
Abstract
Climate change will increase the frequency of droughts over the next century, with severe consequences for ecosystem function in semi-arid grasslands. The shortgrass steppe (SGS) experiences some of the largest interannual variation in precipitation among terrestrial biomes and exhibits extremely high sensitivity to drought. Yet despite decades of research describing the consequences of drought for ecosystem function in the SGS, we currently have little information regarding the impact of drought on bioavailability of important nutrients other than nitrogen, the contribution of herbivores to bioavailable concentrations of these nutrients, and whether drought alters herbivore-derived nutrient cycling. To quantify the impacts of long-term drought and small-bodied herbivores on nutrient cycling and aboveground net primary production (ANPP), we factorially manipulated rainfall and herbivore presence in the SGS of northern Colorado. Specifically, we measured the impacts of drought and herbivores on bioavailability of ten important nutrients: aluminum, calcium, iron, potassium, magnesium, manganese, nitrate, phosphorus, sulfur, and zinc. We then correlated these nutrients with grass production to determine whether reduced plant growth under drought conditions causes a belowground buildup of nutrients. Drought reduced ANPP as expected, and also altered concentrations of many nutrients apart from N, which clustered in their drought response. In contrast, small-bodied herbivores did not affect ANPP or soil N. However, they did contribute to the bioavailable soil concentrations of two important nutrients: PO4-P and S. Importantly, drought generally did not modify the contribution of herbivores to nutrient cycling, suggesting that herbivores might be a critical component of biogeochemical cycling regardless of precipitation in semi-arid grasslands.
Similar content being viewed by others

Introduction
Climate change will increase the frequency, duration, and intensity of droughts over the next century (IPCC 2014), with severe consequences for ecosystem function in arid and semi-arid grasslands. Dry ecosystems, such as the shortgrass steppe (SGS), undergo the largest interannual variation in precipitation among terrestrial biomes and exhibit the highest sensitivity to drought (Huxman et al. 2004a; Knapp et al. 2015b). For example, severe drought reduces aboveground net primary production (ANPP) by 50–75% in xeric grasslands (Knapp et al. 2015a), and this collapse of ANPP impacts a number of other important ecosystem functions. Plant nitrogen (N) uptake rates decline during drought as a consequence of lower growth demand, which in turn causes a buildup of bioavailable soil N accessible by plant roots (Evans and Burke 2013). Soil CO2 flux also generally declines during drought due to reduced soil microbial activity, thereby increasing the pool of belowground soil carbon in the SGS (Huxman et al. 2004b; Munson et al. 2010; Evans and Burke 2013). Low ANPP also prefaces long-term shifts in plant community composition, which persist for several years following severe drought (Rondeau et al. 2018). Yet despite decades of research describing the consequences of drought for ecosystem function in the SGS, several important knowledge gaps remain. We currently have little information regarding (1) the impact of drought on bioavailability of important nutrients other than N, (2) the contribution of herbivores to bioavailable concentrations of these nutrients, and (3) whether drought alters herbivore nutrient cycling in semi-arid grasslands.
Although N has received the most attention as a limiting nutrient in grassland ecosystems, ANPP and ecosystem function depend on several other important, but often overlooked, nutrients (Fay et al. 2015). Phosphorus (P) and potassium (K) can limit ANPP as strongly as N (Fay et al. 2015; Koerner et al. 2016), and both nutrients potentially affect grassland resilience to drought. For example, accumulation of soil P disrupts mycorrhizal-plant mutualisms, thereby weakening plant tolerance of water stress and destabilizing long-term ANPP (Augé 2001; Yang et al. 2014; Shantz et al. 2016). Potassium, on the other hand, can mitigate plant water stress during drought by acting as an antioxidant and facilitating stomatal regulation of water loss (Egilla et al. 2005). The minerals magnesium (Mg), iron (Fe), and zinc (Zn) are all integral components of photosynthesis, enabling capture of light energy and facilitating transport of light energy through the photosystem (Balakrishnan et al. 2000). Soil P and K concentrations decline during drought in forests (Sardans and Peñuelas 2007), and it is likely that nutrient availability in grasslands also depends on soil moisture and ANPP. Thus, drought-induced changes in any number of macro- or micronutrients can potentially impact ecosystem function, but the extents to which drought alters soil concentrations of P, K, or other micronutrients are unknown.
In addition to precipitation, nutrient cycling and ANPP in the SGS might be affected by the presence of herbivores. Herbivores reduce ANPP through direct consumptive effects (Bakker et al. 2006; La Pierre et al. 2015) or stimulate ANPP by converting refractory detritus into labile organic matter and bioavailable soil nutrients (Belovsky and Slade 2000; Nitschke et al. 2015). Unfortunately, herbivore-driven nutrient cycling in grasslands, specifically the contributions of invertebrate and small mammal herbivores, has received relatively little attention. Limited evidence hints that these herbivore groups might contribute substantially to soil nutrient availability. Soil N availability in tallgrass prairies doubles in the presence of large mammal grazers like cattle or bison (Bakker et al. 2009). Since bison biomass density (g m−2) in tallgrass prairies is comparable to that of grasshoppers (Branson et al. 2006), invertebrate herbivores represent a potentially large nutrient cycling pathway via frass deposition. Indeed, insect frass deposition accelerates N cycling by 42–600% in temperate forests (Reynolds et al. 2000; Frost and Hunter 2004) and accounts for over a quarter of N and P soil inputs in tropical forests (Metcalf et al. 2014). In a cold, mixed grass prairie, grasshoppers increased soil N concentrations by ~ 5% (Belovsky and Slade 2000). Many other nutrients [e.g., K, Mg, calcium (Ca)] also positively correlate with grasshopper density, and grasshopper frass contains high concentrations of N, P, and sulfur (S) (Joern et al. 2012; Nitschke et al. 2015). Yet the contribution of insects and small mammals to bioavailable soil nutrients is unknown in grasslands.
