of strata. 1–4 Aulophyllum fungites (Fleming, 1828). 1–3 specimen IDM2/8–1, 1: longitudinal section, 2: transverse section in juvenile stage, 3: transverse section in mature stage. 4. Specimen IDM2/8-13, longitudinal section 5. Dibunophyllum bipartitum McCoy, 1849, specimen IDM2/8-21, transverse section 6–7 Koninckophyllum interruptum Thomson and Nicholson, 1876, specimen IDM2/8-23, 6: transverse section, 7: longitudinal section 8–10 Arachnolasma sp. 8, specimen IDM2/8-22, transverse section 9, IDM2/9-14B, transverse section 10, IDM2/9–29, longitudinal section 11–13, Haplolasma aff. parvicarinatum Semenoff-Tian-Chansky, 1974, specimen IDM8/2–11, 11: transverse section, 12: longitudinal section 13: detail of the fibrous microstructure. 14. Clisiophyllum sp., specimen IDM2/9–26, transverse section 1 cm scale bar, same for all pictures, except Fig. 4:13 (1 mm scale bar)
Dibunophyllum bipartitum (McCoy, 1849) (Fig. 4:5)—Only one fragmentary specimen has been recorded. This species has a long stratigraphic record, being already recorded in the lower Bashkirian from Tindouf Basin (Rodríguez et al. 2013).
Koninckophyllum interruptum Thomson and Nicholson, 1876 (Fig. 4:6–7)—Only a single specimen recorded. It shows typical features of the nominal species.
Arachnolasma sp. (Fig. 4:8–10)—Five specimens have been recorded and all show a poor preservation that prevents a specific identification, but the generic diagnostic features such as the thick columella and small axial structure without well-defined boundaries are present in all specimens.
Clisiophyllum sp. (Fig. 4:13)—One single fragmentary specimen. It shows close similarities to Clisiophyllum sp. 1 of Said et al. (2008). It has been recorded at the top of the Tizra Formation (upper Brigantian), in the Akerchi Formation (uppermost Brigantian) and in lower levels of the Idmarrach Formation (Serpukhovian).
Haplolasma aff. parvicarinatum Semenoff-Tian-Chansky, 1974 (Fig. 4:11–12)—Only a single specimen recorded. Smaller than most species of the genus. Its size is comparable to the Bashkirian species from China H. lingwueense (Lee and Chu, 1934), but the Chinese species has a lower number of septa and lacks a cardinal fossula. It shows also clear similarities to the smallest specimens of the Viséan species from the Sahara, H. parvicarinatum and H. lamelliferum (see Semenoff-Tian-Chansky 1974). The microstructure of the septa is fibrous; consequently, it is closer to H. parvicarinatum, despite the fact that it does not show carinae in the dissepimentarium.
Siphonodendron tindoufense Rodríguez et al., 2013 (see details in the assemblage from Tirhela at a similar stratigraphical level)—Three specimens show a very poor preservation, but their features and size fit well with the species from the Tindouf Basin (Rodríguez et al. 2013).
Siphonodendron sp. (Fig. 5:1–2)—Two recorded specimens show an intermediate size and number of septa between the two Bashkirian species (S. tindoufense and S. ouarkzizense), recorded in the Tindouf Basin (Rodríguez et al. 2013) and share with them the main diagnostic features, mostly flat tabulae and discontinuous axial columella.

Colonial rugose corals of the upper beds from Idmarrach-2 section 1–2 Siphonodendron sp., specimen IDM2/8-25, 1: transverse section, 2: longitudinal section of one single corallite. 3–5 Lithostrotion decipiens McCoy, 1849, 3: specimen IDM2/9-32, transverse section 4: specimen IDM2/9-10A, oblique section showing interruptions in the growth. 5: Specimen IDM2/9-30B, longitudinal section of one corallite. 6 Diphyphyllum fasciculatum, Fleming, 1828, specimen IDM2/9-16, broken branches showing transverse and longitudinal sections. 7–8. Solenodendron furcatum Smith, 1925, specimen IDM2/9–20, 7: broken branches showing transverse section. Note the parricidal increase (upper right). 8: Detail of the deflected inner border of septa building the aulos. 9. Siphonodendron scaleberense Nudds and Somerville, 1987, specimen IDM2/9-11A, broken branch showing transverse section 10–13 Undetermined petalaxid, specimen IDM2/9-12. 10: longitudinal section, 11: detail of the longitudinal section showing periaxial cones (arrows), 12: transverse section; note the presence of lateral lamella (arrow), 13: oblique section; note the presence of lateral lamella (arrow). 1 cm scale bar, same for all pictures, except Fig. 5:10 (5 mm scale bar) and Fig. 5:8, (1 mm scale bar)
Siphonodendron scaleberense Nudds and Somerville, 1987 (Fig. 5:8)—Only one branching fragment of a fasciculate colony has been recorded in a debris bed (unit 9). It is smaller in size than the type material (9 mm in tabularium diameter), although the number of major septa (36) is the same. The cardinal fossula is conspicuous which distinguishes it from all other species in the genus. The minor septa do not usually reach the inner dissepimentarium/tabularium border, creating inosculate or herringbone-type inner dissepiments, a feature developed in S. multiradiale Nudds and Somerville, 1987, but differs from it in having less densely packed major septa. The only other comparable species is S. sociale (Phillips, 1836), but that species has fewer major septa (29–34) for corallites of the same size (see Nudds 1980; Poty 1981).
