
Abstract
The reverse seasonal flooding that resulted from the construction of the Three Gorges Project has induced a drastic degradation of draw-down zone vegetation and the seed bank in the reservoir. The composition and distribution of seed banks and their similarities to standing vegetation were determined through field investigations and germination experiments on the vegetation as well as soil seed bank data collected from three restoration sites in Zigui county, Wanzhou city, and Zhongxian county to uncover the effects of vegetation restoration on seed bank development in the Three Gorges Reservoir region. The results showed the soil seed banks were composed of 44, 43, and 50 herbaceous species, and an average seed density of 9418.79 ± 804.92, 4283.44 ± 427.89, 6539.28 ± 432.22 seeds/m2, respectively. The soil seed bank was mainly composed of annual and perennial herbaceous plants. Most of the dominant species of soil seed banks came from introduced species. Reverse seasonal flooding induced the spatial variation of species diversity and seed density of the soil seed bank. The species diversity in the upper part of the draw-down zone was higher than that in the middle and lower parts. The highest seed density occurred in the middle upper parts of the draw-down zone, followed by the lower part. The similarity coefficient between the soil seed bank and the extant vegetation community was 0.495, 0.526, and 0.550 at Zigui, Wanzhou, and Zhongxian restoration sites, respectively.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
The soil seed bank and its relationship with above-ground vegetation plays a decisive role in the succession of a plant community (Liu et al. 2009a; Kettenring and Galatowitsch 2011). On the one hand, the composition and structure of a soil seed bank determines the composition, structure, and distribution of the extant plant community to a certain extent (Liu et al. 2014; O’Donnell et al. 2016). The soil seed bank significantly affects the regeneration of the plant community, especially in areas that are frequently disturbed (Liu et al. 2005; Landman et al. 2007). On the other hand, species composition and spatial structure as well as plant growth and development of the above-ground vegetation influence the composition and spatial variation of the soil seed bank (Wang et al. 2011a). Replanting can effectively promote the restoration process of riparian vegetation; however, the stability and sustainability of the restored primary ecosystem is relatively low (Hong et al. 2012; Zhang et al. 2017). The composition of a soil seed bank and its relationship with the extant vegetation is considered crucial evidence for vegetation development (Wang et al. 2010). Research conducted on the composition and spatial variation of soil seed banks and their relationship with extant vegetation provides useful information for the prediction of the succession direction of restored vegetation and the improvement of vegetation management (Nishihiro et al. 2006; Liu et al. 2009b).
The construction of the Three Gorges Dam, China’s largest hydro-power and water conservation project on the Yangtze River, significantly raises the water level in the reservoir area. Many terrestrial ecosystems became riparian ecosystems. As a result, a draw-down zone with a total area of 450 km2 with a 30 m flooding amplitude appeared. The construction of the Three Gorges Dam also changed the flooding regime of the riverbank from summer flooding–winter drought to summer drought–winter flooding, the absolute opposite of the natural flood rhythms of the Yangtze River, for electricity production and downstream flood control (New and Xie 2008; Chen et al. 2016). The flooding duration varies along altitude gradient: the lower part (150–155 m), middle part (160–165 m) and upper part (170–175 m) of the draw-down zone are subject to approximately 240, 180, and 90 days of flooding, respectively (Lu et al. 2010a). Hence, many plants disappear from the riverbank because they cannot adapt to the drastic change in flooding (Chen and Xie 2009; Chen et al. 2013). Consequently, the composition and structure of riparian communities have degraded dramatically and change along the altitude gradient (Lu et al. 2010b; Wang et al. 2011b). The degradation and spatial change of riparian vegetation caused by the reverse seasonal flooding further induced a degradation and special spatial variation of the riparian soil seed bank during the completion of the Three Gorges Dam (Wang et al. 2010; Lu et al. 2010a). The last field investigation showed that the degradation and spatial heterogeneity of the soil seed bank became more prevalent (Zhang et al. 2016).
