
Abstract
African teak (Milicia excelsa (Welw.) C.C. Berg) is an endangered multi-use species. Understanding the impact of climate change on the distribution of this species may improve the ability to anticipate or recognize its decline or expansion and to take appropriate conservation measures if necessary. Ecological niche modeling was projected in geographical space to study the current and future distribution of M. excelsa in Bénin. MaxEnt was used to estimate the potential geographic distribution of the species under two Representative Concentration Pathways (RCP). Miroc 5 summaries and two RCP 4.5 and RCP 8.5 scenarios were used as predictor variables for projections of the geographic potential of this species. The performance of the model was assessed by the area under the curve (AUC), true skill statistics (TSS) and partial receiver operating characteristics (Partial ROC). From the results, M. excelsa was more a secondary species in the Guinean climatic zone and part of the Sudanian-Guinean and Sudanian climatic zone. The projections show a significant decrease in suitable habitats for the species from the two RCP scenarios. Only a part of the Guinean climatic zone remained suitable and few protected areas will conserve in situ M. excelsa. For the sustainable conservation of M. excelsa, it is essential to strengthen the protection of sacred forests located in the Guinean climatic zone.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Present global plant extinction rates are 100–1000 times greater than pre-human levels and could increase tenfold by the next century (Pimm et al. 1995; Ricketts et al. 2005). Native species re-introduction is a widely used method to conserve endangered species which often face population growth challenges due to dispersal limitations and transient seed banks (Thompson 1997; Clarke et al. 2007). Despite its widespread use, re-introduction success rates are low due to the difficulty in identifying suitable habitats for restoration (Godefroid et al. 2011; Drayton and Primack 2012; Questad et al. 2014; Kakpo et al. 2018). Therefore, there is a need for methods to identify suitable habitats for endangered species and evaluate environmental factors that influence their distribution (Rovzar et al. 2016).
Knowledge of the spatial and temporal distribution of a species is key for evaluating extinction risks and forecasting possible future threats from factors such as climate change (Pacifici et al. 2015; Urban 2015; Ganglo and Kakpo 2016). Ecological niche models (ENMs) are used to understand ecological requirements of species, predict geographic distributions, select areas for conservation and forecast effects of environmental change (Peterson et al. 2011; Ganglo et al. 2017; Djotan et al. 2018). Studies of geographic distributions of vector species using ENM techniques relate occurrence records and environmental characteristics across species distributions (Altamiranda-Saavedra et al. 2017).
The application of ENMs in biodiversity research is well-known for many economically important species in West Africa, especially in Benin. For instance, Fandohan et al. (2015) modeled the ecological niche of Thunbergia atacorensis Akoègn. & Lisowski to investigate how climate change could affect future geographical range of suitable habitats in Benin and Togo. Moutouama et al. (2016) studied the impact of climate change on the range of suitable habitats for Haematostaphis barteri Hook.f. in Benin. Idohou et al. (2016) evaluated the potential spatio-temporal range dynamics for eight economically important wild palms under present and future climate regimes across West Africa. Dotchamou et al. (2016) evaluated climate change effects on the spatial distribution of the African locust bean (Parkia biglobosa (Jacq.) R.Br. ex G. Don) for better conservation management in Benin. Adjahossou et al. (2016) assessed the effectiveness of protected areas in Benin for the conservation of favorable priority habitats for the following tree species of socio-economic importance: Afzelia africana Smith ex Pers., Anogeissus leiocarpa (DC) Guill. & Perr., Burkea africana Hook., Daniellia oliveri (Rolfe) Hutch. & Dalziel, Detarium microcarpum Guill. & Perr., Prosopis africana (Guill. & Perr.) Taub. and Khaya senegalensis (Desr.) A.Juss. Gbètoho et al. (2017) used species distribution modeling (SDM) to study pioneer species such as Lonchocarpus sericeus (Poir.) Kunth ex DC. and the African birch (Anogeissus leiocarpa (DC.) Guill. & Perr. in secondary forests of Benin.