Lithostrotion decipiens (McCoy, 1849) (Fig. 5:3–5)—It is quite common in the last beds (unit 9) of the Idmarrach Formation. Seven specimens have been collected. They are small cerioid colonies that show some areas changing to phaceloid (subcerioid) surrounded by siliciclastic material mixed with micrite. Five specimens show a tabularium diameter and number of septa that are typical for the species (14–17 major septa and 2–3 mm in tabularium diameter). The other two specimens show slightly larger measurements (19–20 major septa and 3–3.5 mm in tabularium diameter) but are included in the same species.
Diphyphyllum fasciculatum (Fleming, 1828) (Fig. 5:6)—Several branching fragments of fasciculate colonies have been recorded in a debris bed (unit 9). They are broken and compressed. The features of the individual corallites (19–20 major septa, 4.5 mm in diameter, short septa, and absence of axial structure) fit well with those of the type species.
Solenodendron furcatum (Smith, 1925) (Fig. 5:7)—It has been recorded only as broken branches, and in the same debris bed as D. fasciculatum (unit 9). The individual corallites show 20–21 major septa, 4 mm in diameter, a well-developed aulos and parricidal increase. All these features are identical to the nominal species (see Somerville et al. 2016).
Corwenia tirhelensis sp. nov. (see details in the systematic section)—Several branches included in coral debris have been identified as belonging to the genus Corwenia. Their features are identical to those of specimens recorded in the Tirhela Formation (see below).
Undetermined petalaxid (Fig. 5:9–12)—One single colonial specimen. This coral has a fasciculate habit, with closely packed corallites. It shows similarities with several genera of the family Petalaxidae and also with corals of the family Lonsdaleiidae, but it does not fit completely with any of them. (See the systematic section).
Tirhela formation
The coral assemblages of the lower beds of the Tirhela Formation have been described in Rodríguez et al. (2015, 2016). Only the coral assemblage recorded in the upper sandy limestone beds (Figs. 6, 7:1) is described here. The upper beds of the Tirhela section (Fig. 1) represent transitional beds to continental siliciclastic beds of the Migoumess Formation, comprising thick red shales, calcareous sandstones and breccias (Fig. 6). Corals in the upper beds of the Tirhela Formation are abundant and can form biostromes (Fig. 7:2–3). However, the diversity is quite low (5 genera), with only four colonial genera and one solitary genus, and five species recorded (Appendix Table 2). In spite of the short distance (about one kilometre) between the Tirhela and Idmarrach sections (Fig. 1), and similar transitional marine to non-marine facies, the Tirhela and Idmarrach assemblages only have three rugose species in common, and the abundance of each species is different in both sections. The dominant species in Tirhela are Siphonodendron tindoufense and Corwenia tirhelensis sp. nov, that are very scarce in Idmarrach, and the most common species from the latter section (Aulophyllum fungites) is absent in the former (Appendix Table 2).

adapted from Cózar et al. 2011)
Log of the upper levels of the Tirhela section, showing the distribution of corals (

The Tirhela section 1. General view of Tirhela 2 section with steeply dipping beds to the south, showing massive and well-bedded limestones passing up to nodular bedded limestones and shales and sandy limestones (> 20 m thick). 2. Detail of one bed showing abundant fasciculate coral colonies. 3. Biostromal bed composed mainly of broken branches of Siphonodendron tindoufense Rodríguez et al., 2013
The coral assemblage (Appendix Table 2) is composed of:
Siphonodendron tindoufense Rodríguez et al., 2013 (Fig. 8:1–4)—It is particularly abundant, being the dominant species in some beds (Fig. 7:3). Most specimens are in growth position or almost. Eleven specimens have been collected. All of them show corallites with a discontinuous columella, horizontal to slightly conical tabulae, 3.5–4.5 mm in diameter and 18–22 major septa, which are identical features to the specimens from the Tindouf Basin (Rodríguez et al. 2013).

