
Abstract
The core of solid tumors is characterized by hypoxia and a nutrient-starved microenvironment and has gained much attention as targets of anti-cancer drugs. In the course of search for selective growth inhibitors against the cancer cells adapted to nutrient starvation, epidithiodiketopiperazine DC1149B (1) together with structurally related compounds, trichodermamide A (2) and aspergillazine A (3), were isolated from culture extract of marine-derived Trichoderma lixii. Compounds 1 exhibited potent selective cytotoxic activity against human pancreatic carcinoma PANC-1 cells cultured under glucose-starved conditions with IC50 values of 0.02 µM. The selective index of the compound 1 was found to be 35,500-fold higher for cells cultured under glucose-starved conditions than those under the general culture conditions. The mechanistic analysis indicated that compound 1 inhibited the response of the ER stress signaling. In addition, these effects of compound 1 could be mediated by inhibiting complex II in the mitochondrial electron transport chain.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Solid tumors contain hypoxic and nutrient-starved regions due to the abnormal cell proliferation coupled with the defective and disorganized vascular supply [1]. The cancer cells that have adapted to this tumor microenvironment are assumed to develop aggressive phenotype, impaired angiogenesis, and drug resistance [2, 3]. Since the hypoxic and nutrient-starved tumor microenvironment differs significantly from the normal tissues, the compounds with selective cytotoxicity against the cancer cells in tumor microenvironment would have great therapeutic potential.
The marine flora and fauna are a rich source of therapeutic drugs because of their chemical and biological diversity. Some of the marine-derived compounds have been reported to inhibit the growth of cancer cells adapted to the hypoxic or nutrient-starved conditions. For example, furospinosulin-1 (furanosesterterpene) and dictyoceratins-A and -C (sesquiterpene phenols) isolated from the marine sponge Dactylospongia elegans were shown to exhibit the selective growth inhibitory activities against the hypoxia-adapted human prostate cancer cell line DU145 [4, 5]. Recently, we reported the cytotoxic activities of polybrominated diphenyl ethers, N-methylniphatin A (new 3-alkyl pyridine alkaloid) and biakamides (unique new polyketides) isolated from Indonesian marine sponges of Dysidea sp., Xestospongia sp., and Petrosaspongia sp., respectively, against human pancreatic carcinoma cell line PANC-1 adapted to glucose-deficient growth conditions [6,7,8]. In this study, we present the isolation of epidithiodiketopiperazine DC1149B (1), and structurally related but inactive compounds, trichodermamide A (2) and aspergillazine A (3) isolated from the culture extract of marine-derived Trichoderma lixii., and the cytotoxic activity of compound 1 on nutrient-starved cancer cells, and propose the plausible mechanism of its action.
Materials and methods
Materials
Dulbecco’s Modified Eagle’s medium (DMEM), WST-8 colorimetric reagent, and KCN were purchased from Nacalai Tesque, Inc. (Kyoto, Japan). Fetal bovine serum (FBS) and Dialyzed FBS were purchased from Equitech-Bio Inc. (Kerrville, TX, USA) and Thermo Fisher Scientific Inc. (Waltham, MA, USA), respectively. Anti-Akt, Anti-phosphorylated Akt, anti-GRP78, and anti-β-actin antibodies were obtained from Cell Signaling Technology, Inc. (Danvers, MA, USA). Horseradish peroxidase (HRP)-linked anti-rabbit IgG antibody (GE Healthcare Life Sciences, Buckinghamshire, UK) was used as secondary antibody. Mito Check Complex Activity Assay Kit (Cayman Chemical, Ann Arbor, MI, USA) was used to evaluate the effect of compound 1 on the mitochondrial complex I–V. Oxygen consumption of cells was measured by Oxygen Consumption Rate (OCR) Assay Kit (Cayman Chemical, Ann Arbor, MI, USA). Rotenone, thenoyltrifluoroacetone (TTFA), carbonyl cyanide m-chlorophenylhydrazone (CCCP), antimycin A, and oligomycin A were obtained from Tokyo Chemical Industry Co., LTD. (Tokyo, Japan), Wako Pure Chemical Industries, Ltd. (Osaka, Japan), Sigma-Aldrich (St. Louis, MO, USA), LKT Laboratories, Inc. (St. Paul, MN, USA), and Cayman Chemical (Ann Arbor, MI, USA), respectively. Other chemicals were purchased from Sigma-Aldrich (St. Louis, MO, USA) or Kishida Chemical Co., Ltd. (Osaka, Japan).