Milicia excelsa Welw. C.C. Berg, (Moraceae family) is a large tree 30–50 m in height with a diameter at maturity of 1.7–2.0 m, growing in dense forests and forest galleries as well as savannahs (Ouinsavi and Sokpon 2010). In Benin, M. excelsa is commercially known as “Iroko” and is a sacred species, especially within the villages. It is of economic importance adapted to tropical environments but critically endangered in Benin (Adomou et al. 2010). This study estimates current and future suitable areas for M. excelsa and assesses conservation gaps of the species for better conservation in Benin.
Materials and methods
Study area
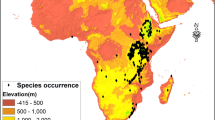
The study was carried out in West Africa with a focus on the Benin Republic located between 6°10′ and 12°50′ N and 1°–3°40′ E (Fig. 1).

Geographic distribution of Milicia excelsa across West Africa
The country’s climatic profile shows two contrasting zones, (Guinean and Sudanian) and a transitional zone (Sudano-Guinean). The Guinean zone (between 6°25′ and 7°30′ N) is characterized by a sub-equatorial climate with four seasons (two rainy and two dry). The rainfall is approximately 1200 mm per year, beginning mid-March to July and a second period from September to November. Temperatures vary between 25 and 29 °C, and relative humidity between 69 and 97%. The Sudanian zone (9°45′–12°25′ N) has a tropical dry climate with two seasons of equal length (rainy and dry). Mean annual rainfall is often less than 1000 mm and occurs mainly from May to September. Relative humidity varies from 18 to 99% and temperatures from 24 to 31 °C. The transitional Sudano-Guinean zone (7°30′ to 9°45′ N) has two rainy seasons merging into a unimodal regime. Annual rainfall fluctuates between 900 and 1110 mm, temperatures are between 25 and 29 °C and relative humidity from 31 to 98%.
Data collection
A total of 1856 records, (georeferenced occurrences), were downloaded from the Global Biodiversity Information Facility portal (GBIF 2016). After cleaning, 209 occurrence points were used for the West African region.
Current (1950–2000) climate data were obtained from WorldClim version 1.4 (available at www.worldclim.org/bioclim). Future climate projections for 2055 were downloaded from AFRICLIM version 3.0 (available at www.york.ac.uk/environment/research/kite/resources/; Platts et al. 2015). For future climatic conditions, predictions from the Interdisciplinary Research on Climate Change (MIROC5) were used. The projections were run under Representative Concentration Pathway (RCP) 4.5 and RCP 8.5 for the 2055-time horizon. By the mid-twenty-first century, RCP 4.5 projects temperatures to rise above industrial levels by at least 1.5 °C in West Africa, with atmospheric CO2 reaching 500 ppm (IPCC 2013). Under the more extreme RCP 8.5 projections, temperatures are predicted to rise by 2.8 °C and atmospheric CO2 to be over 550 ppm (IPCC 2013). RCPs are third generation scenarios and are preferred to the Special Report on Emissions Scenarios (SRES) because they allow more flexibility (and reduced costs) in modeling processes (van Vuuren et al. 2011). RCPs imply collaboration between impacts, adaptation, and vulnerability research, and climate and integrated assessment modeling (van Vuuren and Carter 2014). These scenarios have been developed to explore different combinations of scenario context (demographic, socioeconomic, land use, and technology) (Moss et al. 2010). RCP 4.5 and RCP 8.5 have already been used for studying West African ecosystems (Ayihouenou et al. 2016; Gbètoho et al. 2017). These climate projections were statistically downscaled to match the bioclimatic variables using the delta method (Peterson and Nyari 2008; Ramirez-Villegas and Jarvis 2010).
Modeling of species and model evaluation
The maximum entropy species distribution model algorithm (MaxEnt, version 3.3.3 k) was used for habitat suitability modeling. During the modeling process, occurrence data were cleaned up by removing duplicate records in grids and records which did not correspond to the description of its locality (country) in order to reduce sampling bias which may result from the oversampling of some sites in the study area (Elith et al. 2006).