Riparian vegetation plays a critical role in biodiversity conservation, pollutant decomposition, soil and water conservation, and riverbank protection (Hong et al. 2012; Zhang et al. 2017). A series of experimental and demonstrational projects on riparian vegetation restoration were applied after the completion of the Three Gorges Dam to improve the ecological environment and maintain the ecological safety of the Three Gorges Reservoir area. Investigation of the extent community indicated that these projects promoted the reestablishment of the riparian plant community (Tang et al. 2012; Chen et al. 2016). Some studies have reported the characteristics of the soil seed bank composition and spatial variation of the natural draw-down zone (Lu et al. 2010a; Wang et al. 2010; Zhang et al. 2016). However, the characteristics of the soil seed bank of the rehabilitated draw-down zone and the effects of vegetation plantation on the soil seed bank are not detailed enough. We chose three typical restoration sites for the case study and conducted field investigations and germination experiments of the soil seed banks. The objectives of our research were (1) test the species composition of the soil seed banks and analyze the characteristic composition of seed banks across sites restored with different species, (2) quantify the variation of soil seed banks in species biodiversity and seed density along altitude gradients, (3) examine the similarity between soil seed banks and standing vegetation, (4) evaluate the change of vegetation on the restored draw-down zone in the Three Gorges Reservoir area.
Methods
Study zone
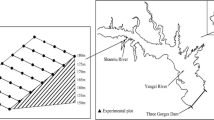
Three restoration sites were chosen as field experimental plots in Zigui county in Hubei province, and Wanzhou city and Zhongxian county in Chongqing municipality which are located at the head, girdle, and tail sections of the Three Gorges Reservoir area, respectively (Fig. 1). The geographical environment and vegetation restoration measures are shown in Table 1.

Location of three vegetation rehabilitation sites in the Three Gorges Reservoir area
Seed bank sampling
Three sample belts, including the lower part (150–155 m), middle part (160–165 m), and upper part (170–175 m) of the draw-down zone, were designed along an altitude gradient on each plot. Ten quadrats (1 × 1 m2) were randomly distributed in each sample belt. Five sub-samples of soil seed banks were collected in each quadrat and pooled as a sample. The surface vegetation was removed, and the sub-samples of the soil seed banks were obtained using a soil collector which collected samples sizes 8 cm in diameter with a depth of 15 cm. Thirty soil seed bank samples were collected from each experimental field plot during riverbank exposure, which occurred from March to May of 2014.
Vegetation sampling
Field vegetation investigations were conducted on the plots and sample belts in September of 2014. Ten woody quadrats (5 × 5 m2) were randomly distributed in each sample belt. Five herbaceous quadrats (1 × 1 m2) were placed at the center and four corners of the woody quadrats. Plant number, coverage, and average height of each species were recorded within each woody and herbaceous quadrat.
Examination of soil seed banks
The seedling-emergence method (ter Heerdt et al. 1996; Lu et al. 2010a) was used to quantify seed bank species composition and density. Soil samples were dried naturally in a laboratory. Debris was removed from the samples, then the samples were crushed, and stones and impurities were removed with a 4 mm sieve. A 2-cm layer of each sieved soil sample was plated on a drought sand substrate (thickness 3 cm) in a germination tray (20 cm × 15 cm × 5 cm). The sand was used as a substrate and had been dried for 12 h at 120 °C in an oven before being used to ensure that any seeds present in the sand matrix were inactivated. The germination trays were randomly placed in artificial climate chambers. Soil seed bank samples were germinated at a temperature of 25 °C, a light measurement of 24,000 lx, and at a humidity level of 60%. Samples were watered every day to keep the soil moist while germinated seedlings were identified and removed. The species and number of seedlings were recorded at the same time. The unidentified seedlings were transplanted into pots to grow and develop until they were identified. Species and number of germinated seeds in each soil sample were recorded for further analysis. The germination experiment lasted 5 months.
Data analysis
Each soil sample was used as a replicate for statistical analysis. The species and species life-form of the soil samples were analyzed. The importance value of each species in soil seed bank was calculated with relative density and relative frequency within each sample belt and restoration site respectively, and was further used to calculate the Simpson dominance index, Shannon–Wiener diversity index, and Pielou evenness index (Wang et al. 2010). Characteristics of species diversity and species spatial variation along the altitude gradient in the restoration sites were analyzed. Furthermore, one-way ANOVA analysis was applied using the altitude gradient as the independent variable and the seed density as the dependent variable to investigate the effects of elevation on seed density of soil banks (Lu et al. 2010a). Tukey’s tests were further employed to conduct multiple comparisons if a significant difference was detected (P < 0.05).
