
Abstract
The industrial footprint of oil and gas extraction in Alberta, Canada dissects the landscape in a network of linear and polygonal disturbances such as roads and well pads. The changes to biotic and abiotic factors beyond the boundaries of these disturbances into the adjacent ecosystem are called edge effects. A common assumption is that edge effects are proportional to the width of the disturbance, with wider disturbances leading to deeper effects into the forest. The objective of this study is to compare edge effects on plant composition, seed dispersal distance, seed weight, and microclimate across three disturbance types: narrow (< 20 m) and wide (> 20 m) roads, and well pads (> 100 m; 1.9 ha) in upland boreal forests in the oil sands region of north-eastern Alberta. We sampled 1 m2 plots at six distances (0 m, 10 m, 15 m, 25 m, 50 m, and 75 m) along transects running perpendicular from the disturbance edge into the forest. Edge effects for plant communities were contained at the disturbance edge for all disturbance types considered. Understory species richness was lower at the disturbance edge and was dominated by introduced species and species with lighter seeds and with medium-distance dispersal mechanisms. Disturbance edges were characterized as having higher soil moisture content, higher soil temperature, and a thinner organic matter layer compared to the forest interior (> 10 m). With no effect of disturbance width on width of edge effects, our study does not support the assumption that narrower disturbances will necessarily result in narrower edge effects for vegetation and abiotic metrics.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
In Canada, forested landscapes are increasingly experiencing disturbances associated with industrial exploration and extraction of non-renewable resources including oil, gas, and minerals (Venier et al. 2014). These disturbances occur in various shapes and sizes but are generally categorized into linear and polygonal disturbances. Examples of linear disturbances include access roads, seismic lines for oil and gas exploration, and pipelines for oil and gas transportation. Polygonal disturbances include mines for extraction of minerals and oil, and well pads for installation of infrastructure for oil and gas in situ extraction. Linear and polygonal disturbances include some combination of removal of trees and other vegetation, removal of the organic layer, soil compaction, and addition of rock and gravel to form a stable operating surface. The direct effects of these mechanical disturbances in forests have been the focus of many research projects (Davidson et al. 2020; Finnegan et al. 2018; Godefroid and Koedam 2004; Lovitt et al. 2018). However, these same disturbances also have indirect effects on the adjacent forest that are not immediately evident or well understood.
These indirect effects, also called edge effects, are defined as changes in abiotic and biotic factors, such as environmental conditions and species composition, at the interface of disturbed areas and the adjacent forest (Gehlhusen et al. 2000; Murcia 1995). For example, in forests cleared for seismic lines in linear corridors, the altered environmental conditions on the seismic line can extend into the adjacent uncut stands, resulting in increased exposure of the stand edges to wind and light, which can affect soil moisture and temperature, and tree blowdown (Dabros et al. 2017; Harper et al. 2005; Stern et al. 2018). Changes in environmental conditions can impact populations of individual species and community structure near disturbance edges (Dabros et al. 2017). Many native forest plant and animal species require undisturbed interior forest habitats to persist and do not fare as well near stand edges adjacent to disturbances (Bender et al. 1998; Harper and Macdonald 2002). Forest edges can also be prone to invasion by non-native and invasive species that spread from disturbed areas into the forest where they can compete with resident species, cause destruction of habitats, and alter ecological processes (Franklin et al. 2021; Gignac and Dale 2007; Harper et al. 2005; Neher et al. 2013). Therefore, the overall industrial footprint on forest landscapes is likely larger than that represented by the mechanically disturbed area because of edge effects.
In the Canadian province of Alberta, calculations of interior habitat area, or the amount of habitat not subject to human disturbances, considers that disturbances extend beyond the mechanically disturbed area into the forest (Alberta Environment and Parks 2022a). The amount of interior habitat adjacent to wide linear disturbances (> 20 m wide) and polygonal disturbances is reduced by a 200 m wide buffer zone, assumed to be the zone of edge effect (Alberta Environment and Parks 2022a). However, the 200 m buffer zone is discounted in forests adjacent to narrow linear disturbances (< 20 m wide) (Alberta Environment and Parks 2022a). The width of these adopted buffers includes a major inherent assumption that narrow linear features have narrower edge effects (Alberta Environment and Parks 2022a).
The assumption that the width of edge effects vary according to disturbance width is not consistently supported by scientific evidence from the boreal forest. Adjacent to wide linear disturbances, increases in tree height, species diversity, and light intensity extended farther into the forest compared to narrow linear disturbances (Abib et al. 2019; Echiverri et al. 2022; Franklin et al. 2021). However, other studies have found the opposite, with a greater magnitude of edge effects for tree basal area and density for roads (60 m wide) compared to well pads (80 m wide) (Jackson et al. 2023). Echiverri et al. (2022) found that there was an edge effect on plant community composition for narrow linear disturbances, but not for wide linear disturbances. The range of environmental and biotic responses to disturbance width across these studies underscores the complexity of edge effects and reveals potential knowledge gaps.