Isolation of compounds 1–3
The marine-derived fungus 15G49-1 was isolated from an unidentified marine sponge collected at Mentawai, Indonesia in 2015. The strain was identified as Trichoderma lixii by Techno Suruga Laboratory Co., Ltd. (Shizuoka, Japan) based on the morphology and 5.8S rDNA sequence. The Trichoderma lixii 15G49-1 was cultured in the rice medium (totally 500 g of unpolished rice and 1000 ml of artificial sea water) under static condition at 30 °C for 2 weeks. The culture was extracted by acetone and a mixed organic solvent of acetone/MeOH/EtOAc (4:2:1), followed by combining and evaporating the organic solvents under reduced pressure to obtain a crude extract. The extract was partitioned into a water/EtOAc mixture. The active EtOAc soluble portion (13.6 g) was further partitioned into an n-hexane/90% aq. MeOH mixture. On the guidance of bioassay, the 90% MeOH-soluble portion [8.7 g, IC50 (Glucose Deficient Medium) = 9.0 µg/ml, IC50 (General Glucose Medium) = > 100 µg/ml] was fractionated by silica gel column chromatography [eluted with n-hexane:EtOAc] to obtain seven fractions (Fr.M1 ~ Fr.M7). Among these fractions, the Fr.M3 (1.2 g, eluted with n-hexane:EtOAc = 1:5) showed selective growth inhibition on the PANC-1 cells adapted to glucose starvation [IC50 (Glucose Deficient Medium) = 2.5 µg/ml, IC50 (General Glucose Medium) = > 100 µg/ml]. The active Fr.M3 was then separated by open ODS column chromatography (eluted with MeOH:H2O) to obtain six fractions (Fr.M3-1 ~ Fr.M3-6). The active Fr.M3-3 [289 mg, eluted with MeOH:H2O = 2:3, IC50 (Glucose Deficient Medium) = 0.3 µg/ml, IC50 (General Glucose Medium) = > 100 µg/ml] was further purified by reversed-phase HPLC [Cosmosil 5C18-MS-II (10 mm id × 250 mm); eluted with MeOH:H2O = 1:1] to give DC1149B (1, 39 mg) [9, 10], trichodermamide A (2, 9.5 mg) [11, 12] and aspergillazine A (3) containing fraction (Fr.M3-3-3, 46 mg). The Fr. M3-3-3 was recrystallized with CHCl3/MeOH (3:1) to give aspergillazine A (3, 10.3 mg) [12, 13]. Compounds 1–3 were identified using ESI–TOF–MS and NMR analyses, and comparison with authentic spectral data [9,10,11,12,13].
Cell culture and bioassay
Human pancreatic carcinoma cell line PANC-1 was maintained in the DMEM supplemented with heat-inactivated 10% FBS and kanamycin (50 µg/ml) in a humidified atmosphere with 5% CO2 at 37 °C. To induce nutrient starvation, PANC-1 cells were cultured in the Glucose Deficient Medium [Basal Medium (25 mM HEPES buffer (pH 7.4) supplemented with 6.4 g/l NaCl, 700 mg/l NaHCO3, 400 mg/l KCl, 265 mg/l CaCl2·2H2O, 200 mg/l MgSO4·7H2O, 125 mg/l NaH2PO4, 0.1 mg/l Fe(NO3)·9H2O, 15 mg/l Phenol red, 10 mL/l MEM vitamin solution (X100) (GIBCO, Carlsbad, CA), 200 mmol/l l-glutamine solution (GIBCO, Carlsbad, CA), and 25 mg/l kanamycin) containing 10% dialyzed FBS]. The General Glucose Medium [Basal Medium supplemented with 10% FBS and 2.0 g/l glucose (final 25 mM)] was also used for control cells.