The relationship between the soil seed bank and plant community were analyzed according to the similarity coefficient and importance value percentage of common species. The Sørensen–Dice index was calculated as follows: SC = 2w/(a + b) where, “w” is the number of species occurring both in the soil seed bank and plant community, “a” is the number of species occurring only in the soil seed bank, and “b” is the number of species occurring only in vegetation (Cui et al. 2016). The importance value of each species in the plant community was calculated with relative density, relative coverage, and relative frequency according to the field investigation data of the extant plant community (Enright et al. 2007). The percentage of importance value of each species was further calculated both in the soil seed bank and plant community (the percentage of importance value of a species refers to the percentage of the species’ importance value accounting for the total importance value of the soil seed bank or plant community) (Cui et al. 2016). The common species occurring both in the soil seed bank and plant community were classified as four types using the Whipple (1978) method: a, species accounting for a large proportion both in the soil seed bank and plant community; b, species accounting for a large proportion in the soil seed bank but only a small proportion in the plant community; c, species accounting a large proportion in the plant community but only a smaller proportion in the soil seed bank; d, species accounting for a smaller proportion both in the soil seed bank and plant community.
Results
Species composition of the soil seed bank
The species composition of soil seed bank changed across the three restoration sites (Table 2). At the Zigui restoration site, 44 species from 22 families and 38 genera germinating from soil seed bank samples were identified, many of which came from the Compositae family. There were 17 species whose seed number accounted for over 1% of the total number of seeds in the soil seed bank. Nine of the 20 introduced species successfully established and produced seeds. Paspalum distichum, an introduced species, appeared as the dominant species in the community with a seed density of 1113.32 ± 209.30/m2. At the Wanzhou restoration site, 43 species from 21 families and 37 genera germinated from soil seed bank samples, many of which were from the Gramineae family. There were 19 species whose seed numbers accounted for over 1% of the total number of seeds from the soil seed bank. Twelve species of the 26 introduced species successfully established and produced seeds. Setaria viridis occurred as the dominant species in the community, with a seed density of 1333.60 ± 223.71/m2. At the Zhongxian restoration site, 50 species from 29 families and 45 genera germinated from soil seed bank samples, many of which belonged to the Gramineae family. There were 19 species whose seed numbers accounted for over 1% of the total number of seeds from soil seed banks. Eight species of the 16 introduced species successfully established and produced seeds. Hemarthria altissima, an introduced species, became the dominant species in the community, with a seed density of 1131.90 ± 201.26/m2.
The life-form composition of the soil seed bank was mainly composed of annuals and perennial herbs in the three restoration sites. Annual herbs accounted for 68.20, 67.40, and 48.00% of the total soil seed banks in the Zigui, Wanzhou, and Zhongxian restoration sites respectively, whereas perennial herbs were 18.20, 30.20, and 46.00% of the total soil seed banks. There were 13.63, 2.33, and 10.00% shrubs and trees species occurring at the three restoration sites, respectively (Table 3).
Species diversity and spatial variation of soil seed banks
The species diversity varied among the three restoration sites. The Zhongxian restoration site had the highest species number while the Wanzhou restoration site had the highest species diversity index, dominance index, and evenness index (Table 4). The distribution of species diversity changed along the altitude gradient among the three restoration sites. The species number, species diversity index, and dominance index of the soil seed bank within the upper and middle riparian zones were all higher than those within the lower riparian zones in Zigui and Wanzhou, and those indicators that were within the upper and lower riparian zones were higher than within the middle riparian zone in Zhongxian. The evenness index changed irregularly.
Seed density and spatial variation of the soil seed bank
There was a significant difference in the seed density of soil seed banks among the three restoration sites (F = 8.107, n = 89, P < 0.01). The average seed density was 9418.79 ± 804.92, 6539.28 ± 432.22, and 4283.44 ± 427.89/m2, ranging within 4,164.0–14,609.0, 4,743.0–8,297.0, and 2,140.0–6,218.0/m2 in the Wangzhou, Zhongxian, and Zigui restoration sites, respectively. The quantitative distribution of soil seed banks changed significantly along the altitude gradient at all three restoration sites (P < 0.01) (Fig. 2). Seed density first increased and then decreased with increase in altitude for the Wanzhou and Zhongxian restoration sites. The highest seed density in the soil seed bank occurred in the middle riparian zone, where the seed number accounted for 47.8 and 38.0% of the total number of soil seed banks. Meanwhile, seed density increased continually with the increase in altitude for the Zigui restoration site, and the highest seed density of soil seed banks occurred at the upper riparian parts, accounting for 42.40% of the total number of soil seed banks.

