
Abstract
Diet often plays a vital role in defining social divisions within and between social groups and thus can be used to understand the social paradigms of archeological cultures. During the Early Avar period (568–630 A.D.), burial evidence indicates that there were strong demarcations of social stratification and divisions between sexes and age groups; however, the symbols of intra-population heterogeneity become increasingly rare during the Late Avar period (680–822 A.D.). In this study, we investigate social differences expressed through diet in the cemetery population from Nuštar, eastern Croatia (eighth to early ninth century), to determine whether dietary social disparities existed during the Late Avar period in this region. Stable isotope analysis of carbon (δ13C) and nitrogen (δ15N) from bone and dentine collagen shows no dietary differences, neither between high, middle, and low status individuals, nor between males, females, and juveniles. These results likely reflect the outcome of the social homogenization process that began after the failed Avar attack on Constantinople in 626 A.D. Geographical patterning is visible when the data from Nuštar is compared to data from other Middle and Late Avar sites. While Avar sites in the southern and south-eastern frontiers of the Avar qaganate do not display dietary differences between sexes, previous isotopic work on populations in Lower Austria shows that males consumed a higher proportion of animal protein than females. This is likely the result of Frankish influence and reflects diversity in social practices within the Avar qaganate itself during the Middle and Late Avar periods.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Diet is often an accurate proxy for social status, especially in Europe, where food has been extensively linked to social roles (Bourbou et al. 2011; Hakenbeck et al. 2010; Lightfoot et al. 2012; Privat et al. 2002). Human social organization is directly impacted by behavioral predispositions, which include the tendency to form intra-group social identities and the quest to seek status, both of which have been linked to dietary practices. This is exemplified by the popular phrase “you are what you eat”, which has both a social and a biophysical basis (Wiessner and Schiefenhövel 1996). It has been widely accepted that people tend to consume foods deemed to be suitable and desired by their society and social group. Evaluations of what is deemed palatable are socially subjective. As Nick Fiddes states: “The foods we select reflect our thought, including our conception of our actual or desired way of life and our perceptions of the food choices of people with whom we wish to identify … We feed not only our appetite but also our desire to belong” (Fiddes 1991, pp. 33–34). Therefore, by studying what specific groups within societies ate, it is possible to discern past social divisions, including status, age, and sex.
Through cultures what is eaten serves as a way to define the boundaries of group identities. Consumption and the rituals associated with eating are parameters that define the value and status that individuals obtain through the act of eating (Grant 2002). It is ethnologically recognized that “some foods confer high status on the eaters” (Fieldhouse 1986, p. 77). However, the kinds of food that attain more or less value, and a specific item’s relationship to social status vary cross culturally (Wiessner and Schiefenhövel 1996). For example, the perceived value of swans in Medieval England was such that by the twelfth century, the royal crown had asserted prerogative rights over them (Sykes 2014). Similarly, wealthy individuals buried at the Anglo-Saxon cemetery in Berinsfield, Oxfordshire consumed a higher proportion of terrestrial herbivores while their lower class counterparts resorted to aquatic and/or omnivore protein sources. Economically, domesticate animals required a lot more time and resources to produce so that their consumption spoke to the consumer’s ability to afford such luxury (Privat et al. 2002). Thus, diets with a higher proportion highly valued items conferred a higher status. This theory can be applied to Nuštar, as it is clear from burial rites that sex, age, and social status were clearly defined through grave goods. The question of whether these divisions existed during life will be tested through a dietary analysis.
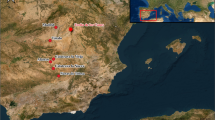
The Avar political structure was nomadic in nature. They were original to the Euroasian Steppe. In 568 A.D., they arrived and settled in the Carpathian Basin, bringing all the local groups under one rule (Curta 2006; Daim 1984; Stadler 2008). Although not much is known of them historically compared to contemporary cultures such as the Franks and Byzantines, recent archeological work has exposed many aspects of Avar culture. The Avar tradition of depositing ornamental grave goods and food offerings in burials allows for social status of the individual to be inferred. The present study aims to correlate status differences (based upon funerary analysis) and dietary patterns during life through an individualized dietary analysis. This study analyzes the skeletal remains of 44 individuals and 17 animals recovered from the Late Avar cemetery at Nuštar, eastern Croatia. It is hypothesized that diet showed strong differences between social groups, as food is deeply entrenched in the formation and demarcation of social identities. To test this hypothesis, the bone and dentine δ13C and δ15N values and the enamel δ18O values will be determined for the population buried at the Nuštar cemetery, and the results will be compared against the finds from other contemporary sites in the Carpathian Basin (Fig. 1). By comparing our results to data from other regions within the qaganate, it is possible to determine large-scale cultural trends within the Avar culture, rather than isolated examples.

