
Abstract
Coastal lagoons and other estuarine habitats are increasingly exposed to the negative effects of growing human populations with associated intensifications in nutrient loading, harmful algal blooms, pollution, and habitat degradation. We examined population dynamics of Gulf pipefish Syngnathus scovelli in estuarine waters of the Indian River Lagoon, on the Atlantic coast of Florida. Substantial declines in abundance of this sentinel species were concurrent with significant losses of seagrass habitat associated with ongoing harmful algal blooms, and other perturbations during the study period spanning from 1998 to 2018. Moderate declines in S. scovelli were observed with early downward trends in seagrasses ahead of the onset of the precipitous seagrass reductions observed. The massive decline of seagrass habitats in the Indian River Lagoon in recent years had negative influences and was directly linked to population declines we observed in the S. scovelli population. Lack of seagrass habitat essential to this and related syngnathid species may reduce optimum seagrass-associated prey, increase predation by lack of appropriate cover, and increase energetic costs which may be realized through reduced growth rates and potential reproductive impairment. Identification and monitoring of population trends of S. scovelli and related sentinel fish species allow for early implementation of management actions that reduce the impact of anthropogenic pressures on the services that estuarine systems provide to the fishes and fisheries they support.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Estuaries provide critical habitats for many fish species and are among the most productive yet most vulnerable coastal ecosystems (Kennish 2002). Habitat loss and anthropogenic stressors negatively influence estuaries globally (Lotze et al. 2006). Shallow coastal lagoons and other estuarine habitats are increasingly susceptible to the impacts of changing human populations and urbanization via increased nutrient loading, contaminants, and related anthropogenic influences (Kennish 2002; McGlathery et al. 2007; Carstensen et al. 2011). The Indian River Lagoon system (IRL) on Florida’s Atlantic coast is no exception (Adams et al. 2019). Human population growth along the IRL has put a substantial amount of pressure on the lagoon’s ecological dynamics. Estimates of the human population of the five coastal counties of the IRL region increased from 1,351,946 in 2000 to 1,724,438 in 2018, and recent projections estimate that it will increase to approximately 2,024,353 by 2030 and 3,667,815 by 2045 (FBEBR 2011, 2021).
Coastal systems worldwide are experiencing increased frequency and duration of harmful algal blooms due to eutrophication, pollution, and climatic and nonclimatic phenomena which together may have a synergistic effect (Glibert 2020; Gobler 2020; Griffith and Gobler 2020). Eutrophication and long-duration harmful algal blooms have been prominent in the IRL during recent years (Lapointe et al. 2015; Phlips et al. 2015). An extensive Pyrodinium bahamense bloom in the IRL coincided with pufferfish toxicity events in 2002 (Phlips et al. 2004; Landsberg et al. 2006). Beginning in 2011, a widespread and long-duration phytoplankton bloom, termed the superbloom, occurred within the IRL system. This superbloom was intensively monitored by the Florida Fish and Wildlife Conservation Commission’s Fish and Wildlife Research Institute’s (FWC-FWRI) Harmful Algal Bloom group and other agencies. The following year, and again in 2016, widely distributed Aureoumbra lagunensis blooms, also known as brown tides, impacted IRL waters (Phlips et al. 2015; Lapointe et al. 2020; Lopez et al. 2021; FWC–FWRI unpublished data). Sporadic and variable-scale blooms of the dinoflagellate P. bahamense and an array of other nanoplankton blooms have been recorded in the northern IRL since and are monitored by FWC–FWRI during weekly standardized water-sampling transects, as well as by ongoing efforts of other state agency and university groups (Lopez et al. 2021).
FWC–FWRI established a Fish Kill Hotline as a means of allowing other governmental entities and the public to report fish kills and other observed fish anomalies (Abbott et al. 2009). More than 220 fish kills were reported in the Indian River Lagoon system from 2008 through 2019 (FWC–FWRI Fish Kill Hotline database), although this number likely underrepresents the full extent of fish kills, as many go unreported, or the observations lack pertinent data for evaluation (Phelps et al. 2019). Hypoxic events related to eutrophication and subsequent harmful algal blooms were responsible for some of these fish kills, but, for many others, the causes remain unexplained. Concurrently, large-scale seagrass die-offs have been observed in the northern IRL, associated with decreased light penetration resulting from prolonged harmful algal blooms (Morris et al. 2018; Lapointe et al. 2020). Within the time frame of our study, declines in mean percent cover of seagrasses in the northern IRL were observed at the onset in 1998 (Morris et al. 2021). Seagrass declines greater than 95% have been recorded on several survey transects in the IRL since 2009 (Morris et al. 2018). In the northern Banana River Lagoon, an IRL basin that has experienced severe seagrass die-offs, long-term transect results reveal greater than 99% loss of seagrasses and an overall decline in seagrass species diversity from 2009 through early 2020 (D.M. Scheidt, Kennedy Space Center Ecological Program, personal communication, January 2020).
Declines in pipefishes and seahorses related to habitat losses, overexploitation, pollution, and climate change have been a continuing global concern (Pollom et al. 2021). Our initial observations, based on data from the FWC-FWRI long-term Fisheries-Independent Monitoring program, suggested that abundance levels of some fish species, including syngnathids, may be reduced along with changes in community structure and species diversity in the IRL. These changes may be associated with the significant losses of seagrass habitat, ongoing harmful algal blooms, and other perturbations in the system (e.g., reduced water quality, hypoxia, sporadic raw sewage spills, and other pollution) (FWC–FWRI unpublished data). Seahorses and pipefishes have been used as flagship species in many areas worldwide for strategic conservation and management endpoints (Shokri et al. 2009; Vincent et al. 2011) and have the potential to serve as sentinel species to detect sometimes cryptic ecosystem changes. Pipefishes and seahorses are also commercially harvested in Florida waters as part of the Marine Life category; however, current landings of pipefishes are not substantial (FWC-FWRI Commercial Fishery Landings Data through Batch 1515, extracted March 25, 2022. Marine Fisheries Information System. St. Petersburg, FL).
To investigate further, we analyzed population trends of the Gulf pipefish Syngnathus scovelli (Evermann & Kendall 1896), a common species in the southeastern United States that is resident in the lagoon with a limited home range and is sensitive to habitat perturbations, with well-understood life history parameters and habitat preferences (Joseph 1957; Brown 1972; IUCN 2013). Effective sentinel species respond rapidly to ecosystem variation, are common species or are effectively monitored within the study system, are typically resident to the study area with known life history parameters, and have documented sensitivity to an array of environmental stressors (Bortone et al. 2006; Adams et al. 2010).