To optimize model performance, several parameters in MaxEnt were tested. These parameters are regularization multiplier, max number of background points and default prevalence (Fig. 2). MaxEnt models were developed using: regularization of 10,000 background points, regularization multiplier of 0.5, default prevalence of 0.7, and 10 replicates. A jackknife test was performed on the selected bioclimatic variables to determine the ones that best contribute to the model prediction. Owing to common collinearity and non-independence of climate dimensions (Zuur et al. 2010), we examined correlation patterns among variables to select those not closely correlated using environmental niche modeling tools (ENMTs). Hence, we included only a subset of variables with Pearson correlation coefficients below 0.80 (Elith et al. 2010). To avoid geographic bias, a bias surface using a kernel density estimate was generated (Galdino et al. 2016). This resulted in a raster where cells with lower values represented places with lower bias. The bias surface was used to account for sampling intensity and potential sampling bias (Jarnevich et al. 2015). All maps were converted to binary via a conservative least presence thresholding approach consisting of the lowest predicted value (0.405) corresponding to an occurrence record of the species in the calibration data set (Barve et al. 2011). All GIS-related work, (layers clipping and conversion, map composition), was carried out in QGIS 2.18.0.

Fluctuation of AUC in function of regularization multiplier (reg), number of background points (bkpt) and default prevalence (prev)
The performance of the model was assessed using the area under the receiver operating characteristic curve (AUC) (Elith et al. 2006), true skill statistic (TSS) (Allouche et al. 2006; Elith et al. 2006) and Partial ROC (Peterson et al. 2008). The AUC is the probability that a randomly chosen presence point of the species will be ranked as more suitable than a randomly chosen absence point (Elith et al. 2006). A model is considered as having a good fit when its AUC is close to one (AUC ≥ 0.75) (Elith et al. 2006). The TSS is the capacity of the model to accurately detect true presences (sensitivity) and true absences (specificity). A model with TSS ≤ 0 indicates a random prediction, while one with a TSS close to 1 (TSS > 0.5) has a good predictive power (Allouche et al. 2006). For Partial ROC, the evaluation dataset was bootstrapped and probabilities obtained by direct count of the area under the curve (AUC) ratios falling ≤ 1 via a Visual Basic script (https://shiny.conabio.gob.mx:3838/nichetoolb2/), with 100 iterations (Altamiranda-Saavedra et al. 2017). Model performance was assessed by dividing the occurrence data of M. excelsa randomly into calibration (70%) and evaluation (30%) subsets.
Results
Model validation
The correlations analysis and Jackknife AUC test identified five bioclimatic variables as contributing more to the model across the study region: Isothermality, Annual Precipitation, Temperature Annual Range, Mean Diurnal Range, and Precipitation of Wettest Month (Fig. 3).

Contribution of bioclimatic variables to the model
Model evaluations indicated that the model was robust (AUC = 0.979). The TSS value was 0.86 (Fig. 4) and AUC ratios were well above 1.0 (Fig. 5). Therefore, the model showed excellent performance.

Partial AUC distribution

TSS value distribution
Suitable habitats in present and future
The analysis revealed that about 54,009 km2 corresponding to 47.1% of Benin’s surface area (114,763 km2) is currently suitable for the cultivation of M. excelsa (Table 1). M. excelsa is more secondary in the Guinean climatic zone and part of the Sudanian-Guinean and Sudanian climatic zones (Fig. 6). For the RCP 4.5 and RCP 8.5, future projections showed a high decrease in suitable habitat for the species. The Miroc 5 model predicts under the RCP 4.5 a loss of 42.3% of the currently suitable habitats, and about 39.4% under the RCP 8.5 by 2055 (Table 1, Fig. 6). Only a part of the Guinean climatic zone will remain suitable.

Milicia excelsa distribution models in Benin mapped under current and future climatic conditions
The protected areas of Benin illustrate a good potential to conserve actual or current populations of M. excelsa. In fact, many of the protected areas had suitable habitat for M. excelsa in all climatic zones. In the future, according to the two scenarios, only the Lama and Djigbé forests will remain suitable for the conservation of the species.