Corals from the upper carbonate beds at the Tirhela section 1–4 Siphonodendron tindoufense Rodríguez et al., 2013 1–2, specimen TIR2/2-1, 1: transverse section, 2: longitudinal section 3–4, specimen TIR2/2-7, 3: longitudinal section, 4: transverse section 5–6 Syringopora sp. 5: specimen TIR2/2-27, transverse section of a part of the colony. 6: specimen TIR2/2-23B, detail of the calyx of a corallite showing the presence of sclerites similar to those of Termieralcyon (see Fernández-Martínez et al., 2018). 7 Palaeosmilia murchisoni Milne-Edwards and Haime, 1848, specimen TIR2/2-28, transverse polished section 5 mm scale bar for Fig. 8:1–4, scale bar 2 mm for Fig. 8:5, scale bar 1 mm for Fig. 8:6 and scale bar 1 cm for Fig. 8:7
Corwenia tirhelensis sp. nov. (Fig. 9:1–8)—Also very abundant and being dominant in some beds. Twenty-four specimens have been collected. Some colonies are entire and in growth position, but some others are broken fragments showing only a few branches. (See the systematic section).

Corwenia tirhelensis sp. nov. 1: specimen IDM2/9-14A (paratype), transverse section 2: specimen TIR2/2-12 (paratype), transverse section 3–4, specimen TIR2/2-13, 3: longitudinal section, 4: transverse section 5–6: specimen TIR2/2-21 (holotytpe), 5: longitudinal section showing an offset, 6: transverse section 7–8, specimen TIR2/2-24 (paratype), 7: longitudinal section, 8: transverse section 5 mm scale bar for all pictures
Lithostrotion decipiens (McCoy, 1849) (see details in Idmarrach assemblage)—One single colony has been recorded in Tirhela showing a mean corallite diameter of 3 mm and 19–20 major septa.
Syringopora sp. (Fig. 8:5–6)—Two specimens that grew together with specimens of Corwenia sp. have been recorded. They have corallites 2 mm in diameter, show infundibuliform tabulae and common septal spines and show symbiosis with the alcyonarian Termieralcyon (Fernández-Martínez et al., 2018).
Palaeosmilia murchisoni Milne-Edwards and Haime, 1848 (Fig. 8:7)—Two very large specimens have been collected (more specimens have been identified in the field). The coralla are 70 and 85 mm in diameter and have 103 and 106 septa, respectively. Both specimens show a naotic peripheral dissepimentarium. They are the only solitary corals recorded in the upper beds of the Tirhela Formation.
Systematic palaeontology
Phylum Coelenterata Frey and Leuckart, 1847
Subphylum Cnidaria Hatschek, 1888
Class Anthozoa Ehrenberg, 1834
Subclass Rugosa Milne-Edwards and Haime, 1850
Family Aulophyllidae Dybowski, 1873
Genus Corwenia Smith and Ryder, 1926
Corwenia tirhelensis sp. nov.
Figure 9:1–8
Holotype. Specimen TIR2/2-21. Upper beds of the Tirhela Formation. All other specimens of the collection are regarded as paratypes.
Material. 27 specimens have been collected; 24 from Tirhela, 3 from Idmarrach (Appendix Table 2).
Location, horizon and age. Upper beds from the Tirhela and Idmarrach formations, Adarouch region, Moroccan Meseta. Lower Bashkirian.
Diagnosis. Fasciculate coral. The corallites have typically 34–37 major septa and 13.5–17 mm in diameter. The corallites show a large dibunophylloid axial structure, long septa that reach the axial structure, slightly or not thickened in the tabularium. Minor septa irregular or absent, substituted by angulate to inosculate dissepiments.
Description. Fasciculate coral showing densely packed corallites. The alar diameter of colonies varies between 13.5 and 17 mm, the maximum being 20 mm; the tabularium diameter of colonies typically varies between 9 and 12 mm, the maximum being 15 mm; the axial structure diameter of colonies varies between 3 and 5 mm, the maximum being 6 mm. The number of major septa varies between 34 and 37, the maximum being 39.
The wall is thin and smooth, but it may show slight local thickenings. Where corallites are in contact, the two walls are distinctly separated (Fig. 9:2, 6). The septa are long; most of them reach the axial structure and some of them are in continuity with radial lamellae. They are thin and irregular in the dissepimentarium, very slightly thickened in the outer tabularium and straight, slightly tapering axially (Fig. 9:2, 4, 6). The cardinal and the counter septa are not different from other septa. Both are joined to the median lamella (Fig. 9:1, 4, 6). The cardinal fossula is absent. The minor septa are short and irregular or absent (Fig. 9:4), only in the specimens from Idmarrach are there some short minor septa that reach the dissepimentarium/tabularium boundary (Fig. 9:1). In juvenile, stages the septa are somewhat shorter and only the cardinal and the counter septa are joined to a simple median lamella (Fig. 9:2, 8). The axial structure is typically dibunophylloid, composed of a somewhat thickened median lamella, 6–10 thin and irregular radial lamellae, and abundant, densely packed conical tabellae. It occupies about 1/3–1/4 of the diameter (Fig. 9:1, 2, 4, 6). The dissepimentarium occupies 1/3–1/4 of the radius. In mature stages, it is composed of 3–7 rows of angulate to inosculate, less commonly regular dissepiments (Fig. 9:2, 6). The inner row may be slightly thickened (Fig. 9:1, 4). In longitudinal section, they are globose to slightly elongated and very steeply declined (60–90°, Fig. 9:3, 5, 7). The tabulae are thin, incomplete and divided in two series of tabellae. The axial tabellae are conical, convex and declined 30°–60° to the periphery; 5–8 of them are intersected in transverse section. The periaxial tabellae are also convex, but less densely packed than the axial tabellae, and slightly less steeply declined to the periphery (Fig. 9:3, 7). The microstructure of the wall is microlamellar. The tabulae and dissepiments are microgranular with fibrous thickenings. The septa show mesoplasm with fibers organized in fascicles and fibronormal stereoplasm.