Collectively, ongoing ecosystem perturbations in the Indian River Lagoon may influence pipefish populations as these species are particularly sensitive to habitat loss, habitat degradation, and reduced water quality (Vincent et al. 2011). Syngnathus scovelli has not been assessed by the International Union for Conservation of Nature and Natural Resources (IUCN) Red List of Threatened Species since 2012, and at that time was classified as “Least Concern” with limited or uncertain population data suggesting a relatively stable or slowly declining overall population trend (IUCN 2013). Considering all the major perturbations in the IRL over the past 20 years, we sought to answer three questions related to S. scovelli in the IRL: (1) How has the population abundance of S. scovelli in the IRL changed over time? (2) How do changes in the population relate to variations in habitat over time? (3) Is the distribution of the IRL population of S. scovelli stable over time?
Materials and methods
Study area
The sampling area identified as the northern Indian River Lagoon (IRL) system is a narrow estuary located along the central Atlantic coast of Florida and extends from the northern terminus of the Indian River Lagoon proper south to Vero Beach. The northern IRL is connected to the Atlantic Ocean by one permanent inlet (Sebastian Inlet) and one intermittently open conduit via the Canaveral Locks at Port Canaveral that links the Banana River Lagoon to the Atlantic just south of Cape Canaveral. The limited natural freshwater inflow comes primarily from the St. Sebastian River and from numerous creeks located mainly along the lagoon’s western shoreline (Bergman and Donnangelo, 1998). Shoreline vegetation consists largely of fringing mangrove (e.g., red mangrove Rhizophora mangle), invasive Brazilian pepper Schinus terebinthifolius, and marsh grasses (e.g., cordgrasses, Spartina spp.). Bottom substratum types are typically characterized as sand or mud mixed with shell hash and occasional oyster reefs. Seagrasses, primarily shoal grass Halodule wrightii and manatee grass Syringodium filiforme have historically been the dominant rooted vegetative cover in the northern IRL, although five other seagrass species also occur in the system, though some have diminished over time (Steward et al. 2006; Morris et al. 2021).
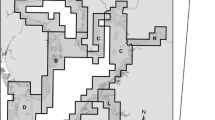
The FWC-FWRI Fisheries-Independent Monitoring program has conducted intensive sampling of fish species in the northern IRL since 1990. To standardize gear types and have consistent geographic coverage in this study, we focused our analyses on Fisheries-Independent Monitoring program data collected from 1998 through 2018. The area sampled for this study was divided into 3 geographically defined estuarine zones based on distinct basin areas—Banana River Lagoon (BRL), Indian River Lagoon north (IRLN), and Indian River Lagoon south (IRLS) (Fig. 1)—that are representative of this segment of the IRL system.

Maps of study area with distribution and relative abundance of Syngnathus scovelli in the Indian River Lagoon during 1998–2018 presented in 3-yr segments (1998–2000; 2001–2003; 2004–2006; 2007–2009; 2010–2012; 2013–2015; 2016–2018). Bubble sizes indicate magnitude of abundance classes: (·, 0 individuals), low ( , 1–10 individuals), medium (
, 1–10 individuals), medium ( , 11–50 individuals), and high (
, 11–50 individuals), and high ( , >50 individuals) catches of pipefish during each time segment. PC, Port Canaveral.
, >50 individuals) catches of pipefish during each time segment. PC, Port Canaveral.  Green shading represents areal extent of seagrass during each segment. Seagrass layers for Brevard and Indian River counties obtained from St. Johns River Water Management District Geospatial Open Data website https://data-floridaswater.opendata.arcgis.com/
Green shading represents areal extent of seagrass during each segment. Seagrass layers for Brevard and Indian River counties obtained from St. Johns River Water Management District Geospatial Open Data website https://data-floridaswater.opendata.arcgis.com/
Experimental design
The FWC-FWRI Fisheries-Independent Monitoring program uses a stratified-random sampling design. The IRL was divided into sampling zones based on geographic and logistical criteria, and each zone was subdivided into 1 minute of latitude × 1 minute of longitude grids. Each grid was further subdivided into 100 microgrids of equal area that were randomly selected for sampling. Sampling grids were stratified by habitat and depth. A single 21.3-m seine sample was collected at each randomly selected site. Monthly sampling for this study was conducted in BRL, IRLN, and IRLS using standardized 21.3-m center-bag seines. The number of monthly samples collected in each zone was proportional to the number of grids in the zone that could be sampled with that gear type.
A 21.3-m center-bag seine (1.8 m deep, with 3.2-mm knotless nylon mesh) was used to sample fishes in water ≤1.5 m deep and selected sites were stratified by the presence or absence of submersed aquatic vegetation in or the presence of a shoreline directly adjacent to the area sampled. Each seine haul was standardized with the aid of guidelines attached to each seine pole and secured to the substratum at the starting point (9.1-m length) with a separate guide line (15.5-m length) connected between seine poles. This technique allowed for a uniform sample coverage area (140 m2) during each haul. All sampling was conducted during daytime hours (1 h after sunrise to 1 h before sunset).
At each site, water temperature (°C), salinity (ppt), pH, and dissolved oxygen (mg/l) were recorded with a multiprobe water-quality instrument (Hydrolab Surveyor or YSI 600 series). Comprehensive fish habitat information, including percent cover of submersed aquatic vegetation (hereafter refers to seagrasses and attached algae Caulerpa spp.), was also assessed visually at each sampling site according to standardized FWC-FWRI Fisheries-Independent Monitoring program procedures (FWC-FWRI 2016). Sample processing was standardized. All fishes captured were identified to the lowest practical taxonomic level, counted, and a random sample of at least 10 individuals measured (standard length to the nearest mm).
