
Abstract
The seagrass Posidonia oceanica forms extensive meadows in the Mediterranean Sea. Studies on their associated highly diverse invertebrate assemblages are limited to the western Mediterranean. The eastern Mediterranean, however, is a basin undergoing rapid change due to the synergistic effects of climate warming, biological invasions and other human stressors that are driving native biodiversity to regional-scale collapses. We here surveyed the shelled molluscan assemblage of a Posidonia oceanica meadow in Plakias, south-western Crete, the first such study in the eastern Mediterranean Sea. This area has increased its yearly mean temperature by 1 °C in the last 20 years and is under heavy pressure by Lessepsian species. We sampled across a 5- to 20-m depth gradient, in two seasons to capture intra-annual variation and the leaf and rhizome strata separately. Against our expectations, the molluscan assemblage proved to be highly diverse, with species richness, dominant species and trophic guilds comparable to healthy western Mediterranean ones, and with a negligible non-indigenous component. The diversity of the native community (following the biotic resistance hypothesis) and oxygen supersaturation in the meadow may cause greater resistance to biological invasions and warming, respectively, suggesting that Posidonia oceanica meadows may act as a precious refugium for native biodiversity in the fast changing eastern Mediterranean Sea.
Similar content being viewed by others


Introduction
Biological invasions and global warming are among the main pressures affecting marine biodiversity in the eastern Mediterranean Sea. After the opening of the Suez Canal in 1869, hundreds of species entered the basin and established abundant populations (Galil 2009; Zenetos et al. 2012; Nunes et al. 2014) in the so-called Lessepsian invasion. Some species have exerted major impacts on native biota. For example, rabbitfishes overgraze algae (Sala et al. 2011) and the lionfish Pterois miles disrupts local marine communities (Savva et al. 2020). Additionally, the seawater temperature in the Mediterranean Sea has been increasing for the last few decades, but the increase in the Aegean and Levantine Seas has been the highest in the Mediterranean reaching 0.048 ± 0.006 °C/year over 1982–2018 (Pisano et al. 2020). This abrupt increase has likely triggered the disappearance of native species from the warmest areas like the Israeli shelf (Yeruham et al. 2015; Rilov 2016; Albano et al. 2021a). In such most affected areas, biological invasions and climate warming are acting in concert substantially modifying not only taxonomic composition but also ecosystem functioning (Rilov et al. 2018; Peleg et al. 2019; Yeruham et al. 2019; Steger et al. 2021a,b).
The South Aegean is in a special position in the eastern Mediterranean Sea because it bridges the Levantine Sea, its easternmost sector and the most affected by the Lessepsian Invasion and seawater warming, with the Ionian Sea. It is among the most invaded areas in Greece (Zenetos et al. 2011). A review of non-indigenous species in Crete is not available, but the recent demographic explosion of invasive Lessepsian species such as Pinctada radiata and Fistularia commersonii (Zenetos et al. 2008; Zenetos 2015) shows that this island is heavily affected similarly to other areas in the eastern Mediterranean (e.g. Dimitriadis et al. 2020).
The Mediterranean endemic plant Posidonia oceanica forms meadows that host a very high biodiversity and productivity (Boudouresque et al. 2006). However, research on its assemblages has been conducted mostly in the western Mediterranean (Idato et al. 1983; Templado 1984; García-Raso 1990; Gambi et al. 1992, 1995; Belgacem et al. 2011; Albano and Sabelli 2012; Urra et al. 2013; Bedini et al. 2015) and in the Adriatic Sea (Solustri et al. 2002; Beqiraj et al. 2008). Molluscs in particular have never been surveyed quantitatively in Posidonia oceanica meadows in the eastern part of the basin, notwithstanding they are one of the most diverse taxa in the Mediterranean.
We here describe a molluscan assemblage from a Posidonia oceanica meadow in Plakias, south-western Crete, along a depth transect from 5 to 20 m. We sampled in spring and autumn, to capture intra-annual variation, and both the leaves and the rhizomes. The latter have often been neglected but host the richest Posidonia oceanica assemblages (Albano and Sabelli 2012; Albano and Stockinger 2019). We expected a poor native assemblage and diverse and abundant populations of non-indigenous species, but our results suggest the opposite, calling for stronger conservation measures of Posidonia oceanica meadows in this over-stressed basin.
Materials and methods
Study area and sampling methods
The study was carried out in the bay of Plakias, on the south-western coast of Crete, (35.1796° N, 24.3957° E, Figure 1). The bay is 1.5 km long and extends from NNW to SSE. It is bordered on the south side by a long rock wall called “Paligremnos Wall”, which extends to the south-west into the sea for approximately 800 m. At the bottom of this wall, a sandy substrate stretches over the entire bay partly covered with a Posidonia oceanica meadow. The meadow is mostly found parallel to the wall, starting patchily at just 30-cm water depth, and growing into a dense meadow at a depth of 5 m continuing to reach more than 30 m. In the year of sampling, the sea surface temperature in the bay ranged from a minimum of 15 °C in March to a maximum of 27 °C in August.

Geographic location of the study site Plakias, south-western Crete, in the eastern Mediterranean Sea (left) and detail of the sampling area in Plakias Bay (right), where the dashed line indicates the area covered by the Posidonia oceanica meadow. Squares mark the four different stations (5 m, 10 m, 15 m, 20 m depth)
Samples were taken while SCUBA diving from both the leaf and the rhizome strata (Figure 2) of the meadow at four different depths (5 m, 10 m, 15 m, 20 m) in May and in September 2017 to capture intra-annual variation. For the leaf stratum, samples were collected using a hand-operated net according to the technique described by Ledoyer (1962), modified and standardized by Russo et al. (1985). The hand net consists of a metal frame (40 × 20 cm) and has a 500-μm mesh. For each replicate, 60 strokes were given against the base of the leaves; the net is then pulled upwards to collect the mobile fauna crawling on the leaves. At every depth, we collected two and four replicates in spring and in autumn, respectively.