Remarks. The specimens from Idmarrach show slight differences with those from Tirhela: they are slightly larger, the minor septa are better developed and the dissepimentarium is somewhat narrower. But all other features are identical and those differences are considered as local variations and adaptations.
The features of this species do not fit with any previously described species of the genus. It is similar to the type species Corwenia rugosa (McCoy, 1849) in the dibunophylloid axial structure, in the length and thickness of septa and in the structure of the tabularium, but it is larger, possesses more septa and lacks a conspicuous cardinal fossula. Other species are even smaller, including those recorded in the Bashkirian (Corwenia longiseptata Fomichev, 1953, Corwenia cantabrica De Groot, 1963). Only the Serpukhovian species C. densicolumellata Bykova, 1966 shows a similar size (even larger) and number of septa, but it differs clearly in having a smaller axial structure and thicker median plate. The Brigantian unnamed species (Corwenia sp.) described by Said et al. (2008) from the Tizra Formation shows also a similar size, but the number of septa is lower, the axial structure is also smaller and more irregular and the juvenile stages lack an axial structure.
Family Petalaxidae Fomichev, 1953
Gen. et sp. unknown
Figure 5:9–12
Material. A single specimen from Idmarrach IDM2/9-12 (Appendix Table 1).
Description. Phaceloid colony. Corallites closely spaced and irregularly distributed; wall generally thin, sinuous in longitudinal section, somewhat thickened by septal bases, septal furrows well marked. In mature stage (Fig. 5:11, 12) number of septa normally 22–23, at corallite diameter 4.5–5.0 mm; major septa long, reaching or almost the axial structure, slightly thickened near the wall and in the outer part of the tabularium, radial, commonly interrupted by lonsdaleoid dissepiments; cardinal septum straight, slightly thickened, connected to median lamella of axial structure, but locally detached from it; minor septa developed only in adult stages, short, thickened at wall, adaxially tapered, forming ridges on wall in early adult stages, becoming slightly longer in maturity and just penetrating the outermost tabularium, discontinuous in dissepimentarium. Axial structure well developed, with variably thickened median lamella, bearing few short septal lamellae and axial tabellae. Dissepiments lonsdaleoid forming almost complete rings in some corallites (Fig. 5:11, 12). In longitudinal section, large dissepiments, moderately convex, steeply inclined in one or more rarely two rows. Thick deposit on their inner border. Tabulae moderately spaced (8–10 in 1 cm corallite length), thin, complete, concave or planar, moderately elevated adaxially, more strongly elevated at median lamella, which is irregular and sinuous. Periaxial cones well developed (Fig. 5:10).
Remarks. It is a phaceloid coral that shows many typical features of the family Petalaxidae. However, the only fasciculate genera included in that family (Hill 1981; Bamber et al. 2017) are Lytvophyllum, and Schoenophyllum.
Lytvophyllum is known from the early Bashkirian in Donets and Novaya Zemlya (Kossovaya 1996), but a recent paper by Fedorowski (2021) stated that the Ukrainian specimens and probably also those from Novaya Zemlya are solitary or protocolonial and included them in his new family Krinkaphyllidae. It lacks the slightly complex axial structure, the lateral lamellae and the periaxial cones that are present in the specimen from Idmarrach. Some species of the genus Schoenophyllum show similar development of the lonsdaleoid dissepimentarium and densely packed corallites, but they have a more simple axial structure and a very peculiar lateral increase that distinguish that genus from our specimen. A close genus is Cystolonsdaleia, to which this specimen was previously tentatively assigned (Rodríguez et al. 2016) which shows also all those features, but it is cerioid. The specimen from Idmarrach has very densely packed corallites and in part of the colony some of them share walls showing a subcerioid habit, but it is clearly fasciculate, not massive. It could be regarded as a derivation of Cystolonsdaleia that developed a fasciculate habit. Alternatively, it could be considered as a possible ancestor of cerioid Cystolonsdaleia. It could be also related to the family Lonsdaleiidae, because most of the features present in the specimen from Idmarrach are shared with Lonsdaleia and its subgenus Serraphyllum, which shows intermediate features between fasciculate and cerioid habitus. Hecker (2012) demonstrated the presence of periaxial cones and a biform tabularium in these genera. However, the axial structure of Lonsdaleia is more complex and it does not possess lateral lamellae, which is a typical feature of the Petalaxidae. If the specimen from Idmarrach belongs to the Petalaxidae, it would represent the first record of that family in North Africa, and could imply certain biogeographical relationships with the eastern European Basins (Donets, Moscow, Ural Mountains), as well as the Cantabrian Mountains (N. Spain), where representatives of this family are common in the Bashkirian (De Groot 1963; Rodríguez 1984). Some Petalaxidae have been also described from the Mississippian and Pennsylvanian from North America (Bamber and Fedorowski 1998; Bamber et al. 2017; Sando 1983).
