
Abstract
To enhance the sensitivity of an ongoing Genome Wide Association Study (GWAS) for in vitro shoot regeneration and genetic transformation, a wide range of factors that can affect regeneration rate and, therefore, expression of genetic diversity were studied. Included were explant types and sources; direct versus indirect regeneration; nitrogen salts and micronutrient levels in basal medium; sucrose, auxin, and cytokinin types and levels; light intensity and quality; melatonin and serotonin; antibiotics (to exogenously control contaminants); antioxidants; and the ethylene-mitigating agent silver nitrate. Genotype was a statistically significant source of variance in response to nearly every treatment and for every measure of regeneration noted in shoots and roots, and initial generation of callus. Nonetheless, broad-sense heritability estimates varied widely as a result of variable degrees of environmental modification and statistical interactions with genotype. Compared to leaves, stem and petiole explants were highly correlated in their expression of genetic variation in response to treatments, a likely result of their functional similarity. Based on literature review, this study appears to be the most intensive analysis of plant natural genetic variation in response to in vitro manipulations published. It should help to guide development of improved and highly heritable treatments in other plant species that employ organogenic regeneration systems.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
The capacity for regeneration of plant organs or embryos from somatic tissues is the basis of plant genetic engineering methods. However, there is enormous genetic variation in response to regeneration-inducing treatments such as use of plant hormones or tissue wounding (Lardon and Geelen 2020). A small number of studies have determined quantitative genetic parameters for variation in regeneration rate (Lazar et al. 1984; Nestares et al. 2002), and only a handful of studies have attempted to identify the genes underlying natural genetic variation in regeneration rate (Tuskan et al. 2018; reviewed by Lardon et al. 2020). As part of an effort to identify regeneration methods that would elevate regeneration rates and expression of genetic variation among genotypes for a study of GWAS, we sought to quantify broad-sense heritability and interactions of genotype with diverse in vitro treatments and explant types for a sample of wild genotypes of the model tree Populus trichocarpa Torr. and Gray (Tuskan et al. 2006). GWAS studies in the related in vivo traits of shoot and root regeneration were recently published (Nagle et al. 2022a, b). Quantitative analyses that focused on treatment effects when averaged over genotypes are presented in a companion paper (Ma et al. 2022); here we focus on genetic analyses of the same experiments. The results presented are based on 12 different experiments involving 170 treatments, with each experiment using two to 20 genotypes and two to three replicates per treatment, for a total of 42,648 tested explants. We report that due to the great variation in responses of individual explants to treatments, despite strong genetic differences, estimates of broad-sense heritability are only moderate for most regeneration treatments studied.
Materials and Methods
The overall structure of experiments and methods was presented in detail in Ma et al. (2022). In brief, for each experiment up to 20 genotypes of wild P. trichocarpa from the Pacific Northwest and nearby Canada were analyzed (Fig. 1). These trees are part of a GWAS association population assembled by Oak Ridge National Laboratory (bioenergycenter.org/besc/gwas). The collections of shoots harvested from wild trees were established in a coppiced clone bank growing near Corvallis, Oregon; then, shoots from these transplants were rooted and grown in pots in a greenhouse, or maintained by micropropagation in vitro, to provide explants for experiments.

Origin of Populus trichocarpa Torr. and Gray genotypes used in study. Inset at upper right shows most northern and southern genotypes with circle; genotypes are CA-05–01 in California (not included in map), and KTMA-12–4, KTMC-12–1, and DENC-17–2 in British Columbia.
Quantitative Analysis
To visualize genetic effects, boxplots of each trait as a function of genotype are presented for each explant type averaged over treatments for each genotype. Scatter plots of these averages are presented to show the correlation between different traits as well as between different explant types as expressed by each genotype.
The data in each experiment were analyzed with an ANOVA model, including the three experimental factors, genotype, treatment, and explant type. In the model, a treatment sum of squares is given for all factors and their interactions. To quantify the relative importance of the various factors to trait variance, sums of squares were used to calculate the proportion of overall variance explained by each factor and their interactions. Statistical analyses were performed using R with packages as described in detail in Ma et al. (2022).
