
Abstract
Humans can derive enormous benefit from the natural environment and the wildlife they see there, but increasing human use of natural environments may negatively impact wildlife, particularly in urban green spaces. Few studies have focused on the trade-offs between intensive human use and wildlife use of shared green spaces in urban areas. In this paper, we investigate the impacts of humans and their dogs on wildlife within an urban green space using camera trap data from Hampstead Heath, London. Spatial and temporal activity of common woodland bird and mammal species were compared between sites with low and high frequency of visits by humans and dogs. There was no significant difference in the spatial or temporal activity of wildlife species between sites with lower and higher visitation rates of humans and dogs, except with European hedgehogs (Erinaceus europaeus) which showed extended activity in the mornings and early evenings in sites with lower visitation rates. This may have implications for the survival and reproductive success of European hedgehogs. Our results suggest that adaptation to human and dog activity deserves greater study in urban green spaces, as would a broader approach to measuring possible anthropogenic effects.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
Enjoyment of green spaces in urban areas is important to mental health and wellbeing, and a key aspect of this effect is the interaction with nature and wildlife (Soulsbury and White 2015; Triguero-Mas et al. 2015). However, there is growing evidence that recreational use can be a form of human disturbance affecting the abundance, diversity and behaviour of wild species (Larson et al. 2016; Gaynor et al. 2018). Understanding how wildlife is impacted by human disturbance in urban green spaces will allow us to develop solutions that minimise negative impacts. This will benefit both wildlife and people, who will experience a greater abundance and diversity of wildlife.
While there is widespread evidence that recreation may have a negative impact on wild animal activity and abundance (Larson et al. 2016; Gaynor et al. 2018), most studies to date have focused on non-urban settings. It could be argued that negative effects will be particularly apparent in urban settings, especially if wildlife populations there have more limited space and resources, and experience more intense and continuous human disturbance. However, wildlife in urban ecosystems may also be more adapted to, and tolerant of, human presence. There is evidence, for instance, that wild birds forage less in city parks with greater human activity (Fernández-Juricic and Tellería 2000; Fernández-Juricic et al. 2001), but also evidence that the distance at which birds take flight when approached is less in urban than rural areas (Matsyura et al. 2015; Bötsch et al. 2018). Some urban wildlife may be adapted to anthropogenic sources of food (Storch and Leidenberger 2003; Ditchkoff et al. 2006), which could make them more likely to be seen in areas of high human activity. The effect of human activity on the presence and enjoyment of urban wildlife may therefore be complex and species-specific.
To explore this, we investigate the impact of human activity on a range of diurnal and nocturnal wildlife species on Hampstead Heath, a large, continuously accessible and intensively visited green space in central London, UK. Our study exploits the fact that different parts of the Heath’s 275 ha experience very different levels of local human activity, due to the distribution of footpaths and fencing. Many people who use Hampstead Heath walk dogs, usually off-lead. Because there is evidence that dogs, like humans, can have a negative impact on wildlife activity (Fernández-Juricic and Tellería, 2000; Dowding et al. 2010; Banks and Bryant 2007; Reilly et al. 2017), our study considers the combined effect of humans and their dogs on the presence and temporal activity of bird and mammal species which use the Heath’s woodlands.
We used camera traps to examine the effect of human and dog visits on wildlife activity. Camera traps provide a unique opportunity to understand small scale patterns in temporal and spatial overlap and interactions across species within vertebrate communities (Rowcliffe and Carbone 2008) and are increasingly used at a range of scales from local communities to global-scale meta-analyses (Steenweg et al. 2016). For this study, we utilised images from a 2018 camera trap survey set up to study European hedgehogs by the London HogWatch Project (Carbone and Cates, 2018). This survey captured activity of species foraging on woodlands based on a systematic grid-based camera survey (see Rowcliffe et al. 2014). By recording continuous photographic data across multiple independently selected woodland sites, we were able to obtain unbiased spatial and temporal information on wildlife occurrence and activity in areas that differed in levels of human and dog visitation.