Discussion
The coral assemblages recorded in the uppermost rocks of the Tirhela and Idmarrach formations are composed mainly of rugose species, usually regarded as Mississippian. However, as the Bashkirian age of those rocks is well established using foraminifers and conodonts (see Cózar et al. 2011), it implies that some of the species are being recorded for the first time in the Bashkirian: Lithostrotion decipiens, Aulophyllum fungites, Koninckophyllum interruptum, Diphyphyllum fasciculatum, Siphonodendron scaleberense, Solenodendron furcatum and the genus Clisiophyllum extending their stratigraphical distribution to the lower Bashkirian. In addition, some other species and genera typically recorded in the Mississippian (Palaeosmilia murchisoni, Dibunophyllum bipartitum, Arachnolasma sp.) were previously recorded in the lower Bashkirian from the Tindouf Basin (Rodríguez et al. 2013; Cózar et al. 2014b) and hence their Bashkirian distribution is confirmed here. The rugose genera Corwenia and Haplolasma and the tabulate Syringopora have long stratigraphical ranges and are common in Bashkirian rocks (Lee and Chu 1934; Gorsky 1951; De Groot 1963; Rodríguez 1984; Fedorowski 2004, etc.).
The presence of many Mississippian-type corals in the Bashkirian rocks from Adarouch confirm the presence of a refuge for corals in North Africa (Cózar et al. 2014a) and indicates that it was not restricted to the Saharan basins, but also expanded to northern North African basins (Adarouch area of central Morocco Meseta). There were also other similar refuges where Mississippian corals reached the Bashkirian: Palaeosmilia and Diphyphyllum in Novaya Zemlya (Kossovaya 1996), Dibunophyllum, Palaeosmilia, Lonsdaleia and Tizraia in Ellesmere Island (Fedorowski et al. 2012) and Dibunophyllum, Caninostrotion, Nemistium and Kizilia in the Cantabrian Mountains (Rodríguez 1984).
The species Siphonodendron tindoufense was found in a similar stratigraphical level at the top of the marine carbonate succession in several Saharan basins: Reggan-Ahnet, Taoudenni (Semenoff-Tian-Chansky 1985), Tindouf (Fabre 1955; Rodríguez et al. 2013). Semenoff-Tian-Chansky (1985) proposed an identical stratigraphical position for the Siphonodendron tindoufense beds in all those basins, as uppermost Serpukhovian. As it was demonstrated that in Tindouf it is lower Bashkirian, the age in Reggan-Ahnet and Taoudenni must be checked; but in those basins, as in Tindouf, the cited species is related to the brachiopod-rich Titanaria beds and a similar if not identical stratigraphic position could be assumed. Consequently, it could be considered as an index fossil for the lower Bashkirian in North Africa. The presence of that species indicates also a communication between the Sahara and the Moroccan Meseta up to and including the Bashkirian, in spite of the rising of the Atlas Mountains, Reguibat Shield and Ougarta High (Cózar et al. 2014c). Some other species are also recorded in both the Tindouf Basin and Adarouch: Dibunophyllum bipartitum and Palaeosmilia murchisoni, but it is not significant, since both species are cosmopolitan. On the other hand, some of the most common species in the Bashkirian from the Tindouf Basin, such as Axophyllum moroccoense and Actinocyathus sarytschevae are not present in Adarouch and Corwenia and Syringopora that are common in Adarouch have not been recorded in the Bashkirian from the Tindouf Basin.
Conclusions
The coral assemblages from the upper part of the Idmarrach and Tirhela formations, which have been identified as lower Bashkirian age based on foraminifers and conodonts, comprise some typical Mississippian species, such as Aulophyllum fungites, Dibunophyllum bipartitum, Koninckophyllum interruptum, Lithostrotion decipiens, Siphonodendron scaleberense, Solenodendron furcatum, Diphyphyllum fasciculatum and Palaeosmilia murchisoni. In addition, some unidentified species of typical Mississippian genera have been also recorded (Arachnolasma sp., Clisiophyllum sp.). It implies the extension of the stratigraphical ranges of those genera and species. The long-ranging genera Syringopora, Corwenia and Haplolasma are also recorded, as well as a new species of Corwenia, C. tirhelenese sp. nov.
In addition, a single colonial specimen identified as belonging to the family Petalaxidae has been recorded in the Idmarrach Formation. It could represent an undescribed genus; the family Petalaxidae, is common in the Bashkirian from the Western Palaeotethys (Eastern European basins and Cantabrian Mountains), indicating a possible connection with those basins.