Discussion
Credibility of the model
In light of global climate changes, it is essential to identify species with high risk of extinction and to diagnose risk driver factors to reverse the decline of biodiversity (Darrah et al. 2017). Information on changes to species ecological niche is important for assessing species geographic range and their realized niche (Breiner et al. 2017). Among several programs for ecological niche modeling and habitat suitability prediction, MaxEnt is one of the most used with respect to our type of data, presence-only data. This modeling tool using presence-only data is one of the best performing algorithms among those using climate modeling approaches (Phillips et al. 2006), and is relatively robust for small sample sizes (Pearson et al. 2007). MaxEnt is a machine learning method that estimates species distribution across a study area by calculating the distribution probability of maximum entropy, subject to the constraint that the expected value of each feature under this estimated distribution should match its empirical average (Phillips et al. 2006). However, MaxEnt cannot indicate the realized niche because of its weaknesses, including uncertainties related to the projections (Elith et al. 2006; Schwartz 2012), failure to account for relevant variables or for geographic barriers (Anderson et al. 2006), bias due to unequal sampling (Fourcade et al. 2014), overfitting, and issues related to evaluation data and methods (Radosavljevic and Anderson 2014). However, the migration of species is a determinant factor in the potential impact of climatic change on their habitats (Elith et al. 2006). When constraints of dispersion are not considered, temperature increase could expand the distribution area of certain species (Sharma and Jackson 2008; Buse and Griebeler 2011). In the current study, indirect variables such as soil and vegetation were not considered. According to Guisan and Zimmermann (2000), direct parameters such as temperature and precipitation are more efficient when the modeling is over large areas. This is contrary to indirect parameters which are not more efficient but are more inclined to introduce errors into the model.
Ecological niche modeling and conservation of M. excelsa
Ecological niche modeling of M. excelsa showed that at present the species is more marginal in the Guinean climatic zone and parts of the Sudanian-Guinean and Sudanian climatic zones. Although the species can be found in dry zones, it reaches its ecological optimum in wetlands, mainly in dense forests and gallery forests (Ouinsavi and Sokpon 2010). In each climatic zone of Benin, at least one protected area is located in a suitable habitat of the species. In the Guinean climatic zone, there are the forests of Lama, Agrimey, Djigbé, and Sakété. In the Sudanian-Guinean climatic zone, there are the forests of Dan, Atchérigbé, Dogo-Ketou, Pénéssoulou, Ouémé Superieur, Boko, N’Dali, Sérou and Sakarou, and in the Sudanian climatic zone, there are the forests of Donga, Kilir, Soubroukou, Birni, Mékrou, and Atchérigbé. Therefore, the protected area network of Benin conserves the species.
In contrast to the present species distribution, future projections show a significant decrease in suitable habitat under RCP 4.5 and RCP 8.5. Only Lama and Djigbé forests retain suitable habitats under both scenarios. This decrease in habitat suitability may be explained by the significant changes projected for bioclimatic parameters, mainly precipitation and temperature. M. excelsa is adapted to temperatures ranging between 25 and 35 °C and precipitation between 1150 and 1900 mm per year (Daïnou et al. 2012). According to Busby et al. (2010), fluctuations in climate variables such as precipitation and temperature will have an impact on biological diversity and on the geographical distribution of suitable habitats. Doxa et al. (2017) suggest that protected areas a principal tool of in situ conservation of biodiversity. Based on future projections, this study shows that few protected areas are key to the conservation of M. excelsa. Therefore, it is important to take the action to conserve effectively M. excelsa by including sacred forests of the Guinean climatic zone in the protected area network. M. excelsa is a sacred tree and is found in many sacred forests.
Conclusions
This study estimated current and future distribution of suitable habitats for M. excelsa so as to assess conservation needs of the species. The suitability prediction will decrease in Benin area under both representative concentration pathway scenarios. At present, M. excelsa is suitable for the Guinean climatic zone and portions of the Sudanian-Guinean and the Sudanian climatic zones. Under future projections, the area of suitable habitat decreases. Among all protected areas, only the Lama and Djigbé reserves will still provide suitable habitats for the species.
