
Abstract
Understanding what environmental factors are genetically linked to a phenological event is critical for predicting responses to climate change. Photosynthetic phenology often varies among a species of evergreen conifers due to local adaptation. However, few empirical studies have revealed relevant relationships between climatic factors in provenance environments and photosynthetic phenology. This study evaluated the effects of environmental conditions of the growing site and seed source provenance on the seasonal changes in maximal photochemical quantum yield of photosystem II (Fv/Fm) in a common garden experiment with 2-year-old seedlings of Sakhalin fir (Abies sachalinensis), a representative species with local adaptation, from four seed source provenances. A logistic model was constructed to explain the seasonal variation of Fv/Fm from July to October and the relationships between the estimated model parameters and representative factors featuring provenance environments were evaluated. The landscape gradient of the detected model parameters responsible for the provenance environments was visualized in a map of the distribution area. The lowest temperature was the most plausible factor in the growing environment to explain the seasonal changes of Fv/Fm. Among the representative meteorological factors of provenance environments, the lowest temperatures in July showed significant relationships with two model parameters, explaining the lower limit of Fv/Fm and the higher sensitivity of autumn Fv/Fm decline. The estimated spatial maps of model parameters consistently showed that the higher the lowest temperature in July in the provenance environment, the lower the Fv/Fm in October and the greater the decrease in the autumn Fv/Fm decline. Therefore, the lowest summer temperature could be associated with the local adaptation of autumn photosynthetic phenology in A. sachalinensis.
Similar content being viewed by others

Introduction
The phenology, or seasonal specific phases in life history can be under natural selection (Forrest and Miller-Rushing 2010). The patterns of phenotypic responses to environmental stimuli are genetically governed, which is known as the reaction norm (West-Eberhard 2003). For tree, the phenology as an evolutionally developed pattern of phenotypic responses is critical since they survive in the environment where multiple conditions change in synchrony even with occasional noise. It has been often recognized as properly governing the initial phases and end of the maturation period of a phenotypic trait (Fréchette et al. 2020; Casmey et al. 2022) when climatic conditions drastically change. Understanding the effects of seasonal climatic factors on phenotypes and the consequences of genetic variations in phenotypes will aid in predicting tree responses to climate change (Aitken and Whitlock 2013; Franks et al. 2014).
In evergreen conifers, photosynthetic activity often declines in the autumn (Sakai 1983; Öquist and Huner 2003). At low temperatures, the generation rate of NADP in photochemical reactions often exceeds the carbon fixation rate, resulting in the accumulation of excess light energy that can be lethal to cells and the entire plant (Chang et al. 2021). Photoinhibition caused by different temperature-dependent photosynthetic systems can be regulated by several physiological mechanisms, such as a reduction in light absorption by chlorophyll and nonphotochemical quenching (Ensminger et al. 2004; Verhoeven et al. 2009). As an indicator of seasonal photosynthetic decline, the maximal photochemical quantum yield of photosystem II (Fv/Fm) is widely used because it is easily measured (Fréchette et al. 2020; Chang et al. 2021). Declines in Fv/Fm in the autumn correspond to seasonal temperature dynamics, especially the lowest temperature (Öquist and Huner 2003).
The autumn decline in Fv/Fm can be regulated not only by environmental conditions but also by genetic factors (Fréchette et al. 2020). The local environmental factors for the tested population could be associated with the intraspecific variation (Aitken and Adams 1996), indicating that relevant climatic factors might be responsible for the differentiation of phenotypes. For example, the photosynthetic activity of seedlings of boreal evergreen conifers originating from a northern provenance declines earlier than in seedlings originating from a southern provenance (Fréchette et al. 2020; Noordermeer et al. 2021). As several studies proposed (Aitken and Adams 1996; Öquist and Huner 2003; Fréchette et al. 2020), the lowest temperature in winter and/or summer provenance environments would be a candidate factor associated with genetic variation in the photosynthetic phenology. However, a genetic cline along with original provenances has been associated with an index that integrates multiple climatic factors (Csilléry et al. 2020). Although it is critical to determine the climatic factors that are spatiotemporally associated with genetic variation (Ishizuka et al. 2021), few empirical studies have been done so far.