Map showing the location of Nuštar plus other Avar and Slavic sites mentioned in the study. Map created using ArcGIS version 10.2.2. for PC. Source: US National Parks Service
Background
The Avars
Archeological research, such as funerary analysis, has formed much of our understanding of Avar society and political structure. The presence of princely burials among other poorer graves, the structure of many cemeteries, and the furbishing and depth of the graves demonstrate strong divisions between sexes and social groups during the Early Avar period, but the evidence for this becomes scarce during the Late Avar period (Daim 1984; Vida 2008; Young 1978; Szõke 2003).
The Avars cultivated millet and to a lesser extent barley, while rye was also cultivated during the Late Avar period (Gyulai 2006, 2014). Animal bones in graves show that cattle, pigs, sheep, goats, and poultry were raised and probably consumed (Curta 2006; Vida 2003). Macro-archaeobotanical remains of broomcorn millet (Panicum miliaceum L.), bread wheat (Triticum cf. aestivum (L.) subsp. aestivum), foxtail millet (Setaria italica (L.) P. Beauv.), rye (Secale cereale L.), and barley (Hordeum vulgare L.) have been identified at Nuštar (Rapan Papeša et al. 2015). Dental evidence from three-seventh to eleventh century coastal Croatian sites suggests that carbohydrates played a major role in diet, focusing on hard fibrous foods (Šlaus et al. 2011); however, whether these results are mirrored in the continental region is unknown. Isotopic work from Avar sites in Leobersdorf, Zwölfaxing, Wien-Csokorgasse, Lower Austria (Herold 2008), and Sajópetri, Hungary (Noche-Dowdy 2015) suggests a mainly C3 diet with some contribution of C4 plants (likely millet) and terrestrial animal protein (Table 1).
It is expected that Avar elites consumed significant amounts of animal protein while their lower status counterparts subsisted on diets composed of predominantly plant resources. Archeologically, this pattern has been identified across cultures through stable isotope analyses, which show elites enjoying a higher proportion of animal protein in their diet in a variety of different contexts (Ambrose et al. 2003; Privat et al. 2002; Reitsema and Vercellotti 2012). Within Middle and Late Avar sites in Lower Austria, isotopic differences have been found between males and females, with males eating a higher proportion of animal protein than females, but no dietary differences were observed between age groups (Herold 2008). However, contradictory results were obtained from Sajópetri, Hungary, a Middle and Late Avar site where no isotopic differences were found between sexes nor social status groups, which were defined by grave goods (Noche-Dowdy 2015). Previous studies therefore present an ambiguous picture; the present study seeks to contribute to this discussion with data from the Late Avar cemetery at Nuštar.
Archeological work at Nuštar
The site is located in the southern outskirts of Nuštar village, eastern Croatia. Only two thirds of the cemetery was excavated (140 × 100 m), leaving the south and eastern limits unexplored. The excavations produced over 1500 very well-preserved artifacts, 196 graves, and 188 skeletons.
Based on preliminary grave good analysis, the overall population at Nuštar can be regarded as high middle status group dated to the Late Avar period, but some graves might belong to the Middle Avar period (see Online Resource). Out of the 188 individuals excavated, 65 were males, 44 females, 35 adults where sex could not be determined, 42 children and 2 individuals for which neither sex nor age could be identified. These were organized in ten fairly regular burial rows. The length of the graves ranged between 0.93 and 2.57 m (1.79 m average), the width ranged between 0.42 and 1.45 m (0.54 m average), and the depth ranged between 0.05 and 1.44 m. All graves were oriented west-east. Most graves housed one individual, but seven multiple burials were found. All of the deceased were buried in a supine position with their arms laid next to the body. Two male individuals were found buried with horses, and likely represent wealthy individuals.
Scientific background
The carbon and nitrogen isotopic signatures of bone collagen reflect the isotopic signature of the food and drink consumed during the last 10 years of an individual’s life, while teeth capture the isotopic signatures at the time of tooth formation, that is childhood (AlQahtani et al. 2010; Brown and Brown 2011; Hadjidakis and Androulakis 2006; Smith 1991). A comparison of these tissues exposes dietary changes during an individual’s life. Similarly, metabolic differences between apatite (i.e., tooth enamel) and protein (i.e., bone and dentine collagen) expose different dietary information (Koch et al. 1997), with body protein being biased towards the protein component of the diet, but apatite reflecting whole diet (Lee-Thorp 2008).