Statistical analysis
Annual indices of relative abundance (IOA) were computed using generalized linear models. To focus on the adult population and reduce confounding effects of young-of-the-year recruitment, we analyzed S. scovelli ≥60 mm standard length. This size conservatively corresponds to known adult sizes from throughout the range of this species (Joseph 1957; Brown 1972; Jones and Avise 1997; Gasparini and Teixeira 1999) as well as our observations in the IRL of sexually mature males with a developed ventral brood pouch and of mature females with complete silver trunk bars (Brown 1972; Jones and Avise 1997). We analyzed adult length-frequency distributions over the entire study period to determine changes in or stability of fish length over time. Mean fish per haul for positive S. scovelli collections in habitats with differing submersed aquatic vegetation coverage (100% and 0% coverages; moderate to high coverages [≥25%] and sparse coverage [1–24%]) were analyzed using a Mann-Whitney rank-sum test.
The FWC-FWRI Fisheries-Independent Monitoring program’s stratified-random sampling design generates count data, the distribution of which is bounded by zero. Often, the frequency distribution of these counts is highly nonnormal. Therefore, a negative binomial distribution was used to create IOAs. Location, time, and environmental variables were treated in the analyses either as classification variables (IRL zone, year, month, shore type, substratum type, and submersed aquatic vegetation) or covariates (water temperature, salinity, and depth). The GLIMMIX procedure (SAS Institute Inc. 2006) was used to complete all analyses. To normalize the data, we natural log–transformed [ln(X+1)] water temperature, salinity, and depth before analysis. Akaike’s information criterion (AIC) was used to determine whether variables contributed a substantial amount of explained variability to the model. With the exception of sampling year, all nonsignificant (P > 0.05) variables that did not improve the fit of the model based on AIC were eliminated, and the analysis was repeated sequentially until only significant variables remained or until AIC values increased. The final model was completed using classification variables and covariates with the lowest AIC.
Relative abundance was calculated for each year as the median number of fish per set. Median values were determined from least-squares-adjusted means by multiplying the standard error by a random normal deviate (μ = 0, σ = 1) and adding it to the least-squares mean. These data were then back-transformed (ex). The process was repeated 500 times for each year to create a sampling distribution of back-transformed values, and summary statistics (25 and 75 percentiles) were then calculated and plotted to allow visualization of annual trends in IOAs (Sokal and Rohlf 1981).
Count data used in the IOA analysis were plotted to allow examination of changes in spatial distribution and abundance within the IRL study area through time, and with changes in seagrass coverage (seagrass mapping layers for the IRL obtained from St. Johns River Water Management District Geospatial Open Data website https://data-floridaswater.opendata.arcgis.com/). Data were partitioned into 3-year segments and plotted on a graduated scale with four abundance classes, representing zero, low (1–10 individuals), medium (11–50 individuals), and high (>50 individuals) catches of pipefish along with the latest year seagrass coverage mapping layer during each time segment.
Results
A total of 7059 standardized 21.3-m seine hauls were completed in our study area within the NIRL, SIRL, and BRL from 1998 to 2018. A total of 10,291 S. scovelli (≥60 mm SL) was collected across all samples. The largest catch of this species in a single seine haul (n = 96) was made in 1998 in full-coverage seagrass habitat (80% widgeon grass Ruppia maritima, 20% shoal grass, Halodule wrightii). Our extensive sampling in the IRL indicates that R. maritima has been rare or nonexistent within the study area since 2011 and that seagrass species richness has generally decreased during that time (FWC–FWRI data; Morris et al. 2021). The areas that have had the greatest seagrass loss in the NIRL either have experienced an increase in Caulerpa spp. or have remained bare substratum (FWC–FWRI data; L. Hall, St. Johns River Water Management District, personal communication, October 2021). Relative abundance of S. scovelli varied by year and generally displayed a declining trend beginning in 2002, concurrent with large-scale declines in seagrass abundance in the estuary (Figure 2). The longest period of consistent year-to-year decline lasted from 2008 through 2013, followed by minor increases in 2016 and 2018 and the lowest 21-year relative abundance observed in 2017. General linear model analyses identified important environmental factors that were significantly related to the relative abundance of S. scovelli, including submersed aquatic vegetation, substratum type, temperature, and depth; submersed aquatic vegetation was the most important (Table 1). During the study period, mean annual water temperature in the IRL increased approximately 0.9°C. Mean annual dissolved oxygen increased approximately 0.3 mg/L, but was frequently reduced in specific areas at times during algal blooms, and salinity increased 7.6 ppt. Mean overall water temperature in the IRL was 24.9°C (± 4.9 SD), mean dissolved oxygen was 7.2 mg/L (± 1.8 SD), and mean salinity was 26.1 (± 6.4 SD).

Annual least-squares means (± SE) of Syngnathus scovelli relative abundance (fish per haul) based on seine monitoring surveys conducted in the Indian River Lagoon, 1998–2018 (black circles). Mean percent cover of seagrass during standardized summer transect sampling in the northern Indian River Lagoon by the St. Johns River Water Management District (gray triangles)
For positive S. scovelli collections (where 1 or more pipefish were collected), significantly more fish per haul were collected in habitats where submersed aquatic vegetation was 100% coverage than in habitats with 0% coverage (P < 0.001, Mann-Whitney rank-sum test). Mean fish per haul for positive S. scovelli collections was 6.39 (± 0.226 standard error) in habitats where submersed aquatic vegetation was 100% coverage and 3.0 fish per haul (± 0.459 standard error) in habitats with 0% coverage. Significantly more fish per haul were collected in habitats where submersed aquatic vegetation was classified as moderate to high coverage (≥25%) than in habitats with sparse coverage (1–24%, P < 0.001, Mann-Whitney rank-sum test). Mean S. scovelli per haul was 4.60 (± 0.163 standard error) in habitats where submersed aquatic vegetation was ≥25% coverage and 2.99 fish per haul (± 0.235 standard error) in habitats with 1–24% coverage. The majority of S. scovelli were collected in depths below 1 m, with 3984 fish caught in the shallowest depth range of 0.2–0.5 m and 5454 fish in the 0.6–0.9 depth range. A total of 850 S. scovelli were collected in the 1.0–1.3 m depth range and only 3 fish collected in depths ≥ 1.4 m. This species was slightly more seasonally abundant during the Summer (June–August, n = 3149 fish) than during Spring (March–May, n = 2724 fish), Fall (September–October, n = 2101 fish), and Winter (December–February, n = 2317 fish).
