
Abstract
The Caenorhabditis Intervention Testing Program (CITP) is an NIH-funded research consortium of investigators who conduct analyses at three independent sites to identify chemical interventions that reproducibly promote health and lifespan in a robust manner. The founding principle of the CITP is that compounds with positive effects across a genetically diverse panel of Caenorhabditis species and strains are likely engaging conserved biochemical pathways to exert their effects. As such, interventions that are broadly efficacious might be considered prominent compounds for translation for pre-clinical research and human clinical applications. Here, we report results generated using a recently streamlined pipeline approach for the evaluation of the effects of chemical compounds on lifespan and health. We studied five compounds previously shown to extend C. elegans lifespan or thought to promote mammalian health: 17α-estradiol, acarbose, green tea extract, nordihydroguaiaretic acid, and rapamycin. We found that green tea extract and nordihydroguaiaretic acid extend Caenorhabditis lifespan in a species-specific manner. Additionally, these two antioxidants conferred assay-specific effects in some studies—for example, decreasing survival for certain genetic backgrounds in manual survival assays in contrast with extended lifespan as assayed using automated C. elegans Lifespan Machines. We also observed that GTE and NDGA impact on older adult mobility capacity is dependent on genetic background, and that GTE reduces oxidative stress resistance in some Caenorhabditis strains. Overall, our analysis of the five compounds supports the general idea that genetic background and assay type can influence lifespan and health effects of compounds, and underscores that lifespan and health can be uncoupled by chemical interventions.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Elaboration of clinical strategies that delay health declines associated with aging is anticipated to markedly improve life quality for the elderly and their families. One research area focused on such a goal seeks to identify compound interventions that can prolong life and/or promote healthy aging. Toward this end, model organisms are invaluable in initial compound screening as these models offer low expense, ease of culture, short lifespans, and often simple health assays. One of the most widely studied aging models, Caenorhabditis elegans, has proven a useful system for identifying and characterizing compounds that robustly extend lifespan and healthspan [1,2,3].
The Caenorhabditis Intervention Testing Program (CITP), an NIH-funded research consortium consisting of investigators at three independent sites (Rutgers University, the University of Oregon, and the Buck Institute for Research on Aging), is tasked to identify pharmacological interventions with the potential to extend Caenorhabditis lifespan and healthspan in a robust manner. The founding principle for the CITP is that compounds with positive effects across a genetically diverse population engage conserved biochemical pathways that promote healthy aging. The CITP is distinctive in testing compounds across a genetically diverse panel of Caenorhabditis strains and species to identify interventions that promote lifespan extension independent of genetic background [4]. Another distinctive feature of the CITP effort is that studies are replicated as closely as possible at the three geographically distinct sites. To date, the CITP has reported on the lifespan and healthspan effects of more than 27 compounds in more than 250,000 individuals across nearly 280 trials [5,6,7,8,9,10].
The initial CITP studies focused on longevity as the sole endpoint for anti-aging intervention evaluation [5,6,7,8]. For the current suite of test compounds, we expanded and modified our workflow [5] to evaluate the potential of compounds to extend lifespan and/or healthspan across Caenorhabditis species and strains (Supplemental Fig. 1).
Here, we report results from five compounds in the CITP testing pipeline: 17α-estradiol, acarbose, green tea extract (GTE), nordihydroguaiaretic acid (NDGA), and rapamycin. 17α-estradiol, a weak endogenous steroidal estrogen, has been reported to alleviate age-related metabolic dysfunction and inflammation in male mice [11], to protect against neuro-degeneration in cell and animal models of Parkinson’s disease [12], and to extend lifespan in genetically heterogenous male mice [13, 14]. Acarbose, an anti-diabetic drug that inhibits alpha-glucosidase, has been shown to prevent age-related glucose intolerance [15] and to limit postprandial hyperglycemia in mice [16, 17]. Likewise, acarbose has been shown to extend median lifespan in genetically heterogenous mice [13, 14]. GTE is rich in antioxidant polyphenols, and has been reported to reduce the risk of coronary heart disease and certain forms of cancer [18], as well as to provide neuroprotection against diseases such as Alzheimer’s [19]. GTE increases lifespan in both flies [20, 21] and mice [22], and its primary constituent flavonoid, epigallocatechin gallate (EGCG), has been shown to extend mean lifespan in C. elegans [23], although that result may be context dependent [24, 25]. NDGA, a lignin found in the creosote bush, possesses both antioxidant and anti-inflammatory properties [26, 27], and has been shown to increase lifespan in mice [13, 28]. Finally, rapamycin, an mTOR kinase inhibitor, has been shown to increase lifespan in a variety of model organisms, including mice and flies [29,30,31].
Our studies of 17α-estradiol, acarbose, GTE, NDGA, and rapamycin underscore the complexities of assessing biological outcomes of candidate lifespan extending treatments. Using our standardized protocols, we find that the antioxidants GTE and NDGA extend Caenorhabditis lifespan in a species-specific manner. GTE and NDGA tests also revealed some assay-specific outcomes—in certain genetic backgrounds we found decreased survival in manual longevity assays, whereas we measured extended lifespan when we determined outcomes using the automated C. elegans Lifespan Machines (ALM). GTE and NDGA affected swimming ability in a strain-specific manner, and GTE lowered oxidative stress resistance in some Caenorhabditis strains. Lifespan and healthspan could thus be uncoupled as evaluated by our approach, with outcomes influenced by genetic background. Overall, our findings on this test set of interventions underscore how impactful genetic background, selected health assay, and protocol details are in the assessment of intervention effects. Interventions that meet the high bar of efficacy across a broad range of genetic backgrounds and across multiple experimental approaches may prove the exception but would establish definitive priority for testing in mammalian models.