To gain insight into the replicability of regeneration phenotypes for specific genotypes across experiments that were conducted at different times but with the same explant sources (in vivo or in vitro), subsets of data that utilized the same genotype/treatment/explant type combinations were extracted for analysis and subjected to ANOVA. To help visualize replicability across experiments, line plots showing each phenotype across experiments were produced for each of these combined datasets. The graphical analyses were then summarized by recording the numbers of plots in which there was at least one case of consistency, defined as no rank changes of genotypes between treatments, vs. inconsistency (rank changes), vs. strong inconsistency (major rank changes), as tabulated and explained in Table S1. For example, in supplemental Figure G1 1 (Supplemental Figures, E13 Cross-experiment replication), there are four plots but only the left three are informative about relative rank changes for genotypes, so only those were considered in Table S1 (column called “N useful plots”). The leftmost pair of genotypes and the third from the left (note the two color lines, one almost obscuring the other) show no rank changes, whereas the second from the left shows a rank change that is also considered a “big cross” as tabulated.
Results
A series of 12 different experiments, several subdivided into sub-components that were described in detail earlier (Ma et al. 2022), were conducted and all traits were subjected to analysis of variance, including determination of variance components to evaluate the relative importance of the different sources of variation (Table S2 sheet 1; heat map coloring given in sheet 2). Heritabilities (all were broad-sense, although not always identified as such) were also estimated (Table S3) for each set of treatments where there were multiple genotypes studied. Sheet 1 of Table S3 shows heritability estimates and their statistical significance, and sheet 2 shows heritability estimates with a heat map to help identify treatments that gave the strongest values.
E1: Direct vs. Indirect Regeneration
ANOVA showed that both genotype and treatment had highly significant variance components, and interacted strongly (variance components shown as percentages in Table S2). Heritability estimates for both treatments were also high, ranging from 0.41 to 0.87, showing a large effect of genotype in relation to total variance (Table S3). Expression of genetic variation, partially captured in the heritability estimates, was also substantially increased by the indirect (callus-based) compared to the direct treatments of root and shoot regeneration (Figs. S1–S3 E1). Both explant types (leaf and petiole) also showed statistically significant genotype × environment interactions (Table S2). Under indirect regeneration, the number of genotypes that regenerated callus was unsurprisingly much higher than with direct regeneration, but also the number of genotypes that regenerated shoots was much higher with indirect regeneration (Fig. 2). Callus and shoot regeneration rates were moderately correlated among genotypes (averaged over treatments and explant types), as was root regeneration (though it was highly non-linear due to very low rates in the direct treatment) (Figs. S4–S6 E1). Callus regeneration rates were very weakly correlated between direct and indirect treatments, but shoot regeneration was more highly correlated (r = 0.44 vs. 0.14; Figs. S7 and S8 E1). Unsurprisingly, heritability estimates for callus production were higher with indirect vs. direct regeneration (Fig. S9 E1).

Mean percentage of Populus trichocarpa Torr. and Gray petiole and leaf explants capable of generating callus or shoots in 20 genotypes. Explants generating calluses via (A) indirect regeneration and (B) direct regeneration. Explants generating shoots via (C) indirect and (D) direct regeneration.
E2: Basal Medium Variation
For experiment E2, where a large number of variations to basal medium composition were studied in two genotypes, explant type had the major and highly statistically significant influence on rate of shoot regeneration, whereas genotype had the largest, and statistically significant, influence on rate of shoot regeneration. Heritability was not estimated as only two genotypes were studied, and interactions generally accounted for less than 10% of the variance (Table S2).
E3: Replication to Select Final Basal Media–In Vitro–Derived Explants
Six genotypes were studied for indirect regeneration under the five best basal media identified in experiment E2. Genotypes, explants, and combined interaction terms (including genotype by explant, genotype by treatment, treatment by explant, and genotype, treatment, explant interactions) were the main sources of phenotypic variance (Table S2). Stem explants tended to give the highest rate of regeneration and expression of genetic variation, followed by petioles and then leaves. Root regeneration, on the other hand, was highest and showed the highest proportion of genetic variation in leaves followed by petioles and stems (Table S3). Rates of callus generation were highly variable based on genotype, but had similar levels among all the explant types, though high replicate to replicate variation with leaf explants led to the lowest heritability estimates (Figs. S1–S6 E3a). Shoot regeneration was most strongly correlated among explant types, especially stem and petiole, compared to that for root or callus generation (Figs. S8–S12 E3a). Among pairs of basal media, genotype responses for generation of callus, roots, and shoots were consistently positive and often strongly correlated (above 0.5 and often near to 0.9; Figs. S13–S16 E3a). Heritability estimates tended to be lowest for callus from leaf explants, and highest for root regeneration from the same explant type (Figs. S17–S24 E3a).