The abundance of the species Siphonodendron tindoufense that has been previously described from Bashkirian rocks in the Saharan Tindouf Basin, has a twofold importance: (1) This species may be regarded as an index fossil for the lower Bashkirian in North Africa (to be confirmed in the Reggan-Ahnet and Taoudenni basins), (2) it proves a communication between the Saharan Basins and the Moroccan Meseta during the Bashkirian.
References
Aretz, M. 2011. Corals from the Carboniferous of central Sahara (Algeria): The collection “Marie Legrand-Blain.” Geodiversitas 33: 581–624.
Atif, K.F.T., M. Aretz, M. Legrand-Blain, A. Bouzid, and M. Aimouche. 2016. Brachiopods and rugose corals in an upper Serpukhovian (Mississippian) biostrome: Preliminary results from the Djebel Arhlal (Béchar Basin, Algeria). Boletín Geológico y Minero 127(2/3): 345–360.
Bamber, E.W., and J. Fedorowski. 1998. Biostratigraphy and systematics of upper Carboniferous cerioid rugose corals, Ellesmere Island, Arctic Canada. Geological Survey of Canada Bulletin 511: 1–127.
Bamber, E.W., S. Rodríguez, B.C. Richards, and B.L. Mamet. 2017. Uppermost Viséan and Serpukhovian (Mississippian) rugose corals and biostratigraphy, Canadian Cordillera. Palaeontographica Canadiana 36: 1–169.
Berkhli, M. 1999. Sédimentologie, biostratigraphie et stratigraphie séquentielle du NE de la Méséta occidentale marocaine pendant le Carbonifère inférieur (Viséen-Serpoukhovien), 1–290. Unpublished Thèse État, Université Moulay Ismaïl de Meknès.
Berkhli, M., and D. Vachard. 2001. New biostratigraphical data from the Early Carboniferous sequences of the Adarouch area (NE Central Morocco). Newsletter on Stratigraphy 39: 33–54.
Berkhli, M., and D. Vachard. 2002. Le Carbonifère du Maroc central: Les formations de Migoumess, de Tirhela et d’Idmarrach. Lithologie, biostratigraphie et conséquences géodynamiques. Comptes Rendus Geoscience 334: 67–72.
Bykova, M.S. 1966. Niznekamennougolnye korally vostocnogo kazakstana. Trudy Instituta Geologicheski Nauk Kazakstan, Institut Geologicheski Kazakskoi SSR 1966: 1–214.
Cózar, P., D. Vachard, I.D. Somerville, M. Berkhli, P. Medina-Varea, S. Rodríguez, and I. Said. 2008. Late Viséan-Serpukhovian foraminiferans and calcareous algae from the Adarouch region (central Morocco), North Africa. Geological Journal 43: 463–485.
Cózar, P., I. Said, I.D. Somerville, D. Vachard, P. Medina-Varea, S. Rodríguez, and M. Berkhli. 2011. Potential foraminiferal markers for the Visean-Serpukhovian and Serpukhovian-Bashkirian boundaries—a casestudy from Central Morocco. Journal of Paleontology 85: 1105–1127.
Cózar, P., D. Vachard, I.D. Somerville, P. Medina-Varea, S. Rodríguez, and I. Said. 2014a. The Tindouf Basin, a marine refuge during the Serpukhovian (Carboniferous) mass extinction in the northwestern Gondwana platform. Palaeogeography, Palaeoclimatology, Palaeoecology 394: 12–28.
Cózar, P., P. Medina-Varea, I.D. Somerville, D. Vachard, S. Rodríguez, and I. Said. 2014b. Foraminifers and conodonts from the late Viséan to early Bashkirian succession in the Saharan Tindouf Basin (southern Morocco): Biostratigraphic refinements and implications for correlations in the western Palaeotethys. Geological Journal 49: 271–302.
Cózar, P., A. García-Frank, I.D. Somerville, D. Vachard, S. Rodríguez, P. Medina-Varea, and I. Said. 2014c. Lithofacies and biostratigraphical correlation of marine Carboniferous rocks in the Tindouf Basin, NW Africa. Facies 60: 941–962.
De Groot, G.E. 1963. Rugose corals from the Carboniferous of Northern Palencia (Spain). Leidse Geologische Mededelingen 29: 1–123.
Dedok, T.A., and H. Hollard. 1980. Brachiopodes du Carbonifère inférieur du Maroc central. Notes et Memoires du Service Géologique du Maroc 285: 185–230.
Dybowski, W.N. 1873. Beschreibung einiger neuen oder wenig bekennten Arten der Zoantharia rugosa aus der Kohlenformation Russlands. Zapadnogo Imperialnogo Minerali Obchestva 8: 102–116.