Smaller-scale reductions in spatial distribution of this species were observed from 2004 through 2018 than in previous years (Figure 1). The observed reductions in the different zones of the IRL were remarkably consistent, with an absence of high catches (>50 individuals) and only a few medium catches (11–50 individuals) of pipefish beginning in 2004–2006, followed by a period of slight decline in 2007 and a return to relative stability in 2008 and 2009 across all IRL zones. The greatest change in spatial distribution and abundance throughout the entire study occurred between the 2007–2009 and 2010–2012 time periods. This sharp abundance decrease and spatial contraction were concurrent with the steep decline (~13,000 ha loss) in seagrass coverage between 2009 and 2011 (Figure 1). This was then followed by consistently low catches through the remainder of the study period. Only a few medium catches (~2/year) throughout the entire IRL study area and high catches were nonexistent from 2010 through 2018, with the majority of the seine hauls yielding no S. scovelli (Figure 1). This species was strikingly absent in the BRL from 2010 through 2018 concurrent with overall seagrass reductions or total seagrass losses throughout much of this zone. While the overall spatial distribution of S. scovelli in the IRL was vastly compressed over this recent time interval, the core distribution of the species remained relatively stable, at significantly lower abundance levels, throughout the period of abundance decline compared to pre-2004 years.
Discussion
Substantial negative effects on estuarine fish populations have been directly related to anthropogenic stressors in perturbed estuaries, with many fish species now less likely to occur in more polluted or stressed estuaries (Toft et al. 2018). The population dynamics and responses of sentinel fish species can reveal underlying and sometimes cryptic or unobserved complications in an ecosystem. Syngnathus scovelli is an effective sentinel species in the IRL and would be useful for similar ecosystem applications in estuaries throughout the southeastern United States.
Dramatic long-term declines in the S. scovelli population were observed in the IRL during the study period. While the population declines were concurrent with large-scale declines in seagrass coverage, habitat, and water-quality degradation (Morris et al. 2018, 2022; Lapointe et al. 2020), moderate declines in S. scovelli were observed with moderate downward trends in seagrasses before the onset of the precipitous seagrass reductions observed during the middle of the study period (2010–2012). The declines observed in this sentinel species indicate chronic impairment to the estuary that may have pushed the system beyond a functional threshold for many fish species, including those that support regional fisheries, despite ongoing efforts to restore lost habitat and reduce excess nutrient inputs (Adams et al. 2019).
Pipefish species are particularly sensitive to anthropogenic stressors due to their unique reproductive strategy: they are live bearers with males carrying developing offspring. In estuarine systems, live bearers, such as pipefishes, have exhibited a higher positive correlation with anthropogenic disturbance than egg guarders, egg scatterers, and broadcast spawners (Iacarella et al. 2018). In addition, environmental contaminants can significantly affect sexual selection in pipefish and influence population viability (Partridge 2009) and male S. scovelli receptivity and fitness (Rose et al. 2013). Persistent organic contaminants (e.g., PCBs, organochlorine pesticides, herbicides) have been detected in pipefish from other lagoon ecosystems (Roche et al. 2009), and laboratory studies with pipefish from the U.S. Atlantic demonstrated that exposure to PCBs may have negative physiological effects on reproduction during embryo development (Ripley and Foran 2008). Similarly, synthetic estrogen found in human contraceptives and widely distributed within aquatic ecosystems from wastewater treatment effluent caused reproductive failure in male S. scovelli in laboratory exposure studies (Rose et al. 2013). An array of known endocrine-disrupting compounds has been found in other fishes in the IRL (Weijs et al. 2015; Xiaohong et al. 2017; Bangma et al. 2018), but the direct effects of these contaminants on S. scovelli reproductive output and population viability in the IRL are not well understood.
Reproductive impairment has been directly observed with regard to reduced brood sizes in mature male S. scovelli during our monitoring efforts in the IRL. Typical brood sizes of mature male S. scovelli can range from 30 to 92 eggs or embryos (Gasparini and Teixeira 1999). In Florida waters, this species has been described as breeding year-round (Joseph 1957; Brown 1972) and most males are pregnant during the majority of the breeding season (Flanagan et al. 2014). We have observed reduced brood sizes in several mature males, including one of 73 mm standard length (specimen UF 244074), collected on March 1, 2018, in the IRL, carrying only 3 late-term embryos (Supplemental Material Figure S1). Further research is needed to determine whether these are isolated abnormalities or if they are an increasing phenomenon, i.e., what is the extent and what are the causes of these reduced broods for S. scovelli in the IRL, as well as, whether reproductive fitness and abundances observed in the population have been reduced as a result.
Factors such as turbidity can also have negative effects on the viability of pipefish populations (Sundin et al. 2010). Turbidity due to algae blooms directly impairs male pipefishes’ choice of larger and possibly more fecund females (Sundin, et al. 2016). This finding focused on the reduced ability to function where sexual selection can behaviors rely on vision, however more recent research suggests that negative effects of visibility reduced by algal blooms may be tempered when pipefish can use other senses (Sundin et al. 2017). In addition, pipefish and the closely related seahorses are visual feeders that have been found to feed mostly during the crepuscular portions of the day and rarely to never at night (Howard and Koehn 1985; Felício et al. 2006). Just as feeding by pipefish is reduced during periods of hypoxia (Ripley and Foran 2007), which sporadically occur in the IRL, food intake is also likely reduced with the diminished water clarity caused by long-duration harmful algal blooms, which occur in the IRL, as visual detection of prey and feeding effectiveness may be lessened. Effective prey detection may have been further exacerbated by the subsequent loss of seagrass habitat. Large reductions in seagrasses can potentially contribute to a loss of sediment stabilization and decreased water clarity in the study area (de Boer, 2007).