In Vivo–Derived Explants
Similar patterns were seen as for in vitro explants, though rates of regeneration were often lower. Genotypes, explants, and combined interaction terms were again the main sources of phenotypic variance (Table S2). Stem explants tended to give the highest rate of shoot regeneration and expression of genetic variation, followed by petioles, and then leaves (Table S3). Root regeneration was again highest and showed the highest proportion of genetic variation in leaves, but stems had comparable levels. Rates of callus generation were again highest in stems, but there was little genetic variation and thus, heritability estimates were zero, whereas petioles and leaves showed great genetic variation and high but variable heritability estimates (Figs. S1–S6 E3b). Correlations among explant types were weaker than for in vitro explants, especially for root regeneration (Figs. S7–S11 E3b). Heritability estimates were again highly variable and imprecise, with standard error bars often overlapping zero (Figs. S18–S23 E3b).
E4: Sucrose Level
The responses of four British Columbia P. trichocarpa genotypes to three sucrose levels in two types of basal media using in vitro–derived explants were studied. As discussed by Ma et al. (2022), there was no evidence of a general benefit from changes in sucrose level. Genotypes were statistically significant and were the major source of phenotypic variance (Table S2). Expression of genetic variation was associated with rates of regeneration, the latter varying widely among explant types but not among sucrose and basal media combinations. Leaf explants had the highest regeneration and expression of genetic variation in root regeneration, and stems had slightly higher rates of callus and shoot generation (Figs. S1–S6 E4). Stems and petioles were most correlated in their regeneration responses, especially for shoots and callus (Figs. S7–S12 E4). Despite the modest differences between treatment and media tested, the correlations among experiments were high for callus and shoot generation traits (Figs. S19–S22 E4). Heritability estimates were modest and generally correlated with rate of regeneration, being highest for stem explants and shoot regeneration, and for leaf explants and root regeneration (Figs. S23–S28 E4).
E5: Auxin Types and Levels
Effects of variation in auxins applied to 6 genotypes (Figs. S1–6 E5ai, E5aii; Table S4) for in vitro explants in two basal media were studied. To anticipate application of these data to GWAS using in vivo–derived explants, responses of 20 genotypes (Figs. S1–6 E5b) were studied, using in vivo explants and the same treatments. Treatments and combined interaction terms were by far the largest sources of phenotypic variance (Table S2). As discussed by Ma et al. (2022), the two auxin types studied, 1-naphthaleneacetic acid (NAA) and 2,4-dichlorophenoxyacetic acid (2,4-D), had distinct effects, with NAA tending to enhance root, callus, and shoot formation from leaves, stems, and petioles, and 2,4-D tending to enhance callus and shoot generation (e.g., Fig. S5 E5b). The large majority of auxin treatments studied gave rise to differential genotypic responses in shoot regeneration (e.g., Fig. 3; Fig. S4 E5b). Correlations among explant types were highest for callus and shoot generation traits compared to root regeneration. This is likely a reflection of the much lower rates of root regeneration (Figs. S7–S12 E5b). Stem to petiole correlations tended to again be stronger than for other explant pairs (e.g., Fig. 4 for callus size). Correlations among treatments were moderate to high within treatments that used the same hormone types (2,4-D or NAA). Conversely, there were low correlations between treatments that used different hormone types (2,4-D vs. NAA; Figs. S13–S18 E5b). Heritability estimates varied widely without clear patterns between explants or treatments (e.g., Figs. S19–S24 E5b).

Differential genotypic response of Populus trichocarpa Torr. and Gray shoot regeneration to auxin treatments. Bar plots shown of (A) number of shoots per stem explant and (B) number of shoots per petiole explant, in relation to varying concentrations in mgL−1 of 1-naphthaleneacetic acid (NAA) and 2,4-dichlorophenoxyacetic acid (2,4-D). Data from in vivo stem and petiole explants in experiment E5.