The bioassay was carried out according to a method described previously [6]. Briefly, PANC-1 cells (1 × 104 cells/100 µl in 96 well plastic plate) were pre-incubated in the DMEM supplemented with 10% FBS for 24 h. The medium was then replaced with either the General Glucose Medium or Glucose Deficient Medium to induce cells adaption to the nutrient starvation. After 12-h incubation, the serially diluted samples were added, and the cells were incubated for an additional 12 h in a humidified atmosphere with 5% CO2 at 37 °C. The cell proliferation was detected using the WST-8 colorimetric reagent. The IC50 value was determined by linear interpolation from the growth inhibition curve. We assessed the selectivity of the cytotoxic activity (Selective Index, S.I.) based on the difference in the IC50 values obtained for experiments with General Glucose Media and Glucose Deficient Media.
Western blotting analysis
PANC-1 cells (3 × 105 cells/2 ml in 6-well plastic plate) were pre-incubated in the DMEM supplement with 10% FBS for 24 h. The medium was then replaced with either General Glucose or Glucose Deficient Medium. After 12-h incubation, compound 1 (0.02–0.3 µM) or antimycin A (0.3 nM, positive control) was added to each well and the cells were incubated for an additional 12 h in a humidified atmosphere with 5% CO2 at 37 °C. Subsequently, the cells were rinsed with ice-cold PBS and lysed in the lysis buffer [50 mM Tris–HCl (pH 8.0) containing 150 mM NaCl, 5 mM EDTA, 1% glycerol, 1% NP-40, 1% protease inhibitor cocktail, and 1% phosphatase inhibitor cocktail]. The cell lysate was analyzed by SDS-PAGE and transferred onto PVDF membranes (GE Healthcare Life Sciences Buckinghamshire, UK). The membranes were then incubated with appropriate primary antibodies followed by HRP-conjugated secondary antibodies, and the immunopositive bands were visualized using an ECL kit (GE Healthcare Life Sciences). The luminescent signals were analyzed using an Image Quant LAS4010 Scanner (GE Healthcare Life Sciences).
Effects of compound 1 on the oxygen consumption of PANC-1 cells
Oxygen consumption rate of PANC-1 cells was assayed using OCR Assay Kit (Cayman Chemical, Ann Arbor, MI, USA) as per the manufacturer’s instructions. Briefly, pre-cultured PANC-1 cells (8.0 × 104 cells) in the black, clear bottom 96-well plate (Corning Incorporated, NY, USA) were incubated with General Glucose Medium for 18 h at 37 °C. The cells were replenished with 140 µl of fresh medium, and the test compound was added. This was followed by adding the phosphorescent probe to measure the oxygen consumption. 100 µl of mineral oil was added in each well to restrict oxygen diffusion. The signals were measured by an Infinite M1000 microplate reader (Tecan Group Ltd., Mannedorf, Switzerland) using time-resolved mode at Ex 380 nm and Em 650 nm for 180 min with 1-min time interval. The linear regression was applied after subtracting the blank, and the oxygen consumption rate was indicated by the slope of each signal profile.
Statistical analysis
Data are shown as means ± standard errors of n = 3 independent experiments, and the differences between data sets were assessed by Dunnett’s test. Differences with p values of less than 0.05 were considered significant.
Results and discussion
Cytotoxic activity of compounds 1–3 against the PANC-1 cells cultured under both glucose-deficient conditions and general culture conditions
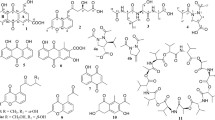
The bioassay-guided separation of the active 90% MeOH-soluble portion from culture extract of marine-derived Trichoderma lixii 15G49-1 led to the isolation of DC1149B (1), trichodermamide A (2), and aspergillazine A (3) (Fig. 1). We then evaluated the cytotoxic activities of compounds 1–3 against the PANC-1 cells cultured under both glucose-deficient and general culture conditions. Antimycin A, which inhibits the growth of PANC-1 cells adapted to the nutrient-starved conditions, was used as the positive control [14].