Methods
Study area
Hampstead Heath is a 275 ha urban green space, which is listed as a site of Metropolitan Importance for Nature Conservation and situated in a densely populated part of Central London (City of London 2019). Hampstead Heath comprises a mosaic of habitats including woodland, grassland, scrub and open water, which are managed to maintain a biodiverse landscape while ensuring access to visitors (City of London 2010; City of London 2019). Local wildlife surveys show woodlands, including an ancient woodland designated as a Site of Special Scientific Interest (Natural England 1990), to be particularly rich in breeding bird and mammal species (City of London, unpublished). These woodlands are criss-crossed by a network of walking paths used by visitors. A significant proportion of visitors bring dogs to Hampstead Heath, usually off-lead, which is typical across green spaces in England (Taylor et al. 2005).
Data collection
Between 18th April and 5th July 2018, camera traps were uniformly distributed over Hampstead Heath across 150 sites of various habitat types using a 150 m2 cell-size grid overlay (Monterroso et al. 2013). Each camera trap operated on average for 15 days (range: 9—23 days) and recorded continuously 24 h a day. Survey lengths varied due to limited storage on memory cards, batteries running out of charge and camera failures. Camera traps were attached 20–50 cm above the ground to suitable features nearest to the ideal coordinates, for example fence posts or trees. Traps were angled slightly downwards to increase the probability of recording wildlife. Traps minimised disturbance by passively detecting wildlife using heat signatures (Welbourne et al. 2016) and were not baited. 104 camera traps across four models were used: 64 Browning Strike Force HD-Pro BTC-131 5HDPs, 20 Reconyx HyperFire HC-500 s, 10 Reconyx Hyperfire HC-600 s and 10 Reconyx Hyperfire PC-800 s. The Browning model took one image per trigger with a recovery time of approximately 0.6 s, while the Reconyx models took ten images per trigger with a recovery time of one second between photos (specifications for these cameras can be found at www.trailcampro.com).
Camera site selection
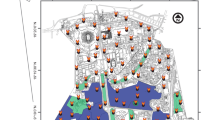
For the purpose of this study, a subset of 26 camera traps were selected from woodland sites across Hampstead Heath which recorded wildlife between 18th April and 10th June 2018 (Fig. 1; Online Resource-Table 1). Camera traps were selected from areas with the vegetation classification: “at least 84% broadleaved woodland” (City of London 2009; Online Resource-Table 2). One camera trap was selected from a site classified as “semi-neutral improved grassland” but was located in a broadleaved woodland copse within that area with a similar composition to the other selected woodland sites. Broadleaved woodland sites were dominated by English oak (Quercus robur) and less frequently by common beech (Fagus sylvatica) and sessile oak (Quercus petraea), with an understory of bramble (Rubus fruticosus), holly (Ilex aquifolium) and other small trees, and a ground cover of common ivy (Hedera helix) or bracken (Pteridium aquilinum) (See Online Resource-Table 2 for full habitat descriptions by site). Care was taken to select woodland sites with similar plant composition and microhabitats in order to reduce the confounding effects of factors like food availability and ground cover. Within this habitat, all camera traps were positioned to give a clear view of open leaf litter to at least five metres with little ground cover under a closed canopy. This improved the detection probability for all wildlife species, especially European hedgehogs which can go undetected in tall vegetation (Sollmann et al. 2013). Importantly, the distribution of cameras across the Heath meant some were close to, and others distant from, foot paths. Some were in the few fenced off woodland areas on the Heath, which people and dogs rarely visited. This created a broad range in frequency of human and dog visits recorded between traps.