Correlations among Populus trichocarpa Torr. and Gray explant types for genotypic means of callus size index score per plate from experiment E5b (auxin treatments on in vivo explants). R values were calculated to assess correlations between stem, petiole, and leaf explants. Methods for callus size index scores are referred to in a companion paper (Ma et al. 2022).
E6: Auxin and Cytokinin Combinations
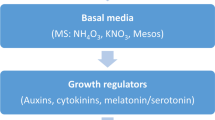
To further identify the best treatments for enhancing expression of genetic variation for GWAS, in vivo–derived explants were used to study a wide variety of auxin-cytokinin combinations. Combined interactions were the largest source of variance, followed by explants and treatments (Table S2). As discussed by Ma et al. (2022), shoot regeneration was generally highest from stem explants, which generally also showed high levels of genetic variation and heritability (Table S3; Fig. S1 E6a). Genotypes and explants responded to the treatments differentially, in relation to both regeneration and necrosis rate (Fig. 5). Stem and petiole explants had the highest correlations in callus and shoot generation responses (Figs. S9 E6a and S10 E6b; root regeneration was zero or very low). Heritability estimates tended to be highest for stem explants (e.g., Figs. S17 and S18 E6a). Some treatments, such as those involving NAA and 2-isopentenyladenine (2iP) (Fig. S4 E6d) or NAA and benzyladenine (BAP) (Fig. S4 E6e), gave very high but nearly identical rates of shoot regeneration per explant among genotypes, thus would be useless for GWAS of this trait. Even modest variations in media composition were important to optimizing expression of genetic variation, with media DB1, DB2, and DB5—but not DB3, DB4, or DB6 (Table S5)—chosen as among the optimal types with respect to suitability for GWAS (Fig. 6).

Examples of Populus trichocarpa Torr. and Gray genotypic variation and genotype × explant (G × E) interactions for shoot regeneration and necrosis in relation to shoot regeneration media and explant type. Data from experiment E6. Explant types in each plate, from top to bottom, are leaf, stem, and petiole. See Table S1, and Table S3 from Ma et al. (2022) for experimental details, medium composition, and mean regeneration rates (see Table S3).

Mean number of Populus trichocarpa Torr. and Gray shoots formed on stem explants averaged over treatments. (A) DB1 basal medium and (B) DB6 basal medium from experiment E6, and (C) heritability estimates and their 95% confidence intervals for six selected media. Shoot induction media tested varied in relative amounts of 2,4-dichlorophenoxyacetic acid (2,4-D) and benzyladenine (BAP); compositions are given in Tables S1 and S3 in Ma et al. (2022). Also see Table S3 from this manuscript.
E7: Melatonin and Serotonin Combinations
Using in vivo–derived explants from 19 genotypes, several concentrations of melatonin and serotonin independently, and then the effect of combined moderate levels in a single medium were tested. As discussed by Ma et al. (2022), there was no evidence of a general benefit from melatonin or serotonin at any levels. Increasing concentration of either chemical clearly reduced the rate and expression of genetic variation in shoot regeneration (Table S3), with a similar but milder effect for callus generation (root regeneration was very low on this medium, so no trend was clear). Heritability estimates were highly variable and imprecise, but also tended to decrease with increasing concentrations (for example, stems and shoot number per explant: Fig. S24 E7).
E8: Antibiotic Treatments
The effects of different concentrations in the fungicide/biocides PPM (Plant Preservative Mixture) and benomyl (benlate) on in vivo–derived explants from 8 genotypes were tested. Experiments under both LED and fluorescent lighting in two different kinds of growth chambers were conducted. Explant type and the sum of interactions were the dominant sources of variance in these experiments, with the three-way genotype × treatment × explant (G × T × E) interaction being especially large; this suggests that different explants from different genotypes had very different reactions to these biocides (Table S2).
LED Conditions
The effects of PPM or benomyl on callus generation or expression of genetic variation were essentially nil (Figs. S1 and S2 E8b). Differential genotype × explant (G × E) responses can be most readily seen for shoot regeneration from stem explants as concentrations of both benomyl and PPM were increased (Fig. S6 E8b). Perhaps as a result of the extensive interactions observed, and the low rates of shoot and root regeneration seen in this experiment, the correlations among explants were lower and more variable than seen for other experimental treatments (Figs. S7–S12 E8b), and estimates of heritability were low, imprecise, and often could not be estimated (Figs. S13–S15 E8b).
