
Abstract
Preserving animal breeds is key to a country’s economy, environment and culture. The present study collects all the available information on the Massese sheep breed and analyzes their demo-genetic trends in Pisa since 1976. Data were provided by Asso.Na.Pa. Descriptive statistics were analysed using Jmp software. The pedigree parameters of a total of 10,393 animals (whole population-WP) were analyzed using CFC, ENDOG and Pedigree viewer software. Animals born in the last fifteen years represented the Reference Population (RP), while animals with one or two unknown parents constituted the Base Population (BP). Demographic results showed an inconstant trend of offspring registration in the Herd book with two periods of a dramatic decline in lamb registrations and a substantially short productivity with a mean number of registered newborns of 1.5 and 18.2 for ewes and rams, respectively. A total of 6456 animals represented the RP, of which only 1059 animals (16.4%) had both known parents. The BP consisted of 7156 animals, thus highlighting the lack of sire and dam registration which prevented an accurate evaluation of the genetic parameters. The average inbreeding coefficient in the WP was 0.78%, and 0.046% in the RP. The number of inbreds was 575 in the WP and 17 in the RP. The present study represents a tool for breeders and technicians aimed at conserving and exploiting the genetic resources of Massese sheep through efficient breeding plans.
Similar content being viewed by others

1 Introduction
A global decline in sheep farming has been reported, especially in Europe following the abandonment of farmlands (Morris 2017). The preservation of local breeds is fundamental for safeguarding animal biodiversity and sustainability. In fact, sheep farming is mainly based on extensive systems that are often carried out on marginal land. For this reason, this activity represents a tool to safeguard biodiversity, to maintain soil functionality, to preserve water quality, and to sustain the social community. An extensive sheep farming system also minimizes the competition with human nutrition and facilitates animals behavioral patterns that are essential for maintaining a high level of animal health and welfare (Braghieri et al. 2015). In addition, safeguarding genetic resources provides an opportunity to combat new diseases and deal with environmental changes (Kurar et al. 2011).
The Massese sheep is an autochthonous breed from the Forno valley, in the Apuan Alps (province of Massa Carrara) which is mostly bred in Tuscany for its good milk production, as well as satisfactory meat production. Massese sheep are reared in mountainous and hilly areas and exploit natural resources. The breeding system is semi extensive: the animals are fed on pasture during the day and in the evening they are kept in barns, where they receive food supplements, are milked and checked regarding their health status (Parrini et al. 2021). In the past, the Massese was one of the major Italian sheep breeds reared for milk purposes (Pugliese et al. 1999). Today, thanks to its valuable characteristics, it is diffuse throughout Tuscany and also in other regions of Central Italy (Liguria, Emilia-Romagna and Lazio), however, the number of animals has contracted in recent years. At the end of the last century, there was a total population of 180,000, decreasing to 55,000 in 2006 and progressively declining so that, in 2011, the regional government in Tuscany reported it as being threatened with the risk of extinction (Regione Toscana 2020, PSR 2014–2020). Data from December 2022 from the National Data Base (BDN), report 17,976 animals, of which 3403 were reared in the province of Pisa (Sistema informatico Veterinario 2023).
The Massese sheep has the typical characteristics of the "dairy breed": light head, elongated and thin neck, long and straight back-lumbar line, wide chest with high thoracic capacity, wide rump, and slightly sloping back. The limbs are strong, guaranteeing easy grazing activity, and the hind legs are well spaced to ensure the development of the mammary gland. The udder is generally well developed however, a great variability is still reported because selection has not focused on nipple shape and size (Verità and Martini 2006). The Massese sheep has its first lamb at about 15–18 months, and is characterised by good fertility and prolificacy. Thanks to these characteristics and the fact that the females show oestrous cycles nearly all year round, breeders try for three births every two years. This is different from the usual sheep breeding system in the Mediterranean area which involves one lambing per year starting in late spring or in early autumn (Todaro et al. 2015). Deliveries generally take place in late summer or autumn. The birth season lasts until May–June, however, it is frequently interrupted after a few months by another pregnancy (Pugliese et al. 1999). In fact, 60% of ewes lamb three times in two years, and the remaining ones have a one-year lambing interval, which is why the lactation length is very variable (Moioli and Pilla 1994).