Map showing the distribution of the 26 camera trap sites across Hampstead Heath, the camera trap model utilised at each site and whether the site was designated as having low or high visits of human and dogs
Photo review
All vertebrate species within camera images were manually identified using ExifPro v2.1 (Kowalski 2013). When an animal enters a camera’s field of view, the camera triggers and takes a series of successive images of the animal. For this study, we measured discrete visits to the site by individuals of particular species, which we call “contacts”. Contacts are indicated by successive images showing an individual appearing, being present, and then disappearing from the camera’s field of view. A new contact was recorded if eleven seconds had elapsed between successive images of that particular species. This removed differences between cameras in the way in which they recorded animals as they entered the camera field. Eleven seconds was chosen as it would take approximately nine seconds for each camera model to take ten successive photos of an animal that was continuously in the camera view. Considering the camera's recovery times, adding two seconds ensured that the next image was of an individual of that species coming newly into the field of a view, and not a continuation of the current visit.
Wildlife selection
Across the 26 camera traps, images of 18 distinct species of wildlife were captured, in addition to images of humans and dogs. Cats were recorded only twice and excluded from analysis. Some species of wildlife were particularly common (Table 1): diurnal wildlife species included a range of ground-foraging birds and grey squirrels (Sciurus carolinensis), while nocturnal species included mostly red foxes (Vulpes vulpes) and European hedgehogs (Erinaceus europaeus). For our analysis of wildlife activity, we put species in groups (‘All Birds’ and ‘Non-diurnal Mammals’) and also investigated individual species where we had enough contacts. A minimum of 25 contacts was required at both low visit sites and high visit sites (see site definition below) for a wildlife species to be included in the analysis, as this was the minimum needed to generate an activity schedule for the temporal analysis.
More frequently observed species were also better represented over a range of sites with different levels of humans and dog visits. Based on total contacts, we selected four bird species for individual analysis: common blackbirds (Turdus merula), Eurasian magpies (Pica pica), song thrushes (Turdus philomelos), and wood pigeons (Columba palumbus). We also created for analysis the category “All Birds” in order to incorporate these species and the individually rare but collectively significant diversity of ‘Other Birds’ (Table 1). Three species of mammals were analysed individually: European hedgehogs, grey squirrels, and red foxes. As nocturnal and crepuscular mammals are likely to respond differently to daytime visits by humans and dogs (Gaynor et al. 2018), we analysed “Non-diurnal Mammals” as a group which comprised of European hedgehogs, red foxes, wood mice (Apodemus sylvaticus), European rabbits (Oryctolagus cuniculus) and muntjac (Muntiacus reevesi) (Online Resource-Fig. 1). Grey squirrels were the only diurnal mammal species analysed, therefore a “Diurnal Mammal” group was not included in the analyses. Camera trap records of brown rats showed them to be predominantly diurnal, but their numbers were so few that they were excluded from the analysis.
Statistics
Establishing sites of low and high human and dog visits
Total contacts of people and dogs were used as an indicator of anthropogenic disturbance at a particular site. Human and dog contacts, separately recorded at each site, were summed and divided by the time of operation of the camera at each site to generate Relative Abundance Indices (RAIs) in contacts per day (Carbone et al. 2001). We justify the use of RAIs as we chose sites of the same habitat with similar fields of view, and examined activity for a period of 2.5 months, a relatively constrained window of observation. This increases the likelihood that our ability to detect particular species was similar across sites. We justify the use of these “Human + Dog RAIs” on the basis that dogs were not seen without humans, and the effects of both on wildlife could not be easily separated. There was a statistically significant positive association between human RAIs and dog RAIs (simple linear regression with logged RAIs: b = 0.67, p-value < 0.001, R2 = 0.70; Online Resource-Table 4 and Fig. 2).

Median Relative Abundance Indices of Humans + Dogs in Low and High Visit Confidence Intervals
Human + Dog RAIs per site ranged from zero for cameras in remote, fenced areas to 127 contacts day−1 for cameras near busy paths. The distribution of Human + Dog RAIs was strongly skewed, with many sites having very low values and few sites having very high values, with no clear gap between the two (Online Resource-Fig. 3). Therefore, sites were divided into “low” and “high” visit sites using the median Human + Dog RAIs (2.18 contacts day−1). Sites with fewer than the median humans and dogs contacts per day were defined as low visit sites (n = 13, median = 1.47 contacts day−1 (95% CI[0.18, 2.10])), while sites with greater than the median human and dogs contacts per day were defined as high visit sites (n = 13, median = 11.61 contacts day−1 (95% CI[3.14, 41.29])) (Fig. 2) (for mean values, see Online Resource-Fig. 4 and Table 5).