Carbon isotope values recorded in body tissues reflect different plant photosynthetic pathways, C3, and C4. Most plants, including wheat and barley, use the C3 pathway and have δ13C values varying between − 36 and − 24‰ (Farquhar et al. 1989; Lee-Thorp 2008; van der Merwe and Medina 1991). C4 plants have δ13C values of approximately − 12.5‰ with little variation (Brown and Brown 2011; Lee-Thorp 2008). During the Avar period, millet was the only C4 domesticate and wild C4 plants were (and are) rare. Differences in δ13C values also arise through the consumption of marine foods due to differential isotopic baselines between marine and terrestrial ecosystems (Barrett et al. 2001; Tauber 1981). Given the location of Nuštar, this is unlikely to be a significant factor in this study. δ15N values can be used to study the relative proportion of animal protein consumed by individuals. Note, however, that δ15N values cannot distinguish between different types of animal protein, such as meat and dairy (O’Connell and Hedges 1999). Nitrogen isotope values are affected by biological factors such as pregnancy, growth, nutritional stress, breastfeeding, and pathologies (Fogel et al. 1989; Fuller et al. 2005; Katzenberg and Lovell 1999; Mekota et al. 2006; Waters-Rist and Katzenberg 2010). Due to environmental isotopic variation, which can alter δ13C and δ15N in an ecosystem, local animal isotopic values should be used as references for the human samples (Ambrose 1991; Sealy et al. 1987).
An individual’s δ18O values are ultimately derived from precipitation and ingested via drinking water and water found in food (Sponheimer and Lee-Thorp 1999). Isotopic studies of migration using δ18O values are possible thanks to the different geospecific isotopic ranges of different biomes, which tend to follow climate and weather patterns ultimately linked to geographical factors such as latitude, altitude, distance from the sea, and temperature (Budd et al. 2004). Thus, δ18O values can be used as a proxy for migration (Dupras and Schwarcz 2001; White et al. 1998). In this research, δ18O enamel values will be used to identify immigrants, as these values would have been recorded during childhood when the individual was in a different location to where they were buried.
Materials and methods
A total of 44 individuals divided into approximately equal groups by sex, age, and social status were selected for the study (See Online Resource). Forty-four bone samples, 31 dentine samples and 35 enamel samples were collected for analysis. All but six of the bone samples were taken from ribs, with the remainder taken from long bones. When possible, a permanent tooth from the same individual was sampled for dentine collagen and enamel apatite. Premolars and second molars were targeted as to avoid nursing effects (Fogel et al. 1989; Smith 1991); but canines, first and third molars were sampled when the former were not available. Bone samples of 17 animals (Bos taurus, Ovis/Capra, Equus caballus, Sus, and Canid) present in the graves were also sampled.
Bone samples averaged 0.5 g and were sampled using a dremel hand-held drill with a diamond cutting wheel. All samples were sandblasted to remove environmental contaminants prior to extraction. The collagen was extracted following the standard laboratory protocol of the Department of Archeology, University of Cambridge, as described in Privat et al. (2002). The bone samples were demineralised in 0.5 M aqueous HCl for up to 2 weeks and then gelatinised (75 °C for 48 h in pH 3 water). The sample was then frozen and freeze-dried. Each sample was weighed and analyzed using a Costech automated elemental analyzer coupled in continuous-flow mode to a Finnigan isotope-ratio mass spectrometer (Delta V). Carbon and nitrogen isotopic ratios were measured on the delta scale relative to the international standards of the VPDB for carbon and AIR for nitrogen. Analytical error was < 0.2‰ for carbon and nitrogen based on repeated measurements of international and in house standards. Collagen samples were run in triplicate, and the average of the three measurements was used for the statistical analyses.
Between 6 and 10 mg of enamel were sampled by drilling enamel powder using a dremel hand-held drill with a diamond-tipped drill bit. Sampling was performed in a vertical line along the crown. Enamel surfaces were cleaned by ablation prior to sampling. In one case (NUH19), a piece of enamel was ground manually. Enamel powder was pre-treated following Balasse (2002); samples were treated with 2–3% aqueous sodium hypochlorite (0.1 ml/mg) for 24 h at 4 °C and then rinsed. They were then treated with 0.1 M acetic acid (0.1 ml/mg) for 4 h at room temperature and rinsed again. They were then freeze-dried. Four modern horse dental standards were treated and analyzed in the same manner. The samples were analyzed using an automated gas bench interfaced with a Thermo Finnigan MAT253 isotope-ratio mass spectrometer. Carbon and oxygen isotopic ratios were measured on the delta scale against the international standard VPDB. Analytical error for this instrument has been recorded as less than 0.10‰ for oxygen and 0.08‰ for carbon.
The statistical analyses were performed using the SPSS Statistics 23 package. Normality was tested using Shapiro-Wilk and Kolmogorov-Smirnov tests. Homogeneity of variance was assumed when a Levene’s test p value was greater than 0.05. One-way ANOVA (with Bonferroni post hoc) tests and paired t tests (p > 0.05) were employed to analyze those datasets that followed the assumptions of parametric data. A migrant is defined as an individual whose isotope value lies more than 1.5 times the interquartile range below quartile 1 or above quartile 3.