Chemical structures of compounds 1–3
As shown in Table 1, DC1149B (1) showed a higher cytotoxic activity against the PANC-1 cells adapted to glucose starvation (IC50 = 0.02 µM) when compared with cells cultured with General Glucose Medium (IC50 = 710 µM). The S. I. value of compound 1 was evaluated to be more than 35,000. On the other hand, the cytotoxic activities of trichodermamide A (2) and aspergillazine A (3) were very weak under glucose-starved conditions. These results indicated that compounds 1 had selective cytotoxic activity against the PANC-1 cells adapted to nutrient starvation.
DC1149B (1) was first reported as a secondary metabolite of Trichoderma longibrachiatum and showed weak cytotoxic activity against rat adrenal pheochromocytoma cell line PC12 (IC50 = 21.9 µg/ml, 42.4 µM) and human cervix epithelioid carcinoma HeLa cells (IC50 = 50.1 µg/ml, 97.1 µM) [9]. H. Yamazaki et al. also showed that compound 1 exhibited moderate cytotoxicity on the human leukemic Jurkat cell line with IC50 value of 5.1 µM [10]. For the first time, DC1149B (1) was shown to exhibit the preferential cytotoxic activity against PANC-1 cells adapted to glucose starvation. We next explored the underlying mechanism of DC1149B (1) cytotoxicity.
Effects of compound 1 on the Akt signaling and induction of GRP78
Recent studies on cancer cells adapted to nutrient starvation have revealed that the activation of PI3k/Akt/mTOR signaling pathway and the unfolded protein response (UPR) such as induction of glucose-regulated protein 78 (GRP78) were important for the adaptation of cancer cells to nutrient starvation [15, 16]. Therefore, these processes have attracted much attention as possible drug targets for cancer chemotherapy. This observation prompted us to investigate the effect of compound 1 on the Akt signaling and the induction of GRP78 by utilizing a western blotting method (Fig. 2). Antimycin A was used as a positive control for comparison. The PANC-1 cells cultured in the Glucose Deficient Medium increased the expression levels of phosphorylated Akt and GRP78 proteins compared with cells cultured in the General Glucose Medium (Fig. 2, lanes 1 and 2). Treatment with DC1149B (1) at the lowest test concentration of 0.02 µM decreased the expression of GRP78 in the nutrient-starved cells to the basal level of cells cultured in General Glucose Medium (Fig. 2, lanes 5–10). Also, treatment with DC1149B (1) reduced the levels of Akt and phosphorylated Akt in the nutrient-starved cells in a dose-dependent manner (Fig. 2, lanes 5–10). These results indicated that DC1149B (1) inhibited response of ER stress signaling. Besides, compound 1 exhibited similar effects with antimycin A on the PANC-1 cells adapted to the nutrient-starved conditions (Fig. 2, lanes 3 and 4). The present results, therefore, suggest that compound 1 showed a similar behavior with that of antimycin A against the PANC-1 cells cultured in the Glucose Deficient Medium.

Effects of compound 1 on the Akt signaling and induction of GRP78. Glc (−): Culture in the presence of Glucose Deficient Medium. Glc (+): Culture in presence of General Glucose Medium. The PANC-1 cells cultured in the Glucose Deficient Medium were treated with indicated concentration of compound 1. Cell lysate was resolved by using SDS-PAGE and detected with antibodies against the indicated proteins
Effect of compound 1 on the mitochondrial electron transport chain
Antimycin A is a known inhibitor of complex III in the mitochondrial electron transport chain [17]. Therefore, we next examined the effect of compound 1 on the function of mitochondrial complex I–V using the Mito Check Complex Activity Assay Kit. As shown in Table 2, compound 1 inhibited complex II with IC50 values of 19 µM, while other complexes remained unaffected up to 30 µM. It is known that inhibitors of the mitochondrial electron transport chain affect the oxygen consumption of the treated cells [18]. Since the inhibitory effect of compound 1 on the complex II was weak compared with that on the PANC-1 cells adapted to glucose-starved conditions (Table 1), we explored whether DC1149B (1) inhibited the oxygen consumption on the PANC-1 cells (Fig. 3). As a result, positive control of Antimycin A inhibited the oxygen consumption on the PANC-1 cells cultured in the General Glucose Medium, while the carbonyl cyanide m-chlorophenylhydrazone (CCCP, negative control), which is uncoupling substance of mitochondria, showed the opposite effect to antimycin A [18]. Compound 1 also inhibited the oxygen consumption on the PANC-1 cells under general culture conditions (Fig. 3). This result indicated that compound 1 inhibits complex II in the mitochondrial electron transport chain. We also evaluated the effect of compound 1 on the oxygen consumption of PANC-1 cells cultured in the Glucose Deficient Medium. However, the oxygen consumption rate of PANC-1 cells could not be determined, as the oxygen consumption of PANC-1 cells in the Glucose Deficient Medium was significantly lower than that in the General Glucose Medium. Collectively, our results indicate that the selective cytotoxicity of compound 1 might be mediated by the inhibition of complex II in the mitochondrial electron transport chain, while further study is necessary to consider whether complex II is major target for DC1149B (1) as a selective inhibitor against cancer cells adapted to glucose-starved conditions.