Accurately estimating the extent of edge effects is especially important in areas with persistence of ubiquitous industrial disturbances such as those found in the Cold Lake region of north-eastern Alberta. This region has a long history of oil sands development resulting in an extensive and dense network of linear and polygonal disturbances, most of which are still evident on the landscape today (Government of Alberta 2023a). The Cold Lake in situ oil sands development area covers approximately 68800 ha containing over 6000 km of linear disturbances (seismic lines, roads, pipelines, and transmission lines) and over 6500 ha of overall direct human footprint developed between 1946 and 2019 (ABMI 2019; Alberta Environment and Parks 2020). The interior habitat indicator values show that less than 30% of the Cold Lake area is currently classified as undisturbed (Alberta Environment and Parks 2022b). The long industrial history resulting in extensive disturbance and habitat fragmentation in the Cold Lake region has had adverse effects on biodiversity; however, the full extent of effects on the biota are only superficially understood (Ficken et al. 2019). More research is needed to measure and predict the persistence of viable populations of disturbance-sensitive species on this highly disturbed landscape and their capacity to disperse and colonize disturbed areas during the ecological recovery process.
This study aims to test the assumption that edge effects of wider disturbances on vegetation and environmental conditions extend further into the adjacent forest than narrower disturbances. We compare the edge effects of narrow linear disturbances (hereafter referred to as narrow roads) to wider linear (wide roads) and polygonal disturbances (well pads) within the upland boreal forest of the Cold Lake oil sands region. Ultimately, this study will deliver knowledge on the extent of edge effects on baseline biotic and abiotic components in boreal ecosystems, which is crucial for planning successful restoration of industrial footprints.
Methods
Site description
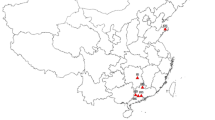
The study sites were located in the upland areas of the dry mixedwood subregion of the boreal forest, at the Imperial Oil lease, near Cold Lake in north-eastern Alberta (~ 54.7053° N, 110.5131° W; Fig. 1). The Cold Lake region has vast oil sands reserves with a long history of extraction beginning in the 1950s (Alberta Environment and Parks 2021; Government of Alberta 2023a). Sites are dominated by trembling aspen (Populus tremuloides), white spruce (Picea glauca), and balsam poplar (Populus balsamifera). The average temperature and precipitation over the growing season (May to September) in the Cold Lake area is 13.8 °C and 302 mm, respectively (Government of Canada 2017). Upland soils in the study area are classified as gray luvisols on medium textured till (Alberta Agriculture and Forestry 2019). All sites are located within the Cold Lake First Nations traditional territory (Government of Alberta 2012).

Study area in north-eastern Alberta, showing the location of sample sites adjacent to well pads (red), narrow roads (pink), and wide roads (yellow). The extent of the study area is displayed in the inset map in purple. Sample sites consist of transects perpendicular to disturbance edges
Experimental design
Three different disturbance types were chosen for this study: (i) narrow roads (NR; < 20 m wide), (ii) wide roads (WR; > 20 m wide), and (iii) well pads (WP; > 100 m wide and an average area of 1.9 ha; Fig. 2). ‘Narrow’ and ‘wide’ roads are relative terms to compare the two road types in this study, and not a description of a standard disturbance type. The width of the disturbance provided above includes the road or well pad disturbance and the ditch or vegetated edge to the forest edge (Fig. 2). The forest edge for each disturbance was delineated by the beginning of the stand of mature trees that were not disturbed or removed during road or well pad construction.

Typical layout of the three disturbance types: a narrow roads, b wide roads, and c well pads, denoting disturbance width. Data collection occurred from the forested edge into the adjacent forested area of each disturbance type
Narrow roads had one or two lanes and a gravel or dirt surface, while wide roads had two or more lanes, a gravel surface, and were maintained with vegetation mowing along the ditches and grading. Wide roads were sampled on the side of the road with a pipeline. Roads and well pads were constructed between 1973 and 2007 (Supplementary Table 1). The year of disturbance was based on the oldest disturbance of the natural landscape that was visible on aerial imagery (Government of Alberta 2023b).
All data collection was completed during the 2021 growing season, except canopy cover, which was measured in 2022. For each disturbance type, two 75 m-long transects spaced 45 m apart were run perpendicular to the edge of the disturbance into the adjacent forest (Fig. 3). There were eight replicate sites for each disturbance type, with four edges having mainly eastern exposure and four mainly western exposure from each disturbance type (3 disturbance types × 8 replicates = 24 sites).

Schematic of sampling transects and quadrats set up for a single site (not to scale). Plant and environmental variables were measured at every 1 m2 quadrat. The black rectangle illustrates a plot, which includes quadrats on both transects at a given distance
At each site, vegetation was sampled along the two transects (Fig. 3) in six 1 m2 quadrats. The quadrats were placed at the forest edge (0 m) and at distances of 10 m, 15 m, 25 m, 50 m, and 75 m towards the forest interior (Fig. 3), for a total of 288 quadrats (6 distances × 2 transects × 24 sites).
Within each quadrat, the percent cover of plants below 1.5 m was estimated visually to the nearest 0.5% and species were identified in the field to the lowest taxonomic level possible. Percent cover was not estimated for plants above 1.5 m due to difficulties in estimating cover within a 1 m2 quadrat from below the canopy. Nomenclature for vascular species follows the most recent records from the Database of Vascular Plants of Canada (VASCAN) (Brouillet et al. 2010 +) and the Integrated Taxonomic Information System (ITIS 2023) for non-vascular species. Plants were grouped into two strata according to their height, as overstory (> 1.5 m) and understory (< 1.5 m) species.
