
Abstract
Purpose
This study deals with the identification of CH4 and CO2 sources in the high-carbonate wetland Lake Podpeč in the Ljubljana Marshes, Slovenia.
Materials and methods
Lake Podpeč is situated on the periphery of the Ljubljana Marshes in central Slovenia. A combination of chemical analysis and natural abundance analysis of stable carbon and hydrogen isotopes of CH4, along with analysis of dissolved inorganic carbon (DIC), was employed in an incubation experiment.
Results and discussion
The isotopic composition of dissolved inorganic carbon (δ13CDIC) suggests three main processes occurring during incubation: oxic degradation of organic matter (OM), anoxic OM degradation, and methanogenesis. During oxic degradation of OM, the δ13CDIC values slightly decrease from − 13.2 to − 14.5‰. However, after 50 days, the δ13CDIC values started to increase, reaching − 12.2‰ by the end of the experiment. 13C enrichment coincided with the formation of CH4, which began to increase simultaneously. The CH4 produced had an average δ13CCH4 value of − 67 ± 1‰ and δ2HCH4 value of − 389 ± 3‰, suggesting that CH4 is formed through acetate fermentation. The contribution of calcite dissolution to DIC increased during the degradation of OM by 53%. However, during methanogenesis, there was no significant change in the concentrations of Ca, and the estimated contribution to DIC was only 3%.
Conclusions
This study enhances our understanding of methane production in wetland Lake Podpeč and its relevance in the context of other high-carbonate lakes. The findings offer insights into the complex interactions between OM degradation, methane production pathways, and carbonate dissolution, which has implications for the global carbon cycle and greenhouse gas emissions.
Similar content being viewed by others


Explore related subjects
Find the latest articles, discoveries, and news in related topics.Avoid common mistakes on your manuscript.
1 Introduction
Methane (CH4) is a potent greenhouse gas that plays a significant role in the Earth’s climate system. Natural sources of CH4 include wetlands (102–179 Tg CH4 year−1), freshwater systems (117–212 Tg CH4 year−1), oceans (9–22 Tg CH4 year−1), fauna (4–18 Tg CH4 year−1), and other land sources (13–54 Tg CH4 year−1), each of which will respond differently to a changing climate (Saunois et al. 2016, 2020). Biogenic CH4, produced through the anaerobic decay of organic matter (OM), accounts for approximately one-third of global CH4 production. In lakes, a significant fraction of biogenically produced CH4 can accumulate in the anoxic part of the water column. However, the CH4 can be rapidly emitted to the upper water column and ultimately to the atmosphere through ebullitive transport, plant-mediated flux, or storage flux via advective processes, particularly during periods of seasonal lake mixing (Borrel et al. 2011; Lehmann et al. 2015). Therefore, it is essential to acknowledge that significant uncertainties accompany current CH4 budgets and emission estimates, and one of the most substantial knowledge gaps revolves around the rates of aquatic CH4 formation and release (Rosentreter et al. 2021).
The production of CO2 in wetland lakes is another critical aspect of their role in the carbon cycle. As wetlands are characterized by high organic content, low oxygen levels, and high-water table, they become conducive environments for both CH4 and CO2 production. Unlike other wetlands, carbonate wetland lakes such as Lake Podpeč are rich in calcium and magnesium carbonate minerals, which affect their water chemistry, alkalinity, and pH levels. According to Marcé et al. (2015), 57% of the Earth’s surface area is occupied by lakes and reservoirs, particularly in tropical and temperate latitudes that exhibit higher alkalinity, which influences CO2 production. The primary sources of CO2 production in carbonate wetland lakes include biological and geochemical processes. Biological sources stem from the decomposition of organic matter, such as plants and algae, by microbial activity. Organic matter accumulates in the sediments and serves as a nutrient source for methanogens, which produce methane as an end-product of their metabolic processes. Simultaneously, the decomposition of organic matter also results in the release of CO2. On the other hand, geochemical sources arise from the interaction of lake water with the underlying carbonate bedrock, leading to the dissolution of carbonates and subsequent release of CO2 into the water. Thus, understanding the factors controlling CH4 and CO2 turnover rates is vital, especially considering the significant contributions of aquatic sources and wetlands to greenhouse gas emissions (Dean et al. 2018).