Our results provide scientific rationale for planning for and implementing M. excelsa conservation. Despite the considerable potential of M. excelsa and its socio-economic attributes, the species resources are still obtained from the wild. M. excelsa has not been included in various agroforestry and conservation programmes of Benin. Currently, the species is endangered due to over-exploitation, deforestation, numerous human activities and climate change. However, for its sustainable management, forest managers should strengthen the protection of the Lama and Djigbé forests, especially the natural areas, by constructing firewalls and by organizing regular night patrols. Special attention should be given to the sacred forests located in the Guinean climatic zone by making enrichment planting of the species and classifying them as protected area. In addition, the species can be used in agroforestry systems during re-forestation campaigns in the Guinean climatic zone.
Although ecological niche modeling can estimate habitat suitability, it cannot predict species response in term of adaptation, dispersion, migration, or extinction. Thus, suitable habitats inside a protected area does not imply that species will migrate there naturally because the reaction of competing species is not considered. Future research on the conservation of M. excelsa should consider its phenotypic diversity since species conservation implies the conservation of all its genetic diversity.
References
Adjahossou SGC, Gouwakinnou GN, Houehanou DT, Sode AI, Yaoitcha AS, Houinato MRB, Sinsin B (2016) Effectiveness of protected areas for the conservation of favourable and priority habitats for valuable tree species in Benin. Bois et Forets des Tropiques 328(2):67–76
Adomou AC, Sinsin B, Akoégninou AA, van der Maesen J (2010) Plant species and ecosystems with high conservation priority in Benin. In: Burgt X, Maesen J, Onana J-M (eds) Systematics and conservation of african plants. Royal Botanic Gardens, Kew, pp 429–444
Allouche O, Tsoar A, Kadmon R (2006) Assessing the accuracy of species distribution models: prevalence, kappa and true skill statistic (TSS). J Appl Ecol 43:1223–1232
Altamiranda-Saavedra M, Arboleda S, Parra JL, Peterson AT, Correa MM (2017) Potential distribution of mosquito vector species in a primary malaria endemic region of Colombia. PLoS ONE 12(6):e0179093. https://doi.org/10.1371/journal.pone.0179093
Anderson RP, Dudík M, Ferrier S, Guisan A, Hijmans RJ, Huettmann F, Leathwick JR, Lehmann A, Li J, Lohmann LG (2006) Novel methods improve prediction of species distributions from occurrence data. Ecography 29:129–151
Ayihouenou EB, Fandohan AB, Sodé AI, Gouwakinnou NG et Djossa AB (2016) Biogéographie du néré (Parkia biglobosa (Jack.) R. Br. ex. Don.) sous les conditions environnementales actuelles et futures au Bénin. Bulletin de la Recherche Agronomique du Bénin (BRAB) (Agronomie, Société, Environnement & Sécurité Alimentaire), pp 1840−7099
Barve N, Barve V, JimeÂnez-Valverde A, Lira-Noriega A, Maher SP, Peterson AT, Soberón J, Villalobos F (2011) The crucial role of the accessible area in ecological niche modeling and species distribution modeling. Ecol Model 222(11):1810–1819. https://doi.org/10.1016/j.ecolmodel.2011.02.011
Breiner FT, Guisan A, Nobis MP, Bergamini A (2017) Including environmental niche information to improve IUCN red list assessments. Divers Distrib 23:484–495. https://doi.org/10.1111/ddi.12545
Busby JW, Smith TG, White KL, Strange SM (2010) Locating climate insecurity: where are the most vulnerable places in Africa? Austin, TX, USA: University of Texas, The Robert Strauss Center for International Security and Law, Climate Change and African Political Stability (CCAPS) Programme
Buse J, Griebeler EM (2011) Incorporating classified dispersal assumptions in predictive distribution models—a case study with grasshoppers and bush-crickets. Ecol Model 222:2130–2141
Clarke LE, Edmonds JA, Jacoby HD, Pitcher H, Reilly JM, Richels R (2007) Scenarios of greenhouse gas emissions and atmospheric concentrations. Sub-report 2.1a of synthesis and assessment product 2.1. Climate change science program and the subcommittee on global change research, Washington DC