The milking procedures are mostly carried out mechanically often without a proper pre-milking area, and hand milking is still carried out on in small farms. Generally, farms do not have a lambing area, but ewes are kept with the offspring after birth to improve mother-progeny bonding (Gastaldo et al. 2015). The Massese breed has a notable aptitude for milk production. In 1971 the Italian Sheep and Goat Breeders Association (Asso.Na.Pa., Rome, Italy) estimated a milk production of 107 kg for primiparous sheep and 128 kg for adult sheep in about 236 day of lactation. Casoli et al. (1989) reported 216 kg of milk for the first lactation and 268 kg in the third, whereas Pugliese et al. (1999) described a milk production of 200–300 kg of milk per lactation. The influence of the seasonality of milk production and the variability of the duration of lactation can be a problem for cheese production (Pugliese et al. 1999; Todaro et al. 2015). Massese milk in fact is successfully used for the production of cheese and this cheesemaking aptitude has been widely investigated (Acciaioli et al. 2000; Martini et al. 2008a, b). In addition, the “Pecorino della Montagna Pistoiese” made only from the milk of Massese sheep reared in the mountains near Pistoia is part of the Slow Food movement. The quality of milk has been widely studied in terms of its chemical and nutritional properties. Diet and breed are the most important factors that influence milk fat (Conte et al. 2022). The fatty acid composition of Massese milk was found to be richer in short-chain fatty acids and lower in oleic and linoleic acid content compared to Sarda milk (Secchiari et al. 2001). Massese breed is able to take advantage of a forage-rich diet with an increase of some milk mono and polyunsaturated fatty acid (trans11-C18:1, total CLA, EPA and DHA), useful for human health (Martini and al. 2010). Several studies have focused on the macrostructure of the fat globules in Massese milk. One study on the quality of milk from the main Italian sheep breeds showed that Sarda and Massese presented a higher amount of small milk fat globules than other breeds, and more globules per milliliter of milk (Martini et al. 2016).
The meat production of the Massese breed consists of three types of lamb slaughtered in early autumn, late autumn, and late spring (Acciaioli et al. 2011). This is due to its specific reproductive pattern which enables three lambings in two years (Todaro et al. 2015). Lambs are often slaughtered after a suckling period of 30 days (Parrini et al. 2021) and a live weight of between 11 and 14 kg (Serra et al. 2009). Suckling lamb meat is characterised by a low lipid content and specific fatty acid composition. The feeding system has an influence on the meat composition of lambs. Weaned lambs fed on concentrate were shown to have a higher intramuscular fat content than animals fed on pasture (Parrini et al. 2021). A study on the differences in meat quality in Massese lamb meat in three farming systems (Parrini et al. 2021), revealed differences in fatty acid composition in lambs reared in barns, at pasture and in a mixed technique reared for up to 69 days. The highest oleic acid and PUFA n3 contents were identified in lambs reared at pasture, while no differences between rearing techniques were found for C18:2 n-6 and C18:3 n-6 C20:4 n-6 acids and PUFA n6. A study by Parrini et al. (2021) showed that lambs reared at pasture after the age of 30 days showed a lower growth rate, less carcass fat and more chewable meat texture compared with semi-free range lambs and those reared indoors.
The Herd Book of Massese breed was established in the last century and is managed by Asso.Na.Pa. The aim was to direct the genetic selection to the improvement of milk production, without neglecting the characteristics of robustness and rusticity. Knowledge of the genetic variability is essential to preserve and improve biodiversity. The first step in any safeguarding biodiversity program and in genetic improvement strategies, is to understand the demographic and genealogical structure. The study of the genetic variability of a population using pedigree data is still a common method used for both livestock (Figueredo et al. 2019; Hashemi and Hossein-Zadeh 2020; Bramante et al. 2022) and pets (Cecchi et al. 2016a; Ciccarelli et al. 2021; Michels et al. 2022), despite the widespread use of modern molecular analyses. In fact, the Massese breed was genetically characterized, together with other Italian sheep breeds, with Short Tandem Repeat (STR) markers (Bozzi et al. 2009) or high-density Single Nucleotide Polymorphism (SNP) arrays (Pilla et al. 2011).
The aim of this study was to collect information on the Massese sheep and to analyze the demo-genetic situation in the province of Pisa to develop an effective recovery and valorization plan.
2 Materials and methods
2.1 Sample collection
The study analysed the genealogical data of Massese sheep reared in the province of Pisa from 1976 to 2021 from 26 registered flocks provided by Asso.Na.Pa.. The complete genealogical sampling represented the whole Population—(WP) while data referred to 2007–2021 represented the reference population (RP). The base population (BP) represented animals with one or two unknown parents (absolute founders = ft).