Stable isotopes of CH4 and dissolved inorganic carbon (DIC) can provide valuable information on the origin and biogeochemical processes of CH4 and CO2 in lake sediments. DIC comprises carbon dioxide (CO2), bicarbonate ions (HCO3−) and carbonate ions (CO32−) and can have distinct isotopic compositions depending on the sources and processes involved. During organic degradation in lake sediments, microbial respiration produces CO2, which is enriched in the heavier carbon isotope, 13C. As a result, the remaining DIC pool in the water column becomes relatively depleted in 13C. Calcite dissolution, particularly in carbonate-rich lakes, can release CO2 into the water, decreasing the content of 13C in DIC due to the preferential release of 12C-enriched CO2. The process of methanogenesis often involves the incorporation of 12C-enriched carbon from organic matter into CH4, resulting in DIC with a higher 13C content.
Further carbon isotopes (δ13C-CH4) can be used to distinguish between different sources of CH4, such as biogenic and thermogenic CH4. Both acetotrophic (Eq. 1) and hydrogenotrophic (Eq. 2) methanogens (Conrad 2005) can be found in freshwater lakes, marine environments, and sediments.
The δ13C-CH4 values ranged from − 65 up to − 30‰ if acetate fermentation is a predominant methanogenic pathway in sediments (Whiticar 1999), while δ13C-CH4 values of methane resulting from the CO2 reduction are lower ranging between − 110 and − 60‰ (Whiticar 1999). Hydrogen isotopes (δ2H-CH4) can provide insights into the environmental conditions under which the CH4 was produced. In lake sediments, δ2H-CH4 values can be influenced by the sources of hydrogen, such as water or OM, as well as the isotopic fractionation that occurs during the production and consumption of CH4. For example, CH4 production via CO2 reduction typically results in higher δ2H-CH4 values (typically from − 250 to − 150‰), while CH4 production via acetate fermentation or other pathways can result in lower δ2H-CH4 values, i.e., − 400 to − 300‰ (Whiticar 1999).
This study aimed to investigate and characterize the sources and biogeochemical processes of CH4 and CO2 production in the high-carbonate wetland Lake Podpeč in the Ljubljana Marshes, Slovenia. The primary objectives were to identify the dominant pathways of methane production by analyzing stable carbon isotopes (δ13C-CH4) and hydrogen isotopes (δ2H-CH4) of CH4 in lake sediments. Additionally, the study contributed to understanding the sources of CO2 production in the carbonate wetland lake by examining the isotopic composition of DIC and distinguishing between biological and geochemical sources. By investigating these isotopic signatures, this research aimed to understand the underlying environmental conditions and microbial processes responsible for methane and carbon dioxide emissions in Lake Podpeč. Thus, it contributes to a deeper understanding of greenhouse gas dynamics in this critical high-carbonate wetland ecosystem.
2 Materials and methods
2.1 Study site
This study was conducted in Lake Podpeč, located at the southern edge of Ljubljana marsh, under the Northern slopes of Krim hills (45° 58′ 7.63″ N, 14° 25′ 58.04″ E; Fig. 1). The lake is a small, circular, karstic lake with a longer diameter of 135 m, a shorter diameter of 120 m, and a surface area of approximately 1.2 ha. A depth of up to 51 m makes this lake one of the deepest lakes in Slovenia. Water drains to the lake from seven karst springs from the white limestone and dolomite hillsides at the southeastern side of the plain. The lake waters fill the northwestern part of an 800-m-long and 400-m-wide depression, which is stretched in the Dinaric direction (NW–SE). This depression is filled with thick layers of alluvium and separated from the Ljubljana Marsh basin by a low rock barrier. The peat and alluvium deposits surrounding the lake are thick, most likely reaching the rock base more than 10 m below. The water level in the lake remains consistently at the same height, although it occasionally floods the surrounding swampy plain. In winter, the temperature of the lake remains above 7 °C, while in summer, it can reach up to 20 °C. The entire lake, along with its surrounding marsh plain, is protected due to the presence of a unique flora and fauna that are greatly threatened by human activities. These include a variety of river mussels and fish (pike, perch, chub, and catfish). The organic carbon (OC) concentration is high (51.1 wt%) in the upper 16 cm of the surrounding marsh soil. The C/N ratio was determined to be 49.1, indicating a predominantly terrestrial plant origin of OM, while δ13COC of the soil was − 26.1‰ (Andrič per. com).