Furthermore, herbivore-dependent biogeochemical cycling might vary with abiotic conditions, such as drought (Irisarri et al. 2016). Rain limitation can alter the nutritional content of plants by decreasing plant water content, thereby increasing leaf toughness and reducing plant palatability (Avolio and Smith 2013; Koerner et al. 2014). Lower consumption rates result in lower frass and fecal production rates by insects and small mammals (Lemoine and Shantz 2016), such that herbivore contributions to biogeochemical cycling could be reduced under drought. Drought can also reduce decomposition and leaching rates of insect frass into soils. Indeed, Frost and Hunter (2004) reported that insect frass deposition in an oak forest must be temporally matched with precipitation events for herbivore nutrients to significantly contribute to soil nutrient pools. Given that soil biogeochemcial processes in the SGS largely depend on rainfall (Evans and Burke 2013), it is likely that drought mediates herbivore contributions to nutrient cycling in semi-arid grasslands. However, no study to date has manipulated both herbivory and rainfall to test how drought influences herbivore nutrient cycling.
Here, we factorially crossed growing season drought with herbivore removals to quantitate the impacts of long-term drought and small-bodied herbivores on nutrient cycling and ANPP in the SGS. We tested three hypotheses: (1) Drought should increase the pool of bioavailable nutrients related to plant growth (N, P, and K) and photosynthesis (Fe, Mn, and Zn) because of lower ANPP and nutrient uptake rates under water limitation; (2) herbivore presence will increase soil bioavailability of macronutrients, like N and P, but would have little effect on minerals that exhibit little herbivore turnover, like Mn and Fe; and (3) drought will reduce the importance of herbivore nutrient cycling because herbivore contributions to soil nutrient pools will be relatively small and therefore decrease in importance as baseline nutrient levels increase.
Methods
Study site
Our study took place in the Central Plains Experimental Range (CPER; 40.8°N, − 104.7°W), an experimental station situated in the SGS of northern Colorado. The SGS is a semi-arid grassland with low annual rainfall (mean annual precipitation: 375 mm; mean growing season precipitation: 293 mm) and primary production (mean annual ANPP: 88.9 g m−2) (Knapp et al. 2015a). Up to 90% of ANPP in the Colorado SGS derives from the perennial C4 grass Bouteloua gracilis (Oesterheld et al. 2001), although the graminoids B. dactyloides (C4), Carex spp. (C3), Elymus elymoides (C3), Pascopyrum smithii (C3), and Vulpia octoflora (C3), the forb Sphaeralcea coccinea, and the shrubs Artemisia frigida and Gutierrezia sarothrae also contribute substantially to ANPP.
Our study site at the CPER has not been grazed by cattle for over 15 years, and the primary herbivores in this pasture are invertebrates and small mammals. Grasshoppers (Orthoptera: Acrididae) are the dominant invertebrate herbivore by biomass in the SGS and consist of a diverse assemblage of species. Arphia pseudonietana, Eritettix simplex, Melanoplus sanguinipes, and Opeia obscura are the most abundant early- and late-season orthopterans in northern Colorado along the Front Range, occurring in densities as high as 10–20 individuals per 100 m2 during non-outbreak years (Welch et al. 1991; Craig et al. 1999). Small mammal herbivores are a combination of lagomorphs (Lepus californicus, Sylvilagus audubonii) and rodents (Dipodomys ordii, Onchyomys leucogaster) (Grant et al. 1982), although lagomorph feces were the most common indication of small mammal herbivory in the experimental plots. Pronghorn antelope (Antilocapra americana) are common in the SGS but were excluded from both control and treatment plots by cages.
Drought treatment
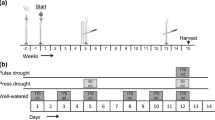
To simulate drought, we took advantage of a long-term rainfall exclusion experiment ongoing in the CPER since 2014. In 2014, twenty 6 × 6 m experimental plots were arrayed into ten blocks. Within each block, the two plots were randomly assigned to either ‘Ambient’ or ‘Drought’ treatments (n = 10 per precipitation treatment). The ‘Ambient’ plot was unmanipulated and received 100% of growing season precipitation each year. Growing season precipitation was reduced by 66% in the ‘Drought’ plots with a passive rainout shelter design (Yahdjian and Sala 2002). Each passive rainout shelter consisted of a 6 × 6 m cold frame greenhouse structure enclosing a 5 × 5 m measurement plot and covered by a roof made of 60 3.7 m × 15 cm clear polycarbonate strips. The plastic strips were evenly spaced across the roof to cover 66% of shelter area and angled to deliver intercepted rainfall into a gutters at the base of each side of the shelter roof. Gutters transported all intercepted rainfall several meters away from each plot via corrugated plastic tubes. All plots, including ‘Ambient’ plots, were trenched to a depth of 0.5 m and lined with plastic to prevent subsurface groundwater movement into the plot. Every year (2014–2017), we installed shelter roofs in May prior to green up and removed the roofs in September at the end of the growing season. This design was successful in reducing annual precipitation by ~ 50% and volumetric soil moisture by ~ 10% within the ‘Drought’ plots compared to ‘Ambient’ plots (Lemoine et al. in prep). Removing 50% of rainfall constitutes an extreme drought in the SGS (< 1% percentile).
In both ‘Ambient’ and ‘Drought’ plots, a 0.5 m buffer around the edge of the plot minimized rainfall blow-in into the central 5 × 5 m measurement plot. The measurement plot was further subdivided into four 2 × 2 m subplots bisected by 1 × 1 m buffer strips. Three randomly chosen subplots were designated sampling units in a different project, so herbivore manipulations in this study occurred within the remaining 2 × 2 m subplot.