The Posidonia oceanica meadow at Plakias, south-western Crete. a The leaf stratum; b The rhizome stratum; note the abundance of molluscs: the vetigastropod Jujubinus exasperatus (top right white cone-shaped shell, grazing on a dead Posidonia leaf) and the nudibranch Felimida luteorosa (violet and yellow, in the centre of the image, identification courtesy G. Furfaro) have been captured by this randomly shot photo
The rhizome stratum was sampled with an air-lift suction sampler that is 1 m long and 8 cm in diameter (Templado et al. 2010). This device consists of a PVC tube on which a net with a mesh size of 500 μm is mounted at one end. A SCUBA tank was attached to the pipe to supply air for the suction force. Sampling was carried out on three 1-m2 quadrats at each depth and season, after defoliation to improve sampling efficacy (Bonfitto et al. 1998). Standardization of sampling intensity was performed by using 100 bar of a 12-l steel tank for each replicate. At each 1-m2 replicate, the shoot density of the Posidonia oceanica meadow was counted on a 40 × 40 cm sub-quadrat (Panayotidis et al. 1981) and the meadow status assessed according to UNEP/MAP-RAC/SPA (2015).
Back in the lab, the samples were kept in saltwater in order to keep the organisms alive and sieved with a 1-mm mesh size. Living shelled molluscs were picked under a stereomicroscope, identified to species level and fixed in 96% ethanol.
Data analysis
We computed sample coverage and estimated richness at perfect coverage (= 1) with the iNEXT R package (Chao and Jost 2012; Hsieh et al. 2016). We then plotted a non-metric multidimensional scaling ordination on square root transformed relative abundances to determine differences between the leaf and the rhizome strata and between the seasons. Such differences were then tested with PERMANOVA (Anderson 2001). For each station and season, we computed the number of specimens (N), the number of species (S), the Shannon diversity index (H’) and the Pielou’s evenness (J). For each species, we computed dominance (%D). The raw data arranged per replicate are available in ESM 1.
We attributed feeding guilds according to the following classification: carnivores (C) feeding on mobile organisms, such as molluscs or polychaetes; scavengers (SC) feeding on remains of dead organisms; deposit feeders (D) feeding on organic particles contained in the sediment; ectoparasites and specialized carnivores (E) feeding on much larger organisms on which they live during their life cycle; filter feeders (F) intercepting nutrient particles with their gills and/or mucous strings; macroalgae grazers (AG); seagrass grazers (SG) ingesting seagrass tissues; microalgal or periphyton grazers (MG) feeding on microalgae (e.g. diatoms); oophagus feeders (O), including species that feed on egg masses of other organisms; and symbiont-bearing species (SY) for those species in which symbiotic bacteria play an important role for obtaining a complementary energy source. This is the same classification of feeding modes and guilds used by Rueda et al. (2009) and Albano and Sabelli (2012) for comparative purposes. Trophic information for all species was obtained from the literature.
We determined the geographic range, with particular focus on endemic and non-indigenous species, from the literature. Molluscan species authorities are reported in the Tables 3 and 6 displaying abundance data; therefore, we do not mention them in the text. The systematic arrangement follows Sigwart et al. (2013) for the polyplacophorans, Bouchet et al. (2017) for the gastropods, Bouchet et al. (2010) for the bivalves, and Steiner and Kabat (2001) for the scaphopods. All data analyses and plotting were carried out in the R statistical environment (R Development Core Team 2019).
Results
Posidonia oceanica bed structure
The shoot density of Posidonia oceanica in the meadow decreases with depth. The highest shoot density of 740 ± 185 shoots m−2 was at 5-m depth. Density decreased to a minimum of 490 ± 52 shoots m−2 at 20-m depth (Table 1). Still, the status of the meadow increases with depth from good to high, because the decrease of shoot density with depth is naturally associated with decreased sun irradiation (Pergent et al. 1995). This trend may suggest that the shallowest sampling stations experience some degree of anthropic disturbance, in contrast to undisturbed deeper ones.
The molluscan taxocoenosis
We collected 9344 specimens, belonging to 109 species: 75 (68.8%) gastropods, 30 (27.5%) bivalves and 4 (3.7%) polyplacophorans. Species richness was 19 and 108 species in the leaf and rhizome stratum, respectively, with a very high sample coverage: 99.7% and 99.6%, respectively. Still, the extrapolated diversity at perfect coverage (= 1) was 23 (+21%) and 169 (+55%) species, respectively (Figure 3).

Sample coverage and estimated species richness at perfect coverage (= 1) of the leaf and rhizome strata of the Posidonia oceanica meadow in Plakias, south-western Crete. a Sample coverage reaches 99.7% and 99.6% in the leaf and rhizome stratum, respectively; b Notwithstanding the high coverage, the extrapolated diversity at perfect coverage is 23 (+21%) and 169 (+55%) species on the leaf and rhizome stratum, respectively
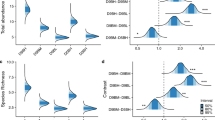
The non-metric multidimensional scaling plot shows that the molluscan assemblages of the leaf and rhizome strata were segregated into two distinct clouds of points (Figure 4). Indeed, these assemblages were significantly different (PERMANOVA, Fd = 29.83, R2=0.39, p=0.001) suggesting that they can be treated as different entities in the following paragraphs. Such differences were reflected in the most abundant species assemblage composition: in each stratum, five of the ten most abundant species were not equally abundant in the other stratum (Figure 5). Additionally, the rhizome stratum hosted most of the sampled individuals (7930 vs 1414 in the leaf stratum) and species (108 vs 19 in the leaf stratum). The differences between the seasons were very clear for the rhizomes and less neat for the leaves, but still statistically significant in both cases (PERMANOVA, leaf stratum: Fd = 5.18, R2 = 0.19, p = 0.001; rhizome stratum: Fd = 6.23, R2 = 0.22, p = 0.001).