Location of Lake Podpeč (wetland lake, Ljubljana Marshes), Slovenia
2.2 Sediment cores and pore water extraction
Sediment cores were collected in the anoxic hypolimnion in the deepest part of the lake in November 2005 (Fig. 1). The bottom water temperature was 9.5 °C. Once collected, the sediment cores were promptly transported to the laboratory for pore water extraction and subsequent geochemical analysis. Additionally, the overlaying water, approximately 5 cm in depth, was collected for further examination. Sediment cores were extruded and sectioned in 1-, 2-, or 4-cm intervals in an N2-filled glove bag. Pore water was extracted by centrifugation at 5000 rpm and filtered through 0.45-µm Millipore HA membrane filters inside the glove bag. The extracted pore water was then analyzed for various parameters, including pH, total alkalinity (TA), isotopic composition of dissolved inorganic carbon (δ13CDIC), and cation analysis (Ca, Mg, Fe, Mn). The sectioned samples were freeze-dried and finely ground into powder form for the determination of δ13C organic carbon (δ13COC) and carbonates (δ13CCa).
2.3 Incubation experiment
In November 2005, during autumn, sediment samples for the incubation experiment were collected following the methodology described in Hall et al. (1989). A SCUBA diver inserted an acrylic chamber (25-cm height, 30-cm ID, 0.6-cm wall thickness) into the sediment, with an acrylic plate attached to the bottom of the chamber and the top hole sealed underwater. The chamber was then carefully transported to the laboratory and stored in the dark in a refrigerator for 111 days at a temperature (9.5 °C) similar to that at the bottom of the lake during sampling.
At the start of incubation, the water above the sediment was replaced with water collected from the bottom of the lake. The sediment depth inside the chamber was 10 cm, with an overlying water volume of 10 L. Periodic (daily) samples were taken using a syringe for chemical and isotopic analysis. The fluxes of dissolved species across the sediment–water interface were determined by linear regression of the solute concentration variation with incubation time, and the fluxes were adjusted for dilution.
2.4 Analyses
2.4.1 Sample collection
Sample aliquots of pore water and incubation experiment collected for chemical analysis were passed through a 0.45-µm nylon filter into bottles and refrigerated until analyzed. Samples for cation (pre-treated with HNO3) and total alkalinity analyses were collected in HDPE bottles. Samples for δ13CDIC analyses were stored in glass bottles filled to leave no headspace. The pH was determined using an Orion™ 3-star pH meter with Orion™ 8102BNUWP ROSS Ultra™ pH combination electrode (Thermo Fisher Scientific Inc., USA) and the concentration of dissolved oxygen using a YSI model 58 m and a YSI 5239 DO probe with high sensitivity membranes.
2.4.2 Chemical analysis
Total alkalinity (TA) was determined on refrigerated water samples using weak hydrochloric acid titration using a Mettler Toledo DL15 auto-titrator, and the data were reanalyzed using the Gran alkalinity method (Stumm and Morgan 1996). Analytical uncertainty for alkalinity was estimated to be 10%. Ten milliliters of water was used to determine the concentration of CH4. The samples were injected into N2-flushed ampoules (20 ml) and treated in an ultrasound bath at 70 °C for 20 min. Headspace gas samples were directly analyzed by gas chromatography (HP 5840 A GC/FID) with an analytical error of ± 5%.
Major cations (Ca, Mg, Fe, Mn—precision ± 2%) in water samples were determined using a PerkinElmer inductively coupled plasma optical emission spectrometry (ICP-OES, DV5300) . The precision of the measurements was ± 3%. Saturation indices (SIs) of calcite and dolomite were calculated from TA, pH, and temperature using the PHREEQC for the Windows program (Appelo and Postma 2005).
2.4.3 Stable isotope analysis
The samples for determining the stable isotopic composition of CH4 (δ13CCH4) were transferred to glass septum tubes previously purged with pure He. After treatment in an ultrasound bath at 70 °C, the isotopic composition was determined directly from the headspace by an Isotope Ratio Mass Spectrometer (IRMS) (Europa Scientific 20–20) with an ANCA-TG preparation module for trace gas samples equipped with a Gilson autosampler. The isotopic composition of H in CH4, δ2HCH4, was measured in the GCA—geochemical laboratory in Iltnu, Germany. The analyses were performed with a GC-C-Finningan Mat Delta plus isotope mass spectrometer. The components were separated on a Poraplot Q column (ID 0.32 mm, 25-m length). The hydrocarbons were reduced to H2 in a reduction oven at 1470 °C (Dumke et al. 1989).
The gas evolution method was used for isotope analyses of dissolved inorganic carbon (δ13CDIC) in water (Atekwana and Krishnamurthy 1998). Briefly, phosphoric acid (105%) was added (200 µl) into glass septum tubes (Exetainer Septum Tubes, Labco Limited, UK) and purged with helium. The water (5 ml) samples were introduced using a syringe, and the acid–water reaction began immediately upon injection. The isotope ratio of CO2 was determined directly from the headspace gas using a continuous-flow IRMS Europa 20–20 with ANCA TG trace gas separation module. The optimal extraction procedure was checked using standard solutions with Na2CO3 with known δ13C values of − 10.8 ± 0.2‰ and − 4.12 ± 0.2‰, respectively.
