
Abstract
Nardus stricta dominated grassland is a specific habitat occurring on the nutrient-poor soils. Its large areas were formed as a result of livestock grazing. However, landscape management underwent significant changes over the last decades including grazing cessation. This triggered successional processes leading to considerable changes in floristic composition reported from numerous European regions. We focused on this phenomenon in the Western Carpathian high mountains, where the issue was not studied sufficiently. Our research, based on pairwise comparison of 19 historical and recent phytocoenological relevés, confirmed changes here. These include (i) decrease in cover of some diagnostic species of Nardetea strictae class, especially Nardus stricta, (ii) increase in competitively strong species with their high biomass productivity, (iii) shift in floristic composition indicating conversion of Nardion strictae vegetation into other communities, especially those of Loiseleurio-Vaccinietea class, (iv) increase in Shannon-Wiener index values and (v) enrichment of originally oligotrophic grasslands with some nutrient-demanding species, mainly at lower altitudes and decrease in light-demanding species.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Low-stem grassland of Nardion strictae typically occur on the nutrient-poor soils from lowlands to alpine zone (Janišová et al. 2007; Kliment 2007a, b; Krahulec et al. 2010; Kliment and Ujházy 2014) and their formation is closely related to grazing (Galvánek and Janák 2008; Korzeniak 2016; Zarzycki et al. 2022). Its dominant species, Nardus stricta, is not favoured by grazing animals due to its hard, indigestible and bristle-like leaves, and thus has a competitive advantage (Šmarda 1963; Sebastiá 2004; Massey et al. 2007). Large areas of this vegetation in Europe were established on the sites of mountain forests and shrublands, which were eradicated by man to extend the pastures. Therefore, the timber line was shifted downward, e.g. by 200–400 m in the Alps (Erschbamer et al. 2003) and 150–300 m in the Western Carpathians (Midriak 1994).
Distribution area of Nardion strictae grassland has decreased and its structure has changed significantly in Europe during recent decades as a result of traditional management changes, mainly grazing cessation (Galvánek and Janák 2008; Kurtogullari et al. 2020). Grazing favours low subtle species, and, on the contrary, inhibits propagation of strong competitors (Puccio et al. 2007; Skarpe and Hester 2008). In addition, thick tufts of Nardus stricta along with soil compaction caused by grazing livestock inhibit successional processes (Sebastiá 2004; Parolo et al. 2011).
If this vegetation is grazed, it is considered stable and long-lasting (Velev and Apostolova 2008). After grazing cessation, it loses its competitive advantage and gradual regeneration of primary communities is triggered. The pattern of this process depends on the specific vegetation structure, grazing intensity, altitude, slope inclination and climate (Tasser and Tappeiner 2002; Vandvik and Birks 2002; Erschbamer et al. 2003; Korzeniak 2016). Nardus stricta is gradually being replaced by competitively stronger graminoids, forbs and woody plants (Erschbamer et al. 2003; Grigoriu and Alda 2004; Velev and Apostolova 2008; Korzeniak 2016).
The changes in this vegetation may take effect almost immediately (Mišić et al. 1978). However, as reported by Korzeniak (2016), the speed of those processes may be mitigated by climate. The most crucial changes in the vegetation structure occur during the first decades after grazing cessation (Velev and Apostolova 2008). Witkowska-Żuk and Ciurzycki (2000) found that 30 years of secondary succession in Nardion strictae grassland leads to the Vaccinium myrtillus dominated stands, tall grassland of Calamagrostion villosae or Pinus mugo stands. After 50–100 years from pasture abandonment, about 40% of grazing indicators disappear (Helm et al. 2006).
In Slovak Western Carpathians, the issue of Nardus stricta grassland dynamics and changes resulting from land use changes that occurred over the last decades, was studied especially in the montane belt (e.g. Šomšák and Balkovič 2002; Hrivnák and Ujházy 2005), while there is a lack of data from the higher altitudes. Therefore, the aim of this article is to identify and characterize Nardus stricta grassland changes in subalpine and alpine belts in the highest mountain ranges of the Western Carpathians, where one of the major land use change is grazing cessation linked to declaration of national parks.