To explore the relationship between understory vascular plant traits and disturbance width, we evaluated seed weight, seed dispersal distance, and introduced status. Seed dispersal mechanisms were obtained from the Traits of Plants in Canada (TOPIC) database, which are derived from records in the literature (Aubin et al. 2012). Species were categorized as short-distance dispersed (gravity, ant, explosive, small animal ingestion, or carried externally), medium-distance dispersed (wind dispersed herbs), or long-distance dispersed (wind dispersed woody species, bird dispersed). Seed weight records were compiled from the Seed Information Database (SID) (SER et al. 2023) and the number of introduced species was based on the list of introduced species in Alberta from the VASCAN database (Brouillet et al. 2010 +).
Environmental variables and canopy cover were collected within 10 m of each 1 m2 quadrat (Fig. 3). Canopy cover was estimated using a spherical convex densiometer, with a reading taken facing each cardinal direction and later averaged. Soil moisture (% volumetric water content) was measured at a depth of 12 cm using a portable time-domain reflectometer (TDR; Field Scout TDR-300, Spectrum Technologies Inc., Plainfield Illinois, USA). Soil temperature was measured at a depth of 3 cm using a General Industries digital thermometer. Soil moisture and temperature measurements were repeated three times throughout the summer. Depth of the organic layer was measured from the surface to the mineral layer with a ruler.
Data analyses
For all univariate and multivariate analysis, variables were averaged between the two transects within each site at each distance. The average values at each site-distance combination are referred to subsequently as ‘plots’ (Fig. 3). As observations within sites are not independent, sites were treated as a random effect in the statistical models. Species richness (number of identifiable taxa) was calculated separately for the understory and overstory. The mean seed weight, number of short-, medium-, and long-distance dispersed species, and the number of introduced understory vascular species in each plot were calculated. For each trait, all species were weighted equally, regardless of cover. Prior to calculating the mean seed weight of species in each plot, seed weight was log-transformed.
Aligned-Rank Transform Analysis of Variance (ART ANOVA) was used to test (α = 0.05) for differences in understory and overstory species richness, canopy cover, organic matter depth, seed weight, and the number of short-, medium-, and long-distance dispersed species and introduced species as a function of disturbance type and distance from the disturbance edge (Wobbrock et al. 2011). Soil moisture and soil temperature were also analysed using ART ANOVA, but only as a function of distance from the disturbance edge. No analyses were carried out on disturbance types for these two variables because sites were sampled on different days under different meteorological conditions. ART ANOVA was chosen because the data did not meet the assumptions of parametric tests and could not be addressed with transformations (Harrar et al. 2019). An analysis of deviance based on type III Wald F tests with Kenward-Roger degrees of freedom was used to assess which effects and interactions were significant. Post hoc tests using the ART-C procedure with Holm's adjustment for multiple comparisons were used to identify differences for significant fixed factors (Elkin et al. 2021).
Constrained ordination using Redundancy Analysis (RDA) was performed to test whether understory species composition differed as a function of distance from the edge and disturbance type (Legendre and Legendre 2012; Rao 1964). The Hellinger transformation was applied to the cover data prior to RDA to improve the model (Legendre and Gallagher 2001). Species with less than three occurrences or less than 3% cover across all plots were removed from the analysis. Model significance was assessed after 999 permutations. Confidence interval ellipses (75% CI) were constructed around group centroids for distance and treatment classes separately and overlaid on the ordination plots to compare groups. Post hoc tests on distance used 75 m as a reference for interior forest conditions with the permutest function allowing observations to be permuted randomly within sites. The 75 m distance was selected as the reference for interior forest conditions based on other studies in the Canadian boreal forest, which have found that edge effects do not typically extend past 60 m (Dabros and Higgins 2023; Echiverri et al. 2022; Harper et al. 2015).
All analyses were performed using R software version 4.3.0 (R Core Team 2023). The vegan package was used for RDA (Oksanen et al. 2022), ARTool package for ART ANOVA (Kay et al. 2021), and emmeans package for univariate post hoc tests (Lenth et al. 2023).
Results
Species richness and canopy cover
Woody overstory species richness and canopy cover did not differ significantly among the disturbance types or distances from the disturbance edge (Table 1; Supplementary Fig. 1). Three woody overstory species were found on average per plot (Supplementary Fig. 1a), with a dominance of P. tremuloides, P. glauca, and P. balsamifera. Understory species richness differed by distance, but not across disturbance types (Table 1), with less species found at the edge (0 m) compared to all other distances further into the forest (10–75 m) (Fig. 4).