Herbivore exclosures
We tested our hypotheses regarding the effects of herbivores on ecosystem function by comparing plots exposed to herbivores to plots protected from herbivores (Lemoine et al. 2017b). We excluded herbivores using cages constructed of a 0.5 × 0.5 × 0.5 m PVC frame. The frame was anchored in place with rebar spikes inside each hollow PVC leg and covered with 14 mm mesh aluminum screen. The window screen had a 20-cm skirt fastened tightly to the ground using common garden staples to prevent invertebrate herbivore immigration into cages. In May 2017, we haphazardly placed two cages in each 2 × 2 m subplot (n = 10 per drought × herbivore treatment) while attempting to minimize differences in plant species compositions between cages and avoiding cacti where possible. For each pair of cages, one cage was randomly designated the ‘No Herbivores’ treatment, and remained fully enclosed. The ‘Herbivores’ cage had large holes cut in the window screen to allow herbivore movement into the cage, while allowing the screen to mimic the reduction in light imposed by the ‘No Herbivores’ treatment (La Pierre and Smith 2016). Since both herbivore treatments were present in every subplot, this design was a split-plot design with precipitation (ambient vs. drought) as the whole-plot factor and herbivore presence (No herbivores vs. herbivores) as the subplot factor.
Every week throughout the growing season, we checked cages for damage and repaired window screen when needed. We also removed any invertebrates found within the ‘No Herbivores’ cages. However, only one grasshopper was ever found within an exclusion cage throughout the course of this experiment. Furthermore, small-bodied herbivores regularly occurred within the ‘Herbivores’ cages, as evidenced by small mammal dung (usually lagomorphs) regularly observed within the open cages.
Bioavailable nutrients
We quantified soil nutrient bioavailability in each drought × herbivore treatment using Plant Root Simulator (PRS) probes (Western Ag, Saskatoon, Saskatchewan, CA) (Evans and Burke 2013). PRS probes consist of a pair of plastic stakes (15 × 2 × 0.5 cm), each with a 17.5 cm2 charged ion membrane. The ionic membrane of one stake in the pair is made of a general purpose R-NH4+ cation base that absorbs anions. The other stake in the pair has a general purpose R-SO3− anion base to absorb cations. We buried two pairs of PRS probes within each cage in early August prior to the period of maximum insect activity (T. Joern—pers. comm.). Pairs of probes were buried at opposite corners of the cage by making a small slit in the soil surface with a garden knife. Probes were then inserted into each slit at a 30°–35° angle and gently driven into the ground using a rubber mallet. Loose soil was back-filled into the slit to maximize contact between the ion membrane and the soil. After 30 days, we removed probes from the cages and pooled the two probe pairs within a single cage for nutrient analysis. Nutrient analyses, conducted by Western Ag, provided uptake rates (μg 10 cm−2 day−1) for aluminum (Al), calcium (Ca), iron (Fe), potassium (K), magnesium (Mg), manganese (Mn), nitrate-derived nitrogen (NO3-N), phosphate-bound phosphorus (PO4-P), sulfur (S), and zinc (Zn).
ANPP sampling
We estimated annual ANPP in each plot via destructive biomass harvest at the end of the growing season. In late August, cages were removed, and a 20 × 50 cm (0.1 m2) metal frame was placed within the footprint of each cage. We harvested all biomass within the metal frame, sorted it by grass, forb, and woody functional types, and dried it at 60 °C for 48 h. Because the site is not annually burned, some harvested biomass from each cage was dead production from previous years. To avoid measuring past production, we removed any stems, shoots, or leaves with > 50% dead tissue. We weighed the remaining biomass to the nearest 0.1 g and calculated ANPP as g m−2 for each functional type.
Statistical analyses
We tested our hypothesis that drought should modify herbivore contributions to nutrient cycling using a split-plot MANOVA (Lemoine et al. 2017b). The MANOVA included all ten nutrients as a multivariate response matrix. The predictor variables were herbivore treatment and the drought × herbivore interaction at the subplot level, with the main effect of drought estimated at the whole-plot level. We used a similar split-plot ANOVA model to analyze the effects of drought and herbivore presence on forb and grass ANPP. Forb and grass ANPP were analyzed separately because six plots were missing forb data (n = 34), and a multivariate analysis would have required removing grass ANPP observations from those plots. Since grass and forb ANPP were only weakly correlated (r = − 0.16), we chose to use a univariate model to incorporate all available data. Forb data were log-transform prior to analysis to mitigate heteroscedasticity.
Many nutrient uptake rates were correlated, and it appeared that nutrients clustered by their response to drought. To better visualize the correlations among nutrients, we used principal component analysis to the reduced variation in soil nutrient bioavailability to two orthogonal axes. We then constructed a distance biplot of the first two principal component axes, overlaid with vectors of nutrient uptake rates. Distance biplots preserve the relationship among plots (and thus accurately representing separation among treatments), while slightly distorting the correlation among nutrient uptake rates. However, nutrient uptake rates clustered strongly in this case, such that any distortion in correlations among nutrient bioavailability had little impact on the results.
We hypothesized that soil nutrient bioavailability would be negatively correlated with ANPP. To test this hypothesis, we calculated the partial correlation coefficient between grass ANPP and nutrient uptake rates for all ten nutrients. We used MCMC sampling to generate a posterior distribution from 40,000 samples for the correlation matrix R containing the correlations among all variables. For each posterior sample, we calculated the partial correlation coefficients as:
where ri,j is the partial correlation coefficient between the ith and jth variables accounting for all other correlations. For example, rGrass,Al is the partial correlation between grass ANPP and Al uptake rates, accounting for any correlations between grass ANPP and all other nutrients and Al uptake rates and all other nutrients.