Syngnathus scovelli is often clearly associated with seagrass ecosystems (Diaz-Ruiz et al. 2000; Matheson et al. 2008; Hughes et al. 2009), and a higher catch per unit effort in seine hauls over habitat with 100% or moderate to high (≥25%) seagrass coverage observed in this study further illustrate that association. In mesocosm experiments, S. scovelli was referred to as a habitat generalist among Gracilaria spp., Caulerpa spp. and the seagrass Halodule wrightii, but had a strong preference for seagrass habitats (Krejci 2012). While others have observed a positive relationship between the abundance of S. scovelli and that of submersed aquatic vegetation in the core range of this species (Thayer and Chester 1989; Kulczycki et al. 1981; Rydene and Matheson, 2003; Matheson et al. 2008), S. scovelli does occur to a limited degree in other habitat types (Targett 1984; Matheson et al. 2008). Our data indicate that S. scovelli does inhabit areas in lagoon basins with low seagrass cover or no seagrass cover and may use alternative habitats (e.g., Caulerpa spp., drift algae Gracilaria spp., sponge reefs). This species is also found in seagrass areas with damage from boat propellers and may occur in higher densities there than in unscarred areas with less edge effect (Bell et al. 2002). Seagrass habitats with boat propeller scarring are commonly observed during FWC-FWRI Fisheries-Independent Monitoring. Optimal prey may occur in seagrass habitat for this often seagrass-associated species, although some species of pipefish have been found in higher abundance at the edges of seagrass habitats than in the middle of seagrass beds, which was found to be related to prey availability and associated factors (Macreadie et al. 2010; Smith et al. 2008, 2011). Syngnathus scovelli in areas of coastal Georgia, USA, at the northernmost portion of its range, have occurred in areas with no seagrass in comparatively small numbers (n = 34 fish, with only 2 adult males) (Targett 1984), but the fitness and viability of that population, or the available prey base in the region, have not been well studied. Pregnant male S. scovelli under low-food conditions have been shown to have lower growth rates than pregnant males under high-food conditions (Paczolt and Jones 2015). Similarly, reliance on seagrasses for hydrodynamic shelter has been demonstrated for pipefishes and the degradation or loss of seagrass habitats can increase energy expenditures that could contribute to subsequent pipefish population declines (Castejón-Silvo et al. 2021).
Declines in related pipefish species have been observed in other estuarine habitats where a >95% reduction of critical seagrass habitat has occurred (O’Leary et al. 2021). The occurrence of the most threatened syngnathid species in the world, the Critically Endangered estuarine pipefish Syngnathus watermeyeri, was positively related to the availability of submersed aquatic vegetation, and loss of this important habitat has caused local extinction of the species (Claassens et al. 2022). The significantly reduced seagrass habitat in the IRL in recent years most certainly has negative effects on the IRL S. scovelli population and may have directly resulted in the declines we observed. Collectively, the lack of available seagrass habitat may reduce optimum seagrass-associated prey, increase energetic costs, increase predation as a result of lack of appropriate cover, and ultimately lead to negative growth rates and reproductive impairment.
Population genomics of S. scovelli from estuarine waters of the Florida Atlantic coast indicate that specimens collected from the Cape Canaveral area of the IRL to the north and directly to the south of our core study area comprise one genetic population (Flanagan et al. 2016). If the large-scale population declines observed in our study continue the current abundance and distribution trajectories, there may be concern regarding continued maintenance of population connectivity of the northernmost and southernmost portion of the population in estuarine waters of the Florida Atlantic coast. Continued monitoring of this portion of the population and expanded monitoring in the region are of critical importance given the recently declining habitat conditions and water quality-related issues in the IRL ecosystem.
The results of this study provide evidence of a steep decline in a formerly robust S. scovelli population that reflects changes in the overall ecological health of the Indian River Lagoon system. These changes may suggest broader declines around peninsular Florida and in similar estuarine systems throughout the southeastern United States for this and related species. Detection of abundance and distribution trends is only possible where effective long-term monitoring programs exist. There is a critical need for long-term monitoring of syngnathids, as abundance estimates and population changes for these species can rely solely on trade surveys in some parts of the world (Vincent et al. 2011). Expanded research regarding population dynamics of pipefishes in other estuaries and comprehensive study of the mechanics of potential reproductive impairment in this and other syngnathids is warranted. Recommended conservation efforts for syngnathids have focused upon reducing fishing pressure and protecting critical habitat (e.g., seagrasses) supported by effective monitoring efforts (Pollom et al. 2021). For pipefishes such as S. scovelli, directly affected by habitat degradation, reducing seagrass habitat loss needs to be a central management focus. Early detection of decline in sentinel species, including S. scovelli, allows for more rapid implementation of management actions to reduce the effects of anthropogenic pressures on our aquatic systems and the fishes and fisheries they support.
Data availability
The datasets generated and/or analyzed during the current study are available from the corresponding author upon reasonable request.
References