Understory species richness by disturbance type: narrow roads (NR), wide roads (WR), and well pads (WP); and by distance from the disturbance edge. Significant post hoc differences for distance are shown with different letters
Understory plant community
Redundancy analysis revealed a significant difference in understory species composition among distances (F[5,126] = 1.34; p = 0.001), but not disturbance type (F[2,126] = 2.03; p = 0.56) or their interaction (F[10, 126] = 0.84; p = 0.27). Overall, 2.0% of the variability was explained by the fixed factors (adjusted R2; Fig. 5). Understory species composition at the disturbance edge (0 m) was different from 75 m (p = 0.001), but there were no differences detected in the community composition between the other distances and 75 m (p > 0.3). The first axis explains 3.8% of variability and distinguished between edge and other distances. Chamaenerion angustifolium (Chaang), Equisetum pratense (Equpra), and Taraxacum officinale (Taroff), were most closely correlated with axis 1 and the 0 m centroid, indicating their dominance near the disturbance edge (Fig. 5a). Astragalus cicer, Cirsium arvense, and Melilotus spp. were introduced species only found at disturbance edges (0 m) (Supplementary Table 2). Common native forest species Cornus canadensis (Corcan), Mitella nuda (Mitnud), Petasites frigidus var. palmatus (Petpal), and Viburnum edule (Vibedu) were highly correlated with axis 1 and the 75 m distance centroid indicating their dominance in the forest interior (Fig. 5a). In Supplementary Table 2, we provide the average percent cover and frequency of occurrence for all identified understory species and their corresponding dispersal distance class, seed weight, and status (introduced vs. native). The distances from the disturbance edge at which each species is found are also listed in Supplementary Table 2.

RDA of understory species composition for a all distances from the disturbance edge and b all disturbance types: narrow roads (NR), wide roads (WR), and well pads (WP). The species that contribute to 50% of the variability are labelled in red. Species abbreviations are as follows: Aranud = Aralia nudicaulis, Corcan = Cornus canadensis, Chaang = Chamaenerion angustifolium, Equpra = Equisetum pratense, Fravir = Fragaria virginiana, Loninv = Lonicera involucrata, Mitnud = Mitella nuda, Petpal = Petasites frigidus var. palmatus, Rosaci = Rosa acicularis, Rubuda = Rubus idaeus, Symcio = Symphyotrichum ciliolatum, Taroff = Taraxacum officinale, and Vibedu = Viburnum edule
Plant traits
Seed weight, the number of short-distance dispersed species, and the number of introduced species differed significantly with distance, but the number of medium- and long-distance dispersed species did not (Table 2; Supplementary Fig. 2). Species at the edge (0 m) had approximately 50% lighter seeds (0.9 g/1000 seeds) than those in the forest (1.5 g/1000 seeds; Fig. 6a). On average, three species with short-distance dispersal mechanisms occurred at the edge (0 m) compared to about five species in the forest (10–75 m) (Fig. 6b). There were no differences in seed weight, or the number of short-, medium-, or long-distance dispersed species across disturbance types (Table 2).

Plant traits for understory vascular species including a median seed weight and b the number of short-distance dispersed species by disturbance type: narrow roads (NR), wide roads (WR), and well pads (WP); and by distance from the disturbance edge. Seed weight was weighted equally for each species found. The plot-level seed weights were back-transformed to conserve the observed values. Significant post hoc differences for distance are shown with different letters
Introduced species represented a small proportion of the species richness and percent cover in plots, with a maximum of four introduced species and 7% cover per plot. However, more introduced species were found at the edge plots than in the forest (Table 2; Fig. 7). Introduced species were found in 21 out of 24 plots at the edge compared to 23 out of 120 plots in the forest. There was no difference in the number of introduced species by disturbance type (Table 2). Introduced taxa found include: Achillea millefolium, A. cicer, C. arvense, Melilotus spp., Sonchus spp., T. officinale, and Trifolium hybridum (Supplementary Table 2).

Median number of introduced species by disturbance type: narrow roads (NR), wide roads (WR), and well pads (WP); and by distance from the disturbance edge. Significant post hoc differences for distance are shown with different letters
Environmental variables
Soil moisture content (Fig. 8a) and soil temperature (Fig. 8b) were higher at the edge compared to plots further into the forest (10–75 m), except for soil moisture at 50 m. The organic matter layer was thinner at the edge compared to the plots further into the forest (Fig. 8c), with no differences found across the disturbance types (Table 3).

Environmental variables by distance from the disturbance edge: a volumetric water content b soil temperature, and c organic matter depth. Significant post hoc differences for distance are shown with different letters
Discussion
Extent of edge effects across three disturbance types
Edge effects on forest plant communities and environmental variables were present at the immediate edge (0 m) but did not appear to extend further into the forest for any disturbance type, regardless of width, in this study. Thus, our results show that there was no relationship between disturbance width and width of edge effects. This improved understanding of the edge effects of narrow linear and polygonal disturbances could help to inform improvements to biodiversity management frameworks used by the Alberta government, sub-regional land-use management planning, and guidelines for restoration practices.
Wide roads and well pads had more traffic and maintenance, such as mowing and resurfacing, compared to the narrow roads. The increased presence of vehicles and regular road maintenance on these disturbance types seem to favour dispersal of seeds on tires and provide more suitable conditions for the establishment of disturbance tolerant introduced species (Neher et al. 2013; Šerá 2010). Roads with more traffic can also lead to higher heavy metal concentrations, higher salt concentrations, NO2 pollution, and dust in adjacent habitats, favouring nitrogen and salt tolerant plant species and reducing overall species richness compared to roads with lower traffic (Lee et al. 2012; Trombulak and Frissell 2000). In this study, changes to the species composition through invasion of introduced species at forest edges were evident in all disturbance types regardless of usage or maintenance, suggesting the importance of other factors.