Results
Preservation and quality assessment
All samples showed good collagen preservation and were included in the subsequent analyses (see Online Resource): the collagen yields ranged from 2.5 to 19.9% (Hillson 2005); the C:N ratios ranged between 3.0 and 3.3 (DeNiro 1985), and carbon and nitrogen yields were above the acceptable 13 and 4.8%, respectively (Ambrose 1990).
Faunal bone collagen results
Although faunal sample size is too small for reliable statistical analyses, clear isotopic differences exist between taxa (Fig. 2, Online Resource). Faunal δ13C values range between − 21.6 and − 14.2‰ with an average of − 17.7 ± 2.1‰ (n = 17). Their δ15N values averaged 8.1 ± 1.2‰ and ranged between 5.6 and 10.3‰. The 10.3‰ value corresponds to a cattle (NUA04) that has been identified as an outlier and excluded from subsequent analysis. Strict herbivores (Bos taurus, Ovicaprid, and Equus caballus; n = 12) have δ15N values ranging between 5.6 and 8.8‰ with an average value of 7.7 ± 1.0‰. The omnivore’s δ15N values range from 7.3 to 10.2‰, with an average value of 8.7 ± 1.2‰ (n = 4).

δ13C and δ15N of bone collagen for all species included in the analysis
Human results
The results from the isotopic analysis of the human samples plus relevant context information are found in the Online Resource. Bone collagen δ13C and δ15N have average values of − 16.4 ± 0.6‰ (range − 17.7 to − 15.0‰, n = 43) and 10.1 ± 0.6‰ (range 8.9 to 11.7‰, n = 43), respectively. Dentine analysis yielded an average δ13C of − 16.1 ± 0.9‰ (range − 18.2 to − 13.7‰, n = 30) and an average δ15N of 10.4 ± 0.8‰ (range 8.9 to 11.8‰, n = 30) (Table 2). When compared against the herbivore averages, average offsets of 1.9 and 2.4‰ were identified for bone collagen δ13C and δ15N, respectively.
Enamel carbonate δ13C yielded an average of − 8.6 ± 1.1‰, (range − 11.1 to − 6.1‰, n = 33). Enamel δ18O values averaged − 7.6 ± 0.6‰ (range − 8.9 and − 6.4‰), with one outlier detected (NUH19, δ18O = − 5.5‰), lying more than 1.5 times the inter-quartile range above quartile 3. This individual will be excluded from statistical analyses.
Statistical analysis showed that there are no differences in any of the isotopic values between males, females, and juveniles (Tables 3 and 4), thus these groups were combined for statistical analyses of status groups. No statistical differences were found between status groups (Tables 5 and 6). Status differences were not compared within sex and age groups, as sample size would have been too small to yield reliable results.
Both males buried with a horse (NUH31 and NUH32) had isotope values similar to that of the other high status males (Fig. 2).
The δ13C and δ15N bone collagen values obtained for the Nuštar population are similar to previous isotopic work on Avar populations from the Carpathian Basin (Fig. 3 and Table 1) (Noche-Dowdy 2015; Herold 2008). Human-faunal δ15N spacing was not compared between Nuštar and the other comparative sites as these studies did not include faunal material in their analysis (Herold 2008; Noche-Dowdy 2015).

Discussion
Faunal diet
The faunal material informs on animal diet and management and serves as a baseline for comparison with the human δ13C and δ15N values. Some individuals have δ13C values indicative of a diet based solely on C3 resources, while other individuals consumed relatively large proportions of C4 resources, probably millet, and, in the case of the omnivores, animals fed on millet. δ15N values varied between species, particularly the horses are depleted in 15N compared to the other herbivores, consistent with differences in their digestive tract (Sponheimer et al. 2003). The high δ15N value for the cattle outlier (NUA04) may be explained by the individual being a nursing animal (Fogel et al. 1989). The pig and the three dogs have δ15N values above the herbivore range, suggesting an omnivorous diet.