Materials and methods
Study area
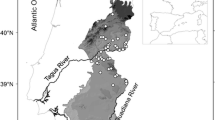
Study area is situated in the alpine landscape of Low and Western Tatras at an altitudinal range of 1545–1870 m (Fig. 1). Geological bedrock includes granitoids (Biely et al. 1992; Nemčok 1994). Relief is relatively smooth thus providing proper conditions for a well-developed subalpine belt (Midriak 1983). The Podzols and Cambisols are the most common soils (Linkeš 1967; Bublinec et al. 1994). The study area is classified as a cold climate region (Lapin et al. 2002). We use climate data from Kasper peak meteorological station located in the Western Tatras (1987 m a.s.l.) and Chopok meteorological station located in the Low Tatras (2005 m a.s.l.) from the period of 1970–2020 to specify the development of the mean annual temperature in the study area (Fig. 2). Study area has been intensively grazed since the Wallachian colonization which took place in the 16th century. Sheep grazing has prevailed over cattle grazing (Häufler 1955). Intensive grazing in the study area is evidenced by the shifting downwards of timberline by at least 50–160 m in the Low Tatras and 100–150 m in the Western Tatras (Midriak 1983). From the 17th to the 20th century, the vegetation was grazed wherever it was possible, including summit areas (Häufler 1955; Bohuš 1966). In 1971, 5,600 ha were grazed above the timberline in the Western Tatras (Bohuš 1994), which, according to Midriak (1983), represents almost 70% of the area with a slope below 35° (including Pinus mugo stands). In the Low Tatras, grazing was even more expanded (Häufler 1955; Midriak 1983). In the Low Tatras, the grazing cessation dates back to the year 1978 when it was declared a national park and to the year 1987 in the Western Tatras when it was included into Tatra National Park.

Location of study sites within the Tatra National Park and the Low Tatras National Park

Average annual temperature in the period of 1970–2020 from Kasper peak and Chopok peak meteorological station with a 5 year moving average
Data collection
Current phytosociological relevés were sampled in 2016–2021 using the Zürich-Montpellier school and its 7-degree cover/abundance scale (Braun-Blanquet 1964) on the plots of historical relevés from Horák (1970), Treskoňová (1972), Kremlová (1974), Turečková (1974), Dúbravcová (1976), Hrabovcová (1976), Pietorová (1977), Králik (1979) and Altmanová (1983). Recent relevés were published in our earlier study (Palaj and Kollár 2022). Plots were located using header data of historical relevés and, in some cases, by maps with relevé locations (Horák 1970). If the field conditions did not correspond to the header data, we selected a site at a maximum distance of 50 m from the likely centre of the historical plot. Originally, we used a dataset of 45 historical relevés, however, 26 of them could not be relocated due to insufficient information in the header data or natural conditions. Thus, only 19 historical relevés were included.
Data analysis
Collected data were stored in the TURBOVEG database (Hennekens and Schaminée 2001) and processed by JUICE software (Tichý 2002). Vascular plant names follow Marhold and Hindák (1998), syntaxonomical classification is according to Jarolímek and Šibík (2008). Frequency and median non-zero cover of species, along with number of years between sampling, altitude, aspect and slope of the plots, is appended as supplementary data (SM_1.pdf). The assessment of vegetation changes was based on a pair comparison of 19 historical and 19 current relevés. Vegetation and site conditions of the study area within both mountain ranges are similar enough (Kliment and Valachovič 2007) to be analyzed together. We used relative cover change to assess the change in species composition with the special emphasis on diagnostic species of Nardetea strictae class (Jarolímek and Šibík 2008): Agrostis capillaris, Carex pilulifera, Hypericum maculatum, Luzula luzuloides, Nardus stricta, Phleum rhaeticum, Potentilla aurea and Trommsdorffia uniflora. For calculation, we used the formula:
CCH = 100 - (CL/CH *100),
where CL represents species cover in the period in which it was lower; CH is species cover in the period in which it was higher. In the case of decline of cover, we added a minus sign to the resulting value. The overall shift in species composition was determined by the Redundancy Analysis (RDA) and we used sampling period as the only variable (historical/current relevé). Vegetation dynamics was evaluated also on the base of species cover changes for individual life forms (Raunkiaer 1934) and ecological strategies (Grime 1979) classified according to the BiolFlor database (Klotz and Kühn 2002). The differences were tested by paired t-test. Changes in site conditions were estimated by comparison of recent and historical values of Shannon-Wiener index and by species cover weighted means of Ellenberg indicator values (EIVs) for light, temperature, moisture, soil reaction and nutrients (Ellenberg et al. 1992). Differences were tested by t-test. Finally, to estimate directions of succession processes, we performed Non-metric multidimensional scaling ordination (NMDS) on log-transformed data using Bray-Curtis dissimilarity matrix. As supplementary variables we used cover of diagnostic species of other classes occurring in the study area, classified according to Jarolímek and Šibík (2008). Based on current and historical data (Horák 1970; Treskoňová 1972; Kremlová 1974; Turečková 1974; Dúbravcová 1976; Hrabovcová 1976; Pietorová 1977; Králik 1979; Altmanová 1983; Palaj and Kollár 2017, 2019), the following classes can be considered adjacent with Nardetea strictae stands in the study area: Betulo carpaticae-Alnetea viridis, Calluno-Ulicetea, Caricetea curvulae, Loiseleurio-Vaccinietea, Mulgedio-Aconitetea, Roso pendulinae-Pinetea mugo, Salicetea herbaceae and Vaccinio-Picetea. If the species was considered diagnostic for more classes, a higher fidelity was decisive. Changes in cover of diagnostic species of all classes were tested by t-test and presented by bar chart. Cover of diagnostic species of other classes was also used as a dependent variable in linear mixed-effect models (Bates et al. 2015). As fixed effects we used altitude, slope and to radians converted aspect degrees, while a random effect included time. For all ordinations we employed R package vegan (Oksanen et al. 2020; RStudio Team 2022), plots were made using ggplot2 (Wickham 2016) and ggtern (Hamilton and Ferry 2018) packages.