Non-metric multidimensional scaling plot of molluscan assemblages in the leaf and rhizome strata of the Posidonia oceanica meadow in Plakias, south-western Crete. The molluscan assemblages differ between the strata and the seasons

Comparison of the ten most abundant species in the leaf (left, green) and the rhizome stratum (right, brown) of the Posidonia oceanica meadow in south-western Crete. The species in bold are those present in only one of the strata among the ten most abundant here illustrated
The assemblage contained 35 (0.3%) individuals of five species endemic to the eastern Mediterranean (Alvania bozcaadensis, Aegeofusinus rolani, Ocinebrina aegeensis, Chlathromangelia loiselieri and Parvicardium trapezium) and 98 (1.0%) individuals of four non-indigenous species (Viriola cf. bayani, Septifer cumingii, Isognomon aff. australica and Pinctada radiata), all of Indo-Pacific origin.
The molluscan assemblage on the leaves
In the leaf stratum, we collected 1414 individuals belonging to 19 species. Sampling saturation was excellent, exceeding a 98% coverage at all sampling events; still, the observed richness at 5-m depth in autumn and at 20-m depth in spring was largely underestimated (Table 2). Nearly all collected individuals are gastropods, except two individuals of bivalves, one belonging to the only non-indigenous species found among the leaves (Pinctada radiata in 20 m depth). The most abundant species were Jujubinus exasperatus, Bittium latreillii and Bittium reticulatum, represented by more than a hundred specimens each. The differences between the spring and autumn assemblages are mostly driven by different proportions of these species that are more dominant in spring. Rissoa violacea was the only species found exclusively in the leaf stratum. A detailed list of species per depth, season and their dominance, is given in Table 3. At the shallowest sites, Shannon index and Pielou’s evenness are markedly lower in autumn than in spring (Table 2). In terms of feeding guilds, the molluscan assemblage of the leaves is dominated by microalgal grazers with a minor component of seagrass-feeding herbivores, filter feeders and carnivores (Table 4).
The molluscan assemblage in the rhizomes
In the rhizome stratum, we collected 7930 individuals belonging to 108 species: 74 (68.5%) gastropods, 30 (27.8 %) bivalves and 4 (3.7%) polyplacophorans. Sampling saturation was excellent, exceeding a 98% coverage at all sampling events; still, the observed richness was largely underestimated, with an estimated richness ~16.7–192.6% larger than the observed one (Table 5). The most abundant species were Bittium latreillii, Alvania mamillata, Bittium reticulatum and Jujubinus exasperatus, with more than a thousand individuals each. The differences between the spring and autumn assemblages are mostly driven by different proportions of these species that are more dominant in spring. A detailed list, of species per depth, season and their dominance, is given in Table 6. Shannon diversity was much higher than in the leaf stratum whereas both Shannon diversity and Pielou’s evenness values were more stable between seasons (Table 5). The trophic structure of the molluscan assemblage in the rhizomes was much more diverse than in the leaves: eight guilds out of ten were represented (Table 7). Only scavengers and oophagus feeders were missing. More than 80% of the assemblage abundance is composed by microalgal grazers, with a non-marginal component of filter feeders.
Discussion
A diverse native molluscan assemblage, devoid of non-indigenous species
The molluscan assemblage of the Posidonia oceanica meadow in Plakias was abundant and diverse and hosted a negligible non-indigenous component. A direct comparison with the few previous works which inspected both the leaf and rhizome strata and published quantitative data (e.g. Templado 1984; Bonfitto et al. 1998; Solustri et al. 2002; Albano and Sabelli 2012) is hampered by the differences in sampling design and methods. Still, the 109 species here reported are only slightly less than the 139 in the methodologically most similar study on the meadow in Cabo de Palos (Murcia, south-eastern Spain) where sampling occurred across a broader depth range and on multiple years (Templado 1984). The Shannon diversity and Pielou’s evenness we recorded on both the leaf and rhizome strata are generally higher than the values recorded at Hvrgada Island in Croatia (Solustri et al. 2002), but lower than on the leaf stratum in the Posidonia oceanica meadow at Ischia Island, central Tyrrhenian Sea, at comparable depths (Idato et al. 1983).
The micrograzers Jujubinus, Tricolia, Bittium and Rissoidae are dominant in the leaf stratum like at Posidonia oceanica meadows in Spain (Templado 1984), in the Tyrrhenian Sea (Idato et al. 1983; Bonfitto et al. 1998; Albano and Sabelli 2012), in the Strait of Sicily (Accardo-Palumbo et al. 1992) and in the Adriatic Sea (Solustri et al. 2002). The rhizome stratum is still dominated by micrograzers, but has a more diverse trophic structure as it hosts also several filter-feeders (mostly bivalves), carnivores and ectoparasites; a typical pattern of Posidonia oceanica meadows (Templado 1984; Bonfitto et al. 1998; Solustri et al. 2002; Albano and Sabelli 2012). Despite some species move along the blade between day and night (so-called nychthemeral migrations (Russo et al. 1991)), all mentioned studies were conducted during the day making comparisons meaningful. Such comparisons suggest that the molluscan assemblage we inspected conforms to the expectations of healthy Posidonia oceanica meadows across the Mediterranean Sea.