Materials and methods
Strains
All natural isolates used were obtained from the Caenorhabditis Genetics Center (CGC) at the University of Minnesota: C. elegans N2, MY16, and JU775; C. briggsae AF16, ED3092, and HK104; C. tropicalis JU1630, JU1383, and QG834. Worms were maintained at 20 °C on 60 mm NGM plates seeded with Escherichia coli OP50-1.
Interventions
Compound intervention treatments were performed as previously described [5]. The compounds used include green tea extract (LKT Laboratories, Inc. G6817, lot #2595901), nordihydroguaiaretic acid (Sigma-Aldrich 74540, lot #BCBQ4489V), α-estradiol (Sigma-Aldrich E8750, lot #016M4175V), rapamycin (LC laboratories R-5000, lot #ASW-135), and acarbose (Sigma-Aldrich A8980, lot #MKBS1059V0). Compounds were obtained as solids and dissolved in either water or DMSO (dimethyl sulfoxide) to obtain stock solutions, with either water or DMSO used to treat control agar plates. DMSO stock solutions (both compound and control) were then further diluted with water to create working solutions to allow for even distribution across the agar plate while maintaining a final concentration of 0.25% DMSO. Agar plates were treated with compound stock solutions such that the final volume was assumed equal to the volume of the agar.
Manual lifespan assay
Per the previously published CITP standard operating procedure [5], synchronized populations were generated via timed egg lays on 60 mm NGM plates. At day one of adulthood, 50 worms were transferred to 35 mm NGM plates containing 51 µM FUdR and compound intervention (or the solvent control). Worms were then transferred to fresh plates and scored as alive or dead on day two and five of adulthood for C. elegans and C. briggsae, or day two and four of adulthood for C. tropicalis. Thereafter, worms were transferred once weekly and scored every Monday, Wednesday, and Friday until dead. Death was defined as a lack of response when stimulated with a platinum wire.
Automated lifespan assay
ALM assays were performed as previously described [6, 32, 33], based on modification of the protocols published for the Lifespan Machine [34]. Briefly, worms were age synchronized and transferred to intervention plates as described above. One week post egg lay, 50 animals were transferred to 50 mm tight-lidded, intervention treated, modified NGM plates containing 51 µM FUdR and 100 µM nystatin and loaded onto the ALMs. Scanner data was collected and analyzed using the Lifespan Machine software (https://github.com/nstroustrup/lifespan; [34] and strain-specific posture files [6]).
Swimming ability assay
Swimming ability was measured per the standard CITP protocol [35, 36], using the C. elegans Swim Test system (CeleST) [37, 38]. Worms were age-synchronized and exposed to compound intervention during adulthood as described above, until swimming measurements were collected at ages 6 and 12 of adulthood (C. elegans and C. tropicalis), or ages 8 and 16 of adulthood (C. briggsae). Videos were processed using the CeleST software (https://github.com/DCS-LCSR/CeleST).
Thermotolerance assay
The ability for animals to withstand heat stress was measured as previously published [35], utilizing a modification of the ALM protocol. Worms were synchronized and aged as adults on compound intervention plates, as stated above, until the desired testing age (adult day 6 and 12 for C. elegans and C. tropicalis, days 8 and 16 for C. briggsae). At this time, animals were placed onto 50 mm plates with modified NGM containing 100 µM nystatin, without FUdR or compound intervention, at a density of 70 worms per plate. Plates without lids were then transferred to Automated Lifespan Machines in an incubator set to 32 °C and 50% humidity. Scanner data were collected using an increased scan speed and reduced resolution to provide proper temporal resolution, needed as a result of the shortened survival under these conditions. Images analyzed using the Lifespan Machine software (https://github.com/nstroustrup/lifespan) [34] and strain-specific posture files [6] and deaths were validated by hand.
Oxidative stress resistance assay
Resistance to oxidative stress was measured per the standard CITP procedure [35]. Worms were prepared and aged as adults on interventions as mentioned above, with the same ages tested as in the swim test and thermotolerance assays. At the desired age, animals were transferred at a density of 70 worms per plate to 50 mm tight-lidded plates with modified NGM containing 40 mM paraquat (or methyl viologen dichloride, from Sigma-Aldrich), 51 µM FUdR, 100 µM nystatin. Plates were then transferred to ALMs at 20 °C and scanner data collection, processing, and analyzing were done with the same methodology mentioned for the thermotolerance assay.