Results
Floristic composition significantly changed during recent decades. In most of the plots, decrease in cover of diagnostic species of Nardetea strictae class was identified. The most noticeable cover decrease was recorded for Nardus stricta, whose cover decreased by almost 60% on average, while Agrostis capillaris and Carex pilulifera disappeared entirely. Other diagnostic species remained without significant change. The shift in cover of the most common species is shown by Fig. 3.

The overall shift in species composition determined by the RDA. The ordination diagram shows only species with a frequency above 5% in both time periods. Out of the total number of 70 species, 44 are displayed. Abbrevations: Agr_rup – Agrostis rupestris, Ant_alp – Anthoxanthum alpinum, Ave_fle – Avenella flexuosa, Ave_ver – Avenula versicolor, Bis_maj – Bistorta major, Cal_vil – Calamagrostis villosa, Cal_vul – Calluna vulgaris, Cam_alp – Campanula alpina, Cam_tat – Campanula tatrae, Car_big – Carex bigelowii, Car_sem – Carex sempervirens, Des_ces – Deschampsia cespitosa, Fes_pic – Festuca picturata, Fes_sup – Festuca supina, Gen_asc – Gentiana asclepiadea, Gen_pun – Gentiana punctata, Hie_alp – Hieracium alpinum, Hie_spe – Hieracium sp., Hom_alp – Homogyne alpina, Hyp_mac – Hypericum maculatum, Jun_tri – Juncus trifidus, Jun_sib – Juniperus sibirica, Lig_mut – Ligusticum mutellina, Luz_alp – Luzula alpinopilosa ssp. obscura, Luz_luz – Luzula luzuloides, Luz_sud – Luzula sudetica, Nar_str – Nardus stricta, Oma_nor – Omalotheca norvegica, Ore_mon – Oreogeum montanum, Phl_rha – Phleum rhaeticum, Pin_mug – Pinus mugo, Pot_aur – Potentilla aurea, Pse_alb – Pseudorchis albida, Pul_sch – Pulsatilla scherfelii, Ran_pse – Ranunculus pseudomontanus, Sem_mon – Sempervivum montanum, Sen_abr – Senecio abrotanifolius, Sen_inc – Senecio incanus ssp. carniolicus, Sol_car – Soldanella carpatica, Sol_vir – Solidago virgaurea ssp. minuta, Tro_uni – Trommsdorffia uniflora, Vac_gau – Vaccinium gaultherioides, Vac_myr – Vaccinium myrtillus, Vac_vit – Vaccinium vitis-idea
Current state promotes competitively strong species with their high biomass productivity. Their coverage increased from 47 to 50%; t = -2.74; p-value = 0.010), while cover of R-selected species has decreased from 8.5 to 6.5% (Fig. 4). However, this change was not significant (t = 1.44; p-value = 0.167). Stress tolerators remain unchanged. These changes are reflected also in vegetation physiognomy, when succession processes are leading to the stands with taller plant species Expansion of dwarf shrubs is noticeable – their frequency increased by 12% and average cover increased from 8.5 to 27.5% (t = -4.36; p-value < 0.001) (Fig. 5). The most expanding species were Vaccinium myrtillus, less also V. gaultherioides, V. vitis-idea and Calluna vulgaris. These are followed by some phanerophytes, such as Juniperus sibirica, Picea abies and Pinus mugo partly forming a low shrub layer. Increase in dwarf shrub cover more-less corresponds to hemicryptophyte decreases (t = -4.37; p-value < 0.001). The hemicryptophyte decrease, which is related mainly to Nardus stricta, is partly compensated by expansion of some competitively strong graminoids, such as Calamagrostis villosa, Carex sempervirens, Deschampsia cespitosa, Festuca picturata, Juncus trifidus and Luzula alpinopilosa ssp. obscura.