Sediment samples for the determination of δ13COC were first treated with 1 M HCl, and an aliquot of dry sample was wrapped in a tin capsule and analyzed after combustion in an O2 atmosphere in a quartz reactor. The C isotopic composition of the CO2 produced was determined using a Europa 20–20 ANCA-SL mass spectrometer. The method’s reproducibility was checked using IAEA-CH-7 polyethylene and NBS22 oil as reference materials. The δ13CCa values were then obtained using a standard method based on the reaction with phosphoric acid. Samples were prepared by grinding into a powder in an agate mortar; 2 mg of the sample was then placed into ampoules, which were capped, flushed with helium, and then reacted with anhydrous H3PO4 for 1 day at 25 °C. The released CO2 was then measured using IRMS with an ANCA-TG preparation module for trace gas samples. NBS 18 and NBS 19 were used as reference materials. All stable isotope results are reported with the conventional delta (δ) notation in per mil (‰) using the general formula (Brand et al. 2014).Footnote 1 Reproducibility of the measurements was ± 0.1‰ for δ13CCa, ± 0.2‰ for δ13CDIC and δ13COC, ± 0.5‰ for δ13CCH4, and ± 3 ‰ for δ2HCH4.
2.5 Calculations
A carbon isotope mass balance calculation was employed to understand better the CO2 and CH4 production. The total DIC flux is formed from the remineralization of OM (Jrem) and the carbonate dissolution flux (JCa) during the oxic degradation of OM (Boehme et al. 1996; Ogrinc et al. 2008):
Consequently, the isotope mass balance equation could be written in the form:
δ13Ctot,ox is the isotopic composition of the total flux during the oxic phase of incubation, δ13CJ-DIC is the isotopic composition of the DIC flux, while δ13Cca is the mean isotope value of carbonate of − 8.2‰. The CH4 flux (JCH4) also contributes to the general isotope mass balance during methanogenesis. Therefore, Eqs. 3 and 4 could be written in the form:
The isotopic composition of DIC (δ13CJ-DIC) and the CH4 (δ13CJ-CH4) fluxes were calculated using the analogous approach applied in other studies (Jahnke et al. 1997; Ogrinc et al. 2002, 2008). The δ13CJ-DIC value was determined for different periods from the slope of the regression line when ([DIC]ti / [DIC]t0) − 1 was plotted against δ13CDIC,i × ([DIC]ti / [DIC]t0), where [DIC]ti is the concentration of DIC at time “i” and [DIC]t0 represents the concentration of DIC at the beginning of the experiment. δ13CDIC,i is the measured δ13CDIC at time ti. Similarly, δ13CJ-CH4 can be calculated from the slope of the line when ([CH4]ti / [CH4]t0) − 1 is plotted vs. δ13CCH4,i × ([CH4]ti / [CH4]t0). Here, [CH4]ti is the concentration of CH4 determined at time i, while [CH4]t0 is the initial concentration of CH4 determined after 50 days of incubation. δ13CCH4 is the measured isotopic composition during the incubation. The unknown isotopic value of degradable sedimentary OM (δ13Crem) was then calculated from these estimates.
3 Results
3.1 Sediment and pore water
The pore water pH decreased from 7.8 to 6.7 at a depth of 18 cm. At the same time, the levels of TA, Ca, and Mg remained constant within the depth profile with an average value of 4.08 ± 0.11 mmol L−1, 1.10 ± 0.14 mmol L−1, and 0.64 ± 0.25 mmol L−1, respectively (Fig. 2). In contrast, δ13CDIC values displayed a high positive gradient from − 11.7 to 3.7‰ from the bottom water layer to the sediment depth of 18 cm, which is typical of methanogenic sediments. Also, Mn decreased from 3.82 to 0.55 μmol L−1, while Fe increased from 5.18 to 27.5 μmol L−1, indicating reducing conditions in the pore water profile (Fig. 2). The study observed low sulfate concentration typical for lake environments with an average value of 0.86 ± 0.20 mmol L−1 . The sediments exhibit an average δ13COC value of − 29.7 ± 0.6‰ and a low δ13CCa average value of − 8.2 ± 0.4‰.