Results
The CITP pipeline for the evaluation of compounds on lifespan and healthspan
We created a workflow plan to evaluate the ability of a chemical intervention to extend lifespan and/or healthspan across Caenorhabditis species and strains (Supplemental Fig. 1). The CITP pipeline involves preliminary testing in a single lab to identify a potentially effective dosage range (phase one study; Supplemental Fig. 2) and to target a bioactive dose across strains (phase two; Supplemental Fig. 3) before we undertake survival assays in all three CITP labs (phase three; Fig. 1). In phase four, we evaluate the effect of bioactive compounds that passed the preliminary tests for robust impact on longevity (phases 1–3) on select health measures (Figs. 2 and 3). We publish our findings on compounds without positive lifespan that exit the pipeline [7,8,9] (Supplemental Fig. 1).

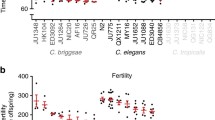
The effect of adult exposure to a 500 μg/mL green tea extract, b 100 μM NDGA, c 50 μM rapamycin, d 30 μM 17α-estradiol, and e 500 μM acarbose on the median lifespan of nine strains across three Caenorhabditis species. Three strains were tested from each species: C. elegans N2, MY16, and JU775 (black text), C. briggsae AF16, ED3092, and HK104 (dark gray text), C. tropicalis JU1373, JU1630, and QG834 (light gray text). Each point represents the change in median lifespan from an individual trial plate (compound treated) relative to the specific control (vehicle only) conducted. The bars represent the mean + / − the standard error of the mean. Replicates were completed at the three CITP testing sites (blue square–Buck Institute, green circle–Oregon, and salmon diamond–Rutgers). Asterisks represent p-values from the CPH model such that ****p < 0.0001, ***p < 0.001, **p < 0.01, and *p < 0.05

The mean adjusted swimming score across Caenorhabditis species after treatment with a GTE, or b NDGA is shown for young and old ages (adult days 6 and 12 for C. elegans and C. tropicalis, adult days 8 and 16 for C. briggsae). Strains tested include the following: three C. elegans strains (N2, JU775, MY16), three C. briggsae strains (AF16, ED3092, HK104), and three C. tropicalis strains (JU1373, JU1630, QG834). Each data point represents the mean from one trial, the bars represent the mean + / − the standard error of the mean, and the colors correspond to the treatment conditions starting at day 1 of adulthood (white–vehicle control, green–500 ug/mL GTE, mauve–100 uM NDGA). Replicates were completed at the three CITP testing sites (square- Buck Institute, diamond- Rutgers, circle- Oregon). Asterisks inside the plots represent p-values from the linear mixed model, and asterisks outside the plot represent p-values from the type III ANOVA indicating a significant compound by age interaction, such that ****p < 0.0001, ***p < 0.001, **p < 0.01, and *p < 0.05

Survival of nine Caenorhabditis strains on 40 mM paraquat after adult exposure to green tea extract (green symbols), or vehicle control (open symbols). Oxidative stress resistance was measured at day 6 and 12 of adulthood (C. elegans and C. tropicalis), or day 8 and 16 of adulthood (C. briggsae). Three strains were tested from each species: C. elegans N2, JU775 and MY16, C. briggsae AF16, ED3092 and HK104, and C. tropicalis JU1373, JU1630, and QG834. Dots represent the median survival of one plate replicate (gray–vehicle only control, green–500 μg/mL GTE). Replicates were completed at three CITP testing sites (circle–Oregon, square–Buck, diamond–Rutgers). Asterisks inside the plots represent p-values from the CPH model, and asterisks outside the plot represent p-values from the type III ANOVA indicating a significant compound by age interaction, such that ****p < 0.0001, ***p < 0.001, **p < 0.01, and *p < 0.05
The antioxidants green tea extract and NDGA extend Caenorhabditis lifespan in a species-specific manner
Using the streamlined pipeline we describe above, we evaluated a test set of compounds for lifespan effects. Among this test set, we found that rapamycin, acarbose, and 17α-estradiol were broadly ineffectual. Rapamycin only altered lifespan for two (C. briggsae HK104 and C. tropicalis JU1373) of the nine tested strains, while 17α-estradiol only had an effect on JU1630 (Fig. 1). We also found that acarbose did not statistically change the lifespan of any of the tested strains.
The two antioxidants we tested, GTE and NDGA, exerted the largest effect on lifespan in manual survival assays (Fig. 1). GTE extended lifespan in two species, C. elegans (two of three strains) and C. tropicalis (all three strains) (Fig. 1a). The effect on lifespan was more pronounced in C. elegans, with mean lifespan extended > 15% in both MY16 and JU775. In the laboratory C. elegans strain N2, we observed lifespan extension via GTE in only one lab, and the pooled results across labs was insignificant. We also observed lifespan extension with GTE in C. tropicalis, with all three strains tested showing small but significant increases in mean lifespan; the largest effect was in JU1630 (~ 11% increase in mean lifespan). In contrast, we found that all C. briggsae strains exhibited a significant decrease in survival when exposed to GTE, with mean lifespan decreasing by 10–19%. Overall, we observe a strong species-specific effect on longevity with GTE exposure.
We found that NDGA also impacted Caenorhabditis longevity in a species-specific manner (Fig. 1b). In C. elegans, much like with GTE, we measured an 8–16% increase in the mean lifespan of strains MY16 and JU775, while N2 had a similar, albeit insignificant, lifespan increase. The effect of NDGA on lifespan was more variable than we recorded for GTE as we observed a significant decrease in survival in five of six C. briggsae and C. tropicalis strains.