Abbott GM, Landsberg JH, Reich AR, Steidinger KA, Ketchen S, Blackmore C (2009) Resource guide for public health response to harmful algal blooms in Florida. Technical report TR-14. St. Petersburg, FL: Florida Fish and Wildlife Conservation Commission, Fish and Wildlife Research Institute. https://myfwc.com/research/redtide/research/scientific-products/tr14/
Adams DH, Sonne C, Basu N, Dietz R, Nam D-H, Leifsson PS, Jensen AL (2010) Mercury contamination in spotted seatrout, Cynoscion nebulosus: an assessment of liver, kidney, blood, and nervous system health. Sci Total Environ 408:5808–5816. https://doi.org/10.1016/j.scitotenv.2010.08.019
Adams DH, Tremain DM, Paperno R, Sonne C (2019) Florida lagoon at risk of ecosystem collapse. Science 365:991–992. https://doi.org/10.1126/science.aaz0175
Bangma JT, Reiner JL, Lowers RH, Cantu TM, Scott J, Korte JE, Scheidt DM, McDonough C, Tucker J, Back B, Adams DH, Bowden JA (2018) Perfluorinated alkyl acids and fecundity assessment in striped mullet (Mugil cephalus) at Merritt Island National Wildlife Refuge. Sci Total Environ 619–620:740–747. https://doi.org/10.1016/j.scitotenv.2017.11.141
Bell SS, Hall MO, Soffian S, Madley K (2002) Assessing the impact of boat propeller scars on fish and shrimp utilizing seagrass beds. Ecol Appl 12:206–217. https://doi.org/10.1890/1051-0761(2002)012[0206:ATIOBP]2.0.CO;2
Bergman MJ, Donnangelo LJ (1998) Simulation of freshwater discharges from ungaged areas to the Sebastian River, Florida. J Am Water Resour Assoc 36:1121–1132. https://doi.org/10.1111/j.1752-1688.2000.tb05715.x
Bortone SA, Martignette AJ, Spinelli JP (2006) Spotted seatrout (family Sciaenidae): growth as an indicator of estuarine conditions in San Carlos Bay, Florida. Florida Sci 69:127–139
Brown JD (1972) A comparative life history study of four species of pipefishes (family Syngnathidae) in Florida. Doctoral thesis. University of Florida
Carstensen J, Sánchez-Camacho M, Duarte CM, Krause-Jensen D, Marbà N (2011) Connecting the dots: responses of coastal ecosystems to changing nutrient concentrations. Environ Sci Technol 45(21):9122–9132. https://doi.org/10.1021/es202351y
Castejón-Silvo I, Terrados J, Nguyen T, Jutfelt F, Infantes E (2021) Increased energy expenditure is an indirect effect of habitat structural complexity loss. Funct Ecol 35(10):2316–2328. https://doi.org/10.1111/1365-2435.13876
Claassens L, de Villiers NM, Seath J, Wasserman J (2022) Distribution and density of the Critically Endangered estuarine pipefish across its range – implications for conservation. Aquat Conserv: Mar Freshw Ecosyst 32:28–41. https://doi.org/10.1002/aqc.3742
de Boer WF (2007) Seagrass-sediment interactions, positive feedbacks and critical thresholds for occurrence: a review. Hydrobiologia 591(1):5–24. https://doi.org/10.1007/s10750-007-0780-9
Diaz-Ruiz S, Aguirre-Leon A, Perez-Solis O (2000) Distribution and abundance of Syngnathus louisianae and Syngnathus scovelli (Syngnathidae) in Tamiahua Lagoon, Gulf of Mexico. Cienc Mar 26(1):125–143. https://doi.org/10.7773/cm.v26i1.567
FBEBR. [Florida] Bureau of Economic and Business Research. (2011) Florida population: census summary. University of Florida, Bureau of Economic and Business Research. https://www.bebr.ufl.edu/sites/default/files/Research%20Reports/census_summary_2010.pdf
FBEBR. [Florida] Bureau of Economic and Business Research. (2021) Projections of Florida population by county, 2025–2045, with estimates for 2020. University of Florida, Bureau of Economic and Business Research. https://www.bebr.ufl.edu/wp-content/uploads/2021/10/projections_2021_asrh.pdf
Felício AKC, Rosa IL, Souto A, Freitas RH (2006) Feeding behavior of the longsnout seahorse Hippocampus reidi Ginsburg, 1933. J of Ethol 24(3):219–225. https://doi.org/10.1007/s10164-005-0189-8
Flanagan SP, Johnson JB, Rose E, Jones AG (2014) Sexual selection on female ornaments in the sex-role-reversed Gulf pipefish (Syngnathus scovelli). J Evol Biol 27:2457–2467. https://doi.org/10.1111/jeb.12487
Flanagan SP, Rose E, Jones AG (2016) Population genomics reveals multiple drivers of population differentiation in a sex-role-reversed pipefish. Mol Ecol. 25:5043–5072. https://doi.org/10.1111/mec.13794
Gasparini JL, Teixeira RL (1999) Reproductive aspects of the Gulf pipefish, Syngnathus scovelli (Teleostei: Syngnathidae), from southeastern Brazil. Braz J Biol 59(1):87–90. https://doi.org/10.1590/S0034-71081999000100011
Glibert, PM (2020) Harmful algae at the complex nexus of eutrophication and climate change. Harm Alg 91:101583. https://doi.org/10.1016/j.hal.2019.03.001
Gobler, CJ (2020) Climate Change and Harmful Algal Blooms: insights and perspective. Harm Alg 91:101731. https://doi.org/10.1016/j.hal.2019.101731
Griffith AW, Gobler CJ (2020) Harmful algal blooms: a climate change co-stressor in marine and freshwater ecosystems. Harm Alg 91:101590. https://doi.org/10.1016/j.hal.2019.03.008
Howard RK, Koehn JD (1985) Population dynamics and feeding ecology of pipefish (Syngnathidae) associated with eelgrass beds of Western Port, Victoria. Aust J Mar Freshwat Res 36:361–370. https://doi.org/10.1071/MF9850361
Hughes AR, Williams SL, Duarte CM, Heck KL Jr, Waycott M (2009) Associations of concern: declining seagrasses and threatened dependent species. Front Eco Environ 7(5):242–246. https://doi.org/10.1890/080041
Iacarella JC, Adamczyk E, Bowen D, Chalifour L, Eger A, Heath W, Helms S, Hessing‐Lewis M, Hunt BP, MacInnis A, O’Connor MI, Robinson VLK, Yakimishyn J, Baum JK (2018) Anthropogenic disturbance homogenizes seagrass fish communities. Glob change bio 24(5):1904–1918. https://doi.org/10.1111/gcb.14090
IUCN (International Union for Conservation of Nature and Natural Resources) (2013) Syngnathus scovelli. The IUCN Red List of Threatened Species 2013: e.T202406A18229413. https://doi.org/10.2305/IUCN.UK.2011.RLTS.T202406A18229413.en
Jones AG, Avise JC (1997) Microsatellite analysis of maternity and the mating system in the Gulf pipefish Syngnathus scovelli, a species with male pregnancy and sex-role reversal. Mol Ecol 6:203–213. https://doi.org/10.1046/j.1365-294X.1997.00173.x