The original disturbance severity might play a more crucial role than the frequency of usage or disturbance width in contributing to edge effects (Hansen and Clevenger 2005). Other studies have found there are many common factors in the construction of linear and polygonal disturbances that can impact the surrounding plant communities including vegetation clearing, topsoil removal, infilling with gravel, and topography modification (Godefroid and Koedam 2004; Neher et al. 2013; Willier et al. 2022). Regardless of disturbance width, these steps can result in soil compaction and have a major impact on local hydrology (Davidson et al. 2021; Lovitt et al. 2018). In Alberta, the interior habitat indictor assumes wide disturbances (> 20 m) are highly engineered, with more surface construction than narrow disturbances (< 20 m; Alberta Environment and Parks 2022a). This assumption about disturbance severity is used to justify discounting the width of edge effects adjacent to narrow linear disturbances when calculating the interior habitat indicator. However, in the current study all disturbance types were heavily engineered, and their construction required many of the same processes and materials, regardless of disturbance width. Therefore, the similarities in disturbance severity across all three disturbance types could help explain the similarities in their plant communities despite differences in disturbance width.
Characteristics of the assessed edges
The three disturbance types shared commonalities in edge characteristics. At all edge types there were more introduced species (Brouillet et al. 2010+), fewer short-distance dispersed species, and species with lighter seeds, lower understory species richness, higher soil temperature, higher soil moisture content, and a thinner organic matter layer. When vegetation is cleared during road and well pad construction, introduced species take advantage of the increase in light availability, soil temperature, and soil moisture (Lee et al. 2012; Ross et al. 2002). Species with lighter seeds or longer distance seed dispersal mechanisms, can also benefit from the increased wind speeds in the open canopy conditions as they aid in seed dispersal, regardless of disturbance width (Resasco et al. 2014; Roberts et al. 2018).
Changes in species composition at the edge caused by the increase in introduced and disturbance-adapted species may also impact litter inputs, possibly leading to changes in the microbial community and decomposition of organic matter (Allison and Treseder 2011; Scheffer et al. 2001). The combined increase in soil compaction from road and well pad construction, changes to litter inputs, and decomposition of organic matter could reduce the organic matter depth at disturbance edges (Davidson et al. 2020).
Despite the possible influx of introduced species on vehicle tires, overall understory species richness was still lower at the edge of all three disturbance types than into the forest. This could be due to competitive exclusion of some forest species by disturbance-adapted species (Lee et al. 2012). For example, C. angustifolium is typically a pioneer species that favours disturbed areas and outcompetes other species by taking advantage of the increased nitrogen availability and acting as a major nitrogen sink (Beese et al. 2022; Ficken et al. 2019; Mitchell et al. 2007). T. officinale, is also characteristic of disturbed conditions and was present at the edges in our study and others (Godefroid and Koedam 2004; Honnay et al. 2002). This non-native forb can take advantage of the higher light availability and temperatures on disturbance edges to outcompete other species (Honnay et al. 2002). Hylocomium splendens and Orthilia secunda are two shade-tolerant (Lieffers 1995) native species that were not found at any disturbance edge plots in this study and may have been adversely affected by disturbance-adapted species and open conditions at disturbance edges.
The altered environmental conditions that favour introduced and disturbance-adapted species did not seem to extend beyond the forest edge in our study. For example, wind dispersal of introduced species decreases past the forest edge as wind speeds decline into the forest interior (20–25 m from the edge) (Roberts et al. 2018; Stern et al. 2018). Although not significant, we observed a reduction in medium-distance dispersed species beyond the edge. One of these medium-distance dispersed species, C. angustifolium, also has few adaptations to survive in the shade of the interior forest (Lieffers 1995; Lieffers and Stadt 1994).
Shade-tolerant native species are better adapted to interior forest conditions compared to disturbance tolerant species (Honnay et al. 2002). V. edule and M. nuda, were the native species most correlated with the interior forest plots in this study and others (Ficken et al. 2019; Gignac and Dale 2007; Harper and Macdonald 2002). Thus, on landscapes highly fragmented by industrial development, it is important that forest fragments of adequate size remain on the landscape to maintain interior forest habitats that can sustain species absent from disturbed areas. Further, the presence of such habitats can serve as sources of propagules for recovery of nearby disturbed areas. This will reduce costs associated with active restoration after road and well pad decommissioning. This study suggests that due to limited edge effects in upland boreal aspen-dominated forests, the overall human footprint on vegetation for this forest type is not much larger than the area directly disturbed. Even small residual patches have value as sources of propagules for restoration of adjacent disturbed areas. However, the findings of this study should not be extrapolated to other forest types nor to other taxa.
Conclusion
Our study does not support the assumption that narrow linear disturbances result in narrower edge effects for vegetation and abiotic metrics, compared to wider disturbances. Changes to the plant community composition, soil moisture content, soil temperature, and organic matter depth did not extend past the immediate edge (0 m) for all disturbance types. For future evaluation of edge effects into the forest, the severity of the adjacent disturbance should be examined in addition to disturbance width. This study can contribute to restoration planning by providing evidence of native plant species propagules within 10 m of road and well pad edges. Also, this study provides evidence that the disturbance footprint for vegetation could be less than previously estimated in the Cold Lake area.