Temporal activity schedules of a Human + Dogs, b All Birds, c Common Blackbirds, d Eurasian Magpies, e Song Thrushes, f Wood Pigeons, g Non-diurnal Mammals, h European Hedgehogs, i Grey Squirrels and j Red Foxes in low visit sites (purple line) and high visit sites (yellow line) with 95% confidence intervals (dashed lines). For number of contacts for each species, see Table 1
Spatial analysis
To examine the impact of human and dog abundances on wildlife species abundances, RAIs were calculated for wildlife species at each site using the same method as for humans and dogs above. Wildlife RAIs were put into low and high visit site groups and the two groups compared using Wilcoxon tests, due to the general non-homogeneity of variances and lack of normality.
Temporal analysis
To examine how the activity schedules of wildlife species are affected by human and dog visits, we compared the activity schedules of each wildlife species between our low and high visit sites. We postulated that at low visit sites, where human and dog disturbance is less, the observed activity schedules are more likely to reflect the natural times of activity for a species. Hence, differences between activity schedules in low and high visit sites suggests human and dogs have disrupted the natural activity schedules of wildlife species.
Activity schedules were estimated for wildlife species by using contacts. A minimum of 25 contacts was required in low visit sites and high visit sites for a wildlife species to be included in the analysis, as this was the minimum number of contacts needed to generate an activity schedule for the temporal analysis.
Timestamps were then converted to ‘solarTime’, using the R package ‘Activity’ (Rowcliffe et al. 2014; Rowcliffe 2022), to standardise how sunrise and sunset varied between 18th April and 10th June across the 26 sites: times at sunrise and sunset were standardised to the mean sunrise and sunset times, respectively. This corrected for changes in timings of behaviour due to the seasonal photoperiod change (Vasquez et al. 2019; Rowcliffe 2022).
Activity schedule estimates were generated using the R package ‘Activity’ (Rowcliffe et al. 2014; Rowcliffe 2022). ‘Activity’ which converts solarTime, in radians, into non-parametric kernel density estimates for each species, i.e. activity schedules (Rowcliffe et al. 2014; Rowcliffe 2022).
To estimate the accuracy of the activity schedules estimates, we generated their standard errors by recalculating the activity schedule estimates 1,000 times using bootstrapping (Rowcliffe 2022).
Activity schedule estimates of each wildlife between low and high visit sites were compared using Watson Wheeler tests using the dedicated function within ‘circular’ package (Agostinelli and Lund 2022). Using a degree of freedom of two, temporal activity schedule estimates from low and high visit sites were significantly different if the chi-squared value was greater than 5.991 (p-value < 0.05).
All plotting and analyses were conducted in R v.3.6.2 (R Core Team 2019).
Results
Across 26 sites, the total trapping effort was 389 days. In total, 2,178 wildlife contacts of 18 species (Table 1), and 3,558 contacts of humans and dogs were recorded, with 1,918 human detections and 1,640 dog detections.
Spatial analysis
There was an eightfold increase in Human + Dog RAIs in high visit sites relative to low visit sites. For all wildlife species and groups, there were no significant differences between RAIs in low visit sites and high visit sites (Table 2).
Temporal analysis
The activity schedule for human and dogs in low sites was not significantly different from that in high visit sites (Fig. 3 and Table 3). There were no significant differences in the activity schedules in low visit sites and high visit sites for all wildlife species and groups with the exception of European hedgehogs (Fig. 3 and Table 3). European hedgehogs showed more activity later in the mornings and earlier in the evenings at low sites (Fig. 3h).