Antioxidants that decrease survival in certain genetic backgrounds extend lifespan in automated C. elegans Lifespan Machine assays
To increase throughput, we have implemented ALMs [34] (Supplemental Fig. 1, Phase Two) at all three CITP sites. The ALM is a lifespan analysis platform built on flatbed scanner technology that enables life-long imaging of animals at 1-h intervals, increasing both throughput and temporal resolution for data sampling. Our previous testing of compounds NP1, resveratrol, propyl gallate, thioflavin t, and α-ketoglutarate revealed that most ALM trials recapitulated outcomes from manual plate-based assays, although we did identify light sensitivity of some compounds as a factor that could change outcome [6]. During preliminary experiments on ALMs, we found that both GTE and NDGA increased lifespan in strains that also reported decreased lifespans in manual assays. More specifically, for GTE assays on the ALM, all C. briggsae strains exhibited increased survival compared to the control. For NDGA treatment, four of the six C. briggsae and C. tropicalis strains exhibited positive lifespan results on the ALM. Difference in outcome might be the consequence of increased light exposure on the ALMs [32], as light is known to induce photooxidative stress [39]. ALM protocols also involve fewer potentially damaging animal transfers, and feature no exposure to freshly treated compound plates in mid- to late-life, in contrast to manual assays [33, 40]. Our highly replicated data, however, emphasize that the methodological approach to lifespan determination is a factor in experimental outcome.
GTE and NDGA impact on swimming ability is strain-specific
With the goal of maintaining health over the lifetime, compounds that slow the age-related decline in swimming ability are of particular interest to the CITP. We previously showed that swimming ability as a measure of locomotion with compound treatment in Caenorhabditis does not always correlate with lifespan, and thus swim locomotion is an assay with potential to identify compounds that may improve healthspan independent of lifespan [35]. Because both GTE and NDGA induced strong species-specific effects on lifespan, we addressed whether these compounds could improve locomotion.
We found that with GTE treatment, all C. elegans strains showed an improvement in swimming ability (Fig. 2a). The improved locomotion effect was age-dependent in MY16 and JU775 (8–9% increase in mean swim score at old age as compared to the control, respectively), while N2 showed an overall robust improvement in locomotion (25% increase at young age, 146% increase at old age). The C. tropicalis strains (in which GTE improved lifespan), exhibited strain-specific responses in swimming ability: strain JU1630 showed no difference in locomotion with GTE treatment, while strain QG834 exhibited a small but significant decrease in swimming ability (2–4% decrease with age). JU1373 exhibited a strong age-related response in which GTE robustly decreased swimming ability at young age (14% decrease) but improved swimming ability during old age (16% increase). The effect of GTE on swimming ability in C. briggsae was minimal but strain-specific as we recorded decreases in swimming ability at young age in strains AF16 (23% decrease) and ED3092 (13% decrease), but a small increase at old age in HK104 (6% increase).
We also tested the effect of NDGA on locomotion in our panel of Caenorhabditis strains. With NDGA treatment, swimming ability improved in five of six C. elegans and C. briggsae strains, but none of the C. tropicalis strains (Fig. 2b). The effect in C. elegans was observed in two strains, with N2 gaining a general increase (11–13% increase in mean swimming with age) and with JU775 exhibiting a reduction in the age-related decline of swimming ability (6% increase at old age). Interestingly, all three C. briggsae strains showed an age-dependent improvement of locomotion at either young age (ED3092, 6% increase), or old age (HK104 and ED3092, 19–21% increase, respectively), irrespective of their reduced lifespan with NDGA treatment. C. tropicalis, which showed either no effect or a negative lifespan effect in response to NDGA, had no difference in swimming ability with NDGA treatment.
Overall, our tests underscore the following: (1) longevity and locomotory health are not well correlated; (2) interventions can elicit marked differences in locomotory healthspan, even in the absence of longevity changes.
GTE reduces oxidative stress resistance in some Caenorhabditis strains
We next assayed oxidative stress resistance with GTE treatment across our panel of strains because GTE exerted the most widespread effect on longevity and locomotion of the two antioxidants tested. In seven of the nine strains, GTE treatment reduced the animal’s ability to resist oxidative stress for at least one age tested (Fig. 3). This effect was most prominent in the C. briggsae strains, which exhibited significant decreases in survival at all ages tested (ranging from a 17% decrease in mean survival in old AF16 to a 65% decrease in young ED3092). Previous exposure to a mild stressor can result in a more robust response to future stressors, a process known as hormesis [41]. Continuous exposure to an antioxidant may subsequently reduce reactive oxygen species that promote an organism’s ability to mount an enhanced oxidative stress response when removed from the compound. While GTE does have antioxidant properties [42], the inability to resist oxidative stress may reflect an anti-hormetic effect. The only exceptions to the failure of GTE to increase oxidative stress resistance were in C. tropicalis JU1373, which exhibited an age-dependent increase in oxidative stress survival (22% increase at old age), and JU1630, with no effect at either age. C. elegans JU775 also had an increase in oxidative stress survival as compared to the control during old age (35% increase), though this increase was paired with a decrease in oxidative stress survival at young age (31% decrease).