Soil seed bank density (mean ± SE) along altitude gradient. The different letters above the color column indicate the significance level
The relationship between the soil seed bank and the extant plant community
The species composition of the extant plant community at the three restoration sites differed from one another. However, the life-form composition was mainly composed of annuals and perennials. The species number changed along the altitude gradient, and the highest biodiversity appeared in the upper part of the draw-down zone in the three restoration sites (Table 5). At the Zigui restoration site, there were 61 species from 25 families and 54 genera. Species from the Gramineae and Asteraceae families accounted for 24.59 and 18.03% of the total species number in the extant plant community, respectively. Cynodon dactylon became the dominant species, with 45.7% of the total importance value of the community. At the Wanzhou restoration site, 52 species from 20 families and 46 genera occurred. Species from the Gramineae family accounted for 25.42% of the total species number of community. Cynodon dactylon also ranked as the dominant species with 23.30% of the total importance value of the community. At the Zhongxian restoration site, 59 species from 28 families and 55 genera were established. Species from the Compositae family accounted for 18.64% of the total species number of the community. Hemarthria altissima dominated the community with 32.00% of the total importance value of the community.
There were 26 common species that were present in the soil seed bank and the extant vegetation, with a similarity coefficient of 0.495 at the Zigui restoration site; hence, there were 25 and 30 common species with a similarity coefficient of 0.526 and 0.550 at the Wanzhou and Zhongxian restoration sites, respectively (Table 6).
Common species were classified a, b, c, and d types according to their abundance in the seed bank and the extant vegetation (Whipple 1978). The composition of common species in soil seed banks and in extant plant communities differed among the three restoration sites (Fig. 3). At the Zigui restoration site, there were six a type species, six b type species, seven c type species, and seven d type species. The most abundant species of each type was Cynodon dactylon, Bidens pilosa, Phytolacca acinosa, and Eclipta prostrate, respectively. At the Wanzhou restoration site, there were seven a type species, eight b type species, six c type species, and five d type species. The most abundant species of each type was Cynodon dactylon, Xanthium sibiricum, Setaria viridis, and Humulus japonicus, respectively. There were nine a type species, six b type species, eight c type species, and seven d type species at the Zhongxian restoration site. Hemarthria altissima, Alternanthera Philoxeroides, Setaria viridis, and Artemisia carvifolia were the most abundant species of each type. Some introduced species, including Cynodon dactylon and Hemarthria altissima, became the dominant species, with relatively high percentage of importance values in the three restored sites.

Importance value percentage accounted for by the species present both in soil seed bank and plant community [a Zigui, b Wanzhou, c Zhongxian]. 1. Cynodon dactylon 2. Setaria viridis 3. Paspalum distichum 4. Abutilon theophrasti 5. Acalypha australis 6. Erigeron annuus 7. Oxalis corniculata 8. Bidens pilosa 9. Polygonum hydropiper 10. Xanthium sibiricum 11. Rumex dentatus 12. Bidens frondosa 13. Solanum nigrum 14. Phyllanthus urinaria 15. Setaria glauca 16. Alternanthera Philoxeroides 17. Digitaria cruciata 18. Digitaria sanguinalis 19. Mazus japonicus 20. Humulus japonicus 21. Eclipta prostrata 22. Artemisia carvifolia 23. Hemarthria altissima 24. Trigonotis peduncularis 25. Gnaphalium affine
Discussion
Effects of vegetation restoration on the development of soil seed banks
The effects of vegetation restoration on soil seed banks differ remarkably from the effects of natural vegetation restoration. It is generally believed that natural vegetation restoration slowly enhances the species diversity of soil seed banks, while vegetation plantation quickly increases species diversity and the seed density of soil seed banks (Baldwin and Derico 1999; Landman et al. 2007). The dramatic change of environments has caused a serious degradation of vegetation in the Three Gorges Reservoir area. Vegetation plantation could rapidly promote vegetation restoration and reconstruction (Tang et al. 2012; Chen et al. 2016). Our previous research on the planted and naturally restored draw-down zone indicated that replanting effectively promoted the development of soil seed banks (Zhang et al. 2017). Seed production of introduced species and other species that were facilitated by the introduced species to establish enriched the biodiversity and increase the seed density of the soil seed bank (Leck 2003; Hong et al. 2012).