We conducted all ANOVAs and MANOVAs in a Bayesian framework so that we could place weakly informative priors of N(0,1) on all parameters (Lemoine et al. 2016a, b). Weakly informative priors constrain effect sizes to reasonable estimates in the presence of small samples sizes and weak effects, thereby making our tests more conservative but less prone to magnitude errors (Button et al. 2013; Gelman and Carlin 2014; Lemoine et al. 2016). Since all variables were standardized prior to analyses, the N(0,1) prior states that effect sizes should be smaller than one standard deviation of the response unless strongly supported by the data. Such a prior is more conservative, but reduces the probability of Type I errors for underpowered studies. To assess the impact of prior choice on our results, we repeated all analyses with noninformative priors, which yield quantitatively identical results to traditional statistics. Qualitative patterns of results did not depend on prior choice ("Appendix 1").
The correlation among responses and the variances of each response were given uninformative LKJ(1) and Cauchy(0, 25) priors, respectively. Models were allowed 10,000 warm-up iterations in four independent MCMC chains. We saved the next 10,000 estimates from each chain for posterior distributions of each parameter (40,000 total samples per parameter). Kernel density plots indicated good convergence among chains. Due to small sample sizes (n = 10 per precipitation × herbivore treatment) and our use of weakly informative priors, we consider any effect with Pr ≥ 0.90 as marginally significant and Pr ≥ 0.95 as statistically significant. A Pr = 0.5 is the lowest value because it indicates that the effect is just as likely to be positive or negative (i.e., centered at 0) (Lemoine and Shantz 2016; Lemoine et al. 2017a, b).
All statistical analyses were conducted in Python v3.6 using numpy, scipy, and pandas modules (Jones et al. 2001; McKinney 2010; van der Walt et al. 2011). Bayesian models were run with STAN v2.17 accessed by PySTAN (Stan Development Team 2016). Unless otherwise stated, all results are reported as mean ± 1 SE.
Results
Bioavailable nutrients
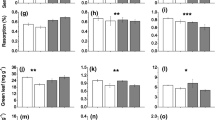
Bioavailable nutrient concentrations varied by several orders of magnitude among nutrients, indicative of considerable variability in nutrient standing stocks in the SGS. Calcium uptake rates, for example, ranged between 200 and 1000 μg 10 cm−2 day−1, while uptake rates of Zn and Mn rarely exceeded 2 μg 10 cm−2 day−1 (Fig. 1). Nutrient bioavailability was highest for alkali/alkaline earth metals Ca, K, and Mg, and lowest for transition metals Fe, Mn, and Zn (Fig. 1).

Nutrient uptake rates (μg 10 cm−2 day−1) of all ten nutrients in each of the drought × herbivore treatments. Note the different scale of each y-axis. Bars denote means ± 1 SE
Long-term drought imposed significant changes in soil nutrient bioavailability, although the direction of change depended on nutrient identity. Ca, K, Mg, NO3-N, PO4-P, and S all decreased uptake rates under drought (Fig. 1, Table 1), and reductions were most severe for Ca (− 54.5 ± 6.8%), Mg (− 61.0 ± 5.6%), and PO4-P (− 65.6 ± 7.8%). In contrast, drought increased uptake rates of both Mn and Zn by + 113.7 ± 231.7% and + 51.53 ± 28.5%, respectively (Fig. 1, Table 1). Neither Al nor Fe was affected by drought (Fig. 1, Table 1). Fe did respond to drought with noninformative priors (Appendix 1"), but given the small sample sizes used here, we prefer the more conservative estimates based on weakly informative priors.
Herbivore presence generally had weak effects on soil nutrient bioavailability, with three notable exceptions. Uptake rates of PO4-P increased with herbivore presence (+ 40.5 ± 21.0%, Fig. 1, Table 1). Likewise, S uptake rates also increased with herbivore presence, but to a lesser degree than did PO4-P (+ 17.5 ± 9.5%, Fig. 1, Table 1). Only Fe exhibited a moderate interaction between drought and herbivore presence (Table 1), suggesting that the effect of herbivore presence on Fe availability depends on rainfall. Under ambient rainfall conditions, herbivore presence decreased Fe uptake rates by ~ 20%, whereas herbivore presence increased Fe uptake rates by ~ 20% under drought conditions (Fig. 1, Table 1).
Principal components analysis revealed strong correlations among nutrient availability, such that the first two principal axes explained over 60% of the variance among nutrients (Fig. 2). Nutrients clustered into three groups along the first principal component, distinguished by their relative availability under drought conditions: nutrients with positive PC 1 loadings demonstrated decreased uptake rates during drought (Ca, K, Mg, NO3-N, PO4-P, S), nutrients with negative PC 1 loadings exhibited increased uptake rates during drought (Mn, Zn), and nutrients that were unrelated to PC 1 for which drought had little effect (Al, Fe). Indeed, PC 1 strongly demarcated plots by rainfall treatment, whereas plots did not cluster by herbivore presence along either PC 1 or PC 2. These results confirm that soil nutrient bioavailability is primarily regulated by rainfall in the SGS, although herbivores do regulate the availability of a small subset of nutrients (e.g., PO4-P, S).

Distance biplots of the first two principal component axes. Plots have been coded by drought × herbivore treatment. The lines are the convex hull enclosing all sites from their respective drought treatments
ANPP
Drought suppressed grass ANPP in the SGS by 32.1 ± 11.0% (Pr(Drought) = 0.99), but had no detectable effect on forb ANPP (Pr(Drought) = 0.61, Fig. 3). Herbivore presence, on the other hand, did not strongly affect either grass (Pr(Herbivores) = 0.71) or forb ANPP (Pr(Herbivores) = 0.53, Fig. 3). Grass ANPP was slightly lower in the presence of herbivores, but this effect was relatively small and highly variable (− 10.4 ± 16.3%), suggesting that insect and small mammal herbivores do not strongly suppress ANPP in this semi-arid system.