2.2 Statistical analysis
Descriptive analysis was performed with JMP software (SAS-2002). Pedigree Viewer software was used for the genealogical data (Kinghorn 1994) to detect errors in the data files and to show the full pedigree structure of the population. The other analyses were carried out with ENDOG v. 4.8 (Gutiérrez and Goyache 2005) and with CFC v.1.0 (Sargolzaei et al. 2006). The analysis of the pedigree data for 2007 to 2021 was carried out only for farms that registered over 100 animals. Several parameters were computed:
-
Genetic importance of the herds. ENDOG computes the effective number (herds with a number of animals greater than 6) and the genetic importance of herds in a population (Vassallo et al. 1986). The herds are classified as: (i) nucleus herds, if breeders use only their own males, and never purchase males but sell their own males; (ii) multiplier herds, when breeders use purchased males and/or own males and also sell males and (iii) commercial herds if they use purchased males but never sell their own males.
-
Pedigree completeness. The percentage completeness of the pedigree was computed. This was also assessed by calculating the mean maximum number of generations, the mean complete number, and the number of equivalent generations traced (Gutiérrez and Goyache 2005). The number of discrete generation equivalent was performed because it is considered as another way to describe pedigree information (Siderits et al. 2013).
-
The number of inbred animals, the average inbreeding coefficient (FPED) and the average numerator relationship coefficient (ANR) in the WP, RP and for birth year, sex and flock were performed using CFC. On the other hand the FPED coefficient, number of inbred animals and average inbreeding coefficient for each traced generation was performed using ENDOG software. FPED coefficient was calculated by the tabular method described by Meuwissen and Luo (1992). The distribution of the inbreeding level in the whole population was analysed, and eight different class levels of inbreeding were considered: 0 < F ≤ 0.05; 0.05 < F ≤ 0.10; 0.10 < F ≤ 0.15; 0.15 < F ≤ 0.20; 0.20 < F ≤ 0.25; 0.25 < F ≤ 0.30; 0.30 < F ≤ 0.35; 0.35 < F ≤ 0.40 (Sargolzaei et al. 2006).
-
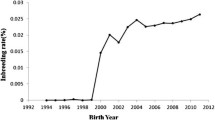
The rate of inbreeding (∆F) was calculated to follow the change in inbreeding over time and was expressed per unit of time (generations, years) (Gutiérrez and Goyache 2005). In the current study, the rate of inbreeding was calculated per generation for the whole population and for the reference population. Individuals who were present in more than one generation were included in all those generations in which they were present.
-
The effective population size (Ne) was calculated using ∆F as Ne = 1/(2∆F) (Gutiérrez and Goyache 2005). In small populations with shallow pedigrees irrespectively of how Ne is computed, it does not fit real populations well thus leading to an overestimation of the actual effective population size. To better characterize this, ENDOG gives three additional values of Ne by computing the regression coefficient (b) of the individual FPED over (i) the number of full generations traced; (ii) the maximum number of generations traced, and (iii) the number of equivalent complete generations, and considering the corresponding regression coefficient as the increase in FPED between two generations, and consequently Ne = 1/(2b) (Gutiérrez and Goyache 2005).
3 Results and discussion
3.1 Demographic data
A total of 9041 lambs were born and registered during the period, comprising 254 males and 8787 females. The number of offspring showed an inconstant trend (Fig. 1). Two periods of dramatic decline were found, the first between 2006 and 2008, the second between 2019 and 2021. From 2000 to 2004, the regional policies of Tuscany promoted an increase in Massese sheep farming. After that, the sector suffered a sharp decline; subsequently, new awareness policies led to a further increase (Belletti et al. 2019). The causes of the crisis are due to generational change, the increase in the number of predators which led to serious animal losses (Dondina et al. 2015; Russo et al. 2020), and the continuous adaptations to health and hygiene regulations which require significant financial resources (AAVV 2009). Furthermore, the second decline may have been affected by the introduction of Legislative Decree n. 52/2018 on the regulation of animal reproduction and which may have led to application problems (Santus 2018).

Offspring/year
Figure 2 shows the number of registered offspring for every ram. Rams were clearly underemployed with evident negative consequences in terms of genetic value assessment. In fact, the mean number of lambs registered per ram was 18 ± 35.7 with a maximum of 254 in only one case. A similar situation appeared for the length of the productive career of the ewes (Fig. 3). The mean registered offspring per ewe was 1.5 ± 0.92 however 1726 ewes (66%) gave birth only once. A career length of nine deliveries was registered for only one ewe. These results do not reflect the common management of sheep which reports four to five lactactions (Giannone 2016).