Human diet
All collagen and enamel δ13C values indicate a mixed diet containing C3 and C4 resources, with most, if not all, individuals consuming enough C4 (or animals fed on C4) plants for it to be evident in their isotope results (Katzenberg 1989; Schoeninger and DeNiro 1984). This agrees with archaeobotanical work from Nuštar, as broomcorn millet was the most common identifiable cereal in the assemblage (Rapan Papeša et al. 2015). This agrees with previous isotopic work in the region (Herold 2008; Lightfoot et al. 2012; Noche-Dowdy 2015), as well as with the archaeobotanical literature (Gyulai 2006). Enamel apatite values indicate that most humans consumed at least some millet directly rather than only consuming animals fed on C4 plants. The relatively low δ15N values indicate a reliance on terrestrial food sources, consistent with the apparent trend for fish avoidance in the region during the Early Medieval period (Hakenbeck et al. 2010; Herold 2008; Lightfoot et al. 2012; Noche-Dowdy 2015). The difference in δ15N values between the humans and the herbivores, is very low (typically 2.4‰) which can be explained in various ways. It is possible that the humans could have consumed a diet that contained a very low proportion of animal protein. It is more likely, however, that the sampled fauna had atypical diets, as previous research at Sajópetri, Hungary, a Middle to Late Avar Site, has shown that sacrificial chickens in human burials enjoyed meals similar to the human they were buried with and likely different diets than animals intended for consumption (Sykes 2014, pp. 134–135). Another possibility is that the Avar community at Nuštar consumed high proportions of horse and/or deer, which tend to have relatively low δ15N values, compared to sheep, goat, cow, and pig (Giblin 2011).
The δ13C and δ15N bone collagen values obtained for the Nuštar population are fairly typical compared to the results of other bone collagen samples from the Carpathian Basin (Fig. 3 and Table 1). The δ15N values fit well with previously published data (although note that no faunal values were available for comparison of human-faunal offset values), while the δ13C values from Nuštar are slightly higher than those at the other sites.
Human migration
One δ18O outlier was identified—NUH19, a middle status group male, who is likely a migrant to this area. His dietary isotopes indicate that both his childhood and adult diet were similar to the rest of the population’s diets, although his dentine δ15N value is relatively high. Although it is possible that other migrations occurred between areas with similar δ18O values in precipitation, these results indicate that all other individuals were likely born in the same place.
Males versus females
In burial, Avar elite men are accompanied by complex belt-sets, often of rare metals; weapons like the reflex-bow with arrows or a saber and whole mounts were buried in harnesses. Women are accompanied by a variety of jewelry. Colorful necklaces of amber or glass-paste beads, bronze dress clasps, and silver or bronze bead earrings are the most common, but sometimes a silver, bronze, or iron bracelet and a metal pendant are found (Daim 1984; Vida 2008). The belt set was the most outstanding symbol of social status among men. At Nuštar, Avar men were often buried with belt sets, knives, and arrowheads, while female burials contained a larger proportion of finger rings, earrings, bracelets, necklaces, and spindle whorls. Note that while some male burials contained jewelry, some female graves contained knives.
No differences were found in either diet or mobility between males, females, and juveniles. This is somewhat surprising, as dietary differences have been observed between males and females at contemporary sites (although, to the best of our knowledge, juvenile Avar isotope data have not previously been reported). We suggest that this pattern can be explained by regional variability during the Late Avar period, with differences in diet between sexes being common in the north-western parts of the qaganate, but not in the southern borders. This variability may explain why clear dietary differences existed at some Late Avar sites like Zwölfaxing, Leobersdorf, and Wien-Csokorgasse, all located in Lower Austria, while no evidence of disparities between the sexes in diet existed at Sajópetri, northwest Hungary or Nuštar, eastern Croatia (Herold 2008; Noche-Dowdy 2015).
The idea of regional variability within the qaganate is supported by variations in material culture from the Avar qaganate (Stadler 2008), with an apparent cultural division along the Danube that appeared after the Frankish invasion in 791 and 795 A.D. Archeological evidence suggests that the north-western regions adopted Frankish fashions, weapons, horse gear, and social structure, while the eastern and southern territories may have fallen under Bulgar rule. Similarly, peripheral pockets of Avar power, such as the regions of Vojvodina, northern Serbia, and eastern Croatia where Nuštar is located, seem to have become practically independent and struggled to maintain their Avar policies until their demise during the ninth century (Curta 2006). The patterning in sex differentials in diet seems to follow this geographical division during the seven-ninth century (Table 1 and Fig. 1), where cemeteries located in the northwest of the Carpathian Basin (such as Leobersdorf and Zwölfaxing, Lower Austria) show statistically significant differences between male and female diets, while those in the east and south (such as Nuštar, Croatia and Sajópetri, Hungary) do not (Herold 2008; Noche-Dowdy 2015).
Social status
Social status was not found to affect diet during the Late Avar period in the population of Nuštar—no dietary differences were found between status groups and the two individuals buried with horses ate fairly typical diets. It is possible that all groups consumed similar proportions of animal protein from the same species but different cuts of meat and/or proportions of meat to secondary products, which would not be visible isotopically (O’Connell and Hedges 1999). Such differences cannot be seen using stable isotope analysis.