Joseph EB (1957) A study of the systematics and life history of the Gulf pipefish, Syngnathus scovelli (Evermann and Kendall). Dissertation, Florida State University
Kennish MJ (2002) Environmental threats and environmental future of estuaries. Environ Conserv 29:78–107. https://doi.org/10.1017/S0376892902000061
Krejci SE (2012) Habitat preferences and the effects of seagrass density on population demographics and feeding ecology of pipefish in the Indian River Lagoon, FL. Dissertation, Florida Institute of Technology
Kulczycki GR, Virnstein RW, Nelson WG (1981) The relationship between fish abundance and algal biomass in a seagrass-drift algae community. Estuar Coast Shelf Sci 12(3):341–347. https://doi.org/10.1016/S0302-3524(81)80130-2
Landsberg JH, Hall S, Johannessen JN, White KD, Conrad SM, Abbott JP, Flewelling LJ, Richardson RW, Dickey RW, Jester EL, Etheridge SM, Deeds JR, Van Dolah FM, Leighfield TA, Zou Y, Beaudry CG, Benner RA, Rogers PL, Scott PS, Kawabata K, Wolny JL, Steidinger KA (2006) Saxitoxin puffer fish poisoning in the United States, with the first report of Pyrodinium bahamense as the putative toxin source. Environ Health Perspect 114(10):1502–7. https://doi.org/10.1289/ehp.8998
Lapointe BE, Herren LW, Brewton RA, Alderman PK (2020) Nutrient over-enrichment and light limitation of seagrass communities in the Indian River Lagoon, an urbanized subtropical estuary. Sci Total Environ 699:134068. https://doi.org/10.1016/j.scitotenv.2019.134068
Lapointe BE, Herren LW, Debortoli DD, Vogel M (2015) Evidence of sewage driven eutrophication and harmful algal blooms in Florida’s Indian River Lagoon. Harmful Algae 43:82–102. https://doi.org/10.1016/j.hal.2015.01.004
Lopez CB, Tilney CL, Muhlbach E, Bouchard JN, Célia Villac M, Henschen KL, Markley LR, Abbe SK, Shankar S, Shea C, Flewelling L, Garrett M, Badylak S, Phlips EJ, Hall L, Lasi MA, Parks A, Paperno R, Adams DH et al (2021) High-resolution spatioemporal dynamics of harmful algae in the Indian River Lagoon (Florida): a case study of Aureoumbra lagunensis, Pyrodinium bahamense, and Pseudo-nitzschia. Front Mar Sci 8:769877. https://doi.org/10.3389/fmars.2021.769877
Lotze HK, Lenihan HS, Bourque BJ, Bradbury RH, Cooke RG, Kay MC, Kidwell SM, Kirby MX, Peterson CH, Jackson JBC (2006) Depletion, degradation, and recovery potential of estuaries and coastal seas. Science 312:1806–1809. https://doi.org/10.1126/science.1128035
Macreadie PI, Hindell JS, Keough MJ, Jenkins GP, Connolly RM (2010) Resource distribution influence positive edge effects in a seagrass fish. Ecol 91(7):2013–2021. https://doi.org/10.1890/08-1890.1
Matheson RE Jr, Adams DH, Keenan SF, Switzer TS (2008) Status, trends, and habitat use by estuarine and marine fishes of greatest conservation need in Florida. Florida Fish and Wildlife Conservation Commission, Fish and Wildlife Research Institute. https://f50006a.eos-intl.net/F50006A/OPAC/Details/Record.aspx?BibCode=1437378
McGlathery KJ, Sundbäck K, Anderson IC (2007) Eutrophication in shallow coastal bays and lagoons: the role of plants in the coastal filter. Mar Ecol Progr Ser 348:1–18. https://doi.org/10.3354/meps07132
Morris L, Hall L, Chamberlain R, Jacoby C (2018) Summary report for the northern Indian River Lagoon. In: Yarbro L, Carlson PR (eds) Seagrass integrated mapping and monitoring report no. 3. Fish and Wildlife Research Institute Technical Report TR-17 version 3. St. Petersburg, FL, pp. 1–10. https://myfwc.com/media/17702/simm3-northern-indian-river-lagoon.pdf
Morris LJ, Hall LM, Jacoby CA, Chamberlain RH, Hanisak MD, Miller JD, Virnstein RW (2022) Seagrass in a Changing Estuary,the Indian River Lagoon, Florida, United States. Front. Mar. Sci. 8:789818. https://doi.org/10.3389/fmars.2021.789818
Morris LJ, Hall LM, Miller JD, Lasi MA, Chamberlain RH, Virnstein RW, Jacoby CA (2021) Diversity and distribution of seagrasses as related to salinity, temperature, and availability of light in the Indian River Lagoon, Florida. Florida Sci 84(2–3):119–137
O’Leary JK, Goodman MC, Walter RK, Willits K, Pondella DJ, Stephens J (2021) Effects of estuary-wide seagrass loss on fish populations. Estuaries Coasts 44(8):2250–2264. https://doi.org/10.1007/s12237-021-00917-2
Paczolt KA, Jones AG (2015) The effects of food limitation on life history tradeoffs in pregnant male Gulf pipefish. Plos One 10(5):e0124147. https://doi.org/10.1371/journal.pone.0124147
Partridge CG (2009) The effect of environmental contaminants on mating dynamics and population viability in a sex-role reversed pipefish. Texas A&M University.
Phelps NB, Bueno I, Poo-Muñoz DA, Knowles SJ, Massarani S, Rettkowski R, Shen L, Rantala H, Phelps PL, Escobar LE (2019) Retrospective and predictive investigation of fish kill events. J Aquat Anim Health 31(1):61–70. https://doi.org/10.1002/aah.10054
Phlips EJ, Badylak S, Lasi MA, Chamberlain R, Green WC, Hall LM, Hart JA, Lockwood JC, Miller JD, Morris LJ, Steward JS (2015) From red tides to green and brown tides: bloom dynamics in a restricted subtropical lagoon under shifting climatic conditions. Estuaries Coasts 38:886–904. https://doi.org/10.1007/s12237-014-9874-6
Phlips EJ, Badylak S, Youn S, Kelley K (2004) The occurrence of potentially toxic dinoflagellates and diatoms in a subtropical lagoon, the Indian River Lagoon, Florida, USA. Harmful Algae 3:39–49. https://doi.org/10.1016/j.hal.2003.08.003
Pollom R, Ralph G, Pollock C, Vincent A (2021) Global extinction risk for seahorses, pipefishes and their near relatives (Syngnathiformes). Oryx 55:497–506. https://doi.org/10.1017/S0030605320000782
Ripley JL, Foran CM (2007) Influence of estuarine hypoxia on feeding and sound production by two sympatric pipefish species (Syngnathidae). Mar Environ Res 63:350–367. https://doi.org/10.1016/j.marenvres.2006.10.003
Ripley JL, Foran CM (2008) Interspecific differences of paternal polychlorinated biphenyl exposure on nutrient availability, egg production and brooding in two Syngnathus species. J Fish Biol 72:1369–1385. https://doi.org/10.1111/j.1095-8649.2008.01803.x