Sakhalin fir (Abies sachalinensis), an evergreen conifer species, is distributed throughout the northernmost large island of Japan, Hokkaido, where gradients of various environmental conditions are present at a landscape scale (Fig. 1, Fig. S1). In this species, intraspecific variation has been widely reported for traits such as home-advantage tree growth, frost tolerance of winter buds, and disease resistance, which are considered as the consequence of local adaptation to winter conditions (Sakai 1983; Eiga 1984; Tsuyama et al. 2020; Ishizuka et al. 2021). Therefore, Sakhalin fir is suitable for evaluating intraspecific variation along a landscape gradient in the local environment (Kitamura et al. 2020; Ishizuka et al. 2021).

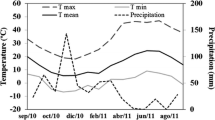
Temperature and daylight hours at the experimental site (a, b) and local environment of original provenance regions (c, d). a Daily highest (light grey dotted), average (grey straight) and lowest temperatures (black dotted). c, d Variation in average environmental conditions in provenance regions of three mother trees: north (black dot), south (grey square), northeastern (white diamond), and eastern edge (white triangle)
This study aimed to address which climatic factors in provenance environments affect seasonal photosynthetic changes in evergreen conifers. We assumed that the earlier the winter lowest temperature occurs in the provenance environment, the faster that the photosynthetic phenology advances, resulting in genetic variation in autumn decline of Fv/Fm. To verify this hypothesis, we (1) explored which growing conditions regulate the autumn decline of Fv/Fm, (2) assessed the association of representative factors in the provenance environments with the autumn photosynthetic dynamics using a logistic model of the photosynthetic phenology (Gonsamo et al. 2012), and (3) visualized a landscape gradient of the detected model parameters responsible for the provenance environments in a map of the distribution area. In relation to the geographical gradient in the autumn photosynthetic decline, we discuss forest management of Sakhalin fir seedlings under regional-specific climate change.
Materials and methods
Experimental design
In a common garden trial in the nursery of the Sapporo Experimental Forest of Hokkaido University (43.068° N, 141.338° E), we planted seedlings of Sakhalin fir from four seed source provenances that had been grown from seeds for 2 years at the nursery of Forestry Research Institute, Hokkaido Research Organization in central Hokkaido (43.290° N, 141.855° E). Open-pollinated seeds were collected from 12 mother trees in several blocks of a seed orchard (42.448° N, 142.477° E) where block separation was introduced along with the seed zoning of Hokkaido. We had four seed source provenances: north region (N), south region (S), northeastern region (NE) and eastern edge region (EE) (Table S1, Fig. S1). In the common garden trial, we selected four seedlings per mother tree to represent a family line. Therefore, we used 48 seedlings (4 seedlings × 3 mothers × 4 seed source provenances). In early May 2018 before the buds burst, the seedlings were transplanted into 4-L pots containing a 1:1 volume ratio of Kanuma soil (i.e., pumiceous soil) and Akadama soil (i.e., claylike red soil) to which 100 mL of commercial liquid fertilizer was applied (HYPONEX, HYPONex Japan Co., Ltd., Osaka, Japan; N = 72.4 mg L−1, P = 284 mg L−1, K = 94.2 mg L−1). The initial height and root collar diameter of the seedlings were then measured using a ruler and electronic caliper. The mean height ± standard error for seedlings in each provenance was 17.6 ± 0.23 cm for the N, 15.6 ± 0.44 cm for the S, 16.5 ± 1.36 cm for the NE, and 15.0 ± 0.28 cm for the EE and the mean collar diameter value ± standard error was 4.48 ± 0.15 mm for the N, 4.27 ± 0.17 mm for the S, 4.56 ± 0.17 mm for the NE, and 4.48 ± 0.22 mm for the EE. All pots were tagged for individual identification, and four groups of 12 combinations by mother tree and origin (12 pots per group) were set. In the nursery of the experimental site, each group was arranged with at least approximately 50 cm between them, and pots within a group were randomly arranged. To prevent any drought stress during the experiment, the plants were irrigated with tap water every 1–2 weeks. Meteorological data at the experimental site was obtained from the Agro-meteorological grid square data (https://amu.rd.naro.go.jp/) where the spatial resolution of grid cells was set as approximately 1 km × 1 km. The environmental conditions of the experimental site and provenance regions are summarized in Fig. 1.