Relative proportion (%) of C-, S- and R-selection for 19 historical and 19 recent relevés. The changes are characterized by a statistically significant increase in C-selected species (t = -2.60; p-value = 0.018)

Comparison of historical and recent average abundance of hemicryptophytes (t = 4.37; p-value < 0.001) and chamaephytes (t = -4.36; p-value < 0.001). Changes in other life forms were not statistically significant
Values of Shannon-Wiener index increased significantly from 1.7 to 2.1 and species richness of communities increased from 15.7 to 16.6, which is not a statistically significant change (Fig. 6). There is a clear trend of expansion of some nutrient-demanding species (Athyrium distentifolium, Bistorta major, Crepis mollis, Leontodon hispidus, Senecio nemorensis agg.), especially at lower altitudes. This, hand in hand with expansion of taller plants, is reflected in light-demanding species decrease (Carex atrata, C. nigra, Dianthus superbus ssp. alpestris, Luzula sudetica, Omalotheca supina, Phleum rhaeticum, Pseudorchis albida). On the other hand, indicator values for moisture, soil reaction and temperature show no significant changes.

Comparison of historical and recent indicator values and diversity parameters. Changes in Shannon-Wiener index (t = 3.02; p-value = 0.007), Ellenberg indicator values for light (t = 5.70; p-value < 0.001) and nutrient (t = -3.92; p-value = 0.001) values were evaluated as statistically significant
Succession processes lead to a higher proportion of diagnostic species of other classes, as indicated by NMDS and bar chart (Figs. 7 and 8). We recorded a significant decrease in average cover of diagnostic species of Nardetea strictae class (from 58.5 to 28.8%; t = 7.0; p-value < 0.001), while those of Roso pendulinae-Pinetea (from 7.1 to 17.7%; t = -3.28; p-value = 0.004) and Loiseleurio-Vaccinietea (from 7.9 to 26.2%; t = -4.26; p-value < 0.001) have expanded in the communities. The overall change in the cover of the diagnostic species of Caricetea curvulae was not evaluated as statistically significant (increase from 11.1 to 13.7%; t = -0.95; p-value = 0.357). Nevertheless, mixed-effect models indicate their expansion on the steeper slopes (t = 2.34; p-value = 0.026), to a lesser extent, their coverage is increasing with increasing altitudes (t = 1.97; p-value = 0.054). On the contrary, lower altitudes seem to be expanded by diagnostic species of Roso pendulinae-Pinetea mugo class, especially by Avenella flexuosa, Calamagrostis villosa and Pinus mugo. However, this phenomenon is not statistically significant (t = -1.858; p-value = 0.073). The succession of most of the stands seems to lead to the communities of Loiseleurio-Vaccinietea class. On the other hand, expansion of dwarf shrubs seems to be indifferent to altitude, slope and aspect. Similar pattern is shown by some diagnostic species of Salicetea herbaceae class, especially by Festuca picturata, Gentiana punctata and Luzula alpinopilosa ssp. obscura. Diagnostic species of other classes remain without significant changes or have very low average cover in communities in both time periods.

NMDS ordination diagram using abundances of diagnostic species of Nardetea strictae and other classes occurring in the study area as supplementary variables. Only classes whose diagnostic species have a high average abundance (more than 7% in both time periods) are shown. Historical relevés are marked with the site number, whereas recent relevés correspond to arrow heads

Comparison of historical and recent average cover (%) of diagnostic species of all classes. A significant decrease in cover of Nardetea strictae species was recorded (from 58.5–28.8%; t = 7.0; p-value < 0.001), on the contrary, cover of species of Roso pendulinae-Pinetea (from 7.1–17.7%; t = -3.28; p-value = 0.004) and Loiseleurio-Vaccinietea (from 7.9–26.2%; t = -4.26; p-value < 0.001) increased. Diagnostic species of other classes remain without significant changes or have very low average abundance in both time periods
Discussion
Our results on enrichment of Nardus stricta grasslands with new species after the pasture abandonment comply with findings of other authors. This higher diversity is explained by negative correlation with coverage of Nardus stricta. As reported by Palaj and Kollár (2022), the richest in species are the current grasslands with Nardus stricta coverage lower than 25%. Increase in species richness along with decrease in cover of Nardus stricta are also reported by Sebastiá (2004) from the south-eastern Pyrenees Mts, Velev and Apostolova (2008) from Central Balkan National Park and Parolo et al. (2011) from Bergamasque Alps.
Current (sub)alpine grasslands in the Western Carpathians are formed by species of a wider ecological spectrum compared to the reference period. Higher proportion of species of Caricetea curvulae class is typical here, since it is a neighbouring vegetation (Kliment 2007a, b). After grazing cessation, these species find their optimum mainly on the higher altitudes with steep relief exposed to constant and strong winds (Dúbravcová and Jarolímek 2007).