Our research showed that soil seed banks were composed of 44, 43, and 50 plant species, including 20, 26, and 13 introduced species, and 35, 31, and 42 native species, at the Zigui, Wanzhou, and Zhongxian restoration sites, respectively. The composition of the soil seed banks was in accordance with the composition of the extant vegetation and was mainly composed of annual and perennial species from the Asteraceae and Gramineae families. In the seed banks and vegetation, it was found that most of the dominant species came from introduced species. These species, which included Cynodon dactylon, Setaria viridis, Hemarthria altissima, and Polygonum hydropiper, adapted to the changed flooding pattern through their life cycle (annual herbs) or had good tolerance to flooding (perennial herbs). All of them had high seed production (Shen et al. 2011; Qin et al. 2014). The natural restored draw-down zone was rehabilitated slowly due to lack of propagule (Zhang et al. 2016, 2017). The introduced species provide additional propagules and enriched the species biodiversity to some extent. However, not all introduced species appeared in the seed banks. Many mature woody species, for example Morus alba, Salix variegata, Distylium chinense, and Leucaena leucocephala, did not form seed banks under the stress of the prolonged reverse seasonal flooding. Seed dormancy and specific germination requirements may also contribute to their failure to germinate (Lucas et al. 2012; Kettenring 2016). As a result, the species biodiversity of soil seed banks was limited at the vegetation restoration sites.
Spatial variation of the soil seed banks at vegetation restoration sites
Flooding significantly influences the species composition and density of soil seed banks (Greet et al. 2012; Yuan et al. 2014). The study by Nicol et al. (2003) indicated that the composition of soil seed banks had a close correlation with flooding. The amplitude and pattern of flooding had a crucial effect on the spatial variation of soil seed banks (Greet et al. 2012). Flooding affects the composition and distribution of soil seed banks through three pathways (Peterson and Baldwin 2004; James et al. 2007; Nielsen et al. 2012). The first is that flooding differentiates the exposed season and duration of a riverbank, which further differentiates the riverbank in the rainy season and the timeline along the altitude gradient (Weiterova 2008; Wang et al. 2013). The second is that flooding causes seeds to disperse, and some seeds that land on the soil surface move with the water flow (Thompson et al. 2014). The third is that flooding induces the heterogeneity of soil environments along the altitude gradient, which further affects seed vigor in soil seed banks. Seeds that suffer long periods of drought and submersion lose their ability to successfully establish (James et al. 2007; Vidal et al. 2014). The habitat heterogeneity of the riverbanks increased the heterogeneity of the soil seed banks in spatial variation (Lu et al. 2010a; Zhang et al. 2016).
A spatial variation with increasing altitude gradient was observed for the diversity and average density of the soil seed bank at the natural restoration sites in the Three Gorges Reservoir area. Species diversity and seed density were highest in the middle part of the draw-down zone, followed by the top part, and were lowest for the lower part (Zhang et al. 2016). This spatial variation was related to the heterogeneity that resulted from the reverse seasonal flooding which induced a variation of flooding duration along the altitude gradient. The exposed season and soil environment were favorable for seed deposition and survival in the middle riparian zone (Zhang et al. 2016, 2017). At the vegetation restoration sites, the species diversity and seed density of soil seed banks also changed with the increase in the altitude gradient. However, the spatial difference along the riverbank decreased significantly. This occurred because the vegetation restoration introduced some species in the lower and upper riparian zones to promote vegetation development, which increased the species diversity and seed density in those places and reduced the spatial heterogeneity of the soil seed bank distribution.
The similarity between soil seed banks and extant vegetation
Flooding increased the similarity coefficient of soil seed banks and plant communities, as it caused the species diversity in both to simplify (Li et al. 2008, Lu et al. 2010b). Many original riparian plants could not establish themselves and survive on the riverbank because their life cycle did not coincide with the regime of the reverse seasonal flooding in the Three Gorges Reservoir area (Yuan et al. 2014). The riparian plant community has degraded to grass and is mainly composed of annual plants and perennial herbs (Lu et al. 2010b; Su et al. 2012), and a few trees and shrubs survived in the upper riparian zone (Chen et al. 2016). Soil seed banks and standing vegetation were quite similar to one another when soil seed banks were mainly composed of annual and perennial herbaceous plants (Touzard et al. 2002; James et al. 2007). The degradation of the plant community further under the influence of the reverse seasonal fluctuation induced the simplification of the soil seed bank, which further changed the relationship between soil seed banks and extant vegetation in the Three Gorges Reservoir area (Wang et al. 2010; Lu et al. 2010a).