We have found ALM-based thermotolerance assays to be challenging to reproduce across labs [35]. Still, as we continue to assess how broadly across compounds health measures are impacted, we tested thermotolerance consequent GTE treatment. As we previously observed, there was a high amount of variability within our dataset that made it difficult to identify significant effects on thermotolerance, but our results suggest that GTE may protect against thermosensitivity as most of the strains exhibited lifespan extension with GTE treatment (Supplemental Fig. 4).
Discussion
Examples of interventions that exert varied effects across species
As a group dedicated to identifying highly reproducible pharmacological interventions that extend lifespan, promote fundamental functionality such as locomotory ability, or both, with efficacy that applies over a genetically diverse Caenorhabditis test set, the CITP has first focused on testing compounds published to be effective in extending lifespan in nematodes or other model systems [5,6,7,8]. The five compounds we studied here, 17α-estradiol, acarbose, green tea extract, nordihydroguaiaretic acid, and rapamycin, are representative of that interest class of interventions. We evaluated the five compounds for longevity modulation across a genetically diverse test set and pursued two of the more potent promoters of longevity, GTE and NDGA, for impact on locomotory health and stress resistance. We observed that the antioxidants GTE and NDGA modestly extend Caenorhabditis lifespan in a species-specific manner (Fig. 1). Additionally, we found that two different survival assay protocols we employed (manual and using Automated Lifespan Machines) could report different lifespan outcomes of these antioxidants, with observed decreased survival for certain genetic backgrounds in manual survival assays contrasting with extended lifespan as determined on the ALMs (Fig. 1 and Supplemental Figs. 2, Fig. 3). GTE and NDGA confer strain-specific impact on swimming ability (Fig. 2), although GTE reduces oxidative stress resistance in some Caenorhabditis strains (Fig. 3), suggesting that GTE targets and the underlying cause(s) of diminished health for these assessments may reflect different processes. While lifespan and health can certainly be uncoupled, and both are plausible targets for intervention, this study, combined with previous observations, underscores the complex challenge to finding universal lifespan and healthspan extending interventions.
Implications for identification of conserved pharmacological interventions
Aging is a complex trait with a broad range of associated measurable outcomes. The inter-individual variability in outcome necessitates studies of significant size and replicate number to reliably identify anti-aging interventions. Because of the necessary study sizes, and the need to follow individuals over the lifetime, the initial characterization of compound effects on aging is frequently performed in short-lived animal models. The oft-unspoken assumption is that the near universality of phenotypic aging in multi-cellular animals reflects shared underlying causes of aging, and insights gained in model systems will be generalizable across taxa. At the same time, genetically identical individuals in the same environment can exhibit strikingly different aging trajectories [5, 43, 44]. Whether that variability in outcome reflects differences in aging symptoms due to unidentified environmental differences or stochastic biological events or, importantly, if those differences reflect different causes of aging experienced by individuals is not known.
The CITP attempts to account for issues of inter-individual differences and stochastic events by testing compounds at a large experimental scale using standardized protocols [33, 40], replicating experiments at three independent sites, and statistically partitioning the variance to determine the sources of variability (Supplemental Table 1 for this study supports that lab specific differences account for very little of experimental variation for GTE lifespan). The CITP tests compound interventions across a genetically diverse panel of Caenorhabditis strains and species to address the potential to identify interventions that are effective independent of genetic background. Presumably such broadly acting compounds would engage widely conserved pro-longevity mechanism. The large number of animals studied, and the numbers of independent replicates in CITP assays, increase confidence in reported outcomes.
In reporting outcomes, CITP always emphasizes that our conclusions, although rigorously reproduced, apply under the specific experimental conditions used for our investigation. For any reported CITP outcome, it remains possible that assay under alternative conditions will lead to distinct conclusions. Factors that lead to differences among studies conducted in the field include compound concentration and source, precise media composition, use or not of FUdR to induce sterility, and period of exposure to the intervention. As noted above, in this study, we also document an example in which method of analysis—manual vs. automated longevity assays can produce markedly different outcomes. Furthermore, when testing against a genetically diverse population as is modeled by CITP, the challenge of finding the optimal intervention introduction conditions is amplified by a need for chemical uptake by all strains and metabolism that will engage in similar ways. These challenges, and the expectation that many false negative outcomes will result, are hypothesized to be overcome by the identification of a rare broadly efficacious compound. We did not identify such a compound in the small set of 5 compounds tested in this study.
Regarding the challenge of translation and implications for identifying compounds that slow aging using C. elegans
Considerable interest in the application of TOR inhibitor Rapamycin for pro-longevity outcomes, anchored in both invertebrate and mammalian models, exists in the aging field [45,46,47] and strong genetic data support the importance of mTOR in longevity pathways under stress. That rapamycin was largely ineffective under CITP assay conditions may reflect one of the many conceptual impediments to direct translation success noted above, including intervention bioavailability and uptake, metabolism, and particular endpoints selected for assessment. Because of these considerations, we are careful to point out that absence of effect does not rule out that a compound might work in mammals.
Rather, it is the rare compound that passes out of the rigorous CITP testing that we seek to highlight for translational consideration. Compounds that are effective in promoting longevity in C. elegans and mammals have been reported. Examples of pharmacological interventions identified by C. elegans screening for longevity include anti-hypertension drug Captropil [48] which extended longevity in female mice [49]. Anti-diabetic drug Metformin can promote longevity in C. elegans and locomotory health in both C. elegans and C. briggsae [10] and promotes health and longevity in some mammalian studies [50, 51]. In this study, NDGA, which can extend longevity in male mice [28], extends lifespan across elegans variants.