Data availability
The data presented in this study are available on request from the corresponding author.
References
Abib TH, Chasmer L, Hopkinson C, Mahoney C, Rodriguez LCE (2019) Seismic line impacts on proximal boreal forest and wetland environments in Alberta. Sci Total Environ 658:1601–1613. https://doi.org/10.1016/j.scitotenv.2018.12.244
Alberta Agriculture and Forestry (2019) Agricultural Regions of Alberta Soil Information Database. https://geodiscover.alberta.ca/geoportal/rest/metadata/item/db848869ef74446e93fd0d9622d5b259/html. Accessed May 2023
Alberta Biodiversity Monitoring Institute (ABMI) (2019) Human footprint inventory (HFI): wall-to-wall human footprint inventory Alberta. Alberta Environment. https://www.abmi.ca/home/data-analytics/da-top/da-product-overview/Human-Footprint-Products/HF-inventory.html. Accessed March 2023
Alberta Environment and Parks (2020) Shapefiles for land disturbance due to oil sands projects. http://osip.alberta.ca/library/Dataset/Details/742. Accessed June 2023
Alberta Environment and Parks (2021) The Alberta Human Footprint Monitoring Program (AHFMP) - A historical overview of geophysical exploration in Alberta. Government of Alberta. https://open.alberta.ca/publications/alberta-human-footprint-monitoring-program-historical-overview-geophysical-exploration Accessed June 2023
Alberta Environment and Parks (2022a) Interior habitat indicator for Alberta. Government of Alberta. https://open.alberta.ca/opendata/gda-84c7d1ae-68b1-4cd7-919c-76b57b4f20f8. Accessed June 2023
Alberta Environment and Parks (2022b) Interior habitat indicator for Alberta. https://extranet.gov.ab.ca/srd/geodiscover/srd_pub/environment/InteriorHabitatIndicator.zip. Accessed June 2023
Allison SD, Treseder KK (2011) Climate change feedbacks to microbial decomposition in boreal soils. Fungal Ecol 4:362–374. https://doi.org/10.1016/j.funeco.2011.01.003
Aubin I, Messier C, Gachet S et al (2012) TOPIC – Traits of Plants in Canada database. Natural Resources Canada, Canadian Forest Service. http://www.nrcan.gc.ca/forests/research-centres/glfc/20303. Accessed May 2023
Beese WJ, Sandford JS, Harrison ML, Filipescu CN (2022) Understory vegetation response to alternative silvicultural systems in coastal British Columbia montane forests. For Ecol Manag 504:119817. https://doi.org/10.1016/j.foreco.2021.119817
Bender DJ, Contreras TA, Fahrig L (1998) Habitat loss and population decline: a meta-analysis of the patch size effect. Ecology 79:517–533. https://doi.org/10.1890/0012-9658(1998)079[0517:HLAPDA]2.0.CO;2
Brouillet L, Coursol F, Meades SJ et al (2010+) Database of Vascular Plants of Canada (VASCAN). http://data.canadensys.net/vascan/search. Accessed May 2023
Dabros A, Higgins KL (2023) Vegetation recovery and edge effects of low-impact seismic lines over eight-year period in boreal uplands of northern Alberta. For Ecol Manag 532:120850. https://doi.org/10.1016/j.foreco.2023.120850
Dabros A, James Hammond HE, Pinzon J, Pinno B, Langor D (2017) Edge influence of low-impact seismic lines for oil exploration on upland forest vegetation in northern Alberta (Canada). For Ecol Manag 400:278–288. https://doi.org/10.1016/j.foreco.2017.06.030
Davidson SJ, Goud EM, Franklin C, Nielsen SE, Strack M (2020) Seismic line disturbance alters soil physical and chemical properties across boreal forest and peatland soils. Front Earth Sci 8:281. https://doi.org/10.3389/feart.2020.00281
Davidson SJ, Goud EM, Malhotra A, Estey CO, Korsah P, Strack M (2021) Linear disturbances shift boreal peatland plant communities toward earlier peak greenness. JGR Biogeosci. https://doi.org/10.1029/2021JG006403
Echiverri LFI, Macdonald SE, Nielsen SE (2022) Neighboring edges: Interacting edge effects from linear disturbances in treed fens. In: R Ohlemuller (ed). Appl Veg Sci. https://doi.org/10.1111/avsc.12645
Elkin LA, Kay M, Higgins JJ, Wobbrock JO (2021) An aligned rank transform procedure for multifactor contrast tests. Softw Technol 34:754–768. https://doi.org/10.1145/3472749.3474784
Ficken CD, Cobbaert D, Rooney RC (2019) Low extent but high impact of human land use on wetland flora across the boreal oil sands region. Sci Total Environ 693:133647. https://doi.org/10.1016/j.scitotenv.2019.133647
Finnegan L, MacNearney D, Pigeon KE (2018) Divergent patterns of understory forage growth after seismic line exploration: Implications for caribou habitat restoration. For Ecol Manag 409:634–652. https://doi.org/10.1016/j.foreco.2017.12.010
Franklin CMA, Filicetti AT, Nielsen SE (2021) Seismic line width and orientation influence microclimatic forest edge gradients and tree regeneration. For Ecol Manag 492:119216. https://doi.org/10.1016/j.foreco.2021.119216