Offspring distribution/ram

Offspring distribution/ewe
A total of 26 herds were counted for this population but ENDOG computed 19 effective herds, of these only three (15.8%) were categorized as multipliers, while 16 herds (84.2%) were categorized as commercial. Fifteen herds had over 100 registered animals from 2007 to 2021. For genetic improvements, it is crucial to use rams derived from the Genetic Center of Asciano (Siena, Italy). The present study showed that only five herds used selected rams (a total of 13 sires) that mated with a limited number of females. In only two flocks, the same ram was used 19 and 129 times, reflecting the same low productive exploitation occurring for the females.
3.2 Genealogical data
Table 1 summarizes the main genealogical parameters computed using CFC software. WP resulted in 10,393 records (368 males and 10,025 females). BP included 7156 animals (68.85% of WP): 6338 with two unknown parents, 796 only with the mother and 22 with only the father. As is known in sheep, the correct attribution of parentage is difficult especially due to the simultaneous presence of more than one ram in the flock (Cortellari et al. 2022). A total of 6456 animals were registered in the RP (201 males and 6255 females); 1059 of these (16.4%) had both known parents and only 17 (0.26%) were inbred, since for many animals the depth of the pedigree reached only the first generation. Three hundred and seventy groups of full sibs were detected in the WP, and the average number of full sibs per family was 2.15 (range 2–5).
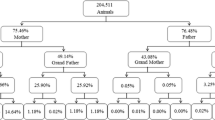
Figure 4 shows the whole genealogy of the population stratified into 12 traced generations, where the first line represents the BP, and the last line reports the animals of the twelfth generation. The details of the pedigree quality of the Massese sheep for the WP and RP are reported in Fig. 4. The percentage of completeness of the pedigree for the WP (Fig. 4a) was computed from 31.2% to 38.7% for the parental generation; from 12.0% to 14.8% for the grandparent generation and from 4.5% to 7.4% for the third generation of great-grandparents. Data regarding the RP (Fig. 4b) were lower: from 16.4% to 28.7% for the parental generation; from 3.4% to 7.0% for the grandparent generation, and from 0.2% to 1.0% for the third generation of great-grandparents. The pedigree completeness level was similar both in the dam and sire pathways. Pedigree completeness that has a strong influence on the validity of the estimated values, determines how accurately the inbreeding coefficient can be estimated.

Pedigree completeness level a in the entire set of pedigree data files (GS and GD, grandparents; GGS and GGD, great grandparents) and b in the reference population
The mean maximum generations and the mean complete generations were 0.99 and 0.40, with an increase in inbreeding equal to 0.76% and 2.51% respectively, while the number of equivalent generations traced was 0.59 with an increase in inbreeding equal to 1.80%. The discrete generation equivalent in this population was 0.22%, indicating that the completeness of the pedigree was at an unsatisfactory level. The number of ancestors that give rise to the RP was 5193, 2168 (42%) of which explained 50% of the genetic variability of the RP, while the effective number of ancestors (fa) was 1948, confirming the low productive exploitation of rams as reported in the “demographic” section. The high number of founders confirmed the lack of complete genealogical information, preventing a realistic analysis of the relationships between the animals and their inbreeding values. It also does not provide a realistic situation of those animals that mainly contributed to the genetic variability of the RP.
3.3 Inbreeding coefficient
The average inbreeding coefficient in the WP was 0.78% and the average numerator relationship was 0.21%. No differences were observed between males and females in the average inbreeding coefficient (0.54% vs 0.79%, respectively). It is not possible to compare our values with those reported in the literature on other breeds since the average FPED in a population varies in relation to the completeness of the data and the pedigree depth. The number of inbreds was 575 (5.53%) with an average inbreeding coefficient of 14.45% (Fmin = 0.2% and Fmax = 37.5%) with no differences between inbred males and inbred females (13.20% vs 14.20%, respectively). Considering the 575 inbred animals (Table 2), 133 had an inbreeding coefficient lower than 0.05 but 196 animals showed values higher than 0.20. The highest values came from four matings between full sibs, 65 matings between half-sibs and seven between parent-offspring.
In the RP population, the average numerator relationship was 0.058% and the average inbreeding coefficient was 0.046% (Fmin 0%—Fmax 25%). Of the 17 inbred animals, 13 had a high coefficient (F > 0.125) with the risk of inbreeding depression. As noted above, this average inbreeding value is not realistic due to the lack of known genealogies and did not enable the real genetic variability of the breed to be quantified. In fact, in a comparison with Italian sheep breeds, Ciani et al. (2014) reported that Massese had the lowest heterozygosity, resulting in one of the most genetically homogeneous breeds.