As CITP generates additional data, cataloging successes in parallel mammalian outcomes will be important for honing screens to that best prioritize compounds for likelihood of benefit in mammals. Possibly, however, a strict adherence to a longevity endpoint is too narrow a net to cast. Along those lines, we note that collective data are converging to suggest the underappreciated value of the extent to which identified longevity compounds for C. elegans promote mammalian health and healthy aging. For example, CITP identified Thioflavin T as highly potent in C. elegans for longevity and mobility [5]. Testing of a more mouse-friendly variant of Thioflavin T, 2-hydroxyphenyl-benzoxazole, revealed a remarkable improvement in old age bone maintenance in mice [52] and protection against ALS pathology [53]. The geroscience concept (since aging predisposes to disease, interventions that target aging will also be effective against disease [54]) predicts such an outcome—interventions that robustly extend lifespan in C. elegans (genetic diversity similar to that among humans) or in Caenorhabditis species (genetic diversity similar to the mouse to human difference) may often confer the capacity to protect against disease and specific conditions of aging in all animals.
Health measures and longevity
The important question of how longevity and health intersect and are controlled by “aging pathways”, and the connection between them, is a matter of controversy [44]. The question of pharmacological impact on health in the context of genetic diversity has not been well investigated in models, despite the importance for considerations of translation in human populations.
Here we monitored Caenorhabditis locomotory features at multiple adult ages to conclude that there is not a strong correlation between longevity and locomotion for the five test compounds of focus. Moreover, assays of oxidative stress resistance and heat resistance did not establish a relationship between longevity and these health indicators across genetic backgrounds. Perhaps an important take home message for health metrics is that often-used metrics of stress resistance are anchored in acute, and likely non-physiological, challenges (i.e., 40 mM paraquat) that might be replaced by assays that more closely approximate native challenges of aging. For example, use of stress reporter gene expression patterns might prove to correlate better with animal health and longevity, providing a more precise assessment tool for intervention potential.
Against a small but expanding test set, we observe that compound genetic background and assay type can give rise to differences in lifespan evaluation and health assessment [5, 6]. The disconnect between lifespan and health measurements, and the genetic background dependency of those effects suggests additional complications that need to be addressed in screening experimental design. Stepping back to offer some perspective, it is likely unrealistic to expect many blockbuster aging interventions using broad based outcome evaluation. Nonetheless, the pursuit of the particular compounds that can positively move outcomes across genetic backgrounds and via multiple measures of heath remains a plausible strategy that spotlights priority interventions for mammalian testing.
Conclusion
Our study of five interventions (NDGA, GTE, 17α-estradiol, acarbose, and rapamycin) underscore the complexities of assessing longevity and health outcomes of candidate aging interventions. Using our standardized protocols, we find that the antioxidants GTE and NDGA extend Caenorhabditis lifespan in a species-specific manner. GTE and NDGA tests also revealed some survival assay-specific outcomes—in certain genetic backgrounds we found decreased survival in manual longevity assays, whereas we measured extended lifespan when we determined outcomes using the automated C. elegans Lifespan Machines. GTE and NDGA affected swimming ability in a strain-specific manner, and GTE lowered oxidative stress resistance in some Caenorhabditis strains. Lifespan and healthspan generally appear uncoupled in our dataset. Overall, our findings on this test set of interventions underscore how impactful genetic background, selected health assay, and protocol details are in the assessment of intervention effects. Interventions that meet the high bar of efficacy across a broad range of genetic backgrounds and across multiple experimental approaches may prove the exception, but such capacity would establish definitive priority for testing in mammalian models.
Data availability
All data, analysis scripts, and detailed SOPs are available as an online supplement [55].
References
Zhang S, Li F, Zhou T, Wang G, Li Z. Caenorhabditis elegans as a useful model for studying aging mutations. Front Endocrinol (Lausanne). 2020;11:1–9.
Arya U, Das CK, Subramaniam JR. Caenorhabditis elegans for preclinical drug discovery. Curr Sci. 2010;99:1669–80.
Lithgow GJ, Driscoll M, Phillips P. A long journey to reproducible results. Nature. 2017;548:387–8.
Teterina AA, et al. Genetic diversity estimates for the Caenorhabditis Intervention testing program screening panel. microPublication Biol. 2022. https://doi.org/10.17912/micropub.biology.000518.
Lucanic M, et al. Impact of genetic background and experimental reproducibility on identifying chemical compounds with robust longevity effects. Nat Commun. 2017;8:14256.
Banse SA, et al. Automated lifespan determination across Caenorhabditis strains and species reveals assay-specific effects of chemical interventions. GeroScience. 2019;41:945–60.
Coleman-Hulbert AL, et al. Caenorhabditis Intervention Testing Program: the tyrosine kinase inhibitor imatinib mesylate does not extend lifespan in nematodes. microPublication Biol. 2019. https://doi.org/10.17912/micropub.biology.000131.
Coleman-Hulbert AL, et al. Caenorhabditis Intervention Testing Program: the creatine analog β-guanidinopropionic acid does not extend lifespan in nematodes. microPublication Biol. 2020. https://doi.org/10.17912/micropub.biology.000207.