Aboveground net primary production (ANPP) for both forbs and grass (g m−2) in each of the drought × herbivore treatments. Note the different scales of each y-axis. Bars denote means ± 1 SE
Correlations between nutrient flux and ANPP
We hypothesized that bioavailable soil nutrient concentrations would be negatively correlated with ANPP due to reduced nutrient uptake by plants. However, many nutrients were positively correlated with ANPP (Table 2). Specifically, NO3-N, S, and Zn uptake rates all increased with ANPP (Pr(> 0) = 0.94, 0.91, 0 and 0.90, respectively). Only K demonstrated a marginally significant, negative correlation with grass ANPP (Pr(< 0) = 0.92), conforming to our initial hypotheses.
Discussion
Drought and herbivores are expected to independently impact ANPP and nutrient cycling in terrestrial SGS grasslands (Huxman et al. 2004b; Munson et al. 2010; Evans and Burke 2013). In our experiment, ANPP and nutrient cycling of many micro- and macronutrients were affected by growing season drought. Interestingly, drought did not have a consistent directional impact on nutrient cycling. Of the ten measured nutrient uptake rates, six decreased, two increased, and two did not change under drought. In contrast, small-bodied herbivores, including both insects and small mammals, did not affect ANPP or soil N in our study. However, they did contribute to the bioavailable soil concentrations of two important nutrients: PO4-P and S. Importantly, drought did not modify the contribution of herbivores to nutrient cycling, suggesting that herbivores might be a critical component of biogeochemical cycling regardless of precipitation in semi-arid grasslands.
Semi-arid grasslands experience high interannual variability in precipitation, making these ecosystems vulnerable to extreme drought (Knapp et al. 2015b). The drought of 2012, for example, caused a 40% decline in annual precipitation throughout US grasslands, resulting in a 50% reduction in ANPP in the SGS. We found a similarly strong effect; ANPP declined by 32.1 ± 11.0% under a 66% reduction in growing season precipitation. However, our study contradicts previous work examining the effects of drought on bioavailable soil nutrients. Evans and Burke (2013) found that drought increased resin-captured NO3-N by 200–400%, whereas we documented a 22.0 ± 17.5% decline in bioavailable NO3-N. This discrepancy might be due to intra- or interannual variation in soil N pools, which can vary by an order of magnitude among years and between early- and late-season periods within a year (Evans and Burke 2013). Other nutrients showed drought sensitivities similar to those reported in other ecosystems. In Mediterranean forests and shrublands, total P concentrations increase during drought, but this increase is caused by an accumulation of plant-unavailable organic P and a decrease in plant-available inorganic P (Sardans and Peñuelas 2004, 2007; Sardans et al. 2008). Inorganic P (PO4-P) declined during drought in our study as well, but did so more severely than in forests. Soil K availability also declined more severely under drought in this study compared to other ecosystems (Sardans and Peñuelas 2007; Sardans et al. 2008). Given the large reductions, or in some cases increases, in soil nutrient bioavailability reported here compared to other ecosystems, semi-arid grasslands like the SGS might be more sensitive to drought in multiple aspects of ecosystem function beyond ANPP.
Changes in soil nutrient availability during drought potentially initiate feedback loops that make semi-arid grasslands more or less susceptible to drought in the future. High P concentrations, for example, disrupt the mycorrhizal-plant mutualism, making plants less tolerants of water stress (Augé 2001; Shantz et al. 2016). Drought-induced declines in soil P might therefore strengthen the mycorrhizal association and assist plants in avoiding drought stress in the future. However, it is unlikely that ambient soil P concentrations in the SGS exceed the threshold required to shut down mycorrhizal mutualisms. Instead, P deficiency under drought conditions could further reduce leaf water potential and water-use efficiency of many plant species, exacerbating the negative effects of water stress (Wu et al. 2018). Likewise, K hinders the formation of reactive oxygen species and protects chloroplasts from damage, such that reduced K concentrations are associated with low photosynthetic rates (Cakmak 2005). Photosynthesis also suffers under sulfur deficiency because sulfur-based proteins comprise nearly 10% of the thykaloid membrane of chloroplasts (Maréchal et al. 1997). Numerous other mineral nutrients help plants maintain physiological function under water stress, including Fe and Mg which are both integral to the photosystem, and Ca, which provides antioxidant protection during abiotic stress (Jiang and Huang 2001). Any drought-imposed reduction in soil concentrations of these important mineral nutrients could potentially make plants more susceptible to drought in the future. Legacy effects are common in the SGS (Sala et al. 1982) and in grasslands worldwide (Sala et al. 2012), and it is possible that drought-induced changes in soil nutrient availability represent an underappreciated legacy effect that might influence the ability of a system to tolerate or recover from drought in the future.
Herbivores might also enhance ecosystem drought tolerance via herbivore nutrient cycling, but the magnitude of herbivore contributions to SGS soil nutrient bioavailability, up to now, remains largely unexplored. Bakker et al. (2009) reported no effect of large mammals on bioavailable N in the SGS, and insect herbivores increased soil N concentrations by only 5% in a dry, cool-season grassland (Belovsky and Slade 2000). These results, coupled with the absence of herbivore effects reported here, suggests that N cycling is largely decoupled from herbivore presence in the SGS. In forests, however, insect frass alone increases soil bioavailable N by 15–500%, likely due to the greater density of insect herbivores and higher frass throughfall (Reynolds et al. 2000; Frost and Hunter 2004; Metcalf et al. 2014). Phosphorus and S in throughfall are also elevated by 200–800% in the presence of insect herbivores (Nitschke et al. 2015). Therefore, herbivore nutrient cycling could be important for other minerals in semi-arid grasslands. In the SGS, herbivores increased soil bioavailable P by nearly 50% and S by almost 20%, more than double the contribution of insects to P deposition in forests (Fonte and Schowalter 2005; Metcalf et al. 2014). Given the importance of P as a determinant of grassland community composition (Koerner et al. 2016), and the roles of P and S in plant photosynthesis and drought tolerance (Maréchal et al. 1997; Wu et al. 2018), herbivores might be an integral component of ecosystem stability and drought tolerance in the face of high interannual rainfall variability.