Effect of compound 1 on the oxygen consumption of PANC-1 cells. Pre-cultured PANC-1 cells (8.0 × 104 cells) in the 96-well plate was incubated in General Glucose Medium for 12 h at 37 °C. The medium was then replaced with each fresh medium, and the test compound was added followed by adding the phosphorescent probe to measure the oxygen consumption. The signals were measured by a Tecan infinite M1000 using time-resolved mode. Differences were considered significant at *p < 0.05
Conclusions
In our search for inhibitors against the cancer cells adapted to nutrient starvation, the epidithiodiketopiperazine DC1149B (1) was isolated from culture extract of marine-derived Trichoderma lixii. DC1149B (1) showed potent selective cytotoxicity against the cancer cells adapted to the glucose-starved conditions, with a S.I. value of 35,500. This study also explains the previously unknown mechanism of action of compound 1 on the cancer cells. Our results revealed that compound 1 inhibited the ER stress signaling, and that these effects of compound 1 could be mediated by the inhibition of complex II in the mitochondrial electron transport chain. Currently, research is underway to synthesize a probe to identify the major target of compound 1 in PANC-1 cells adapted to nutrient-starved conditions, and to observe the distribution of compound 1 in PANC-1 cells.
Change history
11 March 2021
A Correction to this paper has been published: https://doi.org/10.1007/s11418-021-01499-w
References
Vaupel P, Kallinowski F, Okunieff P (1989) Blood flow, oxygen and nutrient supply, and metabolic microenvironment of human tumors: a review. Cancer Res 49:6449–6465
Rohwer N, Cramer T (2011) Hypoxia-mediated drug resistance: novel insights on the functional interaction of HIFs and cell death pathways. Drug Resist Update 14:191–201
Chen S, Rehman SK, Zhang W, Wen A, Yao L, Zhang J (2010) Autophagy is a therapeutic target in anticancer drug resistance. Biochim Biophys Acta 1806:220–229
Arai M, Kawachi T, Kotoku N, Nakata C, Kamada H, Tsunoda S, Tsutsumi Y, Endo H, Inoue M, Sato H, Kobayashi M (2016) Furospinosulin-1, marine spongean furanosesterterpene, suppresses the growth of hypoxia-adapted cancer cells by binding to transcriptional regulators p54nrb and LEDGF/p75. ChemBioChem 17:181–189
Kawachi T, Tanaka S, Fukuda A, Sumii Y, Setiawan A, Kotoku N, Kobayashi M, Arai M (2019) Target identification of the marine natural products dictyoceratin-A and -C as selective growth inhibitors in cancer cells adapted to hypoxic environments. Mar Drugs 17:E163
Arai M, Shin D, Kamiya K, Ishida R, Setiawan A, Kotoku N, Kobayashi M (2017) Marine spongean polybrominated diphenyl ethers, selective growth inhibitors against the cancer cells adapted to glucose starvation, inhibits mitochondrial complex II. J Nat Med 71:44–49
Arai M, Kamiya K, Shin D, Matsumoto H, Hisa T, Setiawan A, Kotoku N, Kobayashi M (2016) N-methylniphatyne A, a new 3-alkylpyridine alkaloid as an inhibitor of the cancer cells adapted to nutrient starvation, from an Indonesian marine sponge of Xestospongia sp. Chem Pharm Bull 64:766–771
Kotoku N, Ishida R, Matsumoto H, Arai M, Tida K, Setiawan A, Muraoka M, Kobayashi M (2017) Biakamides A–D, unique polyketides from a marine sponge, act as selective growth inhibitors of tumor cells adapted to nutrient starvation. J Org Chem 82:1705–1718