Ehrenberg, C.G. 1834. Beitrage zur physiologischen Kenntniss der Corallenthiere im allgemeinen, und besonders des rothen Meeres, nebst einem Versuche zur physiologischen Systematik derselben. Physiologische Abhandlungen der Königlichen Akademie der Wissenschaften zu Berlin 1832: 225–380.
Fabre, J. 1955. Deux polypiers du Viséen dupérieur de Tindouf (Sahara N. W.): Caninophyllum archiaci (M.E. et H.) var. densa n. var. et Carcinophyllum coronatum nov. sp. Annales de la Société Géologique du Nord 75: 8–15.
Fedorowski, J. 2004. Considerations on most Rugosa and the Dividocorallia from de Groot’s (1963) collection. Scripta Geologica 127: 71–311.
Fedorowski, J. 2021. Bashkirian Rugosa (Anthozoa) from the Donets Basin (Ukraine). Part 10. The family Krynkaphyllidae fam. nov. Acta Geologica Polonica 71: 53–101.
Fedorowski, J., E.W. Bamber, and D.V. Baranova. 2012. An unusual occurrence of Bashkirian (Pennsylvanian) rugose corals from the Sverdrup Basin, Arctic Canada. Journal of Paleontology 86: 979–995.
Fernández-Martínez, E., I. Coronado, S. Rodríguez, F. Tourneur, and M. Badpa. 2018. Alcyonacea awakens: palaeobiology and palaeoecology of Palaeozoic octocorals known from their sclerites. Geological Journal 54(6): 3593–3618.
Fleming, J. 1828. A history of British animals, 1–565. Edinburgh: Beel and Bradfute.
Fomichev, V.D. 1953. Rugose corals and stratigraphy of middle and upper Carboniferous and Permian deposits of Donets basin. Trudy Vsesoyuznogo Nauchno-Issledovatelskogo Geologicheskogo Instituta 1953: 1–622. (in Russian).
Frey, H., and R. Leuckart. 1847. Lehrbuch der Anatomie der wirbellosen Thiere, 1–626. Leipzig: L. Voss.
Gorsky, I.I. 1951. Kamennougolnye i permskie korally Novoj Zemli. Trudy Nauchno-Issledovatel’skogo Instituta Geologicheski Arktiki 32: 1–168.
Hatschek, B. 1888. Lehrbuch der Zoologie, eine morphologische Übersicht des Thierreiches zur Einführung in das Studium dieser Wissenschaft, 1–432. Jena: Gustav Fischer.
Hecker, M.R. 2012. Biform tabularia and periaxial cones in Lonsdaleia McCoy, 1849 (Rugosa). Geologica Belgica 15(4): 304–307.
Hill, D. 1981. Rugosa and Tabulata. In Treatise on Invertebrate Paleontology Part F (Supplement 1), ed. C. Teichert, 1–762. Boulder, Colo.: Geological Society of America and Lawrence, Kans.: University of Kansas Press.
Kossovaya, O.L. 1996. The mid-Carboniferous rugose recovery. Geological Society of London, Special Publications 102: 187–199.
Lee, C., and S. Chu. 1934. Note on the stratigraphy of the environs of the Maping City, Central Kwangsi. Bulletin of the Geological Society of China 13(2): 215–232.
McCoy, F. 1849. On some new genera and species of Palaeozoic Corals and Foraminifera. Annals and Magazine of Natural History 2(3): 1–20.
Milne-Edwards, H., and J. Haime. 1848. Recherches sur les polypiers, Premier général. Annales des Sciences Naturelles 9: 37–89.
Milne-Edwards, H., and J. Haime. 1850–1855. A Monograph of the British fossil corals,1–71 (1850); 147–210 (1852); 211–244 (1853); 245–299 (1855). London: Palaeontographical Society.
Nudds, J.R. 1980. An illustrated key to the British Lithostrotionid corals. Acta Paleontologica Polonica 25: 385–394.
Nudds, J.R., and I.D. Somerville. 1987. Two new species of Siphonodendron (Rugosa) from the Viséan of the British Isles. Proceedings of the Yorkshire Geological Society 46(4): 293–300.
Phillips, J. 1836. Illustrations of the geology of Yorkshire, Part 2: The Mountain Limestone district, 1–253. London: John Murray.
Poty, E. 1981. Recherches sur les tetracoralliares et les heterocoralliaires du viséen de la Belgique. Mededelingen Rijks Geologische Dienst 35(1): 1–161.
Rodríguez, S., I.D. Somerville, I. Said, and P. Cózar. 2013. Mississippian-like rugose corals from a Bashkirian biostrome in the Tindouf Basin, S. Morocco. Spanish Journal of Palaeontology 28(2): 253–282.
Rodríguez, S., I.D. Somerville, P. Cózar, I. Coronado, and I. Said. 2015. Description of the Serpukhovian rugose and tabulate corals from Idmarrach and Tirhela Formations (Adarouch, Morocco). Boletin Real Sociedad Española de Historia Natural, Sección Geológica 109: 71–101.