The magnitude of herbivore nutrient cycling depends, however, on the density of herbivores present at a given site in a given year (Schade et al. 2003; Burkepile et al. 2013). Small-bodied herbivore populations in semi-arid grasslands exhibit extreme interannual fluctuations in population densities. Grasshopper densities, for example, vary three- to tenfold over a relatively short time period (Capinera and Thompson 1987; Belovsky and Slade 1995), and rodent population sizes can oscillate over two orders of magnitude from year to year (Garsd and Howard 1981; Brady and Slade 2004). Since herbivore population sizes in semi-arid systems typically track annual precipitation and the concomitant increase and decrease of vegetative cover (Capinera and Horton 1989), the magnitude of herbivore nutrient cycling probably also depends, to some extent, on annual precipitation. Unfortunately, this study was limited to a single growing season, and we are therefore unable to determine the environmental constraints on herbivore nutrient cycling. However, our study occurred during a year of below-average annual and growing season precipitations (293 mm and 221 mm, respectively). We may have therefore underestimated the average, long-term contribution of small herbivores to nutrient cycling in the SGS, and multiyear studies that encompass drier and wetter years provide an opportunity for future research.
Increased frequency and intensity of droughts will no doubt adversely affect ecosystem function of the SGS ecosystem. Numerous previous studies have reported the consequences of rainfall shortages on ANPP, soil N, and soil CO2 fluxes, and here we show for the first time that extreme drought depletes the belowground availability of other important nutrients. Many of these minerals enhance the drought tolerance of plants, and the reduced availability of these nutrients might impose significant drought legacies, initiating a negative feedback loop during recovery or recurrent drought events. Herbivores might be able to offset some of the negative impacts of drought be enhancing bioavailable concentrations of several important nutrients. Thus, our research demonstrates that incorporating consumer effects on nutrient cycles of minerals important to plant growth and drought response could help elucidate the consequences of single or recurrent droughts in semi-arid ecosystems.
References
Augé RM (2001) Water relations, drought and vesicular-arbuscular mycorrhizal symbiosis. Mycorrhiza 11:3–42
Avolio ML, Smith MS (2013) Mechanisms of selection: phenotypic differences among genotypes explain patterns of selection in dominant species. Ecology 94:953–965
Bakker ES, Ritchie ME, Olff H, Milchunas DG, Knops JMH (2006) Herbivore impact on grassland plant diversity depends on habitat productivity and herbivore size. Ecol Lett 9:780–788
Bakker ES, Knops JMH, Milchunas DG, Ritchie ME, Olff H (2009) Cross-site comparison of herbivore impact on nitrogen availability in grasslands: the role of plant nitrogen concentration. Oikos 118:1613–1622
Balakrishnan K, Rajendran C, Kulandaivelu G (2000) Differential responses of iron, magnesium, and zinc deficiency on pigment composition, nutrient content, and photosynthetic activity in tropical fruit crops. Photosynthetica 38:477–479
Belovsky GE, Slade JB (1995) Dynamics of two Montana grasshopper populations: relationships among weather, food abundance and intraspecific competition. Oecologia 101:383–396
Belovsky GE, Slade JB (2000) Insect herbivory accelerates nutrient cycling and increases plant production. Proc Natl Acad Sci USA 97:14412–14417
Brady MJ, Slade NA (2004) Long-term dynamics of a grassland rodent community. J Mammal 85:552–561
Branson DH, Joern A, Sword GA (2006) Sustainable management of insect herbivores in grassland ecosystems: new perspectives in grasshopper control. Bioscience 56:743
Burkepile DE, Allgeier JE, Shantz AA, Pritchard CE, Lemoine NP, Bhatti LH, Layman CA (2013) Nutrient supply from fishes facilitates macroalgae and suppresses corals in a Caribbean coral reef ecosystem. Sci Rep 3:1493
Button KS, Ioannidis JPA, Mokrysz C, Nosek BA, Flint J, Robinson ESJ, Munafò MR (2013) Power failure: why small sample size undermines the reliability of neuroscience. Nat Rev Neurosci 14:365–376
Cakmak I (2005) The role of potassium in alleviating detrimental effects of abiotic stresses in plants. J Plant Nutr Soil Sci 168:521–530
Capinera JL, Horton DR (1989) Geographic variation in effects of weather on grasshopper infestation. Environ Entomol 18:8–14
Capinera JL, Thompson DC (1987) Dynamics and structure of grasshopper assemblages in shortgrass prairie. Can Entomol 119:567–575
Craig DP, Bock CE, Bennett BC, Bock JH (1999) Habitat relationships among grasshoppers (Orthoptera: Acrididae) at the western limit of the Great Plains in Colorado. Am Midl Nat 142:314–327
Egilla JN, Davies FT, Boutton TW (2005) Drought stress influences leaf water content, photosynthesis, and water-use efficiency of Hibiscus rosa-sinensis at three potassium concentrations. Photosynthetica 43:135–140
Evans SE, Burke IC (2013) Carbon and nitrogen decoupling under an 11-year drought in the shortgrass steppe. Ecosystems 16:20–33