Morshead ML, et al. Caenorhabditis Intervention Testing Program: the farnesoid X receptor agonist obeticholic acid does not robustly extend lifespan in nematodes. microPublication Biol. 2020. https://doi.org/10.17912/micropub.biology.000257.
Onken B, et al. Metformin treatment of diverse Caenorhabditis species reveals the importance of genetic background in longevity and healthspan extension outcomes. Aging Cell. 2022;21:e13488.
Stout MB, et al. 17α-Estradiol alleviates age-related metabolic and inflammatory dysfunction in male mice without inducing feminization. J Gerontol A Biol Sci Med Sci. 2017;72:3–15.
Dykens JA, Moos WH, Howell N. Development of 17alpha-estradiol as a neuroprotective therapeutic agent: rationale and results from a phase I clinical study. Ann N Y Acad Sci. 2005;1052:116–35.
Harrison DE, et al. Acarbose, 17-α-estradiol, and nordihydroguaiaretic acid extend mouse lifespan preferentially in males. Aging Cell. 2014;13:273–82.
Strong R, et al. Longer lifespan in male mice treated with a weakly estrogenic agonist, an antioxidant, an α-glucosidase inhibitor or a Nrf2-inducer. Aging Cell. 2016;15:872–84.
Yamamoto M, Otsuki M. Effect of inhibition of α-glucosidase on age-related glucose intolerance and pancreatic atrophy in rats. Metabolism. 2006;55:533–40.
Miyamura M, et al. Effects of acarbose on the acceleration of postprandial hyperglycemia-induced pathological changes induced by intermittent hypoxia in lean mice. J Pharmacol Sci. 2010;114:32–40.
Kim JH, et al. Quercetin attenuates fasting and postprandial hyperglycemia in animal models of diabetes mellitus. Nutr Res Pract. 2011;5:107–11.
Cabrera C, Artacho R, Giménez R. Beneficial effects of green tea–a review. J Am Coll Nutr. 2006;25:79–99.
Weinreb O, Mandel S, Amit T, Youdim MBH. Neurological mechanisms of green tea polyphenols in Alzheimer’s and Parkinson’s diseases. J Nutr Biochem. 2004;15:506–16.
Lopez T, et al. Green tea polyphenols extend the lifespan of male Drosophila melanogaster while impairing reproductive fitness. J Med Food. 2014;17:1314–21.
Li YM, Chan HYE, Huang Y, Chen ZY. Green tea catechins upregulate superoxide dismutase and catalase in fruit flies. Mol Nutr Food Res. 2007;51:546–54.
Kitani K, Osawa T, Yokozawa T. The effects of tetrahydrocurcumin and green tea polyphenol on the survival of male C57BL/6 mice. Biogerontology. 2007;8:567–73.
Abbas S, Wink M. Epigallocatechin gallate from green tea (Camellia sinensis) increases lifespan and stress resistance in Caenorhabditis elegans. Planta Med. 2009;75:216–21.
Zhang L, Jie G, Zhang J, Zhao B. Significant longevity-extending effects of EGCG on Caenorhabditis elegans under stress. Free Radic Biol Med. 2009;46:414–21.
Xiong L-G, et al. Epigallocatechin-3-gallate promotes healthy lifespan through mitohormesis during early-to-mid adulthood in Caenorhabditis elegans. Redox Biol. 2018;14:305–15.
Zhang H, Shen W-J, Cortez Y, Kraemer FB, Azhar S. Nordihydroguaiaretic acid improves metabolic dysregulation and aberrant hepatic lipid metabolism in mice by both PPARα-dependent and -independent pathways. Am J Physiol Gastrointest Liver Physiol. 2013;304:G72-86.
Pardini RS, Heidker JC, Fletcher DC. Inhibition of mitochondrial electron transport by nor-dihydroguaiaretic acid (NDGA). Biochem Pharmacol. 1970;19:2695–9.
Strong R, et al. Nordihydroguaiaretic acid and aspirin increase lifespan of genetically heterogeneous male mice. Aging Cell. 2008;7:641–50.
Harrison DE, et al. Rapamycin fed late in life extends lifespan in genetically heterogeneous mice. Nature. 2009;460:392–5.
Robida-Stubbs S, et al. TOR signaling and rapamycin influence longevity by regulating SKN-1/Nrf and DAF-16/FoxO. Cell Metab. 2012;15:713–24.
Miller RA, et al. Rapamycin-mediated lifespan increase in mice is dose and sex dependent and metabolically distinct from dietary restriction. Aging Cell. 2014;13:468–77.
Abbott M, et al. A simplified design for the C. elegans lifespan machine. J Biol Methods. 2020;7:e137.
Caenorhabditis Intervention Testing Program. CITP Automated Lifespan Machine (ALM) SOPs. figshare. 2019. https://doi.org/10.6084/m9.figshare.c.4580546.v1. https://doi.org/10.6084/m9.figshare.c.4580546.
Stroustrup N, et al. The Caenorhabditis elegans Lifespan Machine. Nat Methods. 2013;10:665–70.
Banse SA, et al. The coupling between healthspan and lifespan in Caenorhabditis depends on complex interactions between compound intervention and genetic background. bioRxiv. 2022. https://doi.org/10.1101/2022.01.15.476462.