The effective population size (Ne) of the RP, computed via the individual increase in inbreeding, was 77, while the Ne, computed via the regression on equivalent generations and that obtained from Log regression on equivalent generations, were 147.3 and 157.3, respectively. These values were higher than 50, thus corresponding to the minimum value suggested by the Food and Agriculture Organization (FAO 1998) to maintain fitness in a breed.
Table 3 reports the WP statistics on the average inbreeding for each generation traced. The highest proportion of inbred animals, with an average inbreeding of 7.42%, was observed in the fifth traced generation. In the last three generations, the data were not reliable. Results by year (data not reported) showed that the animals with both parents known were high up to 2003 (range 97–100%), and then dropped drastically with the total absence of genealogical data in 2007. Registrations then rose slightly reaching a maximum of 45% in 2017. From 2018 registrations increased again. Within the herds considered (data not reported), the limited completeness of pedigree data was confirmed. In fact, in some cases, pedigree data were completely missing and in others, the percentage of animals with both parents known varied from 1 to 39%.
Genealogical information is very important in order to create better mating schemes for genetic improvements, and the use of sires with the lowest ANR coefficient with dams can be an effective strategy to control inbreeding (Gholizadeh and Ghafouri-Kesbi 2016). Excessive inbreeding in a population can lead to a loss of genetic variability, highlighting deleterious recessive alleles and the appearance of an inbreeding depression. The latter affects the productive parameters (Gholizadeh and Ghafouri-Kesbi 2016), morphological parameters (Cecchi et al. 2018) and especially the reproductive parameters such as fitness (Cecchi et al. 2016b), fertility (Gonzalez-Recio et al. 2007) prolificacy and fecundity (Selvaggi et al. 2010) in every species.
Unfortunately, the genealogical data of the Massese sheep reared in the province of Pisa are very poor. To overcome these deficiencies, the only solution is thus to compute the inbreeding coefficient using genomic inbreeding. Different metrics are available to obtain the estimates of genomic inbreeding. These include the runs of the homozygosity (FROH), which is an increasingly important tool to exploit the inbreeding value using traditional pedigree information (FPED). It is also a possible solution when there is a lack of genealogy (Curik et al. 2014).
Genomic data can be used to study inbreeding depression (Bjelland et al. 2013; Saura et al. 2015; Ferenčaković et al. 2017; Howard et al. 2017) without the need for a parentage analysis over many generations, because the two inbreeding coefficients are correlated (Cortellari et al. 2022). In fact, many studies have investigated the accuracy of this estimation (Caballero et al. 2021); Cortellari et al. (2022) showed that a correlation between FPED and FROH exists in Massese sheep with a value of 0.318 (P < 0.0001). In other breeds, where the pedigrees were more complete, the correlation between the two values was higher. However, despite the availability of genomic tools, genealogical data remain the main source of information for monitoring genetic variability in many populations of livestock species (Kasap et al. 2021).
4 Conclusions
The study shows a serious decline in Massese sheep farms enrolled in the Herd book in the province of Pisa. The need for genetic improvement together with breed maintenance requires a plan of action. In the present study, the lack of genealogical information prevented an analysis of the genetic variability of the population and the estimation of important genetic parameters. In fact, it was possible to calculate the inbreeding coefficient only for a few animals with known parents and grandparents. Pedigree information is still the simplest and least expensive method used to compute inbreeding and relationship coefficients, and it is the starting point for the correct genetic management of breeding, which is essential for the preservation of a breed.
References
AAVV (2009) La gestione sostenibile dei sistemi pascolivi italiani. Rete Interregionale per la ricerca agraria, forestale, acquacoltura e pesca, Progetto MASO-GIS. Quaderni della Regione Piemonte.
Acciaioli A, Franci O, Pugliese C, Parisi G, Lucifero M (2000) Variation and repeatability of a.m. and p.m. records for milk yield and composition in Massese ewes. Zoot Nutr Anim 26:253–262
Acciaioli A, Giustini L, Pianaccioli L, Malvezzi G (2011) Survey of feeding techniques in the sheep farms of Pistoia Apennines and milk quality. Sci Tecn LC 62(6):407–412
Belletti G, Conte G, Marescotti A, Mele M, Scaramuzzi S, Serra A (2019) Tutela e valorizzazione dell’agrobiodiversità vegetale e animale in Toscana: analisi e indicazioni di policy. Regione Toscana.