In our study, the differences between seasons markedly varied with depth in the leaf stratum, where both Shannon diversity and evenness were greater in spring than autumn down to 10 m depth, but the opposite occurred at 15- and 20-m depth. These differences match those observed in Ischia Island in the Tyrrhenian Sea (Russo et al. 1991), where the structure of the leaf molluscan assemblage indeed varied with depth, but not with season. In the rhizome stratum, differences in Shannon diversity and evenness with season were minor and without a clear direction, comparable with the results from a very shallow water Posidonia oceanica meadow in the Alboran Sea (Urra et al. 2013). Also the proportions of the trophic guilds are similar between the two seasons in the Alboran Sea, whereas in our study microalgae herbivores and filter feeders showed in the rhizomes considerably greater abundance in spring than in autumn, possibly related to the greater availability of food in the water column during the spring phytoplankton bloom (Evans and Parslow 1985).
Of the four non-indigenous species found, only one, Pinctada radiata, is a long-term host of the Mediterranean Sea. It was recorded already in the second half of the nineteenth century, shortly after the opening of the Suez Canal (Vassel 1896). By now it has established populations all over the Mediterranean basin and reached as far as Spain (López Soriano and Quiñonero-Salgado 2019; Ballesteros et al. 2020; Png-Gonzalez et al. 2021) and the southern Adriatic Sea (Petović and Mačić 2018). Septifer cumingii was first recorded in the Mediterranean in 2001 (Albayrak and Çeviker 2001) and is now broadly distributed in the eastern Mediterranean (Katsanevakis et al. 2009; Albano et al. 2021a), whereas Viriola cf. bayani and Isognomon aff. australica are very recent introductions (Steger et al. 2018; Albano et al. 2021b), suggesting that not only the non-indigenous component is negligible, but that it is also of very recent acquisition.
Finally, yet importantly, in our samples there was a small but taxonomically and functionally diverse component of species endemic to the eastern Mediterranean. Endemic species at the scale of Mediterranean sub-basins were a small share of the diversity also in the Alboran Sea (Urra et al. 2013) and in the Tyrrhenian Sea (Albano and Sabelli 2012), showing on the one hand that Posidonia oceanica meadows host a quantitatively important component of the Mediterranean malacofauna, and on the other hand that this habitat is important for the survival of rare endemic species.
Posidonia oceanica meadows: a refugium against climate warming and deoxygenation?
This healthy assemblage was in sheer contrast with our expectation of an impoverished area heavily affected by biological invasions. Indeed, there is increasing evidence of collapsing native assemblages in other parts of the eastern Mediterranean (Crocetta et al. 2013, 2020; Rilov 2016; Albano et al. 2021a) and Crete is among the most invaded sectors of the Greek seas (Zenetos et al. 2011) and of the Mediterranean basin (Galil 2012; Galil et al. 2014, 2018).
The Mediterranean Sea has a north-west to south-east gradient of increasing sea surface temperature making the eastern Mediterranean its warmest sector (Pisano et al. 2020). On top of this climatological pattern, the eastern Mediterranean is experiencing the highest rates of warming in the basin (Ozer et al. 2017; Pisano et al. 2020). Mediterranean biodiversity is mostly of temperate origin, being the result of the extreme climatic fluctuations of the Pleistocene and especially of its latest glacial-interglacial cycle (Sabelli and Taviani 2014). Consequently, most native species are poorly adapted to the increasing tropical conditions experienced by the basin. Ongoing warming pushes species beyond their thermal tolerance limits, making climate change the main driver beyond the native invertebrate collapse and the changes in fish abundances over the last 20 years recently recorded on the Israeli shelf (Rilov 2016; Givan et al. 2018; Albano et al. 2021a; Steger et al. 2021b). Although the increasing sea surface temperature trend has been recorded for Crete too, the study site currently lies across the 20.5 °C annual isotherm (Pisano et al. 2020). This is almost 1 °C higher than in the 1982–1993 period, but still 2 °C lower than the Israeli shelf where the most dramatic effects have occurred already (Pisano et al. 2020).
Climate warming causes local extinction in multiple ways (Cahill et al. 2012), but in the marine realm, temperatures that exceed the species physiological tolerance are considered a major driver for ectotherms, with species ranges advancing and retracting closely matching suitable isotherms (Sunday et al. 2012). Warming waters increase metabolic oxygen demand and at the same time contain less oxygen because of reduced oxygen solubility and the increased oxygen consumption (Breitburg et al. 2018). Such conditions cause reduced growth and survival (Vaquer-Sunyer and Duarte 2011; Breitburg et al. 2018; Pauly 2021). However, photosynthetic activity causes an oxygen supersaturation that can buffer the effects of warming by significantly increasing the thermal tolerance of marine ectotherm vertebrates and invertebrates (Giomi et al. 2019). Large Posidonia oceanica meadows cause an oxygen supersaturation up to 161% (Hendriks et al. 2014) and thus have the full potential to exert the same buffering effect on thermal tolerance as experimented by Giomi et al. (2019).
Such persistent diverse assemblages in a warming climate may contribute to reduce invasion success (Stachowicz et al. 1999). In marine ecosystems, this resistance is stronger in the subtidal and on hard substrates where both competitive and consumptive biotic resistance occurs (Kimbro et al. 2013). Posidonia oceanica meadows host extensive surfaces acting as hard substrates (e.g. the leaves and the rhizomes) and occur from subtidal down to 40 m depth (Gobert et al. 2006), making them a paradigmatic habitat for biotic resistance.