Chlorophyll fluorescence
On 23 July, 22 August, and 18 October 2018, all 48 seedlings were acclimated in the dark for 3–4 h after sunset (18:00), then a current-year, green needle was collected from the second branch of the stem, and a fixed distance was set between each needle and the light source of a portable chlorophyll fluorometer system (JUNIOR-PAM, Walz, Effeltrich, Germany using a leaf clip. The blue LED light source in the system (maximum emission of 450 nm) was used to measure the minimum fluorescence yield (F0). Then the maximum fluorescence yield (Fm) was measured using a saturating pulse light (> 5000 μmol m−2 s−1 for 0.7 s). The maximal photochemical quantum yield of PSII (Fv/Fm) was calculated as (Fm − F0)/Fm (Genty et al. 1989).
Modeling F v/F m in the growing conditions
To estimate the autumn decline of Fv/Fm, we explored the environmental factors that regulated the seasonal photosynthetic changes in the seedlings. The reaction norm is determined by genetic variation based on the basic relationship between phenotypes and effects of the environment (West-Eberhard 2003). Because the phenotypes should sequentially respond to the environment at variable time scales, the seasonal photosynthetic decline gradually proceeds in response to stimuli from a few days to a few weeks (Öquist and Huner 2003). Based on that concept, we evaluated the relationship between seasonal Fv/Fm changes and the factors in the growth environment with a reasonable period of environmental stimulation. For this process, we tested for a correlation between the seasonal Fv/Fm variation and temperature factors or daylight hour averaged on different days after each measurement date (Fig. S2). The factors temperature and daylight hour were highly relevant to the seasonal photosynthetic decline (Öquist and Huner 2003; Noordermeer et al. 2021) although the seasonal light dynamics indirectly regulated photosynthetic decline in autumn. In fact, reduced sink capacity and growth cessation can be directly linked to seasonal photosynthetic decline (Noordermeer et al. 2021). Since previous studies have evaluated the effects of photoperiod on photosynthetic phenology in the manipulated conditions without clouds or shadows (e.g., Busch et al. 2007; Noordermeer et al. 2021), the daylight hour was adopted in this study as an alternative to photoperiod.
We then established the model response to the environmental factor with a logistic function. Since Fv/Fm saturates at a constant value and does not continue to change monotonically, it is appropriate to describe its seasonal variation for this phenotype using a nonlinear model. By utilizing the selected environmental factors, the Fv/Fm data for each seedling was described by the following logistic function with four model parameters of α1, α2, δ, and β (Gonsamo et al. 2012; Fréchette et al. 2020):
where y(e) is the measured value at a given environmental condition e, α1 is the lower limit value during measurement periods, α2 is the averaged value in summer steady state, δ is the autumn decline normalized slope coefficient and β is the midpoint in the environmental condition of the autumn decline.
Clustering of provenance environments
To select the representative factors explaining the seed source provenance conditions of Sakhalin fir, the meteorological data was obtained from the regional Policy Bureau of the Ministry of Land, Infrastructure, Transport and Tourism of Japan (https://nlftp.mlit.go.jp). The data set included for each month the highest, lowest, and average temperature; precipitation total, snowfall maximum, total daylight hours, and average of the daily total solar radiation, estimated and calculated from observed values for each 1-km mesh for the past 30 years by the Japan Meteorological Agency (http://www.jmbsc.or.jp). The data set was used for a principal component analysis (PCA). Since the values for the different environmental factors have different units, standardization was needed to use them in the analysis. All factors were classified into clusters based on the variability of the principal component loadings, and the k-means method was employed for a nonhierarchical clustering analysis. The most representative factors of the provenance environmental conditions were selected based on the rank of both indices, i.e., the variable of the PC1 loading and the correlation coefficient value between PC1 and each factor, where the factors ranked close in consecutive months were considered to be particularly representative of the corresponding season. The correlation coefficient was calculated using Pearson’s correlation analysis.