Gehlhausen SM, Schwartz MW, Augspurger CK (2000) Vegetation and microclimatic edge effects in two mixed-mesophytic forest fragments. Plant Ecol 147:21–35. https://doi.org/10.1023/A:1009846507652
Gignac LD, Dale MRT (2007) Effects of size, shape, and edge on vegetation in remnants of the upland boreal mixed-wood forest in agro-environments of Alberta, Canada. Can J Bot 85:273–284. https://doi.org/10.1139/B07-018
Godefroid S, Koedam N (2004) The impact of forest paths upon adjacent vegetation: effects of the path surfacing material on the species composition and soil compaction. Biol Conserv 119:405–419. https://doi.org/10.1016/j.biocon.2004.01.003
Government of Alberta (2012) Treaty boundary. https://open.alberta.ca/opendata/gda-bcf1b24c-16e9-4dc6-8729-c5096f302632. Accessed June 2023
Government of Alberta (2023a) Cold Lake - Oil Sands - Alberta's Energy Heritage. http://www.history.alberta.ca/energyheritage/sands/underground-developments/in-situ-development/cold-lake.aspx. Accessed June 2023
Government of Alberta (2023b) Air photos. https://www.alberta.ca/air-photos.aspx. Accessed July 2023
Government of Canada (2017) Historical climate data https://climate.weather.gc.ca/historical_data/search_historic_data_e.html. Accessed March 2023
Hansen MJ, Clevenger AP (2005) The influence of disturbance and habitat on the presence of non-native plant species along transport corridors. Biol Conserv 125:249–259. https://doi.org/10.1016/j.biocon.2005.03.024
Harper KA, Macdonald SE (2002) Structure and composition of edges next to regenerating clear-cuts in mixed-wood boreal forest. J Veg Sci 13:535–546. https://doi.org/10.1111/j.1654-1103.2002.tb02080.x
Harper KA, Macdonald SE, Burton PJ et al (2005) Edge influence on forest structure and composition in fragmented landscapes. Conserv Biol 19:768–782. https://doi.org/10.1111/j.1523-1739.2005.00045.x
Harper KA, Macdonald SE, Mayerhofer MS, Biswas SR, Esseen PA, Hylander K, Stewart KJ, Mallik AU, Drapeau P, Jonsson B, Lesieur D, Kouki J, Bergeron Y (2015) Edge influence on vegetation at natural and anthropogenic edges of boreal forests in Canada and Fennoscandia. J Ecol 103:550–562. https://doi.org/10.1111/1365-2745.12398
Harrar SW, Ronchi F, Salmaso L (2019) A comparison of recent nonparametric methods for testing effects in two-by-two factorial designs. J Appl Stat 46:1649–1670. https://doi.org/10.1080/02664763.2018.1555575
Honnay O, Verheyen K, Hermy M (2002) Permeability of ancient forest edges for weedy plant species invasion. For Ecol Manag 161:109–122. https://doi.org/10.1016/S0378-1127(01)00490-X
ITIS (2023) Integrated taxonomic information system. https://www.itis.gov/. Accessed March 2023
Jackson RS, Dennett JM, Nielsen SE (2023) Effects of oil sands disturbances on shrub and tree structure along forest edges in Alberta’s boreal forest. Can J for Res 00:1–12. https://doi.org/10.1139/cjfr-2023-0005
Kay M, Elkin L, Higgens J, Wobbrock J (2021) ARTool: aligned rank transform for nonparametric factorial ANOVAs. https://doi.org/10.5281/zenodo.594511
Lee MA, Davies L, Power SA (2012) Effects of roads on adjacent plant community composition and ecosystem function: an example from three calcareous ecosystems. Environ Pollut 163:273–280. https://doi.org/10.1016/j.envpol.2011.12.038
Legendre P, Gallagher ED (2001) Ecologically meaningful transformations for ordination of species data. Oecologia 129:271–280. https://doi.org/10.1007/s004420100716
Legendre P, Legendre L (2012) Numerical ecology, 3rd edn. Elsevier, Amsterdam
Lenth RV, Bolker B, Buerkner P, Giné-Vázquez I, Herve M, Jung M, Love J, Miguez F, Riebl H, Singmann H (2023) emmeans: estimated marginal means, aka least squares means. https://cran.r-project.org/web/packages/emmeans/index.html
Lieffers VJ (1995) Ecology and dynamics of boreal understoryspecies and their role in partial-cut silviculture. In: Bamsey CR (ed) Innovative silviculture systems in boreal forests. Clear Lake Ltd, Edmonton, pp 33–39
Lieffers VJ, Stadt KJ (1994) Growth of Picea glauca, Calamagrostis canadensis and Epilobium angustifolium in relation to overstory light transmission. Can J for Res 24:1193–1198. https://doi.org/10.1139/x94-157
Lovitt J, Rahman MM, Saraswati S, McDermid GJ, Strack M, Xu B (2018) UAV remote sensing can reveal the effects of low-impact seismic lines on surface morphology, hydrology, and methane (CH4) release in a boreal treed bog. J Geophys Res Biogeosci 123:1117–1129. https://doi.org/10.1002/2017JG004232
Mitchell AK, Koppenaal R, Goodmanson G, Benton R, Bown T (2007) Regenerating montane conifers with variable retention systems in a coastal British Columbia forest: 10-year results. For Ecol Manag 246:240–250. https://doi.org/10.1016/j.foreco.2007.04.036