Depth profiles of total alkalinity (TA),isotopic composition of dissolved inorganic carbon (δ13CDIC), Ca, Mg, Mn, and Fe in pore water of Lake Podpeč
3.2 Incubation experiment
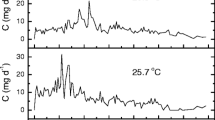
The temporal variations of the studied solutes during incubation are presented in Figs. 3 and 4, while the estimation of the isotopic composition of DIC (δ13CJ-DIC) and CH4 (δ13CJ-CH4) fluxes is presented in Fig. 5. During incubation, linear changes in the concentrations of various electron acceptors and total alkalinity (TA) were observed with time. The regression lines (Figs. 3 and 4) represent the basis for calculating the benthic fluxes of solutes presented in Table 1. Dissolved O2 was present in the incubation chamber for 11 days (Fig. 3) and was related to the oxic degradation of OM. Increases of soluble Mn and Fe were observed during the anoxic phase, present for 50 days, while during methanogenesis, concentrations of Mn decreased and Fe slightly increased (Fig. 3). The levels of Ca and Mg during incubation (Fig. 4) increased in oxic phase due to the dissolution of carbonates also indicated by the calculated saturation indices (SIs) of calcite and dolomite. In parallel, an increase in TA occurred. In the anoxic phase, the levels of Ca and Mg remained relatively stable and TA only slightly increased. After 50 days, CH4 levels began to increase, and the end products of methanogenesis were 13C- and 2H-depleted CH4 and 13C-enriched DIC. The average δ13CCH4 and δ2HCH4 values of CH4 at the end of the experiment were − 67 ± 1‰ and − 389 ± 3‰, respectively.

Concentrations of dissolved oxygen, Mn, and Fe as a function of time in the laboratory-incubated flux chamber experiment in the dark in November 2005. Solid lines represent the linear regression equation used to estimate benthic fluxes for three different periods (i.e., oxic degradation of organic matter, anoxic degradation (“transition”), and methanogenesis) separated by different symbols

Concentrations of Ca, Mg, TA, CH4, δ13CDIC, and δ13CCH4 as a function of time in the incubation experiment performed in November 2005. Solid lines represent the linear regression equations used to estimate benthic fluxes for three different periods (i.e., oxic degradation of organic matter, anoxic degradation (“transition”), and methanogenesis) separated by different symbols

a Determination of the isotopic composition of the CH4 flux (δ13CJ-CH4). [CH4] = concentration of methane during methane formation at t0 and at time ti; δ13CCH4,i = measured δ13C value of methane at time ti; b determination of the isotopic composition of the DIC flux (δ13CJ-DIC). [DIC] = concentration of DIC; t0 and ti represent the time of incubation, where t0 represents the beginning of incubation; δ13CDIC,i = measured δ13C value of DIC at time ti
4 Discussion
The incubation experiment provides valuable information on the temporal dynamics of solutes and isotopic signatures during different phases of biological and geochemical processes (Figs. 3 and 4). Methanogenesis produced CH4 with δ13CCH4 and δ2HCH4 values corresponding predominantly to acetotrophic methanogenesis (Whiticar 1999). The data are comparable to the latest studies on boreal lakes and wetlands (Thompson et al. 2016; Schenk et al. 2021). However, in the study performed by Schenk et al. (2021), isotopic ratios of CH4 in deeper sediments were consistent with mixing/transition between CH4 production pathways, indicating a higher contribution of the CO2 reduction pathway. The study further indicates that the isotopic composition of CH4 sources is consistently higher in littoral sediments than in deep waters across boreal and subarctic lakes. They explain these differences by the variability in organic matter substrates across depths. However, the predominant pathway should be verified using labeled compounds or selective inhibitors (Conrad 2005). In addition, the δ2HCH4 values could not be interpreted conclusively without considering the δ2HCH4 values of coexisting H2O since it was found that the δ2HCH4 value depends on the δ2H values in water in sulfate-poor freshwater environments (Waldron et al. 1999; Wand et al. 2006; Douglas et al. 2021). It was estimated that δ2H–H2O explain approximately 42% of the observed variation in δ2HCH4 globally.
The acetotrophic methanogenesis in Lake Podpeč is further supported by the previous microcosm experiments on soil slurries in the Ljubljana Marshes (Jerman et al. 2009, 2017). In those experiments, Methanosarcinaceae were detected in samples with high CH4 production. The results also showed the most prolonged lag phase for CH4 production in anaerobic soil slurries, suggesting that OM is extensively processed (refractory) and that only a small proportion can be readily mineralized by microorganisms (Stres et al. 2008; Jerman et al. 2009). In addition, iron oxides help to form soil aggregates with OM, reducing the contact between microorganisms and the organic substrate and lowering the rate of its degradation (Roden and Wetzel 2003). In our study, the CH4 production began after 50 days of incubation and can be related to the poorly degradable OM and the presence of only a small portion of easily accessible OM in the sediment. Unfortunately, the concentrations of OC and TN in sediments were not determined. However, we assume that the OC and C/N ratio is similar to the surrounding soil. The concentration of OC in the upper 16 cm of the marsh soil is high, amounting to 51.1 wt% with a C/N ratio of 49.1, indicating a predominantly plant-derived origin of the OM.