Discussion
Camera traps have a history of being used to study rare, widely ranging mammals in remote areas. However it is only in more recent years have they been used in urban green spaces and to study common urban birds and mammals. This study further shows their value to the study of small-scale spatiotemporal patterns of habitat use in response to disturbance, at both the individual species and community level. Using camera traps overcomes the problems of more typical focal observations, including any bias from the presence of a human observer and the limited time period for such observations. This results in more accurate observations of wildlife activity and increases the likelihood of capturing long term changes in behaviour in response to a range of environmental factors.
The spatial analysis revealed no significant differences between the activity of particular species at low and high visit sites. Other studies suggest that human and dog presence could affect the distribution of some of the species we observed, including magpies and blackbirds, the bird species seem most frequently and widely across our study sites (Table 1). For example, in parks and recreational spaces, blackbirds have been shown to be less active in sites with higher human or dog visitation (Fernández-Juricic and Tellería 2000; van der Zande and Vos 1984), while magpies have been reported to be anthropophilic in natural habitats (Gutzwiller et al. 2002; Larson et al. 2016). Foxes too are known to be anthropophilic (Contesse et al. 2004), but were not significantly more active in high visit sites in this study, where they may have found food discarded by people, or dog faeces, on which they are known to feed (Waggershauser et al. 2022). In another study in London using camera traps across six urban green spaces, no differences were found in fox activity rates relative to levels of human activity (Lovell et al. 2022), although temporal differences were observed which were not found in this study.
With respect to temporal activity, combined human and dog activity was similar at high and low visit sites, and showed a distinct, flat peak between mid-morning and mid-afternoon (Fig. 3). Were this affecting diurnal wildlife activity at high visit sites, we would have expected species to show a depressed midday activity, with activity periods shifted to early morning and/or late evening. This was found for no diurnal species (Fig. 3). While we would expect temporal impact of human and dog activity to be less likely for nocturnal species, a significant temporal effect was found for European hedgehogs, a largely nocturnal species which also exhibits some diurnal activity. In high visit sites, European hedgehogs shifted their foraging to earlier periods in the morning and later periods in the evening, suggesting that they may be avoiding humans and dogs. Rast et al. (2019) also found that European hedgehogs exhibit temporal evasion of areas experiencing greater human disturbance. Given the steep recent decline in British European hedgehog populations, particularly in urban areas (Wilson and Wembrige 2018), this evidence of anthropogenic disturbance is noteworthy. Avoidance of humans and dogs may limit foraging activity and, potentially, fitness (Ditchkoff et al. 2006; Larson et al. 2016; Gaynor et al. 2018).
European hedgehogs aside, the results of this study suggest that the presence of humans and dogs does not affect patterns of use of urban green spaces by a range of common wildlife species. Given this evidence, our study suggests that the phenomenon of adaptation by wildlife to recreational activity may be significant and worthy of more study.
This study has a number of limitations. Many factors determine the activity of wildlife species in woodlands, including the local distribution of vegetation, food and water resources, and the location of burrows and nest sites. We measured only one factor, the effects of visits by humans and dogs, and variation in other factors may have obscured anthropogenic effects, particularly if these habitats were not of particular significance to the species observed. We sought to control for other factors by selecting traps sites with specific, and very similar, vegetation and microhabitats. Furthermore, the species we observed do have a strong natural association with broadleaved woodland floors as a foraging site. This is an important microhabitat for common blackbirds and song thrush (Carpenter et al. 2009), and one which is frequently used by wood pigeon (Robinson 2005) and Eurasian magpie (Birkhead 2010). Broadleaved woodland floors are also used for foraging by grey squirrels (Gurnell 1987; Kenward et al. 1998), red foxes (Sidorovich et al. 2006) and European hedgehogs (Riber 2006).