Daïnou K, Doucet JL, Sinsin B, Mahy G (2012) Identity and ecology of Central African timber tree species: the case of Milicia spp. (a review). Biotechnologie Agronomie Société Et Environnement 16(2):229–241
Darrah SE, Bland LM, Bachman SP, Clubbe CP, Trias-Blasi A (2017) Using coarse-scale species distribution data to predict extinction risk in plants. Divers Distrib 23:435–447
Djotan AKG, Aoudji AKN, Yêhouénou Tessi DR, Kakpo SB, Gbètoho JA, Koura K, Ganglo JC (2018) Vulnerability of Khaya senegalensis Desr & Juss to climate change and to the invasion of Hypsipyla robusta Moore in Benin (West Africa). Int J Biol Chem Sci 12(1):24–42. https://doi.org/10.4314/ijbcs.v12i1.3
Dotchamou FT, Atindogbe G, Sode AI, Fonton HN (2016) Density and spatial pattern of Parkia biglobosa under climate change: the case of Benin. J Agric Environ Int Dev 110(1): 173–194. 10.12895/jaeid.20161.447.
Doxa A, Albert CH, Leriche A, Saatkamp A (2017) Prioritizing conservation areas for coastal plant diversity under increasing urbanization. J Environ Manag 201:425–434. https://doi.org/10.1016/j.jenvman.2017.06.021
Drayton B, Primack RB (2012) Success rates for reintroductions of eight perennial plant species after. Soc Ecol Restor 20:299–303
Elith J, Graham CH, Anderson RP, Dudik M, Ferrier S, Guisan A, Hijmans RJ, Huettmann F, Leathwick JR, Lehmann A, Li J, Lohmann LG, Loiselle BA, Manion G, Moritz C, Nakamura M, Nakazawa Y, Overton JM, Peterson AT, Phillips SJ, Richardson K, Scachetti-Pereira R, Schapire RE, Soberon J, Williams S, Wisz MS, Zimmermann NE (2006) Novel methods improve prediction of species’ distributions from occurrence data. Ecography 29:129–151
Elith J, Kearney M, Philips S (2010) The art of modeling range-shifting species. Methods Ecol Evol 1:330–342
Fandohan AB, Moutouama JK, Biaou SSH, Gouwakinnou GN, Adomou CA (2015) Le réseau d’aires protégées Bénin-Togo assure-t-il la conservation de Thunbergia atacorensis (Acanthaceae)? CAMES Science de la vie de la terre et agronomie 3(2):25
Fourcade Y, Engler JO, Rödder D, Secondi J (2014) Mapping species distributions with MAXENT using a geographically biased sample of presence data: A performance assessment of methods for correcting sampling bias. PLoS ONE 9:e97122. https://doi.org/10.1371/journal.pone.0097122
Galdino TVdS, Kumar S, Oliveira LSS, Alfenas AC, Neven LG, Al-Sadi AM, Picanço MC (2016) Mapping global potential risk of mango sudden decline disease caused by Ceratocystis fimbriata. PLoS ONE 11(7):e0159450. https://doi.org/10.1371/journal.pone.0159450
Ganglo JC, Djotan GK, Gbètoho JA, Kakpo SB, Aoudji AKN, Koura K, Yêhouénou Tessi DR (2017) Ecological niche modeling and strategies for the conservation of Dialium guineense Willd (Black velvet) in West Africa. Int J Biodivers Conserv 9(12):373–388. https://doi.org/10.5897/IJBC2017.1151
Ganglo JC, Kakpo SB (2016) Completeness of digital accessible knowledge of the plants of Benin and priorities for future inventory and data discovery. Biodivers Inform 11:23–39
Gbètoho AJ, Aoudji AKN, Roxburgh L, Ganglo JC (2017) Assessing the suitability of pioneer species for secondary forest restoration in Benin in the context of global climate change. Bois et forêts des tropiques 332(2):43–55
GBIF (2016) Occurrence data download. Denmark: global biodiversity information facility. https://doi.org/10.15468/dl.ujt6rx
Godefroid S, Piazza C, Rossi G, Buord S, Stevens AD, Aguraiuja R, Cowell C, Weekley CW, Vogg G, Iriondo JM (2011) How successful are plant species reintroductions? Biol Cons 144:672–682
Guisan A, Zimmermann NE (2000) Predictive habitat distribution models in ecology. Ecol Model 135:147–186
Idohou R, Assogbadjo AE, Glèlè Kakaï R, Peterson AT (2016) Spatio-temporal dynamic of suitable areas for species conservation in West Africa: eight economically important wild palms under present and future climates. Agrofor Syst. https://doi.org/10.1007/s10457-016-9955-6
IPCC (2013) Climate change (2013) The physical science basis. IPCC working group I contribution to the IPCC fifth assessment report. Cambridge University Press, New York, New York, USA.