Further, a carbon isotope mass balance calculation was employed to understand better organic matter (OM) degradation, carbonate dissolution, and CH4 production (Eqs. 3–6). Table 2 presents the calculated fluxes and the isotopic value of degradable sedimentary OM (δ13Crem) for all three periods. Changes in JDIC flux were observed during the three different processes and are associated with the consumption of electron acceptors during the degradation of the OM. It is seen that the contribution of carbonate dissolution to DIC changed during incubation. Initially, 53% of the DIC was derived from carbonate dissolution, while the contribution was only 3% during methanogenesis. The δ13Crem of − 28.1‰ during oxic degradation is comparable with the δ13COC value of − 29.7 ± 0.6‰ determined in sediments, while during anoxic conditions, the δ13Crem of − 18.1‰ is higher, indicating that other OM sources contribute to the C pool. A relatively low δ13Crem value of − 41.2‰ was determined during methanogenesis. There are two possible explanations. The first is related to the oxidation of 13C‐depleted biogenic methane that can contribute to 13C‐depleted dissolved CO2 to the dissolved inorganic carbon (DIC) reservoir. The assimilation of this DIC would lead to the production of 13C-depleted biomass. Alternatively, 13C‐depleted biogenic methane formed in the sediment could enable the expansion of methanotrophic organisms that produced 13C‐depleted biomass (Hinrichs et al. 1999; Orphan et al. 2001; Hollander and Smith 2001).
Drawing comparisons with other impacted high-carbonate lakes, such as the eutrophic subalpine Lake Bled (Ogrinc et al. 2002; Mandič-Mulec et al. 2012) and high-mountain Lake Planina (Ogrinc et al. 2008), provides valuable insights into the eutrophication processes and the sources of greenhouse gases on a global scale. In both lacustrine environments, hydrogenotrophic methanogenesis emerges as the dominant process responsible for CH4 production. The average δ13CCH4 value for CH4 produced in Lake Bled is approximately − 69.5 ± 1.2‰, which closely aligns with the values obtained in Lake Planina with δ13CCH4 value of − 70.1 ± 1.1‰ and Lake Podpeč − 67 ± 1‰. However, the difference in δ2HCH4 determined in Lake Planina compared to Lake Podpeč, i.e., − 208 ± 10‰ vs − 389 ± 3‰, respectively, shows that different processes are responsible for methanogenesis. Unfortunately, it was not possible to measure the δ2HCH4 in Lake Bled; however, the archaeal community supports the hypothesis that hydrogenotrophic methanogenesis is the dominant pathway in the sediment of Lake Bled (Mandič-Mulec et al. 2012). It is also interesting to note that the CH4 flux in Lake Bled was twice as high, averaging 2.2 mmol m−2 day−1, compared to Lake Planina’s JCH4 of 1.1 mmol m−2 day−1 and nine times higher compared to JCH4 obtained in Lake Podpeč (Table 1). These data indicate that in both eutrophic lakes, OM remineralization occurs faster than in Lake Podpeč and is due to the presence of fresh and highly degradable OM in Lake Bled and Lake Planina sediments primarily originating from microalgae and deposited phytoplankton. Further, it was also observed that methanogenesis inhibits calcite dissolution in all three carbonate-rich systems. For example, the contribution of calcite dissolution to DIC decreased during incubation, i.e., from 20 to 7% in Lake Planina (Ogrinc et al. 2008) and from 53 to 3% in Lake Podpeč.
5 Conclusions
This study focused on CH4 and CO2 production in Lake Podpeč, a high-carbonate wetland lake in Ljubljana Marshes, Slovenia. The investigation involved a laboratory-based incubation experiment conducted at in situ temperatures, covering the transition from oxic to anoxic conditions. The experiment revealed three distinct processes: oxic degradation of organic matter (OM), anoxic degradation of OM, and methanogenesis. Further, the CH4 produced had a measured average δ13CCH4 value of − 67 ± 1‰ and δ2HCH4 value of − 389 ± 3‰, indicating that, contrary to alpine lakes, CH4 is formed through acetate fermentation. Moreover, based on carbon isotope mass balance calculations, different biological and geochemical processes influencing the DIC production were evaluated. For instance, OM degradation contributed around 50% to DIC, while methanogenesis contributed about 45%. Interestingly, the contribution of carbonate dissolution to DIC changed significantly during the incubation, from 53 to 3%. This finding highlights the dynamic nature of these geochemical processes and implies that methanogenesis inhibits calcite dissolution.