Hopfensperger (2007) found that standing vegetation and its associated seed bank was the least similar in forest ecosystems, most similar in grasslands, and of intermediate similarity in wetlands. The similarity coefficient was 0.13, 0.37, 0.07, and 0.24 at Cahngshou, Zhongxian, Wushan, and Zigui natural restoration sites during the primary impounding period in the Three Gorges Reservoir area, respectively (Lu et al. 2010a, 2010b). The similarity coefficient was relatively low as most of the surviving wood species in vegetation stopped producing seeds under the reverse seasonal flooding. The similarity coefficient at natural restoration sites was increased after 6 years of restoration, ranging from 0.48 to 0.53, when most woody species disappeared from vegetation, and the composition of seed bank and vegetation became simple (Zhang et al. 2016). In this study, the similarity coefficient at the three restoration sites ranged from 0.49 to 0.55. Those values suggest that vegetation restoration increased the similarity coefficient to some extent, which is in accordance with the results of our previous research on soil seed banks of the natural and planted draw-down zones (Zhang et al. 2017). This occurred because several introduced herbaceous plants with good adaptation became extreme dominant species in both the community and soil seed banks (class a in Fig. 2), increasing species similarities. By comparison, dominance of the dominant species in the natural draw-down zone is lower than that in the planted draw-down zone (Zhang et al. 2016, 2017). The increased similarity and dominant species’ dominance, and the species composition characteristics of the soil seed bank and vegetation limits the vegetation of the draw-down zone to grassland dominated by several introduced herbaceous plants including mostly Cynodon dactylon, Setaria viridis, Hemarthria altissima, and Polygonum hydropiper.
Conclusions
The introduced species in vegetation restoration enhanced species composition of the draw-down zone in the Three Gorges reservoir area. However, the effects of rehabilitation on species diversity of soil seed banks were limited under the influence of reverse seasonal flooding. The introduced annual and perennial herbs survived and produced seeds. Some species from the Asteraceae and Gramineae families adapted to the changed flooding pattern and became dominant species. The compositions of soil seed banks and extant vegetation were mainly composed of annual and perennial herbs. The reverse seasonal flooding also induced the species diversity and seed density of soil seed banks changing with the increase of the altitude gradient. The highest species diversity and seed density occurred at the middle and the upper draw-down zone, respectively. The similarity between soil seed banks extant vegetation ranged from 0.49 to 0.55 at the vegetation restoration sites. The characteristics of species composition of the soil seed bank and extant vegetation, and the reverse seasonal flooding determined whether a plant community remains at the grassland stage with annual and perennial herbs for a long term.
References
Baldwin AH, Derico EF (1999) The seed bank of a restored tidal freshwater marsh in Washington, DC. Urban Ecosyst 3:5–20
Chen FQ, Xie ZQ (2009) Survival and growth responses of Myricaria laxiflora seedlings to summer flooding. Aquat Bot 90:333–338
Chen FQ, Wang CH, Jia GM (2013) Ecology of Salix variegate seed germination: implications for species distribution and conservation in the Three Gorges region. S Afr J Bot 88:243–246
Chen SH, Chen FQ, Zhang M, Wang YJ (2016) Characteristics of artificial and natural restored plant communities within the water-level fluctuation zone along Xiangxi river. J Hydroecol 37:17–23 (in Chinese)
Cui L, Li W, Zhao X, Zhang M, Lei Y (2016) The relationship between standing vegetation and the soil seed bank along the shores of lake Taihu, China. Ecol Eng 96:45–54
Enright NJ, Mosner E, Miller BP, Johnson N, Lamont BB (2007) Soil vs canopy seed storage and plant coexistence in species-rich Australian shrublands. Ecology 88:2292–2304