Statistical analyses and geographic visualization
All statistical analyses were performed using R version 4.2.1 (R Core Team 2022) and Excel ver. 16.61 (Microsoft, Redmond, WA, USA). The significance level was set at 5%. First, a repeated measures ANOVA was used to test for significant differences in the raw values of Fv/Fm between provenance regions and seasonal changes, with provenance regions, measurement months, and its interaction as fixed factors and the individual 48 pots as a random factor with repeated measures. However, the main purpose of this study was to determine which factors in the provenance environments are associated with the genetic variation in the parameters in the reaction norm for modeling the seasonal changes of Fv/Fm. Therefore, a t-test was used to test for effects of the selected representative factors in provenance environments on the parameters of the Fv/Fm model. The relationships between the selected factors in the provenance environments and the model parameters were estimated using a generalized linear model. The landscape maps of model parameters were projected based on the meteorological data of spatial grid cells through the distribution area (latitude: 41.30°–45.50°, longitude: 139.30°–145.90°). The meteorological data were obtained from the Agro-meteorological grid square data, constructed by the Japan Meteorological Agency (http://www.jmbsc.or.jp). The estimated parameters in the projected map were extracted from the environmental range of the four provenances and the growing environments of this experiment during the measurements. QGIS ver.3.4 (QGIS Development Team 2018) was used for the projection.
Results
Variation of F v/F m among provenances and seasons and their correlations with climatic factors at the nursery
The significant decline of Fv/Fm was observed in October (Fig. 2) as demonstrated by repeated measures ANOVA (P < 0.001) although no differences were detected among provenance regions (Table S3). Correlation analyses showed higher correlation coefficients for temperature parameters than for the daylight hour (Fig. 2). Among the temperature parameters, the average values for 3 days consistently had relatively high coefficients. Particularly for the lowest temperature, correlation coefficients remained high (r > 0.84) until the end of the month compared to other temperature parameters. In this study, therefore, the lowest temperature averaged for 3 days was considered as the candidate cue regulating the autumn photosynthetic decline.

Seasonal and regional variation in the average maximum quantum yield (Fv/Fm) after dark acclimatization for at least 3 h. Provenance regions were north region (N), south region (S), northeastern region (NE), and eastern edge region (EE). The relevant statistical results by repeated measures ANOVA were shown in supplemental Table 3
Clustering provenance environments
The PCA results showed that 85.1% of the total profile represented by PC1 and PC2 (Fig. 3, PC1: 65.7%, PC2: 19.4%) where the differences of the four provenance regions were clearly distinguished (Fig. 3). All meteorological factors were classified in two clusters. In cluster 1, mostly composed of the factors of positive PC1 loading, the highest coefficients and PC1 loadings were found for the highest temperatures from November to January. In cluster 2, composed of the factors of negative PC1 loading, relatively lower PC1 loadings were widely found for several factors, and these values did not vary. On the other hand, variations in the low correlation coefficients showed a clear difference between meteorological factors, and the coefficients were lowest for the lowest temperature from June to July. Based on these results, the highest temperature in November for cluster 1 and the lowest temperature in July for cluster 2 were selected as the representative factors in seed source provenance conditions.

a Principal component analysis (PCA) of monthly meteorological factors in four provenance regions, i.e., north region (N), south region (S), northeastern region (NE), and eastern edge region (EE), and b clustering of PC1 loading for each factor and their correlation coefficients with PC1. Abbreviations for all meteorological factors are summarized in Supplemental Table 2
Landscape gradient of reaction norms
Significant relationships were recognized between the lowest temperature of July in provenance environments and several model parameters (Table 1, P < 0.05). A negative relationship with α1 and a positive relationship with δ suggested that the higher the lowest temperature in the provenance environments, the lower the lower limit value during autumn and the earlier the autumn photosynthetic decline. Based on the established models, the landscape gradient of α1 and δ associated with provenance environments were visualized (Fig. 4). On the other hand, there was no significant relationship with the highest temperature of November.

Landscape gradient of the responsible model parameters for describing the seasonal changes of Fv/Fm. The background autumn parameter (α1) shown in a and the decline in the normalized slope coefficient from summer to autumn (δ) shown in b were estimated based on the lowest temperature in July averaged over the past 30 years
Seasonal variation in F v/F m in relation to the seed source provenance climatic factors
The environmental range for extracting the parameters in the projected map was constructed using 339 points in the climatic mesh map. The decline in α1 and the lowest temperature in growing environments was associated with the lowest temperature in the seed source provenance environments (Fig. 5a). Similarly, the δ variation had a linear gradient along with the lowest temperature in provenance environments (Fig. 5b). These variable patterns demonstrated that when the lowest temperature of July decreased by more than 1 °C in the provenance environment, α1 increased by approximately 10%, and δ decreased by approximately 20%.