Fay PA, Prober SM, Harpole WS, Knops JMH, Bakker JD, Borer ET, Lind EM, MacDougall AS, Seabloom EW, Wragg PD, Adler PB, Blumenthal DM, Buckley YM, Chu C, Cleland EE, Collins SL, Davies KF, Du G, Feng X, Firn J, Gruner DS, Hagenah N, Hautier Y, Heckman RW, Jin VL, Kirkman KP, Klein J, Ladwig LM, Li Q, McCulley RL, Melbourne BA, Mitchell CE, Moore JL, Morgan JW, Risch AC, Schütz M, Stevens CJ, Wedin DA, Yang LH (2015) Grassland productivity limited by multiple nutrients. Nat Plants 1:1–5
Fonte SJ, Schowalter TD (2005) The influence of a neotropical herbivore (Lamponius portoricensis) on nutrient cycling and soil processes. Oecologia 146:423–431
Frost CJ, Hunter MD (2004) Insect canopy herbivory and frass deposition affect soil nutrient dynamics and export in oak mesocosms. Ecology 85:3335–3347
Garsd A, Howard WE (1981) A 19-year study of microtine population fluctuations using time-series analysis. Ecology 62:930–937
Gelman A, Carlin JB (2014) Beyond power calculations: assessing Type S (Sign) and Type M (Magnitude) errors. Perspect Psychol Sci 9:641–651
Grant WE, Birney EC, French NR, Swift DM (1982) Structure and productivity of grassland small mammal communities related to grazing-induced changes in vegetative cover. J Mammal 63:248–260
Huxman TE, Smith MD, Fay PA, Knapp AK, Shaw MR, Loik ME, Smith SD, Tissue DT, Zak JC, Weltzin JF, Pockman WT, Sala OE, Haddad BM, Harte J, Koch GW, Schwinning S, Small EE, Williams DG (2004a) Convergence across biomes to a common rain-use efficiency. Nature 429:651–654
Huxman TE, Snyder KA, Tissue D, Leffler AJ, Ogle K, Pockman WT, Sandquist DR, Potts DL, Schwinning S (2004b) Precipitation pulses and carbon fluxes in semiarid and arid ecosystems. Oecologia 141:254–268
IPCC (2014) Summary for policymakers. In: Field CB, Barros V, Stocker TF, Qin D, Dokken DJ, Ebi KL, Mastrandrea MD, Mach KJ, Plattner G-K, Allen SK, Tignor M, Midgley PM (eds) Managing the risks of extreme events and disasters to advance climate change adaptation A species report of working groups I and II of the Intergovernmental Panel on Climate Change. Cambridge University Press, Cambridge, pp 1–19
Irisarri JGN, Derner JD, Porensky LM, Augustine DJ, Reeves JL, Mueller KE (2016) Grazing intensity differentially regulates ANPP response to precipitation in North American semiarid grasslands. Ecol Appl 26:1370–1380
Jiang Y, Huang B (2001) Effects of calcium on antioxidant activities and water relations associated with heat tolerance in two cool-season grasses. J Exp Bot 52:341–349
Joern A, Provin T, Behmer ST (2012) Not just the usual suspects: insect herbivore populations and communities are associated with multiple plant nutrients. Ecology 93:1002–1015
Jones E, Oliphant TE, Peterson P, et al (2001) SciPy: open source scientific tools for Python. www.scipy.org
Knapp AK, Carroll CJW, Denton EM, La Pierre KJ, Collins SL, Smith MD (2015a) Differential sensitivity to regional-scale drought in six central US grasslands. Oecologia 177:949–957
Knapp AK, Hoover DL, Wilcox KR, Avolio ML, Koerner SE, La Pierre KJ, Loik ME, Luo Y, Sala OE, Smith MD (2015b) Characterizing differences in precipitation regimes of extreme wet and dry years: implications for climate change experiments. Glob Change Biol 21:2624–2633
Koerner SE, Collins SL, Blair JM, Knapp AK, Smith MD (2014) Rainfall variability has minimal effects on grassland recovery from repeated grazing. J Veg Sci 25:36–44
Koerner SE, Avolio ML, La Pierre KJ, Wilcox KR, Smith MD, Collins SL, Fridley JD (2016) Nutrient additions cause divergence of tallgrass prairie plant communities resulting in loss of ecosystem stability. J Ecol 104:1478–1487
La Pierre KJ, Smith MD (2016) Soil nutrient additions increase invertebrate herbivore abundances, but not herbivory, across three grassland systems. Oecologia 180:485–497
La Pierre KJ, Joern A, Smith MD (2015) Invertebrate, not small vertebrate, herbivory interacts with nutrient availability to impact tallgrass prairie community composition and biomass. Oikos 124:842–850
Lemoine NP, Shantz AA (2016) Increased temperature causes protein limitation by reducing the efficiency of nitrogen digestion in the ectothermic herbivore Spodoptera exigua. Physiol Entomol 41:143–151
Lemoine NP, Hoffman AM, Felton AJ, Baur L, Chaves F, Gray J, Yu Q, Smith MD (2016) Underappreciated problems of low replication in ecological field studies. Ecology 97:2554–2561
Lemoine NP, Dietrich JD, Smith MD (2017a) Precipitation and environmental constraints on three aspects of flowering in three dominant tallgrass species. Funct Ecol 31:1894–1902
Lemoine NP, Doublet D, Salminen J-P, Burkepile DE, Parker JD (2017b) Responses of plant phenology, growth, defense, and reproduction to interactive effects of warming and insect herbivory. Ecology 98:1817–1828
Maréchal E, Block MA, Dorne AJ, Douce R, Joyard J (1997) Lipid synthesis and metabolism in the plastid envelope. Physiol Plant 100:65–77
McKinney W (2010) Data structures for statistical computing in Python. In: Proceedings of 9th Python science conference, pp 51–56
Metcalf DB, Asner GP, Martin RE, Espejo JES, Huasco WH, Amézquita FFF, Carranza-Jimenez L, Cabrera DFG, Baca LD, Sinca F, Quispe LPH, Taype IA, Mora LE, Dávila AR, Solórzano MM, Vilca BLP, Román JML, Bustios PCG, Revilla NS, Tupayachi R, Girardin CAJ, Doughty CE, Malhi Y (2014) Herbivory makes major contributions to ecosystem carbon and nutrient cycling in tropical forests. Ecol Lett 17:324–332
Munson SM, Benton TJ, Lauenroth WK, Burke IC (2010) Soil carbon flux following pulse precipitation events in the shortgrass steppe. Ecol Res 25:205–211