At lower altitudes, their expansion is limited by a competition for light with taller species (Korzeniak 2016), which are able to utilize the nutrients from accumulated biomass (Dupré et al. 2010). Therefore, due to the easier nutrient availability at lower altitudes, Nardus stricta grasslands are modified mainly by less oligotrophic species (Erschbamer et al. 2003; Kurtogullari et al. 2020), whose frequencies and cover are declining with rising altitudes (Parolo et al. 2011). The marginal parts of pastures along the timber line are the most susceptible to such changes, where the proportion of oligotrophic species was highest in the past due to the relatively less depositions of livestock urinary and excrements (Skarpe and Hester 2008). Mainly at lower altitudes, grazing cessation is followed by an expansion of woody plants (Švajda et al. 2011; Palaj and Kollár 2021), since these are no longer blocked by browsing and trampling (Puccio et al. 2007; Skarpe and Hester 2008).
Due to the expansion of forest and shrub vegetation, the area of grasslands has decreased by 22% in the Tatra part of the study area over the last 50 years (Palaj and Kollár 2021). Higher proportion of species of Loiseleurio-Vaccinietea in current vegetation formed after grazing cessation complies with findings of numerous authors across the European mountains (Meshinev et al. 2000; Witkowska-Żuk and Ciurzycki 2000; Tasser and Tappeiner 2002; Erschbamer et al. 2003; Grigoriu and Alda 2004; Velev and Apostolova 2008; Parolo et al. 2011; Korzeniak 2016). Same trend was found also for less intensively used pastures (Bensettiti et al. 2005). It is necessary to emphasize that climate change has a similar impact on the expansion of dwarf shrubs (and also other shifts in species composition) as grazing cessation (Pauli et al. 2012) and these drivers operate in synergy and in a complex way (Kobiv 2017). Air pollution can play some role, too. For example, in Western Tatras, annual deposition of sulphur was estimated to reach 11 kg/ha and nitrogen 12 kg/ha, what can lead to soil acidification or eutrophication (Halada et al. 2009).
Another particular phenomenon is, regardless of site conditions and floristic composition changes, the accumulation of biomass, which no longer has a consumer. This, together with the subsequent expansion of taller species can be a reason of the decrease in the light-demanding species in the (sub)alpine Nardus grasslands. After the grazing cessation, light becomes a limiting factor (Borer et al. 2014; Gavrichkova et al. 2022), and change in the vegetation structure can lead to the local extinction of lower and competitively weaker species (Oksanen 1990; Alm and Often 1997).
Data Availability
Not applicable.
Code Availability
Not applicable.
References
AIm T, Often A (1997) Species conservation and local people in E Finnmark, Norway. Plant Talk 11:30–31
Altmanová M (1983) Subalpine and alpine vegetation of the Low Tatras and its assessment for the needs of LANDEP. Dissertation Thesis, Institute of Landscape Ecology (in Slovak)
Bates D, Maechler M, Bolker B, Walker S (2015) Fitting Linear Mixed-Effects Models using lme4. J Stat Softw 67(1):1–48
Bensettiti F, Boullet V, Chavaudret-Laborie C, Deniaud J (eds) (2005) Natura 2000 habitat notebooks. Knowledge and management of habitats and species. Volume 4 – agropastoral habitats. La Documentation française, Paris. (in French)
Biely A, Beňuška P, Bezák V, Bujnovský A, Haloužka R, Ivanička J, Kohiit M, Klinec A, Lukáčik E, Maglay J, Miko O, Pulec M, Putis M, Vozár J (1992) Geological map 1: 50 000 of the low Tatras. ŠGÚDŠ, Bratislava. (in Slovak)
Bohuš I (1966) History of the Liptov Tatras. Unpublished report, Tatranská Lomnica. (in Slovak)
Bohuš I (1994) Agriculture. In: Vološčuk I (ed) Tatra National Park. Biosphere Reserve, Gradus, Martin, pp 256–261. (in Slovak)