a Relationship between lowest temperature and variation of estimated background autumn parameter (α1) with the lowest temperature in July at the provenances and b variation of estimated decline in the normalized slope coefficient (δ) with four ranges of the lowest temperature of July in provenances based on the 1-km mesh map of Hokkaido, Japan for 21–23 July, 20–22 August, and 16–18 October 2018. The range of the lowest temperature of July in provenances is denoted by the following colors: 12.0 (purple): below –12.25 °C, 12.5 (blue): 12.25–12.75 °C, 13.0 (green): 12.75–13.25 °C, and 13.5 (yellow): 13.25 °C—above. In b, the number of mesh points selected in each class is indicated in the white box
Discussion
This study revealed the lowest temperature in July in provenance environments as the responsible factor associated with the intraspecific variation in seasonal changes of Fv/Fm and the landscape gradient of the key parameters for this phenology. In evaluating the plastic responses to seasonal conditions in the natural distribution environment, previous common garden experiments traditionally are limited by the meteorological conditions at the experimental site (Csilléry et al. 2020). In the present study, we separately evaluated the interaction between the effects of the conditions in both the experimental and the provenance environments using logistic modeling for the photosynthetic phenology (Gonsamo et al. 2012). The modeling results are scalable for a fine-scale landscape map, uncovering the potential range of phenological responses limited by the experimental site. This novel framework that can be used to address spatiotemporal intraspecific variation (Sang et al. 2021) and predict optimal plantation area and seasonal risks as the climate changes (Franks et al. 2014; Tsuyama et al. 2020; Casmey et al. 2022).
Environmental factor regulating autumn photosynthetic decline
The autumn decline of Fv/Fm was associated with the lowest temperature at the growth site, consistent with the findings of a previous study (Öquist and Huner 2003). The autumn photosynthetic decline has been suggested to be a functional phase to protect against photoinhibition in the winter (Adams and Demming-Adams 1995; Öquist and Huner 2003), and optimal regulation of PSII should contribute to the restart of photosynthesis the next season (Öquist and Huner 2003). We did not find a clear relationship between seasonal Fv/Fm variation and daylight hour with averaged days (Fig. 1; Fig. S2). Photoperiod alone does not affect the seasonal photosynthetic dynamics (Busch et al. 2007). Since we did not evaluate the sink capacity or growth cessation, we do not know how these factors affected the autumn photosynthetic decline in Sakhalin fir. Noordermeer et al. (2021) reported that photoperiod alone did not induce a seasonal photosynthetic decline in Douglas-fir (Pseudotsuga menziesii) under simulated conditions. This may partly support our results that the longer the averaged days, the lower the coefficients of correlation between daylight hour and seasonal Fv/Fm variation (Fig. S2).
Gradient intraspecific variation along with provenance conditions
Contrary to our expectation, the lowest temperature in July for the seed source provenances was the responsible factor associated with the intraspecific variation in the autumn decline of Fv/Fm. Subdividing the parameters of the nonlinear dynamics enabled us to visualize the landscape gradient of intraspecific variation. This is the first empirical data on the factors associated with the intraspecific variations in photosynthetic phenology, consistent with a recent study (Fréchette et al. 2020). Autumn photosynthetic activities are often maintained in evergreen conifers by specific regulation of compounds such as xanthophyll cycle pigments and antioxidant enzymes (Chang et al. 2021), probably for maintaining carbon reduction as long as possible in a differentiated niche. These studies indicate the existence of an adaptive photosynthetic phenology in evergreen conifers (Fréchette et al. 2020; Chang et al. 2021). If the autumn decline of Fv/Fm is adaptive, it may be linked to an acclimation phase for winter (Adams and Demming-Adams 1995; Öquist and Huner 2003). Nevertheless, in Sakhalin fir, a subarctic evergreen conifer, the summer provenance factor, rather than winter, was detected as a more plausible factor associated with genetic variation. This gap may be interpreted by the provenance variation in the length of carbon gain period. It has been proposed that the more unsuitable photosynthetic conditions are, the more carbon gain is compensated by prolonged leaf life span (Kikuzawa and Ackerly 1999) although the leaf life span of the evergreen conifer itself did not show any intraspecific difference (Reich et al. 1996). The seasonal changes in the monthly mean temperature of the provenances might support this assumption since the higher mean temperature in summer, but its relatively quick decline, was observed especially in the northern region (Fig. 1) where the lowest temperature in July was higher than in other regions (Fig. S1). Furthermore, the geographical gradients of the lowest temperature in July showed a significance temperature difference of more than 10 °C at maximum on this island. These points partly imply that populations with a relatively long period of carbon gain and probably a low capacity for carbon gain, might have adapted to local environments where growth conditions were less desirable especially in the summer. On the other hand, we did not measure intraspecific variations in any other parameters such as photosynthetic rates, leaf morphology, and leaf life span. Therefore, the degree to which genetic differentiation for the period of carbon gain is associated with the summer provenance factor is unclear. To verify whether the autumn decline of Fv/Fm is associated with adaptive photosynthetic phenology, further experiments are needed to evaluate the seasonal variations of other morphophysiological traits, gas-exchange capacity, and especially the length of time it can function, but not the leaf life span itself (Reich et al. 1996).