Fluorescent Conditions
Similar results were found under fluorescent lighting (Fig. S1 E8a), with little effect of either biocide on callus generation, but there were suppressive effects of high benomyl on stem regeneration from shoots, with no clear effect from PPM. Correlations among explants were again quite low (Figs. S7–S12 E8a), as were heritability estimates, with the exception of shoot regeneration from stem explants, which were above 0.5 for three of the treatments studied (Figs. S13–S15 E8a).
E9: Lipoic Acid
The effects of a wide range of concentrations of lipoic acid (LA) on in vivo–derived explants from 16 genotypes were studied. As discussed by Ma et al. (2022), there was no evidence of a general benefit from lipoic acid (LA) at any level. Explant type and statistical interactions were the major sources of variance (Table S2). As is obvious from the heat maps in Fig. S3 of Ma et al. (2022), callus generation declined as levels of LA increased, as did the expression of genetic variation. This was true for callus from all explant types, though the LA levels at which callus generation declined were highly variable among explants and genotypes (Figs. S1 and S2 E9), as expected from the large interaction variance. Root regeneration from stem explants was low and only declined at high levels of LA and varied among genotypes (Fig. S5 E9). The decline in shoot regeneration varied widely between stem and petiole explant types, with clear variation in genotype sensitivity to LA (Figs. S4 and S6 E9). Correlations between explant types and experimental conditions tended to be low. This was unsurprising given the high interaction variance (Figs. S7–S16 E9). Heritability, where they were estimable, tended to be low and its confidence interval overlapped zero, which was expected given the strong interaction terms.
E10: Activated Charcoal and Ascorbic Acid
Several levels of activated charcoal (AC) and ascorbic acid or vitamin C (VC) were studied in 16 genotypes from in vivo–derived explants. Explant types and combined interaction terms were the largest sources of variance (Table S2).
Activated Charcoal
As discussed by Ma et al. (2022), there was no evidence of a general benefit from activated charcoal (AC) at any level. AC had essentially no effect on rate of callus generation, but had a strong and depressing effect on the rate and genetic variation in shoot regeneration from both petiole and stem explants; root regeneration was hardly observed in this experiment (Figs. S1–S6 E10a). Correlations among explants and experimental treatments, as well as heritability estimates, were again low or inestimable (Figs. S7–S20 E10a).
Vitamin C
As discussed by Ma et al. (2022), there was no evidence of a general benefit from VC at any level. Shoot regeneration was only marginally affected except at the highest level (200 mg L−1), where a slight and genotype-dependent decline was noted, especially for petiole explants (Figs. S1–S6 E10b). The absolute values of correlations among explant types and treatments were again low, but were largely positive. Heritability estimates for callus and shoot regeneration were moderate. The highest and most precise heritability levels were seen for shoot numbers from petioles at high concentrations of VC, mainly a reflection of low variation among replicate plates (Figs. S22 E10b and S6 E10b).
E11: Silver Nitrate
The effects of 7 levels of silver nitrate (SN) on regeneration of 16 genotypes from in vivo–derived explants were studied. As discussed by Ma et al. (2022), there was no evidence of a general benefit from SN at any level (Table S5). The patterns seen were quite complex; explants and combined interactions were again the major sources of variance (Table S2), and the responses of the various explants to increased levels of SN were distinctive and often genotype-specific. Callus size showed no clear trends, though petioles from one genotype seemed to benefit from the highest level (Fig. S1 E11). The proportion of explants forming shoots was strongly affected by SN level and genotype, with different patterns of decline among the three explant types; leaves were most sensitive followed by petioles and then by stems (Fig. S4 E11). Correlations among explant types were again modest, and heritability was also low or inestimable, and highly variable (Figs. S7–S20 E11), with the exception of shoot number for petioles and stems at some SN concentrations (Fig. S20 E11).