Rodríguez, S., I. Said, I.D. Somerville, P. Cózar, and I. Coronado. 2016. Serpukhovian coral assemblages from Idmarrach and Tirhela Formations (Adarouch, Morocco). Paleontologica Belgica 19(1–2): 29–42.
Rodríguez, S. 1984. Corales Rugosos del Carbonífero del Este de Asturias, 1–529. Unpublished Ph.D. Thesis, Universidad Complutense de Madrid 109/84.
Said, I., and S.S. Rodríguez. 2008. Descripción de los corales aulophyllidos del Viseense y Serpujoviense del área de adarouch (Marruecos). Coloquios de Paleontología 58: 13–40.
Said, I., S. Rodríguez, M. Berkhli, P. Cózar, and A. Gómez-Herguedas. 2010. Environmental parameters of a coral assemblage from the Akerchi formation (Carboniferous), Adarouch Area, central Morocco. Journal of Iberian Geology 36(1): 7–19.
Said, I., S. Rodríguez, I.D. Somerville, and P. Cózar. 2011. Environmental study of coral assemblages from the upper Viséan Tizra Formation (Adarouch area, Morocco): Implications for Western Palaeotethys biogeography. Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen 260(1): 101–118.
Said, I., M. Berkhli, and S. Rodríguez. 2007. Preliminary data on the coral distribution in the Upper Visean (Mississippian) succession from Adarouch area (NE Central Morocco). In Fossil Corals and Sponges. Proceedings of the 9th International Symposium on Fossil Cnidaria and Porifera, eds. B. Hubmann, and W. Piller. Schriftenreihe der Erdwissenschaftlichen Kommission 17: 353–364.
Said, I. 2005. Estudio de los corales rugosos con disepimentos del Mississippiense del NE de la meseta marroquí (sectores de Adarouch y Agouaraï), 1–240. Unpublished Ph.D. Thesis, Universidad Complutense de Madrid.
Sando, W.J. 1983. Revision of Lithostrotionella (Coelenterata, Rugosa) from the Carboniferous and Permian. Geological Survey Professional Papers 1247: 1–73.
Semenoff-Tian-Chansky, P. 1974. Recherches sur les Tétracoralliaires du Carbonifére du sahara Occidental. Editions du Centre Nationale de la Recherche Scientifique (Sér. 6, Sciences de la Terre) 30: 1–316.
Semenoff-Tian-Chansky, P. 1985. Corals. In The Carboniferous of the World, II, Australia, Indian Subcontinent, South Africa, South America and North Africa, eds. R.H. Wagner, C.F. Winkler-Prins, and L.F. Granados. IUGS Publication 20: 374–381.
Smith, S. 1925. The genus Aulina. Annals and Magazine of Natural History 16: 485–496.
Smith, S., and T.A. Ryder. 1926. The genus Corwenia, gen. nov. Annals and Magazine of Natural History 9: 149–159.
Somerville, I.D., S. Rodríguez, and I. Said. 2016. Carboniferous aulate corals from Azrou-Khenifra Basin (Morocco): Distribution and phylogenetic relationships. Geobios 49(4): 303–317.
Termier, H. 1936. Etudes Geologiques sur le Maroc Central et le Moyen Atlas septentrional. Notes et Memoires du Service Géologique du Maroc 33: 1–1566.
Termier, G., and H. Termier. 1950. Paleontologie marocaine II. Invertebres de l’Ere Primaire. Foraminiferes, spongoaires et coelenteres. Notes du Service Geologique du Maroc 73: 1–220.
Termier, G., H. Termier, and D. Vachard. 1975. Recherches micropaléontologiques dans le Paléozoïque du Maroc central. Cahiers de Micropaléontologie 4: 1–99.
Thomson, J., and H.A. Nicholson. 1876. Contributions to the study of the chief generic types of the Palaeozoic corals. Annals and Magazine of Natural History 17(2): 123–128 (17(4): 290–305; 17(6): 451–461; 18(7): 68–73).
Acknowledgements
The present research has been carried out with the funds provided by the Research project CGL2016-78738BTE of the Spanish Ministerio de Ciencia, Innovación y Universidades. This article is a contribution to the IGCP 596. The authors thank the cooperation of the Department of Mines and Development of the Ministère de l’Energie et des Mines of Morocco. The thin sections used for this study have been prepared by Isabel Díaz Mejías. We are grateful for the comments provided by the reviewers Julien Denayer and Jerzy Fedorowski which helped improved the paper.
Funding
Open Access funding provided thanks to the CRUE-CSIC agreement with Springer Nature.
Author information
Authors and Affiliations
Corresponding author
Additional information
Handling Editor: Markus Aretz.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Rodríguez, S., Said, I., Somerville, I.D. et al. Coral assemblages of the Serpukhovian–Bashkirian transition from Adarouch (Morocco). PalZ 97, 847–860 (2023). https://doi.org/10.1007/s12542-021-00586-3
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12542-021-00586-3