Conclusion
This study revealed that the autumn decline of Fv/Fm in Sakhalin fir was regulated by the lowest temperature under growing conditions, and their phenological pattern was associated with the lowest temperature of July in provenance conditions. Our results were provided from a novel framework for visualizing a landscape gradient that considers the environmental conditions of both the growing site and seed source provenance, emphasizing that the lower the lowest temperature in July at the provenances, the higher the Fv/Fm in October and the slower the seasonal photosynthetic decline, and vice versa. In Hokkaido, daily minimum temperatures have been increasing over the long term, and the extent of its increase was greater for daily minimum temperatures than for daily mean temperatures (Sapporo Regional Headquarters 2017). In the future, therefore, Sakhalin fir seedlings, especially those originating from provenances with the lowest temperatures in summer, may not undergo a suitable autumn decline of Fv/Fm, which may lead to lower tolerance of extreme meteorological events such as sudden low temperature in autumn. In order to achieve the healthy growth of seedlings not only in nurseries but also in transplant fields during climate change and extreme weather conditions, it would be critical for forest managers to choose the best combination of seed source provenance and planting areas for the environmental changes expected in each region (Tsuyama et al. 2020; Sang et al. 2021; Casmey et al. 2022).
References
Adams WW, Demmig-Adams B (1995) The xanthophyll cycle and sustained thermal energy dissipation activity in Vinca minor and Euonymus kiautschovicus in winter. Plant Cell Environ 18(2):117–127
Aitken SN, Adams WT (1996) Genetics of fall and winter cold hardiness of coastal Douglas-fir in Oregon. Can J For Res 26(10):1828–1837
Aitken SN, Whitlock MC (2013) Assisted gene flow to facilitate local adaptation to climate change. Annu Rev Ecol Evol Syst 44(1):367–388
Busch F, Huner NP, Ensminger I (2007) Increased air temperature during simulated autumn conditions does not increase photosynthetic carbon gain but affects the dissipation of excess energy in seedlings of the evergreen conifer Jack pine. Plant Physiol 143(3):1242–1251
Casmey M, Hamann A, Hacke UG (2022) Adaptation of white spruce to climatic risk environments in spring: implications for assisted migration. For Ecol Manag 525(1):120555
Chang CYY, Bräutigam K, Hüner NP, Ensminger I (2021) Champions of winter survival: cold acclimation and molecular regulation of cold hardiness in evergreen conifers. New Phytol 229(2):675–691
Csilléry K, Ovaskainen O, Sperisen C, Buchmann N, Widmer A, Gugerli F (2020) Adaptation to local climate in multi-trait space: evidence from silver fir (Abies alba Mill.) populations across a heterogeneous environment. Heredity 124(1):77–92
Eiga S (1984) Ecogenetical study of the freezing resistance of Saghalin fir (Abies sachalinensis MAST.) in Hokkaido. Bull For Tree Bree Inst 2(1):61–107
Ensminger I, Sveshnikov D, Campbell DA, Funk C, Jansson S, Lloyd J, Shibistova O, Öquist G (2004) Intermittent low temperatures constrain spring recovery of photosynthesis in boreal Scots pine forests. Glob Change Biol 10(6):995–1008
Forrest J, Miller-Rushing AJ (2010) Toward a synthetic understanding of the role of phenology in ecology and evolution. Philos Trans R Soc Lond B Biol Sci 365(1555):3101–3112
Franks SJ, Weber JJ, Aitken SN (2014) Evolutionary and plastic responses to climate change in terrestrial plant populations. Evol Appl 7(1):123–139
Fréchette E, Chang CYY, Ensminger I (2020) Variation in the phenology of photosynthesis among eastern white pine provenances in response to warming. Glob Change Biol 26(9):5217–5234