Borer E, Seabloom E, Gruner D, Harpole S, Hillebrand H, Lind E, Adler P, Alberti J, Anderson M, Bakker J, Biederman L, Blumenthal D, Brown C, Brudvig L, Buckley Y, Cadotte M, Chu Ch, Cleland E, Crawley M, Daleo P, Damschen E, Davies K, DeCrappeo N, Du G, Firn J, Hautier Y, Heckman R, Hector A, HilleRisLambers J, Iribarne O, Klein J, Knops J, La Pierre K, Leakey A, Li W, MacDougall A, McCulley R, Melbourne B, Mitchell Ch, Moore J, Mortensen B, O’Halloran L, Orrock J, Pascual J, Prober S, Pyke D, Risch A, Schuetz M, Smith M, Stevens C, Sullivan L, Williams R, Wragg P, Wright J, Yang L (2014) Herbivores and nutrients control grassland plant diversity via light limitation. Nature 508:517–520
Braun-Blanquet J (1964) Pflanzensoziologie. Gründzuge der Vegetationskunde. Springer Verlag, Wien & New York
Bublinec E, Linkeš V, Koreň M (1994) Soil characteristics. In: Vološčuk I (ed) Tatra National Park. Biosphere Reserve, Gradus, Martin, pp 86–105. (in Slovak)
Dúbravcová Z (1976) Subalpine and alpine vegetation in Kamenistá and Gáborova dolina valleys (Western Tatras). Dissertation Thesis, Comenius University (in Slovak)
Dúbravcová Z, Jarolímek I (2007) Caricetea curvulae. In: Kliment J, Valachovič M (eds) Plant communities of Slovakia. 4. High-mountain vegetation. Veda, Bratislava, pp 321–339 (in Slovak)
Dupré C, Stevens CJ, Ranke T, Bleeker A, Peppler-Lisbach C, Gowing DJG, Dise NB, Dorland E, Bobbink R, Diekmann M (2010) Changes in species richness and composition in european acidic grasslands over the past 70 years: the contribution of cumulative atmospheric nitrogen deposition. Global Change Biol 16:344–357
Ellenberg H, Weber HE, Düll R, Wirth V, Werner W, Paulissen D (1992) Zeigerwerte von Pflanzen in Mitteleuropa. Scr Geol 18:1–258
Erschbamer B, Virtanen R, Nagy L (2003) The impacts of vertebrate grazers on vegetation in european high mountains. In: Nagy L, Grabherr G, Korner Ch, Thompson DBA (eds) Alpine biodiversity in Europe. Springer-Verlag, Berlin, pp 377–396
Galvánek D, Janák M (2008) Management of Natura 2000 habitats. 6230 *Species-rich Nardus grasslands. European Commission. Technical Report 2008 14/24
Gavrichkova O, Pretto G, Brugnoli E, Chiti T, Ivashchenko K, Mattioni M, Moscatelli MC, Scartazza A, Calfapietra C (2022) Consequences of Grazing Cessation for Soil Environment and Vegetation in a Subalpine Grassland Ecosystem. Plants 11(16):2121
Grigoriu A, Alda N (2004) Nardus stricta L. meadow phytocoenology, synecology and chorology in the Timis river basin. Contrib Bot 39:95–104
Grime JP (1979) Plant strategies and vegetation processes. Wiley, Chichester
Halada Ľ, David S, Halabuk A (2009) Vegetation structure and Aboveground Biomass at Mt. Salatín Long-Term Ecological Research Site, the West Tatra Mts. Slovakia Ekológia (Bratislava) 28(2):113–126
Hamilton NE, Ferry M (2018) ggtern: Ternary Diagrams using ggplot2. J Stat Softw 87(3):1–17. https://doi.org/10.18637/jss.v087.c03
Häufler V (1955) Mountain areas in Czechoslovakia and their land use. Academia, Prague. (in Czech)
Helm A, Hanski I, Pärtel M (2006) Slow response of plant species richness to habitat loss and fragmentation. Ecol Lett 9:72–77
Hennekens S, Schaminée JHJ (2001) TURBOVEG, a comprehensive data base management system for vegetation data. J Veg Sci 12(4):589–591
Horák J (1970) Geobiocoenosis of timber line. Part I. – Western Tatras, Jamnická and Račkova dolina valleys. University of Agriculture, Brno. (in Czech)
Hrabovcová J (1976) Subalpine and alpine vegetation of Jamnická dolina valley (Western Tatras). Rigorous Thesis, Comenius University (in Slovak)
Hrivnák R, Ujházy L (2005) Changes of the mountain grassland vegetation after abandonment and colonization by Norway spruce. Ekológia (Bratislava) 24(3):231–253
Janišová M, Hájková P, Hegedüšová K, Hrivnák R, Kliment J, Michálková D, Ružičková H, Řezníčková M, Tichý L, Škodová I, Uhliarová E, Ujházy K, Zaliberová M (2007) Grassland vegetation of Slovak Republic – electronic expert system for identification of syntaxa. Institute of Botany SAS, Bratislava. (in Slovak)
Jarolímek I, Šibík J (eds) (2008) Diagnostic, constant and dominant species of higher vegetation units of Slovakia. Veda, Bratislava
Kliment J (2007a) Nardetea strictae. In: Kliment J, Valachovič M (eds) Plant communities of Slovakia. 4. High-mountain vegetation. Veda, Bratislava, pp 343–353. (in Slovak)