Despite that our results shall be deemed preliminary, they may suggest that healthy seagrass meadows constitute a unique refugium for native biodiversity from warming and increasingly invaded waters on the shallow shelf. Future studies may compare meadows with other substrates and measure oxygen production to build up a more robust framework. Still, this perspective would be particularly valuable in the vulnerable eastern Mediterranean that is already experiencing the climate-driven collapse of native biodiversity and reinforces the need to fully protect this habitat that is retreating all over the Mediterranean Sea (Telesca et al. 2015).
Data availability
Quantitative data are included in ESM1 and will be uploaded in OBIS.
References
Accardo-Palumbo MT, Chemello R, Russo GF (1992) Malacofauna di strato foliare di una prateria di Posidonia oceanica dell’Isola di Lampedusa (Canale di Sicilia). Oebalia (Suppl XVII):315–319
Albano PG, Sabelli B (2012) The molluscan assemblages inhabiting the leaves and rhizomes of a deep water Posidonia oceanica settlement in the central Tyrrhenian Sea. Sci Mar 76:721–732. https://doi.org/10.3989/scimar.03396.02C
Albano PG, Steger J, Bakker PAJ, Bogi C, Bošnjak M, Guy-Haim T, Huseyinoglu MF, LaFollette PI, Lubinevsky H, Mulas M, Stockinger M, Azzarone M, Sabelli B (2021b) Numerous new records of tropical non-indigenous species in the Eastern Mediterranean highlight the challenges of their recognition and identification. ZooKeys 1010:1–95. https://doi.org/10.3897/zookeys.1010.58759
Albano PG, Steger J, Bošnjak M, Dunne B, Guifarro Z, Turapova E, Hua Q, Kaufman DS, Rilov G, Zuschin M (2021a) Native biodiversity collapse in the Eastern Mediterranean. Proc R Soc B Biol Sci 288:1–9. https://doi.org/10.1098/rspb.2020.2469
Albano PG, Stockinger M (2019) The rhizome layer of Posidonia oceanica: an important habitat for Mediterranean brachiopods. Mar Biodivers 49:2467–2472. https://doi.org/10.1007/s12526-019-00968-6
Albayrak S, Çeviker D (2001) Two new extra-Mediterranean molluscs from southeast Turkey: Siphonaria belcheri Hanley, 1858 [Gastropoda: Siphonariidae] and Septifer bilocularis (Linnaeus, 1758) [Bivalvia: Mytilidae]. Israel J Ecol Evol 47:297–298. https://doi.org/10.1560/50DQ-KE0G-N9RB-8K9A
Anderson MJ (2001) A new method for non-parametric multivariate analysis of variance. Austral Ecol 26:32–46. https://doi.org/10.1111/j.1442-9993.2001.01070.pp.x
Ballesteros E, Marsinyach E, Bagur M, Sales M, Movilla J, Bolado I, Cefalí ME (2020) The pearl oyster Pinctada imbricata radiata (Leach, 1814) (Bivalvia: Pteriidae) reaches Minorca, Balearic Islands. Bolletí de la Societat d’Història Natural de les Balears 63:97–108
Bedini R, Bedini M, Bonechi L, Piazzi L (2015) Patterns of spatial variability of mobile macro-invertebrate assemblages within a Posidonia oceanica meadow. J Nat Hist 49:2559–2581. https://doi.org/10.1080/00222933.2015.1021872
Belgacem W, Langar H, Hassine OKB (2011) Depth and temporal distribution of vagile fauna associated with Posidonia oceanica meadows in Cap Zebib, north-eastern Tunisian coastline. Afr J Ecol 49:459–470. https://doi.org/10.1111/j.1365-2028.2011.01278.x
Beqiraj S, Kashta L, Kuçi M, Kasemi D, Gace A (2008) Benthic macrofauna of Posidonia oceanica meadows in the Albanian coast. Natura Montenegrina 7:55–69
Bonfitto A, Fellagara I, Gillone G (1998) Sampling techniques and structure of the malacofauna associated to the rhizome zone in Posidonia oceanica (L.) Delile. Bollettino Malacologico 33:83–88
Bouchet P, Rocroi J-P, Bieler R, Carter JG, Coan EV (2010) Nomenclator of bivalve families with a classification of bivalve families. Malacologia 52:1–184
Bouchet P, Rocroi J-P, Hausdorf B, Kaim A, Kano Y, Nützel A, Parkhaev P, Schrödl M, Strong EE (2017) Revised classification, nomenclator and typification of gastropod and monoplacophoran families. Malacologia 61:1–526. https://doi.org/10.4002/040.061.0201
Boudouresque CF, Bernard G, Bonhomme P (2006) Préservation et conservation des herbiers à Posidonia oceanica. Ramoge pub, S.l
Breitburg D, Levin LA, Oschlies A, Grégoire M, Chavez FP, Conley DJ, Garçon V, Gilbert D, Gutiérrez D, Isensee K, Jacinto GS, Limburg KE, Montes I, Naqvi SWA, Pitcher GC, Rabalais NN, Roman MR, Rose KA, Seibel BA et al (2018) Declining oxygen in the global ocean and coastal waters. Science 359. https://doi.org/10.1126/science.aam7240
Cahill AE, Aiello-Lammens ME, Fisher-Reid MC, Hua X, Karanewsky CJ, Yeong Ryu H, Sbeglia GC, Spagnolo F, Waldron JB, Warsi O, Wiens JJ (2012) How does climate change cause extinction? Proc R Soc B Biol Sci 280:20121890–20121890. https://doi.org/10.1098/rspb.2012.1890
Chao A, Jost L (2012) Coverage-based rarefaction and extrapolation: standardizing samples by completeness rather than size. Ecology 93:2533–2547. https://doi.org/10.1890/11-1952.1