Another possible limitation of this study is the manner in which we established high and low visit sites. The majority of the 26 woodland sites had infrequent visits by humans and dogs, and a very few had extremely high visit levels. Separating high and low visit sites based on median contacts was sensible in the absence of any more ecological justification, and created an equal number of high and low visit sites. However, because of the skewed distribution of contacts between sites, the resultant “high” category displayed a greater range of values (2–127 contacts per day) than the low (0–2 contacts/day). Hence, any anthropogenic effects that occur only at very high levels of humans and dog visits may have been obscured by the inclusion in the “high visit” category of sites with lower visit levels.
Furthermore, the general applicability of our findings is limited by two factors: firstly, Hampstead Heath is not a typical urban green space as it is exhibits both high ecological diversity and intense recreational use; and secondly wildlife behaviour was recorded for only two and half months, which prevented analysis on how the year-round behaviour may be being impacted by recreation.
Finally, time spent foraging in specific habitats is not necessarily a good indicator of the fitness and sustainability of wildlife populations in the face of human and dog activity. Predation and disturbance of nests, for instance by dogs off-lead, may be much more significant. The encouragement in urban spaces of anthropophilic species, like Eurasian magpies and red foxes, may also have a negative impact on other wild species with which they compete, or on which they feed. European hedgehogs on Hampstead Heath, for instance, are often found injured or dead due to injuries that may have been caused by dogs or red foxes.
Future studies would benefit from a more experimental approach, matching, or even creating sites with high and low levels of human and dog visits prospectively, and inclusion of a more thorough assessment of impacts of humans and dogs on wildlife fitness.
In conclusion, our study provides evidence that the spatial and temporal activity of common bird and mammal species in one urban woodland is not significantly reduced by increased frequency visits by humans and dogs, with the exception that European hedgehogs were more nocturnal in sites with high levels of human and dog visits. While this does not dispel the possibility that such disturbance is harmful to wildlife communities, it suggests that a broader consideration of possible anthropogenic effects, beyond those on foraging activity, would be desirable, as well as a greater focus on possible behavioural adaptation by wildlife to human disturbance.
Availability of data and material
The original images and resulting dataset are not publicly available to protect the privacy of those recorded.
Code availability
All code is currently available by request from the corresponding author.
References
Agostinelli C, Lund U (2022) R package 'circular': Circular Statistics (version 0.4–95). https://r-forge.r-project.org/projects/circular/
Banks PB, Bryant JV (2007) Four-legged friend or foe? Dog walking displaces native birds from natural areas. Biol Letters 3(6):611–613. https://doi.org/10.1098/rsbl.2007.0374
Birkhead T (2010) The Magpies: The Ecology and Behaviour of Black-billed and Yellow-billed Magpies Poyser Monographs London -10: 1408140241. ISBN-13: 978-1408140246
Bötsch Y, Tablado Z, Scherl D, Kery M, Graf RF, Jenni L (2018) Effect of Recreational Trails on Forest Birds: Human Presence Matters. Front Ecol Evo 6:175. https://doi.org/10.3389/fevo.2018.00175
Carbone C, Christie S, Conforti K, Coulson T, Franklin N, Ginsberg JR, Shahruddin WN (2001) The use of photographic rates to estimate densities of tigers and other cryptic mammals. Anim Conserv 4(1):75–79. https://doi.org/10.1017/S1367943001001081
Carbone C, Cates R (2018) LONDON HOGWATCH Hampstead Heath camera-trap survey April- July 2018. Zoological Society of London. https://democracy.cityoflondon.gov.uk/documents/s122517/Appendix%202%20-%20Hampstead%20Heath%20Survey%202018%20Report.pdf. Accessed 27 Feb 2020
Carpenter R, Charman E, Smart J, Amar A, Gruar D, Grice P (2009) Habitat associations of woodland birds II. RSPB Research Report No. 36. ISBN 978–1–905601–20–2
City of London (2009) Phase 1 Habitat Survey data of Hampstead Heath, London 2008–2009 [data file].