Kliment J (2007b) In: M (ed) Plant communities of Slovakia. 4. High-mountain vegetation. Veda, Bratislava. (in Slovak)
Kliment J, Ujházy K (2014) Nardetea strictae. In: Hegedüšová Vantarová K, Škodová I (eds) Plant communities of Slovakia. 5. Grassland vegetation. Veda, Bratislava, pp 385–444. (in Slovak)
Kliment J, Valachovič M (eds) (2007) Plant communities of Slovakia. 4. High-mountain vegetation. Veda, Bratislava. (in Slovak)
Klotz S, Kühn I (2002) Ökologische Strategietypen. In: Klotz S, Kühn I, Durka W (eds) BIOLFLOR: eine Datenbank mit biologisch-ökologischen Merkmalen zur Flora von Deutschland. BfN, Münster, pp 119–126
Kobiv Y (2017) Response of rare alpine plant species to climate change in the ukrainian Carpathians. Folia Geobot 52:217–226
Korzeniak J (2016) Mountain Nardus stricta grasslands as a relic of past farming –the effects of grazing abandonment in relation to elevation and spatial scale. Folia Geobot 51:93–113
Krahulec F, Chytrý M, Härtel H (2010) Nardus grasslands and heathlands. In: Chytrý M (ed) Vegetation of the Czech Republic. 1. Grassland and heathland. Academia, Praha, pp 281–319
Králik T (1979) Plant communities of the Spálená dolina valley. Diploma thesis, Comenius University (in Slovak)
Kremlová R (1974) Alpine and subalpine vegetation of Žiarska dolina valley (Western Tatras) Diploma thesis, Comenius University (in Slovak)
Kurtogullari Y, Rieder NS, Arlettaz R, Humbert J-Y (2020) Conservation and restoration of Nardus grasslands in the swiss northern Alps. Appl Veg Sci 23:26–38. https://doi.org/10.1111/avsc.12462
Lapin M, Faško P, Melo M, Šťastný P, Tomlain J (2002) Climatic regions. In: Miklós L (ed) Landscape Atlas of the Slovak Republic. MŽP, Bratislava. (in Slovak)
Linkeš V (1967) Soils of the low Tatras. Vedecké práce laboratórií pôdoznalectva 2:23–34 (in Slovak)
Marhold K, Hindák F (eds) (1998) Checklist of non-vascular and vascular plants of Slovakia. Veda, Bratislava
Massey FP, Ennos AR, Hartley SE (2007) Grasses and the resource availability hypothesis: the importance of silica-based defences. J Ecol 95(3):414–424
Meshinev T, Apostolova I, Kachaunova E, Velchev V, Bondev I (2000) Flora and plant communities. In: Popov A, Meshinev T (eds) High-mountain Treeless Zone of the Central Balкan National Park. Biological Diversity and Problems of its Conservation. BSBCP, Sofia
Midriak R (1983) Morphogenesis of high-mountain relief (morphological study with focus on soil destruction in western Carpathians). Veda, Bratislava. (in Slovak)
Midriak R (1994) Timberline. In: Vološčuk I (ed) Tatra National Park. Biosphere Reserve, Gradus, Martin, pp 313–327. (in Slovak)
Mišić V, Jovanović-Dunjić R, Popović M, Borisavljević L, Antić M, Dinić A, Danon J, Blaženčić Ž (1978) Plant communities and habitats of the Stara Planina Mountains. Srpska akademija nauka i umetnosti, Beograd (in Serbian)
Nemčok J (1994) Geological bedrock. In: Vološčuk I (ed) Tatra National Park. Biosphere Reserve, Gradus, Martin, pp 14–24. (in Slovak)
Oksanen L (1990) Predation, herbivory, and plant strategies along gradients of primary production. In: Grace JB, Tilman D (eds) Perspectives on plant competition. Academic Press, San Diego, pp 445–474
Oksanen J, Blanchet G, Friendly M, Kindt R, Legendre P, McGlinn D, Minchin PR, O’Hara RB, Simpson GL, Solymos P, Stevens MHH, Szoecs E, Wagner H (2020) vegan: Community Ecology Package. R package version 2.5-7. https://CRAN.R-project.org/package=vegan. Accessed 1 Mar 2022
Palaj A, Kollár J (2017) Contribution to the knowledge of the alpine vegetation of the western Tatras. Phytopedon (Bratislava) 16:9–13 (in Slovak)
Palaj A, Kollár J (2019) Contribution to the knowledge of the alpine tall-grass vegetation of the western Tatras (Slovakia). Phytopedon (Bratislava) 18:1–6 (in Slovak)
Palaj A, Kollár J (2021) Expansion of phanerophytes above the timberline in the western Carpathians. Biologia 76:1991–2003. https://doi.org/10.1007/s11756-021-00782-1
Palaj A, Kollár J (2022) Contemporary state of Nardus grasslands on the former pastures in the alpine zone of the western Carpathians. Phytopedon (Bratislava) 21:1–10 (in Slovak)