Genty B, Briantais JM, Baker NR (1989) The relationship between the quantum yield of photosynthetic electron transport and quenching of chlorophyll fluorescence. Biochim Biophys Acta Gen Subj 990(1):87–92
Gonsamo A, Chen JM, Price DT, Kurz WA, Wu C (2012) Land surface phenology from optical satellite measurement and CO2 eddy covariance technique. J Geophys Res 117(G3):G03032
Ishizuka W, Kon H, Kita K, Kuromaru M, Goto S (2021) Local adaptation to contrasting climatic conditions in Sakhalin fir (Abies sachalinensis) revealed by long-term provenance trials. Eco Res 36(4):720–732
Kikuzawa K, Ackerly D (1999) Significance of leaf longevity in plants. Plant Species Biol 14(1):39–45
Kitamura K, Uchiyama K, Ueno S, Ishizuka W, Tsuyama I, Goto S (2020) Geographical gradients of genetic diversity and differentiation among the southernmost marginal populations of Abies sachalinensis revealed by EST-SSR polymorphism. Forests 11(2):233
Noordermeer D, Velasco V, Ensminger I (2021) Autumn warming delays downregulation of photosynthesis and does not increase risk of freezing damage in interior and coastal Douglas-fir seedlings. Front For Glob Change. https://doi.org/10.3389/ffgc.2021.688534
Öquist G, Huner NP (2003) Photosynthesis of overwintering evergreen plants. Annu Rev Plant Biol 54(1):329–355
QGIS Development Team J (2018) QGIS geographic information system. Open source geospatial foundation project
R Core Team (2022) R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. Website https://www.R-project.org
Reich PB, Oleksyn J, Modrzynski J, Tjoelker MG (1996) Evidence that longer needle retention of spruce and pine populations at high elevations and high latitudes is largely a phenotypic response. Tree Physiol 16(7):643–647
Sakai A (1983) Comparative study on freezing resistance of conifers with special reference to cold adaptation and its evolutive aspects. Can J Bot 61(9):2323–2332
Sang Z, Hamann A, Aitken SN (2021) Assisted migration poleward rather than upward in elevation minimizes frost risks in plantations. Clim Risk Manag. https://doi.org/10.1016/j.crm.2021.100380
Sapporo Regional Headquarters (2017) Climate change in Hokkaido: past 120 years and projections for the future, 2nd edn, Japan. https://www.data.jma.go.jp/sapporo/bosai/publication/kiko/kikohenka/pdf/report.pdf
Tsuyama I, Ishizuka W, Kitamura K, Taneda H, Goto S (2020) Ten years of provenance trials and application of multivariate random forests predicted the most preferable seed source for silviculture of Abies sachalinensis in Hokkaido, Japan. Forests 11(10):1058
Verhoeven A, Osmolak A, Morales P, Crow J (2009) Seasonal changes in abundance and phosphorylation status of photosynthetic proteins in eastern white pine and balsam fir. Tree Physiol 29(3):361–374
West-Eberhard MJ (2003) Development. In: West-Eberhard MJ, Mary J (eds) Developmental plasticity and evolution. Oxford University Press, pp 89–90
Acknowledgements
We thank Mr. K. Ichikawa and the staff of the Sapporo experimental site in Hokkaido University for managing the nursery and Drs. S Goto, T Nagamitsu, I Tsuyama and the anonymous reviewers for insightful comments that helped to improve the manuscript.
Author information
Authors and Affiliations
Corresponding author
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Project funding: The work was supported by Japan Society for the Promotion of Science, KAKENHI (18J2013908).
The online version is available at http://www.springerlink.com
Corresponding editor: Yanbo Hu.
Guest editor: Yanbo Hu.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Sugai, T., Ishizuka, W. & Watanabe, T. Landscape gradient of autumn photosynthetic decline in Abies sachalinensis seedlings. J. For. Res. 34, 187–195 (2023). https://doi.org/10.1007/s11676-022-01592-0
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11676-022-01592-0