The dietary analysis therefore supports the hypothesis of social homogenization after the transition from the Early Avar period to the Late Avar period. While the Early Avar period was characterized by clearly defined social divisions in burial culture, the evidence of elite burials becomes more scarce and inconsistent after the second half of the seventh century (Daim 1984; Szõke 2003). Similarly, these results agree with those of Sajópetri, Hungary where no dietary differences were identified between social strata (Noche-Dowdy 2015). This suggests that if social status was expressed in diet during the Early Avar period, the homogenization of social groups occurred immediately following the failed attack of Constantinople and during the socio-political downfall that characterized the civil war. However, more research is needed to determine whether social status groups ate isotopically different diets in the Early Avar period, and to determine if this pattern is repeated across the entire of Avar territory.
Conclusion
Broader social patterns are expressed through diet. As such, this study exposes socio-cultural dynamics during the Middle and Late Avar period. The results suggest that some Avar social groups were much more homogeneous than previously thought. Neither social status, as determined by burial furbishment, nor sex were found to relate to diet at Nuštar or at Sajópetri, Hungary (Noche-Dowdy 2015). However, sexual disparities existed at Late Avar sites located in the northwest of the Carpathian Basin (Herold 2008). This geographical patterning supports the hypothesis that regional variation existed within the qaganate at least during the Late Avar period (Stadler 2008). Unfortunately, there is no available data from the northwest Carpathian Basin comparing dietary differences with status group, so it is not possible to determine if the pattern seen with sex across the Avar area is replicated within status groups. Nevertheless, it is likely that the differences between these regions were polarized after the Frankish invasion when the territories in the northwest acculturated into Carolingian traditions, while those in the south-eastern regions adopted Bulgar lifestyles, and pockets of Avar culture continued into the early ninth century in the regions of Vojvodina and northern Serbia (Curta 2006). While more research is clearly needed, this study has shown that Avar society was at once more homogeneous and more heterogeneous than previously assumed, with less diversity seen within sites, but more variation between sites than expected.
References
AlQahtani SJ, Hector MP, Liversidge HM (2010) Brief communication: the London atlas of human tooth development and eruption. Am J Phys Anthropol 142:481–490. https://doi.org/10.1002/ajpa.21258
Ambrose SH (1990) Preparation and characterization of bone and tooth collagen for isotopic analysis. J Archaeol Sci 17:431–451. https://doi.org/10.1016/0305-4403(90)90007-R
Ambrose SH (1991) Effects of diet, climate and physiology on nitrogen isotope abundances in terrestrial foodwebs. J Archaeol Sci 18:293–317. https://doi.org/10.1016/0305-4403(91)90067-Y
Ambrose SH, Buikstra J, Krueger HW (2003) Status and gender differences in diet at Mound 72, Cahokia, revealed by isotopic analysis of bone. J Anthropol Archaeol 22:217–226. https://doi.org/10.1016/S0278-4165(03)00036-9
Balasse M (2002) Reconstructing dietary and environmental history from enamel isotopic analysis: time resolution of intra-tooth sequential sampling. Int J Osteoarchaeol 12:155–165. https://doi.org/10.1002/oa.601
Barrett JH, Beukens RP, Nicholson RA (2001) Diet and ethnicity during the Viking colonization of northern Scotland: evidence from fish bones and stable carbon isotopes. Antiquity 75:145–154. https://doi.org/10.1017/S0003598X00052844
Bourbou C, Fuller BT, Garvie-Lok SJ, Richards MP (2011) Reconstructing the diets of Greek Byzantine populations (6th–15th centuries AD) using carbon and nitrogen stable isotope ratios. Am J Phys Anthropol 146:569–581. https://doi.org/10.1002/ajpa.21601
Brown TA, Brown K (2011) Biomolecular archaeology: an introduction. Wiley, Chichester
Budd P, Millard A, Chenery C, Lucy S, Roberts C (2004) Investigating population movement by stable isotope analysis: a report from Britain. Antiquity 78:127–141. https://doi.org/10.1017/S0003598X0009298X
Curta F (2006) Southeastern Europe in the middle ages, 500-1250. Cambridge University press, Cambridge
Daim F (1984) The Avars: steppe people of Central Europe. Archaeology 37:33–39