Bjelland DW, Weigel KA, Vukasinovic N, Nkrumah JD (2013) Evaluation of inbreeding depression in Holstein cattle using whole-genome SNP markers and alternative measures of genomic inbreeding. J Dairy Sci 96:4697–4706. https://doi.org/10.3168/jds.2012-6435
Bozzi R, Deglinnocenti P, Diaz PR, Nardi L, Crovetti A, Sargentini C, Giorgetti A (2009) Genetic characterization and breed assignment in five Italian sheep breeds using microsatellite markers. Small Ruminant Res 85(1):50–57
Braghieri A, Pacelli C, Bragaglio A, Sabia E, Napolitano F (2015) The Hidden Costs of Livestock Environmental Sustainability: The Case of Podolian Cattle. In: Di Trana et al. The Sustainability of Agro-Food and Natural Resource Systems in the Mediterranean Basin. Springer - Publisher Connector https://files.core.ac.uk/pdf/2612/81562724.pdf
Bramante G, Pieragostini E, Ciani E (2022) Genetic Variability within the Murgese Horse Breed Inferred from Genealogical Data and Morphometric Measurements. Diversity 14(6):422. https://doi.org/10.3390/d14060422
Caballero A, Villanueva B, Druet T (2021) On the estimation of inbreeding depression using different measures of inbreeding from molecular markers. Evol Appl 14:416–428
Casoli C, Duranti E, Morbidini L, Panella F, Vizioli V (1989) Quantitative and compositional variations of Massese sheep milk by parity and stage of lactation. Small Ruminant Res 2(1):47–62
Cecchi F, Giacalone G, Paci G (2016a) Inbreeding depression in Lizard canaries breed estimated by pedigree analysis. Czech J Anim Sci 61(1):15–21. https://doi.org/10.17221/8664-CJAS
Cecchi F, Paci G, Spaterna A, Ragatzu M, Ciampolini R (2016b) Demographic approach on the study of genetic parameters in the dog Braque Français type Pyrénées Italian population. Ital J Anim Sci 15(1):1–7. https://doi.org/10.1080/1828051X.2015.1128689.ISSN1828-051X
Cecchi F, Carlini G, Giuliotti L, Russo C (2018) Inbreeding may affect phenotypic traits in an Italian Population of Basset Hound dog. Atti Accad Naz Lincei Cl Sci Fi Mat Nat 29:165–170
Ciani E, Crepaldi P, Nicoloso L, Lasagna E, Sarti FM, Moioli B et al (2014) Genomewide analysis of Italian sheep diversity reveals a strong geographic pattern and cryptic relationships between breeds. Anim Genet 45:256–266
Ciccarelli J, Macchioni F, Cecchi F (2021) A genealogical survey on the main bloodline of the Australian Cattle Dog in Italy. Rend Fis Acc Lincei 32:357–364
Conte G, Palombo V, Serra A, Correddu F, D’Andrea M, Macciotta NPP, Mele M (2022) Study of the fatty acid profile of milk in different sheep breeds: evaluation by Multivariate Factorial Analysis. Animals 12(6):722
Cortellari M, Negro A, Bionda A, Grande S, Cesarani A, Carta A, Macciotta N, Biffani S, Crepaldi P (2022) Using Pedigree and Genomic Data toward Better Management of Inbreeding in Italian Dairy Sheep and Goat Breeds. Animals 12:2828. https://doi.org/10.3390/ani12202828
Curik I, Ferenčaković M, Sölkner J (2014) Inbreeding and runs of homozygosity: A possible solution to an old problem. Livestock Sci. https://doi.org/10.1016/j.livsci.2014.05.034
Dondina O, Meriggi A, Dagradi V, Perversi M, Milanesi P (2015) Wolf predation on livestock in an area of northern Italy and prediction of damage risk. Ethol Ecol Evol 2(72):200–219. https://doi.org/10.1080/03949370.2014.916352
FAO (1998) Secondary guidelines for development of national farm animal genetic resources management plans. Food and Agriculture Organization, Page 63 in Management of Small Populations at Risk. United Nations Rome, Italy.