Nakano H, Hara M, Meshiro T, Ando K, Saito Y, Morimoto S (1990) Japan Patent Kokai 1990–218686
Yamazaki H, Takahashi O, Murakami K, Namikoshi M (2015) Induced production of a new unprecedented epitrithiodiketopiperazine, chlorotrithiobrevamide, by a culture of the marine-derived Trichoderma cf. brevicompactum with dimethyl sulfoxide. Tetrahedron Lett 56:6262–6265
Garo E, Starks CM, Jensen PR, Fenical W, Lobkovsky E, Clardy J (2003) Trichodermamides A and B, cytotoxic modified dipeptides from the marine-derived fungus Trichoderma virens. J Nat Prod 66:423–426
Liu R, Gu QQ, Zhu WM, Cui CB, Fan GT (2005) Trichodermamide A and aspergillazine A, two cytotoxic modified dipeptides from a marine-derived fungus Spicaria elegans. Arch Pharmacol Res 28:1042–1046
Capon RJ, Ratnayake R, Stewart M, Lacey E, Tennant S, Gill JH (2005) Aspergillazines A–E: novel heterocyclic dipeptides from an Australian strain of Aspergillus unilateralis. Org Biomol Chem 3:123–129
Momose I, Ohba S, Tatsuda D, Kawada M, Masuda T, Tsujiuchi G, Yamori T, Esumi H, Ikeda D (2010) Mitochondrial inhibitors show preferential cytotoxicity to human pancreatic cancer PANC-1 cells under glucose-deprived conditions. Biochem Biophys Res Commun 392:460–466
Jin S, DiPaola RS, Mathew R, White E (2007) Metabolic catastrophe as a means to cancer cell death. J Cell Sci 120:379–383
Lee AS (2007) GRP78 Induction in cancer: therapeutic and prognostic implications. Cancer Res 67:3496–3499
Xia D, Yu CA, Kim H, Xia JZ, Kachurin AM, Zhang L, Yu L, Deisenhofer J (1997) Crystal structure of the cytochrome bc1 complex from bovine heart mitochondria. Science 277:60–66
Weatherly LM, Shim J, Hashmi HN, Kennedy RH, Hess ST, Gosse JA (2016) Antimicrobial agent triclosan is a proton ionophore uncoupler of mitochondria in living rat and humanmast cells and in primary human keratinocytes. J Appl Toxicol 36:777–789
Acknowledgements
The human pancreatic carcinoma cell line, PANC-1 (RCB2095), was provided by the RIKEN BRC through the National Bio-Resource Project of the MEXT, Japan. The authors are grateful to Mr. Izumi Kimura, School of Pharmaceutical Sciences, Osaka University, for technical assistance. This research was funded by Platform Project for Supporting Drug Discovery and Life Science Research (Basis for Supporting Innovative Drug Discovery and Life Science Research [BINDS]) from AMED (Grant No. JP19am0101084), Kobayashi International Scholarship Foundation, and a Grant-in-Aid for Scientific Research B (Grant Nos. 18H02096 and 17H04645) from JSPS to MA.
Author information
Authors and Affiliations
Corresponding author
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
The original online version of this article was revised due to retrospective open access order.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Tang, R., Kimishima, A., Ishida, R. et al. Selective cytotoxicity of epidithiodiketopiperazine DC1149B, produced by marine-derived Trichoderma lixii on the cancer cells adapted to glucose starvation. J Nat Med 74, 153–158 (2020). https://doi.org/10.1007/s11418-019-01357-w
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11418-019-01357-w