Murcia C (1995) Edge effects in fragmented forests: implications for conservation. Trends Ecol Evol 10:58–62. https://doi.org/10.1016/S0169-5347(00)88977-6
Neher DA, Asmussen D, Lovell ST (2013) Roads in northern hardwood forests affect adjacent plant communities and soil chemistry in proportion to the maintained roadside area. Sci Total Environ 449:320–327. https://doi.org/10.1016/j.scitotenv.2013.01.062
Oksanen J, Simpson GL, Blanchet FG et al (2022) vegan: Community ecology package. https://CRAN.R-project.org/package=vegan
R Core Team (2023) R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. https://www.R-project.org/
Rao CR (1964) The use and interpretation of principal component analysis in applied research. Sankhya 26:329–358
Resasco J, Haddad NM, Orrock JL et al (2014) Landscape corridors can increase invasion by an exotic species and reduce diversity of native species. Ecology 95:2033–2039. https://doi.org/10.1890/14-0169.1
Roberts D, Ciuti S, Barber QE, Willier C, Nielsen SE (2018) Accelerated seed dispersal along linear disturbances in the Canadian oil sands region. Sci Rep 8:4828. https://doi.org/10.1038/s41598-018-22678-y
Ross KA, Fox BJ, Fox MD (2002) Changes to plant species richness in forest fragments: fragment age, disturbance and fire history may be as important as area. J Biogeogr 29:749–765. https://doi.org/10.1046/j.1365-2699.2002.00722.x
Scheffer RA, Van Logtestijn RSP, Verhoeven JTA (2001) Decomposition of Carex and Sphagnum litter in two mesotrophic fens differing in dominant plant species. Oikos 92:44–54. https://doi.org/10.1034/j.1600-0706.2001.920106.x
Šerá B (2010) Road-side herbaceous vegetation: life history groups and habitat preferences. Pol J Ecol 58:66–79
Society for Ecological Restoration (SER), International Network for Seed-based Restoration (INSR), Royal Botanic Gardens Kew (RBGK) (2023) Seed information database. https://ser-sid.org/. Accessed July 2023
Stern E, Riva F, Nielsen S (2018) Effects of narrow linear disturbances on light and wind patterns in fragmented boreal forests in northeastern Alberta. Forests 9:486. https://doi.org/10.3390/f9080486
Trombulak SC, Frissell CA (2000) Review of ecological effects of roads on terrestrial and aquatic communities. Conserv Biol 14:18–30. https://doi.org/10.1046/j.1523-1739.2000.99084.x
Venier LA, Thompson ID, Fleming R, Malcolm J, Aubin I, Trofymow JA, Langor D, Sturrock R, Patry C, Outerbridge RO, Holmes SB, Haeussier S, De Grandpré L, Chen HYH, Bayne E, Arsenault A, Brandt JP (2014) Effects of natural resource development on the terrestrial biodiversity of Canadian boreal forests. Enviro Rev 22:457–490. https://doi.org/10.1139/er-2013-0075
Willier CN, Dennett JM, Devito KJ, Bater CW, Nielsen SE (2022) The extent and magnitude of edge effects on woody vegetation in road-bisected treed peatlands in boreal Alberta, Canada. Ecohydrology. https://doi.org/10.1002/eco.2455
Wobbrock JO, Findlater L, Gergle D, Higgins JJ (2011) The aligned rank transform for nonparametric factorial analyses using only anova procedures. In: Proceedings of the SIGCHI Conference on human factors in computing systems. ACM, Vancouver, pp 143–146
Acknowledgements
We thank Philip Hoffman, Jessica Hudson, Kristen Schafer, Leah Crandall, Maeve Warren, Ivy Boddez, and Molly Sharp for field work assistance. We thank Laureen Echiverri for performing and reporting on preliminary data analysis and Celina Waldron for formatting the references and tables. We also thank Curtis Fedor and Michelle Young from Imperial Oil for providing site access and on-site support.
Funding
Open Access funding provided by Natural Resources Canada. This work was supported by the Office of Energy Research and Development (OERD), Natural Resources Canada (NRCan), and the Canadian Forest Service (CFS).
Author information
Authors and Affiliations
Contributions
AD, JP, DL, and JH contributed to the study conception and design. Data collection was performed by JH. The first draft of the manuscript was written by AD, JB, and KH. Statistical analyses were conducted by KH. Graphics were prepared by KH and JB. AD, KH, JH, JP, DL, and JB edited the manuscript, and JB finalized it.
Corresponding author
Ethics declarations
Competing interests
The authors have no relevant financial or non-financial interests to disclose.
Additional information
Communicated by Christina Rinas.
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Buss, J., Dabros, A., Higgins, K.L. et al. Comparison of edge effects from well pads and industrial roads on mixed upland boreal forest vegetation in Alberta. Plant Ecol 225, 331–343 (2024). https://doi.org/10.1007/s11258-023-01393-3
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11258-023-01393-3