Nitschke N, Wiesner K, Hilke I, Eisenhauer N, Oelmann Y, Weisser WW (2015) Increase of fast nutrient cycling in grassland microcosms through insect herbivory depends on plant functional composition and species diversity. Oikos 124:161–173
Oesterheld M, Loreti J, Semmartin M, Sala OE (2001) Inter-annual variation in primary production of a semi-arid grassland related to previous-year production. J Veg Sci 12:137–142
Reynolds BC, Hunter MD, Crossley DA (2000) Effects of canopy herbivory on nutrient cycling in a northern hardwood forest. Selbyana 21:74–78
Rondeau RJ, Decker KL, Doyle GA (2018) Potential consequences of repeated severe drought for shortgrass steppe species. Rangel Ecol Manag 71:91–97
Sala OE, Lauenroth WK, Parton WJ (1982) Plant recovery following prolonged drought in a shortgrass steppe. Agric Meteorol 27:49–58
Sala OE, Gherardi LA, Reichmann L, Jobbagy E, Peters DPC (2012) Legacies of precipitation fluctuations on primary production: theory and data synthesis. Philos Trans R Soc B Biol Sci 367:3135–3144
Sardans J, Peñuelas J (2004) Increasing drought decreases phosphorus availability in an evergreen Mediterranean forest. Plant Soil 267:367–377
Sardans J, Peñuelas J (2007) Drought changes phosphorus and potassium accumulation patterns in an evergreen Mediterranean forest. Funct Ecol 21:191–201
Sardans J, Peñuelas J, Prieto P, Estiarte M (2008) Drought and warming induced changes in P and K concentration and accumulation in plant biomass and soil in a Mediterranean shrubland. Plant Soil 306:261–271
Schade JD, Kyle M, Hobbie SE, Fagan WF, Elser JJ (2003) Stoichiometric tracking of soil nutrients by a desert insect herbivore. Ecol Lett 6:96–101
Shantz AA, Lemoine NP, Burkepile DE (2016) Nutrient loading alters the performance of key nutrient exchange mutualisms. Ecol Lett 19:20–28
Stan Development Team (2016) Stan: a C++ library for probability and sampling, version 2.14. http://www.mc-org/
van der Walt S, Colbert SC, Varoquaux G (2011) The NumPy array: a structure for efficient numerical computation. Comput Sci Eng 13:22–30
Welch JL, Redak R, Kondratieff BC (1991) Effect of cattle grazing on the density and species of grasshoppers (Orthopter: Acrididae) of the Central Plains Experimental Range, Colorado: a reassessment after two decades. J Kansas Entomol Soc 64:337–343
Wu ZZ, Ying YQ, Bin Zhang Y, Bi YF, Wang AK, Du XH (2018) Alleviation of drought stress in Phyllostachys edulis by N and P application. Sci Rep 8:1–9
Yahdjian L, Sala OE (2002) A rainout shelter design for intercepting different amounts of rainfall. Oecologia 133:95–101. https://doi.org/10.1007/s00442-002-1024-3
Yang G, Liu N, Lu W, Wang S, Kan H, Zhang Y, Xu L, Chen Y (2014) The interaction between arbuscular mycorrhizal fungi and soil phosphorus availability influences plant community productivity and ecosystem stability. J Ecol 102:1072–1082
Acknowledgements
We are very grateful for the dedicated and hardworking undergraduates of Colorado State: Abigail Lock, Madison Rode, Holly Perretta, Megan Coyle, Abby Lathrop-Melting, and Sam Rollman, who helped construct cages in a subfreezing April snowstorm, harvest biomass around cacti, and sort dried plant material in the lab. We also thank the EDGE crew: John Dietrich, Maddie Shields, Lauren Bauer, Ava Hoffman, Mao Wei, Tyler Roberts, Gene Halsey, Elsie Denton, and Melissa Johnson, who were responsible for maintaining the drought shelters each year and who helped us deploy insect cages. This work was supported by a United States Department of Agriculture—National Institute of Food and Agriculture—Agriculture and Food Research Initiative postdoctoral fellowship (Grant No. 2016-67012-25169) and an NSF DEB award (1754124) to NPL, as well as an NSF DEB Macrosystems grant to MDS.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by Laura Yahdjian.
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Appendix 1: Comparison of priors for Bayesian models
Appendix 1: Comparison of priors for Bayesian models
Weakly informative priors constrain large effect sizes in the presence of lower statistical power (i.e., small sample sizes, noisy data). These priors are therefore more conservative than noninformative priors, which are analogous to conducting frequentist statistics (the p-values and confidence intervals are identical between noninformative priors and frequentist methods). To assess the impact of prior choice on model output, we conducted all analyses with both weakly informative [N(0,1)] and noninformative [N(0,10000)] on regression coefficients.
MANOVA of nutrient availability
Results from weakly informative priors were qualitatively similar to, but slightly weaker than, noninformative priors. This is to be expected, due to the ability of N(0,1) priors to constrain effect sizes. The biggest difference occurred for Fe, where weakly informative priors negated marginally significant main effects of herbivory and drought and reduced the significant interaction to marginal significance. Thus, the weakly informative priors were more conservative then traditional statistics.
ANOVA of ANPP
Rights and permissions
About this article

Cite this article
Lemoine, N.P., Smith, M.D. Drought and small-bodied herbivores modify nutrient cycling in the semi-arid shortgrass steppe. Plant Ecol 220, 227–239 (2019). https://doi.org/10.1007/s11258-019-00908-1
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11258-019-00908-1