Ferenčaković M, Sölkner J, Kapš M, Curik I (2017) Genome-wide mapping and estimation of inbreeding depression of semen quality traits in a cattle population. J Dairy Sci. https://doi.org/10.3168/jds.2016-12164
Figueredo JS, Cruz JF, Sousa LS, Teixeira Neto MR, Carneiro PLS, Brito ND, Pinheiro RGS, Lacerda KSO, Mottin VD (2019) Genetic diversity and population structure estimation of Brazilian Somali sheep from pedigree data. Small Rumin Res 179:64–69
Gastaldo A, Benvenuti MN, Paganelli O, Rossi P, Giuliotti L (2015) Animal welfare assessment in sheep farms before the application of the Measure 215 “Animal welfare payments” in Tuscany. Large Anim Rev 21(3):137–141
Gholizadeh M, Ghafouri-Kesbi F (2016) Inbreeding depression in growth traits of Baluchi sheep. Small Ruminant Res. https://doi.org/10.1016/j.smallrumres.2016.09.012
Giannone M (2016) Gli ovini da latte. Edagricole, Bologna. https://doi.org/10.1080/1828051X.2020.1869599
Gonzalez-Recio O, Lopez de Maturana E, Gutierrez JP (2007) Inbreeding depression on female fertility and calving ease in Spanish dairy cattle. J Dairy Sci 90:5744–5752
Gutiérrez JP, Goyache F (2005) A note on ENDOG: a computer program for analysing pedigree information. J Anim Breed Genet 122:172–176
Hashemi M, Hossein-Zadeh NG (2020) Population genetic structure analysis of Shall sheep using pedigree information and effect of inbreeding on growth traits. Italian J Anim Sci 19(1):1195–1203
Howard JT, Pryce JE, Baes C, Maltecca C (2017) Invited review: Inbreeding in the genomics era: Inbreeding, inbreeding depression, and management of genomic variability. J Dairy Sci. https://doi.org/10.3168/jds.2017-12787
Kasap A, Ramljak J, Špehar M (2021) Estimation of Population-Specific Genetic Parameters Important for Long-Term Optimum Contribution Selection-Case Study on a Dairy Istrian Sheep Breed. Animals 11:2356
Kinghorn BP, Kinghorn S (1994) Pedigree Viewer-a graphical utility for browsing pedigreed data sets. In 5th World Congress on Genetics Applied to Livestock Production, 22: 85–86. Canada: University of Guelph.
Kurar E, Ozsensoy Y, Dogan M, Bulut Z, Altunok V, Isik A., Camlidag A, Nizamlioglu M (2011) Autosomal and mitochondrial genetic diversity of Turkish native cattle breeds. In RBI 8th Global Conference on the Conservation of Animal Genetic Resources, Tekirdağ, Turkey. Ekim, October 4–11 pp. 185–191
Martini SF, Altomonte I (2016) The macrostructure of milk lipids: the fat globules. Crit Rev Food Sci Nutr 56(7):1209–1221
Martini LGB, Salari F (2010) Effect of forage: concentrate ratio on the quality of ewe’s milk, especially on milk fat globules characteristics and fatty acids composition. J Dairy Res 77(2):239–244
Martini M, Mele M, Scolozzi C, Salari F (2008a) Cheese making aptitude and the chemical and nutritional characteristics of milk from Massese ewes. Ital J Anim Sci 7(4):419–437
Martini M, Scolozzi C, Cecchi F, Mele M, Salari F (2008b) Relationship between morphometric characteristics of milk fat globules and the cheese making aptitude of sheep’s milk. Small Rum Res 74(1–3):194–201
Meuwissen T, Luo Computing Z (1992) Inbreeding coefficients in large populations. Genet Sel Evol 24:305. https://doi.org/10.1186/1297-9686-24-4-305
Michels PW, Distl O (2022) Genetic Diversity and Trends of Ancestral and New Inbreeding in Deutsch Drahthaar Assessed by Pedigree Data. Animals 12:929. https://doi.org/10.3390/ani12070929
Moioli BM, Pilla AM (1994) Genetic Evaluation of Dairy Sheep with an Animal Model for Annual or Partial Lactation Production. J Dairy Sci 77(2):609–615
Morris ST (2017) Advances in sheep welfare. Food Sci Technol Nutr. https://doi.org/10.1016/B978-0-08-100718-1.00002-9
Parrini S, Sirtori F, Acciaioli A, Becciolini V, Crovetti A, Bonelli A, Franci O, Bozzi R (2021) Effect of farming system on meat traits of native Massese suckling lamb. Ital J Anim Sci 20(1):71–83
Pilla F, Ciani E, Maria Sarti F, Moioli B, Carta A, Matassino D, Crepaldi P, Ciampolini R, Marletta D, Acutis PL, Macciotta NPP, Ajmone Marsan P, Portolano B (2011) Genetic diversity and structure of Italian sheep breeds inferred by genome-wide SNP analysis: overview of the “Italian Project for Sheep Biodiversity.” Ital J Anim Sci 10:69–70
Pugliese C, Acciaioli A, Rapaccini S, Parisi G, Franci F (1999) Evolution of chemical composition, somatic cell count and renneting properties of the milk of Massese ewes. Small Rum Res. https://doi.org/10.1016/S0921-4488(99)00070-X
Regione toscana. Piano di sviluppo rurale Fondo europeo agricolo di sviluppo rurale (Feasr), 2014–2020. https://www.regione.toscana.it/psr-2014-2020/cos-e
Russo C, Cecchi F, Accorsi PA, Scampuddu N, Benvenuti MN, Giuliotti L (2020) Investigation on sheep farm characteristics, wolf predation and animal welfare in the Grosseto province (Italy). Atti Soc Tosc Sci Nat Mem Serie B. https://doi.org/10.2424/ASTSN.M.2020.08
Santus E. (2018) Riproduzione animale: nuovi decreti e futuro da costruire. Razza Bruna 3:6–8. http://www.anarb.it/wp-content/uploads/2019/03/RIPRODUZIONE_ANIMALE.pdf
Sargolzaei M, Iwaisaki H, Colleau JJ (2006) CFC (Contribution, Inbreeding (F), Coancestry), Release 1.0. A software package for pedigree analysis and monitoring genetic diversity. Proc 8th World Congress on Genetics Applied on Livestock Production. Belo Horizonte, Brazil, CD-ROM comm. N 27–28.