E12: Light Intensity and Quality
The effects of varied full-spectrum white light intensity with or without far red (FR) light supplementation on four genotypes of in vivo–derived explants were studied. The light treatments were a minor source of variance; genotype and explant were the major sources along with combined interactions, especially for shoot regeneration traits (Table S2). The use of FR light appeared to depress callus growth on petioles and stems in a genotype-specific manner. The correlations among explants were quite strong for shoot regeneration (Figs. S10 and S12 E12), but much less so among treatments (Figs. S13–S18 E12). Heritability estimates were generally modest and had low precision, with the exception of specific treatments that gave low variation among replicate Petri dishes (for example, shoot number from both petiole and stem explants at 40 µmol m−2 s−1 and zero FR supplementation, which had heritability estimates above 0.9 with very small confidence intervals; Figs. S24 E12 and S6 E12).
Correlations Among Explant Types
A major goal of estimating correlations of regeneration rates among explant types in these experiments was to determine which explant types are most physiologically/developmentally similar, as judged by their responses to in vitro manipulations. Despite the high variability in regeneration responses among explants that was observed, due to the very large number of treatments and genotypes studied, the explant types were assessed with high confidence. Correlations in response to treatments (data averaged over genotypes, as presented in the supplemental figures in Ma et al. 2022), and in genotype responses (averaged over treatments) were studied, as presented in the supplemental figures in this study. Based on correlations among explants in response to different treatments (data averaged over genotypes), two-sided Student’s t tests showed that stem-petiole correlations were significantly higher than stem-leaf correlations (p < 0.0003) or petiole-leaf correlations (p < 0.03), whereas stem-leaf and petiole-leaf correlations did not quite differ significantly (p < 0.07). Thus, stems and petioles are clearly the most similar in their responses with a mean of 0.56 (± 0.03, standard error) compared to means of 0.38 (± 0.03) and 0.48 (± 0.03) for stem-leaf and petiole-leaf correlations, respectively (Fig. 7; Table S5). When correlations among explant responses based on covariation among genotypes (after averaging over treatments) were analyzed, we found similar results. Student’s t tests showed that stem-petiole correlations were significantly higher than stem-leaf correlations, (p < 0.03) though were not quite significantly higher compared to petiole-leaf correlations (p < 0.07); stem-leaf and petiole-leaf correlations showed no evidence of a statistical difference (p < 0.76). Stems and petioles were, again, most similar at 0.51 (± 0.04) compared to means of 0.37 (± 0.05) and 0.39 (± 0.05) for stem-leaf and leaf-petiole correlations, respectively.

Mean correlation coefficients among Populus trichocarpa Torr. and Gray explant types. Correlations for measures of generation of callus (size, percent of explants with callus) and shoots (number, percent of explants with shoots) over experiments E2–E11, calculated from data where genotype results are first averaged (“averaged over genotypes”), or treatment results are averaged and genotype means plotted (“averaged over treatments”). Standard error shown above bars. Based on data in Supplementary Table S5.
Repeatability of Genotypic Responses
Although all the experiments included many pseudo-replicates (explants within dishes) and biological replicates (plates randomized in culture rooms), most were not replicated over time since the main goal of the work was a sequential search for conditions that would maximize expression of genetic variance in relation to environmental variance to enhance the sensitivity of GWAS. Indeed, because these experiments were carried out over nearly 3 yr, and most done with in vivo explants taken from trees grown in a greenhouse at various times of the year and of different ages (number of propagation cycles) and physiological histories (greenhouse light, humidity, and temperature conditions), a high degree of repeatability over time would not be expected. Nonetheless, ANOVA (Table S6) showed that despite high experiment to experiment variance, genotypes were usually a major and significant source of variance, as were the two- and three-way interactions between genotype, treatment, and explant. Residuals (variation among replicate plates) were also a large source of variance due to, in part, the high variance among explants within plates (e.g., Fig. 5).
The graphical analyses (Supplemental Figs., E13 Cross-experiment replication) showed a wide range of responses between experiments, genotypes, and regeneration traits (summarized in Table S1). Rank changes among genotypes in different experiments were not observed in two-thirds (68.5%) of the graphical comparisons examined, supporting general consistency of relative genotypic responses. However, there was evidence of rank changes in one-third of the graphs (32.3%), and major rank changes in about one-fifth of the graphs (18.9%). As examples, mean proportion of shoots show high genotype consistency in nearly all graphical comparisons (e.g., supplemental Figs. G1 12–13, G2 10–12, G3 14 and 15 but not G3 13 and G4 13–15).