Jarnevich CS, Stohlgren TJ, Kumar S, Morisette JT, Holcombe TR (2015) Caveats for correlative species distribution modeling. Ecological Informatics 29:6–15
Kakpo SB, Yêhouénou Tessi DR, Gbètoho JA, Aoudji AKN, Ganglo JC (2018) Répartition spatiale de Cola millenii K. Schum., Dialium guineense Wild. et Afzelia africana Smith ex Pers. dans les forêts secondaires du Sud Benin (Afrique de l’Ouest). Int J Biol Chem Sci 12(1):353–362. https://doi.org/10.4314/ijbcs.v12i1.28
Moss RH, Edmonds JA, Hibbard KA, Manning MR, Rose SK, Van Vuuren DP, Carter TR, Emori S, Kainuma M, Kram T, Meehl GA, Mitchell JFB, Nakicenovic N, Riahi K, Smith SJ, Stouffer RJ, Thomson AM, Weyant JP, Wilbanks TJ (2010) The next generation of scenarios for climate change research and assessment. Nature 463:747–756
Moutouama JK, Fandohan AB, Biaou SSH, Amahowe OI, Moutouama FT, Natta AK (2016) Potential climate change favored expansion of a range limited species, Haematostaphis barteri Hook f. J Agric Environ Int Dev 110(2), 397–411. https://doi.org/10.12895/jaeid.20162.516
Ouinsavi C, Sokpon N (2010) Morphological variation and ecological structure of Iroko (Milicia excelsa Welw. C.C. Berg) populations across different Biogeographical Zones in Benin. International Journal of Forestry Research. https://doi.org/10.1155/2010/658396
Pacifici M, Foden WB, Visconti P, Watson JEM, Butchart SHM, Kovacs KM, Scheffers BR, Hole DG, Martin TG, Akcakaya HR, Corlett RT, Huntley B, Bickford D, Carr JA, Hoffmann AA, Midgley GF, Pearce-Kelly P, Pearson RG, Williams SE, Willis SG, Young B, Rondinini C (2015) Assessing species vulnerability to climate change. Nature Climate Change 5:215–225
Pearson RG, Raxworthy CJ, Nakamura M, Peterson AT (2007) Predicting species distributions from small numbers of occurrence records: a test case using cryptic geckos in Madagascar. J Biogeogr 34:102–117
Peterson AT, Nyari AS (2008) Ecological niche conservatism and pleistocene refugia in the thrush-like mourner, Schiffornis sp., in the neotropics. Evolution 62:173–183
Peterson AT, Papes M, Soberon J (2008) Rethinking receiver operating characteristic analysis applications in ecological niche modeling. Ecol Model 2131:63–72
Peterson AT, Soberon J, Pearson RG, Anderson RP, Martínez-Meyer E, Nakamura M, Araújo MB (2011) Ecological niches and geographic distributions. Princeton University Press, Princeton
Phillips SJ, Anderson RP, Schapire RE (2006) Maximum entropy modelling of species geographic distributions. Ecol Model 190:231–259
Pimm SL, Russell GJ, Gittleman JL, Brooks TM (1995) The future of biodiversity. 269(5222):347–350. https://doi.org/10.1126/science.269.5222.347
Platts PJ, Omeny PA, Marchant R (2015) AFRICLIM: high-resolution climate projections for ecological applications in Africa. Afr J Ecol 53:103–108
Questad EJ, Kellner JR, Kinney K, Cordell S, Asner GP, Thaxton J, Diep J, Uowolo A, Brooks S, Inman-Narahari N, Evans SA, Tucker B (2014) Mapping habitat suitability for at-risk plant species and its implications for restoration and reintroduction. Ecological Applications A Publication of the Ecological Society of America 24:385–395