Crocetta F, Bitar G, Zibrowius H, Oliverio M (2013) Biogeographical homogeneity in the eastern Mediterranean Sea. II. Temporal variation in Lebanese bivalve biota. Aquat Biol 19:75–84. https://doi.org/10.3354/ab00521
Crocetta F, Bitar G, Zibrowius H, Oliverio M (2020) Increase in knowledge of the marine gastropod fauna of Lebanon since the 19th century. Bull Mar Sci 96:22. https://doi.org/10.5343/bms.2019.0012
Dimitriadis C, Galanidi M, Zenetos A, Corsini-Foka M, Giovos I, Karachle PK, Konstantinidoy IF, Kytinou E, Issaris Y, Azzurro E, Castriota L, Falautano M, Kalimeris A, Katsanevakis S (2020) Updating the occurrences of Pterois miles in the Mediterranean Sea, with considerations on thermal boundaries and future range expansion. Mediterr Marine Sci 21:62–69. https://doi.org/10.12681/mms.21845
Evans GT, Parslow JS (1985) A model of annual plankton cycles. Biol Oceanogr 3:327–347
Galil BS (2009) Taking stock: inventory of alien species in the Mediterranean sea. Biol Invasions 11:359–372. https://doi.org/10.1007/s10530-008-9253-y
Galil BS (2012) Truth and consequences: the bioinvasion of the Mediterranean Sea. Integr Zool 7:299–311. https://doi.org/10.1111/j.1749-4877.2012.00307.x
Galil BS, Marchini A, Occhipinti-Ambrogi A (2018) East is east and West is west? Management of marine bioinvasions in the Mediterranean Sea. Estuar Coast Shelf Sci 201:7–16. https://doi.org/10.1016/j.ecss.2015.12.021
Galil BS, Marchini A, Occhipinti-Ambrogi A, Minchin D, Narščius A, Ojaveer H, Olenin S (2014) International arrivals: widespread bioinvasions in European Seas. Ethol Ecol Evol 26:152–171. https://doi.org/10.1080/03949370.2014.897651
Gambi MC, Giangrande A, Martinelli M, Chessa LA (1995) Polychaetes of a Posidonia oceanica bed off Sardinia (Italy): spatio-temporal distribution and feeding guild analysis. Sci Mar 59:129–141
Gambi MC, Lorenti M, Russo GF, Scipione MB, Zupo V (1992) Depth and seasonal distribution of some groups of the vagile fauna of the posidonia oceanica leaf stratum: structural and trophic analyses. Mar Ecol 13:17–39. https://doi.org/10.1111/j.1439-0485.1992.tb00337.x
García-Raso JE (1990) Study of a Crustacea Decapoda taxocoenosis of Posidonia oceanica beds from the southeast of Spain. Mar Ecol 11:309–326. https://doi.org/10.1111/j.1439-0485.1990.tb00386.x
Giomi F, Barausse A, Duarte CM, Booth J, Agusti S, Saderne V, Anton A, Daffonchio D, Fusi M (2019) Oxygen supersaturation protects coastal marine fauna from ocean warming. Sci Adv 5:eaax1814. https://doi.org/10.1126/sciadv.aax1814
Givan O, Edelist D, Sonin O, Belmaker J (2018) Thermal affinity as the dominant factor changing Mediterranean fish abundances. Glob Chang Biol 24:e80–e89. https://doi.org/10.1111/gcb.13835
Gobert S, Cambridge ML, Velimirov B, Pergent G, Lepoint G, Bouquegneau J-M, Dauby P, Pergent-Martini C, Walker DI (2006) Biology of Posidonia. In: Seagrasses: biology, ecology and conservation. Springer-Verlag, Berlin/Heidelberg, pp 387–408
Hendriks IE, Olsen YS, Ramajo L, Basso L, Steckbauer A, Moore TS, Howard J, Duarte CM (2014) Photosynthetic activity buffers ocean acidification in seagrass meadows. Biogeosciences 11:333–346. https://doi.org/10.5194/bg-11-333-2014
Hsieh TC, Ma KH, Chao A (2016) iNEXT: an R package for rarefaction and extrapolation of species diversity (Hill numbers). Methods Ecol Evol 7:1451–1456. https://doi.org/10.1111/2041-210X.12613
Idato E, Fresi E, Russo GF (1983) Zonazione verticale della fauna vagile di strato foliare in una prateria di Posidonia oceanica Delile: I - Molluschi. Bollettino Malacologico 19:109–120
Katsanevakis S, Tsiamis K, Ioannou G, Michailidis N, Zenetos A (2009) Inventory of alien marine species of Cyprus (2009). Mediterr Marine Sci 10:109–134. https://doi.org/10.12681/mms.113
Kimbro DL, Cheng BS, Grosholz ED (2013) Biotic resistance in marine environments. Ecol Lett 16:821–833. https://doi.org/10.1111/ele.12106
Ledoyer M (1962) Etude de la faune vagile des herbiers superficiels de zosteracees et de quelques biotopes d’algues littorales. Recueil des travaux de la Station Marine d’Endoume 25:117–235
López Soriano J, Quiñonero-Salgado S (2019) Primera cita de Pinctada radiata (Leach, 1814) (Bivalvia: Pteriidae) para la Península Ibérica. Elona 1:52–56
Nunes AL, Katsanevakis S, Zenetos A, Cardoso AC (2014) Gateways to alien invasions in the European seas. Aquat Invasions 9:133–144. https://doi.org/10.3391/ai.2014.9.2.02
Ozer T, Gertman I, Kress N, Silverman J, Herut B (2017) Interannual thermohaline (1979–2014) and nutrient (2002–2014) dynamics in the Levantine surface and intermediate water masses, SE Mediterranean Sea. Glob Planet Chang 151:60–67. https://doi.org/10.1016/j.gloplacha.2016.04.001
Panayotidis P, Boudouresque CF, Marcot-Coqueugniot J (1981) Microstructure de l’herbier de Posidonia oceanica (Linnaeus) Delile. Microstructure of Posidonia oceanica (Linnaeus) beds. Bot Mar 24:115–124. https://doi.org/10.1515/botm.1981.24.3.115