Discussion
The expression of genetic variation, and the ratio of genetic to environmental variation (as measured by broad-sense heritability), varied extensively among treatments. Unfortunately, for a species that is difficult to regenerate and transform like P. trichocarpa, an extensive search for high heritability and high regeneration conditions was a prerequisite for GWAS. A brief study suggested that indirect regeneration was far superior to direct regeneration for regeneration of most genotypes (Fig. 2). For this reason, and because of the more common use of indirect vs. direct systems for poplar transformation (Gaur et al. 2016), all subsequent studies were focused on indirect systems. Of course, the best choice of regeneration and transformation system depends on many factors and varies widely among species. For example, a comparison of direct and indirect somatic embryogenesis for biolistic transformation of sugarcane cultivar CP 88–1762 found that the highest transformation efficiency was with the indirect procedure (2.2 lines per shoot) but the most rapid production of transgenic plants was with the direct procedure (12 wk from explant to plants in soil) (Taparia et al. 2012).
The large number of treatments and genotypes studied for three distinct explant types allowed us to examine their similarity in regeneration responses. Results showed that genetic covariation was stronger than treatment-induced environmental covariation, and that stems and petioles were most similar in their regeneration responses, followed by leaves and petioles, and then stems and leaves. These results are likely to be explained by the similarity in function of stems and petioles, both primarily serving in vascular and support functions, and often having abundant vasculature-proximal cells capable of entering regeneration pathways upon induction treatments (Atta et al. 2009; Ikeuchi et al. 2019).
The repeatability of genotypic responses to treatments over time was examined by investigating the statistical significance and consistency of relative performance during the multiple year periods when these experiments were conducted. ANOVA showed that genotypes were usually a major and significant source of variance—demonstrating that their effects were generally consistent. However, it was also found that two- and three-way interactions between genotype, treatment, and explant were large, showing that genotype responses were strongly conditioned by experiment (physiology of explants, specific media and hormone components, sterilization conditions) and developmental state (explant type). Residuals were also large, likely due to the high variation among individual explants within plates and the modest number of explants per plate employed (individual explant often responded differently than those directly adjacent). This variation was also expressed in our analysis of rank changes, for which minor or major changes were seen in about half of our graphical analyses. Thus, although genotypes with varying levels of regenerability could readily be identified, their specific behaviors relative to other genotypes vary widely among experiments. This may also help to explain the common observation that genotypes and regeneration responses vary widely among laboratories even when trying to replicate published experimental conditions (Monthony et al. 2021).
Conclusions
The experiments presented appear to be among the largest and most extensive analysis of genetic variation for in vitro regenerative responses for any plant species. It allowed us to identify conditions that gave high genetic variance for GWAS studies, and will help others to develop conditions to improve regeneration and thus ultimately transformation, especially in dicot species with organogenic regeneration systems. The most highly regenerable genotypes that were identified in this study are as follows: SLMB28-1, SKWB24-2, SKWF24-5, CA-05–01, HOMB21-4, LILB26-5, CHWH27-2. They may be useful to others studying regeneration processes.
The heritability estimates, which vary widely in size among experiments, and the repeatability studies and graphic analyses, showed that while genotypes behaved with reasonable consistency even when studied at very different times, their behavior was also strongly affected by high interactions of genotype with explant and experiment, and high explant-to-explant variation in response. Without seeds or inbred lines to standardize developmental state, this high variance will continue to be a major source of error for organogenic studies in highly heterozygous, woody, and vegetatively propagated plant species.