Overall, this study provides valuable insights into mechanisms of CH4 and CO2 production in high-carbonate wetland lakes providing information for further research on similar ecosystems and their role in the global carbon cycle.
Data Availability
The data that support the findings of this study are available from the corresponding author, NO, upon reasonable request.
Notes
δiE = (R(iE/jE)sample / R(iE/.jE)standard) – 1.
where E is the element (H, C) and R is the isotope ratio between the heavier “i” and the lighter “j” isotope (2H/1H, 13C/12C) in the sample and relevant internationally recognized reference standard. The delta values are multiplied by 1000 and expressed in units “per mil” (‰). For hydrogen the Vienna Standard Mean Ocean Water (VSMOW), while for carbon the Vienna Pee Dee Belemnite (VPDB) are used as a reference standard.
References
Appelo CAJ, Postma D (2005) Geochemistry, Groundwater and Pollution. Rotterdam: A. A. Balkema. 536
Atekwana E, Krishnamurthy R (1998) Seasonal variations of dissolved inorganic carbon and δ13C of surface waters: application of a modified gas evolution technique. J Hydrol 205:265–278
Boehme SE, Blair NE, Chanton JP, Martens CS (1996) A mass balance of 13C and 12C in an organic-rich methane-producing marine sediment. Geochim Cosmochim Acta 60:3835–3848
Brand WA, Coplen TB, Vogl J, Rosner M, Prohaska T (2014) Assessment of international reference materials for isotope-ratio analysis (IUPAC Technical Report). Pure Appl Chem 86:425–467
Borrel G, Jézéquel D, Biderre-Petit C, Morel-Desrosiers N, Morel JP, Peyret P, Fonty G, Lehours AC (2011) Production and consumption of methane in freshwater lake ecosystems. Res Microbiol 162(9):832–847. https://doi.org/10.1016/j.resmic.2011.06.004
Conrad R (2005) Quantification of methanogenic pathways using stable carbon isotopic signatures: a review and a proposal. Org Geochem 36(5):739–752
Dean JF, Middelburg JJ, Röckmann T, Aerts R, Blauw LG, Egger M, Jetten MSM, de Jong AEE, Meisel OH, Rasigraf O, Slomp CP, in’t Zandt MH, Dolman AJ, (2018) Methane feedbacks to the global climate system in a warmer world. Rev Geophy 56(1):207–250. https://doi.org/10.1002/2017RG000559
Douglas PMJ, Stratigopoulos E, Park S, Phan D (2021) Geographic variability in freshwater methane hydrogen isotope ratios and its implications for global isotopic source signatures. Biogeosci 18:3505–3527. https://doi.org/10.5194/bg-18-3505-2021
Dumke I, Faber E, Poggenburg J (1989) Determination of stable carbon and hydrogen isotopes of light hydrocarbons. Anal Chem 61:2149–2154
Hall POJ, Anderson LG, Rutgers van der Loeff MM, Sundby B, Westerlund SFG (1989) Oxygen uptake kinetics in the benthic boundary layer. Limnol Oceanogr 34:734–774
Hinrichs K-U, DeLong EF, Hayes JM, Sylva SP, Brewer PG (1999) Methane-consuming archaebacteria in marine sediments. Nature 398:802–805
Hollander DJ, Smith MA (2001) Microbially mediated carbon cycling as a control on the δ13C of sedimentary carbon in eutrophic Lake Mendota (USA): new models for interpreting isotopic excursions in the sedimentary record. Geochim Cosmochim Acta 65(23):4321–4337
Jahnke RA, Craven DB, McCorkle DC, Reimers CE (1997) CaCO3 dissolution in California continental margin sediments: the influence of organic matter remineralization. Geochim Cosmochim Acta 61:3587–3604
Jerman V, Metje M, Mandić-Mulec I, Frenzel P (2009) Wetland restoration and methanogenesis: the activity of microbial populations and competition for substrates at different temperatures. Biogeosci 6:1127–1138
Jerman V, Danevčič T, Mandić-Mulec I (2017) Methane cycling in a drained wetland soil profile. J Soils Sediments 17:1874–1882. https://doi.org/10.1007/s11368-016-1648-2
Lehmann MF, Simona M, Wyss S, Blees J, Frame CH, Niemann H, Veronesi M, Zopfi J (2015) Powering up the “biogeochemical engine”: the impact of exceptional ventilation of a deep meromictic lake on the lacustrine redox, nutrient, and methane balances. Front Earth Sci 3:1–13. https://doi.org/10.3389/feart.2015.00045