Pauly D (2021) The gill-oxygen limitation theory (GOLT) and its critics. Sci Adv 7:eabc6050. https://doi.org/10.1126/sciadv.abc6050
Peleg O, Guy-Haim T, Yeruham E, Silverman J, Rilov G (2019) Tropicalisation may invert trophic state and carbon budget of shallow temperate rocky reefs. J Ecol 108:844–854. https://doi.org/10.1111/1365-2745.13329
Pergent G, Pergent-Martini C, Boudouresque CF (1995) Utilisation de l’herbier a Posidonia oceanica comme indicateur biologique de la qualité du milieu littoral en Méditerranée: état des connaissances. Mésogée 54:3–27
Petović S, Mačić V (2018) New data on Pinctada radiata (Leach, 1814) (Bivalvia: Pteriidae) in the Adriatic Sea. Acta Adriat (Online) 58:357–361. https://doi.org/10.32582/aa.58.2.14
Pisano A, Marullo S, Artale V, Falcini F, Yang C, Leonelli FE, Santoleri R, Buongiorno Nardelli B (2020) New evidence of Mediterranean climate change and variability from sea surface temperature observations. Remote Sens 12:132. https://doi.org/10.3390/rs12010132
Png-Gonzalez L, Aguilo-Arce J, Vázquez-Luis M, Carbonell A (2021) New occurrence of Pinctada imbricata radiata (Leach, 1814) in the Balearic Archipelago (NW Mediterranean Sea). BioInvasions Rec 10(4):853–858
R Development Core Team (2019) R: a language and environment for statistical computing. Vienna, Austria: R Foundation for Statistical Computing. http://www.r-project.org
Rilov G (2016) Multi-species collapses at the warm edge of a warming sea. Sci Rep 6:36897. https://doi.org/10.1038/srep36897
Rilov G, Peleg O, Yeruham E, Garval T, Vichik A, Raveh O (2018) Alien turf: overfishing, overgrazing and invader domination in south-eastern Levant reef ecosystems. Aquat Conserv Mar Freshwat Ecosyst 28:351–369. https://doi.org/10.1002/aqc.2862
Rueda JL, Gofas S, Urra J, Salas C (2009) A highly diverse molluscan assemblage associated with eelgrass beds (Zostera marina L.) in the Alboran Sea: micro-habitat preference, feeding guilds and biogeographical distribution. Sci Mar 73:679–700. https://doi.org/10.3989/scimar.2009.73n4679
Russo GF, Fresi E, Vinci D (1985) The hand-towed net method for direct sampling in Posidonia oceanica beds. Rapports et Proces-Verbaux des Réunions Conseil Internationale pour l’Exploration Scientifique de la Mer Méditerranée 29:175–177
Russo GF, Vinci D, Scardi M (1991) Mollusc syntaxon of foliar stratum along a depth gradient in a Posidonia oceanica bed: 3. A year’s cycle at Ischia Island. Posidonia Newsl 4:15–25
Sabelli B, Taviani M (2014) The making of the Mediterranean molluscan biodiversity. In: Goffredo S, Dubinsky Z (eds) The Mediterranean Sea: its history and present challenges. Springer Netherlands, Dordrecht, pp 285–306
Sala E, Kizilkaya Z, Yildirim D, Ballesteros E (2011) Alien marine fishes deplete algal biomass in the Eastern Mediterranean. PLoS One 6:e17356. https://doi.org/10.1371/journal.pone.0017356
Savva I, Chartosia N, Antoniou C, Kleitou P, Georgiou A, Stern N, Hadjioannou L, Jimenez C, Andreou V, Hall-Spencer JM, Kletou D (2020) They are here to stay: the biology and ecology of lionfish (Pterois miles) in the Mediterranean Sea. J Fish Biol 97:148–162. https://doi.org/10.1111/jfb.14340
Sigwart JD, Stoeger I, Knebelsberger T, Schwabe E (2013) Chiton phylogeny (Mollusca : Polyplacophora) and the placement of the enigmatic species Choriplax grayi (H. Adams & Angas). Invert Syst 27:603. https://doi.org/10.1071/IS13013
Solustri C, Morello E, Sabelli B (2002) Primi dati sulla malacofauna associata ad una prateria di Posidonia oceanica (L.) Delile in Adriatico Orientale (Croazia). Biologia Marina Mediterranea 9:231
Stachowicz JJ, Whitlatch RB, Osman RW (1999) Species diversity and invasion resistance in a marine ecosystem. Science 286:1577–1579. https://doi.org/10.1126/science.286.5444.1577
Steger J, Bošnjak M, Belmaker J, Galil BS, Zuschin M, Albano PG (2021b) Non-indigenous molluscs in the Eastern Mediterranean have distinct traits and cannot replace historic ecosystem functioning. Glob Ecol Biogeogr 31:89–102. https://doi.org/10.1111/geb.13415
Steger J, Dunne B, Zuschin M, Albano PG (2021a) Bad neighbors? Niche overlap and asymmetric competition between native and Lessepsian limpets in the Eastern Mediterranean rocky intertidal. Mar Pollut Bull 171:112703. https://doi.org/10.1016/j.marpolbul.2021.112703
Steger J, Stockinger M, Ivkić A, Galil B, Albano PG (2018) New records of non-indigenous molluscs from the eastern Mediterranean Sea. BioInvasions Records 7:245–257. https://doi.org/10.3391/bir.2018.7.3.05
Steiner G, Kabat AR (2001) Catalogue of supraspecific taxa of Scaphopoda (Mollusca). Zoosystema 23:433–460
Sunday JM, Bates AE, Dulvy NK (2012) Thermal tolerance and the global redistribution of animals. Nat Clim Chang 2:686–690. https://doi.org/10.1038/nclimate1539