SAS Institute Inc. (2002) JMP-5 Statistical Software, Version 5. Cary, USA.
Saura M, Fernández A, Varona L, Fernández AI, de Cara MAR, Barragán C, Villanueva B (2015) Detecting inbreeding depression for reproductive traits in Iberian pigs using genome-wide data. Genet Sel Evol 47:1. https://doi.org/10.1186/s12711-014-0081-5
Secchiari P, Antongiovanni M, Mele M, Buccioni A, Serra A, Ferruzzi G (2001) Effect of dietary fat on milk fat acidic composition of ewes and cows. Liv Prod Sci 70:175
Selvaggi M, Dario C, Peretti V, Ciotola F, Carnicella D, Dario M (2010) Inbreeding depression in Leccese sheep. Small Rumin Res 89:42–46
Serra A, Mele M, La Comba F, Conte G, Buccioni A, SecchiariP. (2009) Conjugated Linoleic Acid (CLA) content of meat from three muscles of Massese suckling lambs slaughtered at different weights. Meat Sci. 81(2), 396–404Siderits M, Baumung R, Fuerst-Waltl B (2013) Pedigree analysis in the German Paint Horse: genetic variability and the influence of pedigree quality. Liv Sci 151:152–157
Siderits M, Baumung R, Fuerst-Waltl B (2013) Pedigree analysis in the German Paint Horse: genetic variability and the influence of pedigree quality. Liv Sci 151:152–157. https://doi.org/10.1016/j.livsci.2012.10.018
Sistema Informatico Veterinario (2023) https://www.vetinfo.it/j6_statistiche/index.html#/ Accessed on 15 March 2023.
Todaro M, Dattena M, Acciaioli A, Bonanno A, Bruni G, Caroprese M, Mele M, Sevi A, Trabalza Marinucci M (2015) Aseasonal sheep and goat milk production in the Mediterranean area: Physiological and technical insights. Small Rum Res 126(1):59–66
Vassallo JM, Díaz C, García-Medina JR (1986) A note on the population structure of the Avileña breed of cattle in Spain. Livest Prod Sci 15:285–288
Verità P, Martini M (2006) Massese. In: Risorse genetiche animali autoctone della Toscana. ARSIA. Regione Toscana.
Funding
Open access funding provided by Università di Pisa within the CRUI-CARE Agreement. This work is supported by a grant from the University of Pisa (Ateneo—Dr. Lorella Giuliotti, Prof. Francesca Cecchi, and Prof. Giovanna Preziuso).
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors report that they have no conflicts of interest. The authors alone are responsible for the content and writing of this article.
Animal studies statement
All animal procedures used in this study were in compliance with the ethical and animal welfare concerns of the Committee on the Ethics of Animal Experiments of Minimally Invasive Surgery Centre and fully complied with recommendations outlined by the Italian laws.
Informed consent
All authors have approved this manuscript, agree to the order in which their names are listed, declare that they have no conflict of interest exist, and have no commercial affiliation.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Giuliotti, L., Benvenuti, M.N., Ventura, E. et al. Massese sheep characteristics and demo-genetic trends in the province of Pisa (Italy). Rend. Fis. Acc. Lincei 34, 1221–1229 (2023). https://doi.org/10.1007/s12210-023-01198-6
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12210-023-01198-6