DeNiro MJ (1985) Postmortem preservation and alteration of in vivo bone collagen isotope ratios in relation to palaeodietary reconstruction. Nature 317:806–809. https://doi.org/10.1038/317806a0
Dupras TL, Schwarcz HP (2001) Strangers in a strange land: stable isotope evidence for human migration in the Dakhleh Oasis, Egypt. J Archaeol Sci 28:1199–1208. https://doi.org/10.1006/jasc.2001.0640
Farquhar GD, Ehleringer JR, Hubick KT (1989) Carbon isotope discrimination and photosynthesis. Annu Rev Plant Physiol Plant Mol Biol 40:503–537. https://doi.org/10.1146/annurev.pp.40.060189.002443
Fiddes N (1991) Meat: a natural symbol. Routledge, London
Fieldhouse P (1986) Food & Nuterition: customs & culture. Croom Helm, Dover
Fogel ML, Tuross N, Owsley DW (1989) Nitrogen isotope traces of human lactation in modern and archaeological populations. Carnegie Institution of Washington Yearbook 88: 111–117
Fuller BT, Fuller JL, Sage NE, Harris DA, O’Connell TC, Hedges REM (2005) Nitrogen balance and δ15N: why you’re not what you eat during nutritional stress. Rapid Commun Mass Spectrom 19:2497–2506. https://doi.org/10.1002/rcm.2090
Giblin JI (2011) Isotope analysis on the great Hungarian plain: an exploration of mobility and subsistence strategies from the Neolithic to the copper age. Dissertation, The Ohio State University
Grant A (2002) Food, status, and social hierarchy. In: Miracle P, Milner N (eds) Consuming passions and patterns of consumption. McDonald Institute for Archaeological Research, Cambridge, pp 17–24
Gyulai F (2006) Historical plant-biodiversity in the Carpathian Basin. In: Jerem E, Mester Z, Benczes R (eds) Archaeological and cultural heritage preservation. Archaeolingua, Budapest, pp 63–72
Gyulai F (2014) Archaeobotanical overview of rye (Secale cereale L.) in the Carpathian-Basin II from the migration period until the Late Medieval Age. Columella-J Agric Environ Sci 1:37–48
Hadjidakis DJ, Androulakis II (2006) Bone remodeling. Ann N Y Acad Sci 1092:385–396. https://doi.org/10.1196/annals.1365.035
Hakenbeck S, McManus E, Geisler H, Grupe G, O’Connell T (2010) Diet and mobility in Early Medieval Bavaria: a study of carbon and nitrogen stable isotopes. Am J Phys Anthropol 143:235–249. https://doi.org/10.1002/ajpa.21309
Herold M (2008) Sex differences in mortality in Lower Austria and Vienna in the Early Medieval period: an investigation and evaluation of possible contributing factors. Dissertation, University of Vienna
Hillson S (2005) Teeth, 2nd ed, Cambridge manuals in archaeology. Cambridge University Press, Cambridge
Katzenberg MA (1989) Stable isotope analysis of archaeological faunal remains from southern Ontario. J Archaeol Sci 16:319–329. https://doi.org/10.1016/0305-4403(89)90008-3
Katzenberg MA, Lovell NC (1999) Stable isotope variation in pathological bone. Int J Osteoarchaeol 9:316–324. https://doi.org/10.1002/(SICI)1099-1212(199909/10)9:5<316::AID-OA500>3.0.CO;2-D
Koch PL, Tuross N, Fogel ML (1997) The effects of sample treatment and diagenesis on the isotopic integrity of carbonate in biogenic hydroxylapatite. J Archaeol Sci 24:417–429. https://doi.org/10.1006/jasc.1996.0126
Lee-Thorp JA (2008) On isotopes and old bones. Archaeometry 50:925–950. https://doi.org/10.1111/j.1475-4754.2008.00441.x
Lightfoot E, Šlaus M, O’Connell TC (2012) Changing cultures, changing cuisines: cultural transitions and dietary change in Iron Age, Roman, and Early Medieval Croatia. Am J Phys Anthropol 148:543–556. https://doi.org/10.1002/ajpa.22070
Mekota AM, Grupe G, Ufer S, Cuntz U (2006) Serial analysis of stable nitrogen and carbon isotopes in hair: monitoring starvation and recovery phases of patients suffering from anorexia nervosa. Rapid Commun Mass Spectrom 20:1604–1610. https://doi.org/10.1002/rcm.2477
Noche-Dowdy, L. (2015) Multi-isotope analysis to reconstruct dietary and migration patterns of an Avar population from Sajópetri, Hungary, AD 568–895. Dissertation, University of South Florida Scholar Commons
O’Connell TC, Hedges REM (1999) Investigations into the effect of diet on modern human hair isotopic values. Am J Phys Anthropol 108:409–425. https://doi.org/10.1002/(SICI)1096-8644(199904)108:4<409::AID-AJPA3>3.0.CO;2-E
Privat KL, O’Connell TC, Richards MP (2002) Stable isotope analysis of human and faunal remains from the Anglo-Saxon cemetery at Berinsfield, Oxfordshire: dietary and social implications. J Archaeol Sci 29:779–790. https://doi.org/10.1006/jasc.2001.0785