Telesca L, Belluscio A, Criscoli A, Ardizzone G, Apostolaki ET, Fraschetti S, Gristina M, Knittweis L, Martin CS, Pergent G, Alagna A, Badalamenti F, Garofalo G, Gerakaris V, Pace ML, Pergent-Martini C, Salomidi M (2015) Seagrass meadows (Posidonia oceanica) distribution and trajectories of change. Sci Rep 5:12505. https://doi.org/10.1038/srep12505
Templado J (1984) Moluscos de las praderas de Posidonia oceanica en las costas del cabo de Palos (Murcia). Investig Pesq 48:509–526
Templado J, Paulay G, Gittenberger A, Meyer C (2010) Sampling the marine realm. In: Eymann J, Degreef J, Häuser C, Monje JC, Samyn Y, VandenSpiegel D (eds) Manual on field recording techniques and protocols for all taxa biodiversity inventories, ABC Taxa, pp 273–302
UNEP/MAP-RAC/SPA (2015) Guidelines for the standardization of mapping and monitoring methods of marine Magnoliophyta in the Mediterranean. UNEP/MAP-RAC/SPA, Tunis
Urra J, Mateo Ramírez Á, Marina P, Salas C, Gofas S, Rueda JL (2013) Highly diverse molluscan assemblages of Posidonia oceanica meadows in northwestern Alboran Sea (W Mediterranean): seasonal dynamics and environmental drivers. Estuar Coast Shelf Sci 117:136–147. https://doi.org/10.1016/j.ecss.2012.11.005
Vaquer-Sunyer R, Duarte CM (2011) Temperature effects on oxygen thresholds for hypoxia in marine benthic organisms. Glob Chang Biol 17:1788–1797. https://doi.org/10.1111/j.1365-2486.2010.02343.x
Vassel E (1896) Sur la Pintadine du Golfe de Gabes. Compte Rendu de la Association Française pour l’Avancement des Sciences 25:458–466
Yeruham E, Rilov G, Shpigel M, Abelson A (2015) Collapse of the echinoid Paracentrotus lividus populations in the Eastern Mediterranean—result of climate change? Sci Rep 5:13479. https://doi.org/10.1038/srep13479
Yeruham E, Shpigel M, Abelson A, Rilov G (2019) Ocean warming and tropical invaders erode the performance of a key herbivore. Ecology 101:e02925. https://doi.org/10.1002/ecy.2925
Zenetos A (2015) ELNAIS: a collaborative network on aquatic alien species in Hellas (Greece). MBI 6:185–196. https://doi.org/10.3391/mbi.2015.6.2.09
Zenetos A, Katsanevakis S, Poursanidis D, Crocetta F, Damalas D, Apostolopoulos G, Gravili C, Vardala-Theodorou E, Malaquias M (2011) Marine alien species in Greek Seas: additions and amendments by 2010. Mediterr Mar Sci 12:95–120. https://doi.org/10.12681/mms.55
Zenetos A, Vassilopoulou V, Salomidi M, Poursanidis D (2008) Additions to the marine alien fauna of Greek waters (2007 update). Mar Biodivers Rec 1:E91. https://doi.org/10.1017/S1755267207009281
Zenetos Α, Gofas S, Morri C, Rosso A, Violanti D, JEG R, Cinar ME, Almogi-Labin A, Ates AS, Azzurro E, Ballesteros E, Bianchi CN, Bilecenoglu M, Gambi MC, Giangrande A, Gravili C, Hyams-Kaphzan O, Karachle PK, Katsanevakis S et al (2012) Alien species in the Mediterranean Sea by 2012. A contribution to the application of European Union’s Marine Strategy Framework Directive (MSFD). Part 2. Introduction trends and pathways. Mediterr Mar Sci 13:328–352. https://doi.org/10.12681/mms.327
Acknowledgements
This research was conducted in the framework of the project “Historical ecology of Lessepsian migration” funded by the Austrian Science Fund (FWF) P28983-B29 (PI: PGA). Martin Zuschin, University of Vienna, supported us throughout the whole project. The Mare Mundi and Dive2gether Marine Field Station on Crete hosted the team during fieldwork. Stefan Duijvestijn, Michael Holzknecht, Nadja Loferer and Denny Morchner supported sampling and lab work. We thank two anonymous reviewers for their useful comments to a first draft of the manuscript.
Funding
Open access funding provided by Austrian Science Fund (FWF). Fieldwork was supported by the KWA–Kurzfristige Auslandstipendien of the University of Vienna.
Author information
Authors and Affiliations
Contributions
MH and PGA conceived the question and designed the study. MH conducted fieldwork and labwork under PGA’s supervision. MH and PGA analysed the data and wrote the manuscript.
Corresponding author
Ethics declarations
Conflict of interest
The authors declare no competing interests.
Ethics approval
All applicable international, national and/or institutional guidelines for sampling were followed.
Sampling and field studies
The fieldwork has been conducted in compliance with all applicable regulations.
Additional information
Communicated by C. Chen
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
ESM 1
Quantitative data (XLSX 38 kb)
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Holzknecht, M., Albano, P.G. The molluscan assemblage of a pristine Posidonia oceanica meadow in the eastern Mediterranean. Mar. Biodivers. 52, 59 (2022). https://doi.org/10.1007/s12526-022-01292-2
Received:
Revised:
Accepted:
Published:
DOI: https://doi.org/10.1007/s12526-022-01292-2