Caenorhabditis Intervention Testing Program. Caenorhabditis Intervention Testing Program: CeleST Data and Composite Score. figshare. 2021. https://doi.org/10.6084/m9.figshare.c.5126579.
Ibáñez-Ventoso C, Herrera C, Chen E, Motto D, Driscoll M. Automated analysis of C. elegans swim behavior using CeleST software. J Vis Exp. 2016;2016:1–9.
Restif C, et al. CeleST: computer vision software for quantitative analysis of C. elegans swim behavior reveals novel features of locomotion. PLoS Comput Biol. 2014;10:e1003702.
De Magalhaes Filho CD, et al. Visible light reduces C. elegans longevity. Nat Commun. 2018;9(1):927.
Plummer WT, et al. Standardized protocols from the Caenorhabditis Intervention Testing Program 2013–2016: conditions and assays used for quantifying the development, fertility and lifespan of hermaphroditic Caenorhabditis strains. Protoc Exch. 2017. https://doi.org/10.1038/protex.2016.086.
Hood WR, Zhang Y, Mowry AV, Hyatt HW, Kavazis AN. Life history trade-offs within the context of mitochondrial hormesis. Integr Comp Biol. 2018;58:567–77.
Forester SC, Lambert JD. Antioxidant effects of green tea. Mol Nutr Food Res. 2011;55:844–54.
Herndon LA, Schmeissner PJ, Dudaronek JM, Brown PA, Listner KM, Sakano Y, Paupard MC, Hall DH, Driscoll M. Stochastic and genetic factors influence tissue-specific decline in ageing C. elegans. Nature. 2002;419:808–14.
Bansal A, Zhu LJ, Yen K, Tissenbaum HA. Uncoupling lifespan and healthspan in Caenorhabditis elegans longevity mutants. Proc Natl Acad Sci USA. 2015;112:E277-286.
Selvarani R, Mohammed S, Richardson A. Effect of rapamycin on aging and age-related diseases - past and future. Geroscience. 2021;43:1135–58.
Hansen M, Kennedy BK. Does longer lifespan mean longer healthspan? Trends Cell Biol. 2016;26:565–8.
Robida-Stubbs S, Glover-Cutter K, Lamming DW, Mizunuma M, Narasimhan SD, Neumann-Haefelin E, Sabatini DM, Blackwell TK. TOR Signaling and rapamycin influence longevity by regulating SKN-1/Nrf and DAF-16/FoxO. Cell Metab. 2012;15:713–24.
Kumar S, Dietrich N, Kornfeld K. Angiotensin converting enzyme (ACE) inhibitor extends Caenorhabditis elegans life span. PLoS Genet. 2016;12: e1005866.
Strong R, et al. Lifespan benefits for the combination of rapamycin plus acarbose and for captopril in genetically heterogeneous mice. Aging Cell. 2022;21: e13724.
Triggle CR, et al. Metformin: is it a drug for all reasons and diseases? Metabolism. 2022;133: 155223.
Kulkarni SA, Gubbi S, Barzilai N. Benefits of metformin in attenuating the hallmarks of aging. Cell Metab. 2020;32:15–30.
Evans DS, et al. Longitudinal functional study of murine aging: a resource for future study designs. JBMR Plus. 2021;5: e10466.
Evans TM, et al. Moderate modulation of disease in the G93A model of ALS by the compound 2-(2-hydroxyphenyl)-benzoxazole (HBX). Neurosci Lett. 2016;624:1–7.
Sierra F, Kohanski RA. The role of the National Institute on Aging in the development of the field of geroscience. Cold Spring Harb Perspect Med. 2023;13(10):a041211.
CITP. Caenorhabditis Intervention Testing Program: data and SOPs for the green tea extract and nordihydroguaiaretic acid manuscript. figshare. 2022. https://doi.org/10.6084/m9.figshare.c.5850182.
Acknowledgements
We acknowledge the members of the Lithgow, Driscoll, and Phillips labs for helpful discussions. We thank the CITP Advisory Committee and Ronald Kohanski (National Institute on Aging) for extensive discussion. We thank Asher Cutter, Marie-Anne Félix, and Christian Braendle for providing strains that they had directly collected. Additional strains were provided by the CGC, which is funded by NIH Office of Research Infrastructure Programs (P40 OD010440).
Funding
This work was supported by funding from National Institutes of Health grants (U01 AG045844, U01 AG045864, U01 AG045829, U24 AG056052) and the Glenn Foundation for Medical Research and the Larry L. Hillblom Foundation.
Author information
Authors and Affiliations
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
This article is published under an open access license. Please check the 'Copyright Information' section either on this page or in the PDF for details of this license and what re-use is permitted. If your intended use exceeds what is permitted by the license or if you are unable to locate the licence and re-use information, please contact the Rights and Permissions team.
About this article

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Cite this article
Banse, S.A., Sedore, C.A., Johnson, E. et al. Antioxidants green tea extract and nordihydroguaiaretic acid confer species and strain-specific lifespan and health effects in Caenorhabditis nematodes. GeroScience 46, 2239–2251 (2024). https://doi.org/10.1007/s11357-023-00978-0
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11357-023-00978-0