Roche H, Vollaire Y, Persic A, Buet A, Oliveira-Ribeiro C, Coulet E, Banas D, Ramade F (2009) Organochlorines in the Vaccarès Lagoon trophic web (Biosphere Reserve of Camargue, France). Environ Pollut 157:2493–2506. https://doi.org/10.1016/j.envpol.2009.03.016
Rose E, Paczolt KA, Jones AG (2013) The effects of synthetic estrogen exposure on pre-mating and post-mating selection episodes in sex-role-reversed Gulf pipefish. Evol Appl 6:1160–1170. https://doi.org/10.1111/eva.12093
Rydene DA, Matheson RE Jr (2003) Diurnal fish density in relation to seagrass and drift algae cover in Tampa Bay, Florida. Gulf Mex Sci 21(1):35–58. https://doi.org/10.18785/goms.2101.04
Shokri MR, Gladstone W, Jelbart J (2009) The effectiveness of seahorses and pipefish (Pisces: Syngnathidae) as a flagship group to evaluate the conservation value of estuarine seagrass beds. Aquat Conserv: Mar Freshw Ecosyst 19:588–595. https://doi.org/10.1002/aqc.1009
Smith T, Hindell J, Jenkins G, Connolly R, Keough M (2011) Edge effects in patchy seagrass landscapes: the role of predation in determining fish distribution. J Exp Mar Biol Ecol 399:8–16. https://doi.org/10.1016/j.jembe.2011.01.010
Smith TM, Hindell JS, Jenkins GP, Connolly RM (2008) Edge effects on fish associated with seagrass and sand patches. Mar Ecol Prog Ser 359:203–213. https://doi.org/10.3354/meps07348
Sokal RR, Rohlf FJ (1981) Biometry, 2nd edn. WH Freeman, New York, pp 859
Steward JS, Virnstein RW, Lasi MA, Morris LJ, Miller JD, Hall LM, Tweedale WA (2006) The impacts of the 2004 hurricanes on hydrology, water quality, and seagrass in the central Indian River Lagoon, Florida. Estuaries Coasts 29:954–965. https://doi.org/10.1007/BF02798656
Sundin J, Aronsen T, Rosenqvist G, Berglund A (2017) Sex in murky waters: algal-induced turbidity increases sexual selection in pipefish. Behav Ecol Sociobiol 71(5):78. https://doi.org/10.1007/s00265-017-2310-8
Sundin J, Berglund A, Rosenqvist G (2010) Turbidity hampers mate choice in a pipefish. Ethol 116:713–721. https://doi.org/10.1111/j.1439-0310.2010.01787.x
Sundin J, Rosenqvist G, Myhren S, Berglund A (2016) Algal turbidity hampers ornament perception, but not expression, in a sex-role reversed pipefish. Ethol 122:215–225. https://doi.org/10.1111/eth.12461
Targett TE (1984) A breeding population of Gulf pipefish (Syngnathus scovelli) in a Georgia estuary. Contrib Mar Sci 27:169–174
Thayer GW, Chester AJ (1989) Distribution and abundance of fishes among basin and channel habitats in Florida Bay. Bull Mar Sci 44(1):200–219
Toft JD, Munsch SH, Cordell JR, Siitari K, Hare VC, Holycross BM, DeBruyckere LA, Greene CM, Hughes BB (2018) Impact of multiple stressors on juvenile fish in estuaries of the northeast Pacific. Glob Change Biol 24:2008–2020. https://doi.org/10.1111/gcb.14055
Vincent ACJ, Foster SJ, Koldewey HJ (2011) Conservation and management of seahorses and other Syngnathidae. J Fish Biol 78:1681–1724. https://doi.org/10.1111/j.1095-8649.2011.03003.x
Weijs L, Briels N, Adams DH, Lepoint G, Das K, Blust R, Covaci A (2015) Bioaccumulation of organohalogenated compounds in sharks and rays from the southeastern USA. Environ Res 137:199–207. https://doi.org/10.1016/j.envres.2014.12.022
Xiaohong X, Xue J, Liu W, Adams DH, Kannan K (2017) Trophic magnification of parabens and their metabolites in a subtropical marine food web. Environ Sci Technol 51(2):780–789. https://doi.org/10.1021/acs.est.6b05501
Acknowledgements
We thank the dedicated scientists of the Fisheries-Independent Monitoring program at the Indian River Field Laboratory of the Florida Fish and Wildlife Conservation Commission’s Fish and Wildlife Research Institute for their assistance in collecting samples. We are grateful to Phil Stevens, Jon Peake, Bland Crowder, and 2 anonymous reviewers for providing valuable feedback and editorial comments. We greatly appreciate the efforts and expertise of Zach Randall, Florida Museum of Natural History, in preparing the S. scovelli computed tomography (CT) scan figure, and guidance from Howie Brown regarding FWC-FWRI’s Fish Kill Hotline database. We also appreciate additional insights and seagrass data provided by Lori Morris and Lauren Hall of the St. Johns River Water Management District.
Funding
This research was supported by the U.S. Department of the Interior, U.S. Fish and Wildlife Service, under the Federal Aid in Sport Fish Restoration Program, and by proceeds from State of Florida saltwater recreational fishing licenses
Author information
Authors and Affiliations
Contributions
DHA conceptualization, Formal analysis, Writing—original DRAFT. AS Formal analysis, Writing—review and editing. RP Formal analysis, Writing–review and editing, Donut administration.
Corresponding author
Ethics declarations
Conflict of Interest
The authors declare no competing interests.
Ethics approval
No animal testing was performed during this study.
Sampling and field studies
All necessary permits for sampling have been obtained with permitting authorized by the Florida Fish and Wildlife Conservation Commission within waters of the State of Florida.
Additional information
Communicated by S.E. Lluch-Cota
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Fig. S1
Top image: computerized tomography scan of a male Syngnathus scovelli (FSBC 28276; UF 244279) collected from the Indian River Lagoon in March 1991, with a typically normal brood size, n = 40 developing embryos (in blue). Bottom image: computerized tomography scan of a male S. scovelli (UF 244074) collected from the Indian River Lagoon in March 2018, with a reduced brood size, n = 3 developing embryos (in blue). Computerized tomography scans by Zach Randall, Florida Museum of Natural History. (JPG 3238 kb)
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

{kind=link}
Cite this article
Adams, D.H., Sebastian, A. & Paperno, R. Population decline of Gulf pipefish (Syngnathus scovelli) in a subtropical estuary: ecosystem changes and habitat loss. Mar. Biodivers. 52, 57 (2022). https://doi.org/10.1007/s12526-022-01289-x
Received:
Revised:
Accepted:
Published:
DOI: https://doi.org/10.1007/s12526-022-01289-x