Parolo G, Abeli T, Gusmeroli F, Rossi G (2011) Large-scale heterogeneous cattle grazing affects plant diversity and forage value of Alpine species-rich Nardus pastures. Grass Forage Sci 66:541–550
Pauli H, Gottfried M, Dullinger S, Abdaladze O, Akhalkatsi M, Alonso JLB, Coldea G, Dick J, Erschbamer B, Calzado FR, Ghosn D, Holten JI, Kanka R, Kazakis G, Kollár J, Larsson P, Moiseev P, Moiseev D, Molau U, Mesa JM, Nagy L, Pelino G, Puşcaş M, Rossi G, Stanisci A, Syverhuset AO, Theurillat JP, Tomaselli M, Unterluggauer P, Villar L, Vittoz P, Grabherr G (2012) Recent plant diversity changes on Europe’s Mountain Summits. Science 336(6079):353–355
Pietorová E (1977) Alpine vegetation of the Račkova dolina valley (Western Tatras). Diploma Thesis, Comenius University (in Slovak)
Puccio C, Gusmeroli F, Della Marianna G, D’Angelo A (2007) Foraging circuits, selectivity and drawing quality in goat grazing in subalpine pasture areas. Quaderno SOZOOALP 4:83–120 (in Italian)
Raunkiaer C (1934) The life forms of plants and statistical plant geography. Clarendon Press, Oxford
RStudio Team (2022) RStudio: Integrated Development for R. http://www.rstudio.com/. Accessed 1 Mar 2022
Sebastiá M-T (2004) Role of topography and soils in grassland structuring at the landscape and community scales. Basic Appl Ecol 5:331–346
Skarpe C, Hester A (2008) Plant traits, browsing and grazing herbivores, and vegetation dynamics. In: Gordon J, Prins HHT (eds) The ecology of browsing and grazing. Ecological studies, vol 195. Springer, Berlin & Heidelberg, pp 217–261
Šmarda J (1963) Secondary Plant Communities in the Tatra National Park. Šport, Bratislava & Tatranská Lomnica. (in Slovak)
Šomšák L, Balkovič J (2002) Cyclic succession and plant biodiversity within the secondary spruce forests in the Hnilec river watershed. Phytopedon (Bratislava) 1:45–51
Švajda J, Solár J, Janiga M, Buliak M (2011) Dwarf pine (Pinus mugo) and selected abiotic habitat conditions in the western Tatra Mountains. Mt Res Dev 31(3):220–228
Tasser E, Tappeiner U (2002) Impact of land use changes on mountain vegetation. Appl Veg Sci 5:173–184
Tichý L (2002) JUICE, software for vegetation classification. J Veg Sci 13:451–453
Treskoňová M (1972) Bald mountains of the central and the western part of Low Tatras. Diploma Thesis, Comenius University (in Slovak)
Turečková J (1974) Subalpine and alpine vegetation of Jamnická dolina valley (Western Tatras). Diploma Thesis, Comenius University (in Slovak)
Vandvik V, Birks HJB (2002) Partitioning floristic variation in norwegian upland grasslands into within–site and between–site components: are the patterns determined by environment or by land–use? Pl Ecol 162:233–245
Velev N, Apostolova I (2008) Successional changes of Nardus stricta communities in the Central Balkan Range (Bulgaria). Phytol Balc 14(1):5–84
Wickham H (2016) ggplot2: Elegant graphics for data analysis. https://ggplot2.tidyverse.org. Accessed 1 Mar 2022
Witkowska-Żuk L, Ciurzycki W (2000) Vegetation succession in the areas excluded from sheep grazing in the Tatra National Park during 1965–1994. Ochr Przyr 57:19–40 (in Polish)
Zarzycki J, Korzeniak J, Perzanowska J (2022) Impact of Land Use Changes on the diversity and conservation status of the vegetation of Mountain Grasslands (polish Carpathians). Land 11(2):252. https://doi.org/10.3390/land11020252
Acknowledgements
The research was financially supported by the VEGA grant agency project 2/0048/22.
Funding
Open access funding provided by The Ministry of Education, Science, Research and Sport of the Slovak Republic in cooperation with Centre for Scientific and Technical Information of the Slovak Republic. This work was supported by the VEGA grant agency project 2/0048/22.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
Authors have no conflict of interest.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Palaj, A., Kollár, J. & Michalová, M. Changes in the Nardus grasslands in the (Sub)Alpine Zone of Western Carpathians over the last decades. Biologia 79, 1081–1090 (2024). https://doi.org/10.1007/s11756-023-01458-8
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11756-023-01458-8