Greet J, Cousens RD, Webb JA (2012) Flow regulation affects temporal patterns of riverine plant seed dispersal: potential implications for plant recruitment. Freshw Biol 57:2568–2579
Hong JM, Liu S, Shi GP, Zhang YQ (2012) Soil seed bank techniques for restoring wetland vegetation diversity in Yeyahu Wetland, Beijing. Ecol Eng 42:192–202
Hopfensperger KN (2007) A review of similarity between seed bank and standing vegetation across ecosystems. Oikos 116:1438–1448
James CS, Capon SJ, White MG, Rayburg SC, Thoms MC (2007) Spatial variability of the soil seed bank in a heterogeneous ephemeral wetland system in semi-arid Australia. Plant Ecol 190:205–217
Kettenring KM (2016) Viability, dormancy, germination, and intraspecific variation of Bolboschoenus maritimus (alkali bulrush) seeds. Aquat Bot 134:26–30
Kettenring KM, Galatowitsch SM (2011) Seed rain of restored and natural prairie wetlands. Wetlands 31:283–294
Landman GB, Kolka RK, Sharitz RR (2007) Soil seed bank analysis of planted and naturally revegetating thermally-disturbed riparian wetland forests. Wetlands 27:211–223
Leck MA (2003) Seed-bank and vegetation development in a created tidal freshwater wetland on the Delaware River, Trenton, New Jersey, USA. Wetlands 23:310–343
Li JM, Xu HL, Zhang ZJ, Ye M, Wang ZR, Li Y (2008) The characteristics of soil seed bank and standing vegetation in differently degraded areas in the lower reaches of Tarim river. Acta Ecol Sin 28:3826–3836 (in Chinese)
Liu GH, Zhou J, Li W, Cheng Y (2005) The seed bank in a subtropical freshwater marsh: implications for wetland restoration. Aquat Bot 81:1–11
Liu M, Jiang G, Yu S, Li Y, Li G (2009a) The role of soil seed banks in natural restoration of the degraded hunshandak sandlands, northern china. Restor Ecol 17:127–136
Liu WZ, Zhang QF, Liu GH (2009b) Seed banks of a river-reservoir wetland system and their implications for vegetation development. Aquat Bot 90:7–12
Liu QY, Jung M, Lü XG (2014) A review of similarity between soil seed bank and above ground vegetation in wetlands. Acta Ecol Sin 34:7465–7474 (in Chinese)
Lu ZJ, Li LF, Jiang MX, Huang DY, Bao DC (2010a) Can the soil seed bank contribute to revegetation of the draw down zone in the Three Gorges Reservoir Region. Plant Ecol 209:153–165
Lu ZJ, Li LF, Huang HD, Tao M, Zhang QF (2010b) Preliminary effects of impounding on vegetation in drawdown zone of the Three Gorges Reservoir Region. J Wuhan Bot Res 28:303–314 (in Chinese)
Lucas CM, Mekdeçe F, Nascimento CMN, Holanda ASS, Braga J, Dias S, Sousa S, Rosa PS, Suemitsu C (2012) Effects of short-term and prolonged saturation on seed germination of Amazonian floodplain forest species. Aquat Bot 99:49–55
New T, Xie ZQ (2008) Impacts of large dams on riparian vegetation: applying global experience to the case of China’s Three Gorges Dam. Biodivers Conserv 17:3149–3163
Nicol JM, Ganf GG, Pelton GA (2003) Seed banks of a southern Australian wetland: the influence of water regime on the final floristic composition. Plant Ecol 168:191–205
Nielsen DL, Podnar K, Watts RJ, Wilson AL (2012) Empirical evidence linking increased hydrologic stability with decreased biotic diversity within wetlands. Hydrobiologia 708:81–96
Nishihiro J, Nishihiro MA, Washitani I (2006) Assessing the potential for recovery of lakeshore vegetation: species richness of sediment propagule banks. Ecol Res 21:436–445
O’Donnell J, Fryirs KA, Leishman MR (2016) Seed banks as a source of vegetation regeneration to support the recovery of degraded rivers: a comparison of river reaches of varying condition. Sci Total Environ 542:591–602
Peterson JE, Baldwin AH (2004) Seedling emergence from seed banks of tidal freshwater wetlands: response to inundation and sedimentation. Aquat Bot 78:243–254
Qin HW, Liu ZX, Zhong Y, Zehng LD, Liu R, Yu J (2014) The physiological and germination responses of Cynodon dactylon seeds to simultaneous submergence. Chin J Grassl 36:76–82 (in Chinese)
Shen JH, Zeng B, Lei ST, Su XL, Huang WJ (2011) Seed submergence tolerance of four annual species growing in the water-level-fluctuation zone of Three Gorges Reservoir, China, and effects of long-term submergence on their seed germination. J Plant Ecol 35:237–246 (in Chinese)