Mandić-Mulec I, Gorenc K, Gams Petrišič M, Faganeli J, Ogrinc N (2012) Methanogenesis pathways in a stratified eutrophic alpine lake (Lake Bled, Slovenia). Limnol Oceanogr 57:868–880
Marcé R, Obrador B, Morguí JA et al (2015) Carbonate weathering as a driver of CO2 supersaturation in lakes. Nat Geosci 8:107–111. https://doi.org/10.1038/ngeo2341
Ogrinc N, Lojen S, Faganeli J (2002) A mass balance of carbon stable isotopes in an organic-rich methane-producing lacustrine sediment (Lake Bled, Slovenia). Glob Plan Chan 33:57–72
Ogrinc N, Žagar M, Faganeli J, Kanduč T, Vreča P (2008) Methane formation in a remote mountain lake (Lake Planina, NW Slovenia). Geomicrobiol J 25:346–356
Orphan VJ, House CH, Hinrichs KU, McKeegan KD, DeLong EF (2001) Methane-consuming archaea revealed by directly coupling isotopic and phylogenetic analysis. Science 293:484–487
Roden EE, Wetzel RG (2003) Competition between Fe(III)-reducing and methanogenic bacteria for acetate in iron-rich freshwater sediments. Microbial Ecol 45:252–258
Rosentreter JA, Borges AV, Deemer BR et al (2021) Half of global methane emissions come from highly variable aquatic ecosystem sources. Nat Geosci 14:225–230. https://doi.org/10.1038/s41561-021-00715-2
Saunois M, Bousquet P, Poulter B, Peregon A, Ciais P, Canadell JG et al (2016) The global methane budget 2000–2012. Earth Syst Sci Data 8(2):697–751. https://doi.org/10.5194/essd-8-697-2016
Saunois M, Stavert AR, Poulter B, Bousquet P, Canadell JG et al (2020) The global methane budget 2000–2017. Earth Syst Sci Data 12:1561–1623. https://doi.org/10.5194/essd-12-1561-2020
Schenk J, Sawakuchi HO, Sieczko AK, Pajala G, Rudberg D, Hagberg E, Fors K, Laudon H, Karlsson J and Bastviken D (2021) Methane in lakes: variability in stable carbon isotopic composition and the potential importance of groundwater input. Front Earth Sci 9:722215. https://doi.org/10.3389/feart.2021.722215
Stres B, Danevčič T, Pal L, Mrkonjić Fuka M, Resman L, Leskovec S, Hacin J, Stopar D, Mahne I, Mandić-Mulec I (2008) Influence of temperature and soil water content on bacterial, archaeal and denitrifying microbial communities in drained fen grassland soil microcosms. FEMS Microbiol Ecol 66:110–122
Stumm W, Morgan JJ (1996) Aquatic chemistry: chemical equilibria and rates in natural waters, 3rd edn. Wiley, New York
Thompson HA, White JR, Pratt LM, Sauer PE (2016) Spatial variation in flux, δ13C and δ2H of methane in a small arctic lake with fringing wetland in Western Greenland. Biogeochem 131:17–33. https://doi.org/10.1007/s10533-016-0261-1
Waldron S, Lansdown JM, Scott EM, Fallick AE, Hall AJ (1999) The global influence of the hydrogen isotope composition of water on that of bacteriogenic methane from shallow freshwater environments. Geochim Cosmochim Acta 63:2237–2245
Wand U, Samarkin VA, Nitzsche H-M, Hubberten H-W (2006) Biogeochemistry of methane in the permanently ice-covered Lake Untersee, central Dronning Maud Land, East Antarctica. Limnol Oceanogr 51:1180–1194
Whiticar MJ (1999) Carbon and hydrogen isotope systematics of bacterial formation and oxidation of methane. Chem Geol 161:291–293
Acknowledgements
This research was conducted in the framework of program activity P1-0143: Cycling of nutrients and contaminants in the environment, mass balances and modeling of environmental processes and risk analysis funded by the Slovenian Research Agency (ARRS).
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare no competing interests.
Additional information
Responsible editor: Philip N. Owens
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Ogrinc, N., Šegedin, U. & Faganeli, J. Methane and CO2 production in the wetland Lake Podpeč (Slovenia). J Soils Sediments 23, 4163–4172 (2023). https://doi.org/10.1007/s11368-023-03622-8
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11368-023-03622-8