Radosavljevic A, Anderson RP (2014) Making better Maxent models of species distributions: Complexity, overfitting and evaluation. J Biogeogr 41(4):629–643
Ramirez-Villegas J, Jarvis A (2010) Downscaling global circulation model outputs: the delta method decision and policy analysis Working Paper No. 1. J Policy Anal Manag 1:1–18
Ricketts TH, Dinerstein E, Boucher T, Brooks TM, Butchart SHM, Hoffmann M, Lamoreux JF, Morrison J, Parr M, Pilgrim JD, Rodrigues ASL, Sechrest W, Wallace GE, Berlin K, Bielby J, Burgess ND, Church DR, Cox N, Knox D, Loucks C, Luck GW, Master LL, Moore R, Naidoo R, Ridgely R, Schatz GE, Shire G, Strand H, Wettengel W, Wikramanayake E (2005) Pinpointing and preventing imminent extinctions. Proc Natl Acad Sci USA 102:18497–18501
Rovzar C, Gillespie TW, Kawelo K (2016) Landscape to site variations in species distribution models for endangered plants. For Ecol Manag 369:20–28
Schwartz MW (2012) Using niche models with climate projections to inform conservation management decisions. Biol Conserv 155:149–156
Sharma S, Jackson DA (2008) Predicting smallmouth bass (Micropterus dolomieu) occurrence across North America under climate change: a comparison of statistical approaches. Can J Fish Aquat Sci 65:471–481
Thompson K (1997) The soil seed banks of North West Europe: methodology, density and longevity, vol 1. Cambridge University Press, Cambridge
Urban MC (2015) Accelerating extinction risk from climate change. Science 348:571–573
van Vuuren DP, Carter TR (2014) Climate and socio-economic scenarios for climate change research and assessment: reconciling the new with the old. Clim Change 122:415–429
van Vuuren PD, Edmonds J, Kainuma M, Riahi K, Thomson A, Hibbard K, Hurtt GC, Kram T, Krey V, Lamarque JF, Masui T, Meinshausen M, Nakicenovic N, Smith SJ, Rose SK (2011) The representative concentration pathways: an overview. Climatic Change 109:5–31
Zuur AF, Ieno EN, Elphick CS (2010) A protocol for data exploration to avoid common statistical problems. Methods Ecol Evol 1:3–14
Acknowledgements
We extend our profound gratitude to JRS Biodiversity Foundation who supported financially the training of several researchers of this study. We would like to thank Professor Townsend Peterson from Kansas University (USA) and Dr. Lizanne Roxburgh from the Endangered Wildlife Trust (South Africa) for their technical support.
Author information
Authors and Affiliations
Corresponding author
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Project funding: The work was supported partially by JRS Biodiversity Foundation.
The online version is available at http://www.springerlink.com.
Corresponding editor: Tao Xu.
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.
About this article

Cite this article
Kakpo, S.B., Aoudji, A.K.N., Gnanguènon-Guéssè, D. et al. Spatial distribution and impacts of climate change on Milicia excelsa in Benin, West Africa. J. For. Res. 32, 143–150 (2021). https://doi.org/10.1007/s11676-019-01069-7
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11676-019-01069-7