Su XL, Zeng B, Huang WJ, Xu S, Lei S (2012) Effects of the Three Gorges Dam on preupland and preriparian drawdown zones vegetation in the upper watershed of the Yangtze River, P. R. China. Ecol Eng 44:123–127
Tang XQ, Wu M, Jin F (2012) Vegetation restoration and reconstruction in the water-fluctuation zone of Three Gorges reservoir area. J Yangtze River Sci Res Inst 29:13–17 (in Chinese)
ter Heerdt GNJ, Verweij GL, Bekker RM, Bakker JP (1996) An improved method for seed-bank analysis: seedling emergence after removing the soil by sieving. Funct Ecol 10:144–151
Thompson SE, Assouline S, Chen L, Trahktenbrot A, Svoray T, Katul GG (2014) Secondary dispersal driven by overland flow in drylands: review and mechanistic model development. Mov Ecol 2:1–13
Touzard B, Amiaud B, Langlois E, Lemauviel S, Clément B (2002) The relationships between soil seed bank, above ground vegetation and disturbances in an eutrophic alluvial wetland of Western France. Flora 197:175–185
Vidal DB, Andrade IL, Andrade ELP, Mielke MS (2014) Effects of submergence in water on seed germination and vigor of the Copaifera lucens (Fabaceae) seedlings. J Forest Res 25:903–908
Wang XR, Cheng RM, Xiao WF, Guo Q, Feng X (2010) Relationship between standing vegetation and soil seed bank in water-level-fluctuating Zone of Three Gorges Reservoir at the beginning after charging water. Acta Ecol Sin 30:5821–5831 (in Chinese)
Wang JC, Zhu B, Wang T (2011a) Characteristics of restoration of natural herbaceous vegetation of typical water-level fluctuation zone after flooding in the Three Gorges reservoir area. Resour Environ Yangtze Basin 20:603–610 (in Chinese)
Wang Q, Yuan XZ, Liu H, Zhang YW, Chen ZL (2011b) Effect of initial impoundment on the vegetation and species diversity in water-level fluctuation zone of the Three Gorges reservoir. J Nat Resour 10:1680–1693 (in Chinese)
Wang YC, Jiang DM, Toshio O, Zhou Q (2013) Recent advances in soil seed bank research. Contemp Probl Ecol 6:520–524
Weiterova I (2008) Seasonal and spatial variance of seed bank species composition in an oligotrophic wet meadow. Flora 203:204–214
Whipple SA (1978) The relationship of buried germinating seeds to vegetation in an old-growth Colorado subalpine forest. Can J Bot 56:1505–1509
Yuan SH, Zeng B, Su XL, Xu JP (2014) Effect of water-level fluctuation discrepancy on the composition of different annuals in Three Gorges Reservoir drawdown zone. Acta Ecol Sin 22:6481–6488 (in Chinese)
Zhang M, Chen FQ, Chen J, Wang YJ, Wang J (2016) Effects of the reverse seasonal flooding on riparian soil seed bank in the Three Gorges Reservoir Region: a case study in Shanmu River. SpringerPlus 5:1–11
Zhang M, Chen FQ, Wu Y, Ma YR, Guan SP, Huang YW (2017) Characteristics of the soil seed bank of planted and natural restored drawdown zones in the Three Gorges Reservoir Region. Ecol Eng 103:127–133
Acknowledgements
We thank Shoupeng Guan and Tianyi Wu for their help with the experiment. We would like to thank Binhe Gu for his patience in refining the manuscript. This work is supported by the National Natural Science Foundation of China (51379105). We thank LetPub (http://www.letpub.com) for its linguistic assistance during the preparation of this manuscript.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
This article is published under an open access license. Please check the 'Copyright Information' section either on this page or in the PDF for details of this license and what re-use is permitted. If your intended use exceeds what is permitted by the license or if you are unable to locate the licence and re-use information, please contact the Rights and Permissions team.
About this article

Cite this article
Zhang, M., Chen, F., Chen, S. et al. The soil seed bank of a rehabilitated draw-down zone and its similarity to standing vegetation in the Three Gorges Reservoir Area. Ecol Res 32, 1011–1021 (2017). https://doi.org/10.1007/s11284-017-1518-4
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11284-017-1518-4