References
Atta R, Laurens L, Boucheron-Dubuisson E, Guivarc’h A, Carnero E, Giraudat-Pautot V, Rech P, Chriqui D (2009) Pluripotency of Arabidopsis xylem pericycle underlies shoot regeneration from root and hypocotyl explants grown in vitro. Plant J 57:626–644. https://doi.org/10.1111/j.1365-313X.2008.03715.x
Gaur A, Kumar P, Thakur AK, Srivastava DK (2016) Review: In vitro plant regeneration studies and their potential applications in Populus spp.: a review. Isr J Plant Sci 63:77–84. https://doi.org/10.1080/07929978.2015.1076982
Ikeuchi M, Favero DS, Sakamoto Y, Iwase A, Coleman D, Rymen B, Sugimoto K (2019) Molecular mechanisms of plant regeneration. Annu Rev Plant Biol 70:377–406. https://doi.org/10.1146/annurev-arplant-050718-100434
Lardon R, Geelen D (2020) Natural variation in plant pluripotency and regeneration. Plants 9(10):1261. https://doi.org/10.3390/plants9101261
Lardon R, Wijnker E, Keurentjes J, Geelen D (2020) The genetic framework of shoot regeneration in Arabidopsis comprises master regulators and conditional fine-tuning factors. Commun Biol 3:549. https://doi.org/10.1038/s42003-020-01274-9
Lazar MD, Baenziger PS, Schaeffer GW (1984) Combining abilities and heritability of callus formation and plantlet regeneration in wheat (Triticum aestivum L.) anther cultures. Theoret Appl Genetics 68:131–134. https://doi.org/10.1007/BF00252328
Ma C, Goddard A, Peremyslova E, Duan C, Jiang Y, Nagle M, Strauss SH (2022) Factors affecting in vitro regeneration in the model tree Populus trichocarpa I. Medium, environment, and hormone controls on organogenesis. In Vitro Cell Dev Biol - Plant (in press)
Monthony AS, Kyne ST, Grainger CM, Jones AMP (2021) Recalcitrance of Cannabis sativa to de novo regeneration; a multi-genotype replication study. PLoS ONE 16:e0235525. https://doi.org/10.1371/journal.pone.0235525
Nestares G, Zorzoli R, Mroginski L, Picardi L (2002) Heritability of in vitro plant regeneration capacity in sunflower. Plant Breed 121:366–368. https://doi.org/10.1046/j.1439-0523.2002.727109.x
Taparia Y, Gallo M, Altpeter F (2012) Comparison of direct and indirect embryogenesis protocols, biolistic gene transfer and selection parameters for efficient genetic transformation of sugarcane. Plant Cell Tiss Org Cult 111:131–141. https://doi.org/10.1007/s11240-012-0177-y
Tuskan GA, DiFazio S, Jansson S et al (2006) The genome of black cottonwood, Populus trichocarpa (Torr. & Gray). Science 313:1596–1604. https://doi.org/10.1126/science.1128691
Tuskan GA, Mewalal R, Gunter LE, Palla KJ, Carter K, Jacobson DA, Jones PC, Garcia BJ, Weighill DA, Hyatt PD, Yang Y, Zhang J, Reis N, Chen J-G, Muchero W (2018) Defining the genetic components of callus formation: a GWAS approach. PLoS ONE 13:e0202519. https://doi.org/10.1371/journal.pone.0202519
Acknowledgements
We thank the National Science Foundation Plant Genome Research Program (IOS # 1546900) for primary support of this project. We also thank the members of the GREAT TREES (Genetic Research on Engineering and Advanced Transformation of Trees) Research Cooperative at Oregon State University for its long-term investment in our transformation and regeneration studies. We thank Middleton Spectral Vision (Middleton, WI) for their customized imaging system used to capture regeneration data, and high-quality imaging system support. We also thank a number of student workers and technicians for their help on the project, including Brett Pierce, Megan McEldowney, Alyssa Andrews, Julie Kucinski, Kate Dow, Anthony Marroquin, Nathan Dean, Bahiya Zahl, Samantha Shelton, Alexa Nino de Rivera, Hikmat A Bittar, Zachary Haralabatos, Abigail Lawrence, Ali Zahir Ahmed Alsulaimawi, I-Chen Chuang, and Daniel Mitchell.
Author information
Authors and Affiliations
Contributions
Ma and Strauss designed the experiments; Ma and Peremyslova oversaw and executed the experiments and data collection; Nagle and Jiang oversaw and executed the data analysis and graphics; Goddard formatted the graphics and tables; Strauss, Ma, and Goddard wrote the manuscript; all authors edited and approved the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Key Message
As a result of wide exploration of regeneration conditions among diverse genotypes of Populus trichocarpa, treatment combinations that substantially elevated heritability estimates of responses were identified.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Ma, C., Duan, C., Jiang, Y. et al. Factors affecting in vitro regeneration in the model tree Populus trichocarpa: II. Heritability estimates, correlations among explant types, and genetic interactions with treatments among wild genotypes. In Vitro Cell.Dev.Biol.-Plant 58, 853–864 (2022). https://doi.org/10.1007/s11627-022-10302-8
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11627-022-10302-8