Rapan Papeša A, Kenéz A, Pető Á (2015) The archaeobotanical assessment of grave samples from the Avar age cemetery of Nuštar (Eastern Croatia). Pril Inst Archeol Zagrub U 32:261–288
Reitsema LJ, Vercellotti G (2012) Stable isotope evidence for sex- and status-based variations in diet and life history at Medieval Trino Vercellese, Italy. Am J Phys Anthropol 148:589–600. https://doi.org/10.1002/ajpa.22085
Schoeninger MJ, DeNiro MJ (1984) Nitrogen and carbon isotopic composition of bone collagen from marine and terrestrial animals. Geochim Cosmochim Acta 48:625–639. https://doi.org/10.1016/0016-7037(84)90091-7
Sealy JC, van der Merwe NJ, Thorp JAL, Lanham JL (1987) Nitrogen isotopic ecology in Southern Africa: implications for environmental and dietary tracing. Geochim Cosmochim Acta 51:2707–2717. https://doi.org/10.1016/0016-7037(87)90151-7
Šlaus M, Bedić Ž, Rajić Šikanjić P, Vodanović M, Domić Kunić A (2011) Dental health at the transition from the Late Antique to the Early Medieval period on Croatia’s Eastern Adriatic coast. Int J Osteoarchaeol 21:577–590. https://doi.org/10.1002/oa.1163
Smith BH (1991) Standards of human tooth formation and dental age assessment. In: Kelley MA, Larsen CS (eds) Advances in dental anthropology. Wiley, New York
Sponheimer M, Lee-Thorp JA (1999) Oxygen isotopes in enamel carbonate and their ecological significance. J Archaeol Sci 26:723–728. https://doi.org/10.1006/jasc.1998.0388
Sponheimer M, Robinson T, Ayliffe L, Roeder B, Hammer J, Passey B, West A, Cering T, Dearing D, Ehleringer (2003) Nitrogen isotopes in mammalian herbivores: hair δ15N values from a controlled feeding study. Int J Osteoarchaeol 13:80–87
Stadler P (2008) Avar chronology revisited, and the question of ethnicity in the Avar qaganate. In: Curta F, Kovalev R (eds) The other Europe in the Middle Ages: Avars, Bulgars, Khazars, and Cumans. Brill, Leiden
Sykes NJ (2014) Beastly questions: animal answers to archaeological issues. Bloomsbury Academic, London
Szõke M (2003) The Late Avar period. In: Visy Z (ed) Hungarian archaeology at the turn of the millennium. Ministry of National Cultural Heritage, Budapest, pp 308–312
Tauber H (1981) 13C evidence for dietary habits of prehistoric man in Denmark. Nature 292:332–333. https://doi.org/10.1038/292332a0
van der Merwe NJ, Medina E (1991) The canopy effect, carbon isotope ratios and foodwebs in Amazonia. J Archaeol Sci 18:249–259. https://doi.org/10.1016/0305-4403(91)90064-V
Vida T (2003) The early and middle Avar period. In: Visy Z (ed) Hungarian archaeology at the turn of the millennium. Ministry of National Cultural Heritage, Budapest, pp 302–308
Vida T (2008) Conflict and coexistence: the local population of the Carpathian Basin under Avar rule (sixth to seventh century). In: Curta F, Kovalev R (eds) The other Europe in the Middle Ages: Avars, Bulgars, Khazars, and Cumans. Brill, Leiden, pp 13–46
Waters-Rist AL, Katzenberg MA (2010) The effect of growth on stable nitrogen isotope ratios in subadult bone collagen. Int J Osteoarchaeol 20:172–191. https://doi.org/10.1002/oa.1017
White CD, Spence MW, Le Q, Stuart-Williams H, Schwarcz HP (1998) Oxygen isotopes and the identification of geographical origins: the valley of Oaxaca versus the valley of Mexico. J Archaeol Sci 25:643–655. https://doi.org/10.1006/jasc.1997.0259
Wiessner PW, Schiefenhövel W (1996) Food and the status quest: an interdisciplinary perspective. Berghahn Books, Oxford, pp 1–2
Young BK (1978) Funeral archaeology and Avar culture: old excavations yield serial data. J Field Archaeol 5:471–477. https://doi.org/10.2307/529498
Acknowledgements
The authors wish to thank Catherine Kneale and James Rolfe (University of Cambridge) for their help with mass spectrometry.
Author information
Authors and Affiliations
Corresponding author
Electronic supplementary material
Online Resource 1
Microsoft Excel spreadsheet showing relevant archeological information, plus detailed Human bone and dentine collagen δ13C and δ15N, and enamel δ18O and δ13C values on Sheet 1. Faunal bone and dentine collagen δ13C and δ15N values are included on Sheet 2. (XLSX 22 kb)
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.
About this article

Cite this article
Vidal-Ronchas, R., Rajić Šikanjić, P., Premužić, Z. et al. Diet, sex, and social status in the Late Avar period: stable isotope investigations at Nuštar cemetery, Croatia. Archaeol Anthropol Sci 11, 1727–1737 (2019). https://doi.org/10.1007/s12520-018-0628-4
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12520-018-0628-4