City of London (2010) Hampstead Heath management plan Part II. Topic Paper 2: The Natural Landscape. City of London
City of London (2019) Hampstead Heath Management Strategy 2018 to 2028. City of London. https://www.cityoflondon.gov.uk/assets/Green-Spaces/hampstead-heath-strategy.pdf. Accessed 14 Jul 2020
Contesse P, Hegglin D, Gloor S, Bontadina F, Deplazes P (2004) The diet of urban foxes (Vulpes vulpes) and the availability of anthropogenic food in the city of Zurich. Switzerland Mamm Biol 69(2):81–95. https://doi.org/10.1078/1616-5047-00123
Ditchkoff SS, Saalfeld ST, Gibson CJ (2006) Animal behavior in urban ecosystems: modifications due to human-induced stress. Urban Ecosyst 9:5–12. https://doi.org/10.1007/s11252-006-3262-3
Dowding CV, Harris S, Poulton S, Baker PJ (2010) Nocturnal ranging behaviour of urban hedgehogs, Erinaceus europaeus, in relation to risk and reward. Anim l Behav 80(1):13–21. https://doi.org/10.1016/j.anbehav.2010.04.007
Fernández-Juricic E, Tellería JL (2000) Effects of human disturbance on spatial and temporal feeding patterns of Blackbird Turdus merula in urban parks in Madrid. Spain Bird Study 47(1):13–21. https://doi.org/10.1080/00063650009461156
Fernández-Juricic E, Jimenez MD, Lucas E (2001) Alert distance as an alternative measure of bird tolerance to human disturbance: Implications for park design. Environ Conserv 28:263–269. https://doi.org/10.1017/S0376892901000273
Gaynor KM, Hojnowski CE, Brashares CNH, JS, (2018) The influence of human disturbance on wildlife nocturnality. Science 360(6394):1232–1235. https://doi.org/10.1126/science.aar7121
Gurnell J (1987) The natural history of squirrels. Christopher Helm, London
Gutzwiller KJ, Riffell SK, Anderson SH (2002) Repeated human intrusion and the potential for nest predation by gray jays. J Wildlife Manage 66:372–380. https://doi.org/10.2307/3803170
Kenward R, Hodder K, Rose R, Walls C, Parish T, Holm J, Morris P, Walls S, Doyle F (1998) Comparative demography of red squirrels (Sciurus vulgaris) and grey squirrels (Sciurus carolinensis) in deciduous and conifer woodland. J Zool 244:7–21. https://doi.org/10.1111/j.1469-7998.1998.tb00002.x
Kowalski M (2013) ExifPro (v 2.1). https://www.exifpro.com/. Accessed 20 Apr 2020
Larson CL, Reed SE, Merenlender AM, Crooks KR (2016) Effects of recreation on animals revealed as widespread through a global systematic review. PLoS ONE 11(12):e0167259. https://doi.org/10.1371/journal.pone.0167259
Lovell C, Li S, Turner J, Carbone C (2022) The effect of habitat and human disturbance on the spatiotemporal activity of two urban carnivores: The results of an intensive camera trap study. Ecol Evol 12:e8746. https://doi.org/10.1002/ece3.8746
Matsyura AV, Jankowski K, Zimaroyeva AA (2015) Escape behaviour of Corvidae in an urban ecosystem of Zhytomyr (Ukraine). Rom J Biol Zool 60:125–134
Monterroso P, Alves PC, Ferreras P (2013) Catch me if you can: diel activity patterns of mammalian prey and predators. Ethology 119(12):1044–1056. https://doi.org/10.1111/eth.12156
Natural England (1990) Hampstead Heath Woods SSSI. https://designatedsites.naturalengland.org.uk/sitedetail.aspx?SiteCode=S1003451&SiteName=&countyCode=&responsiblePerson=&unitId=&SeaArea=&IFCAArea=. Accessed 31 Jul 2020
R Core Team (2019) R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. https://www.R-project.org/
Rast W, Barthel L, Berger A (2019) Music festival makes hedgehogs move: How individuals cope behaviorally in response to human-induced stressors. Anim Open Sccess J MDPI 9(7):455. https://doi.org/10.3390/ani9070455
Reilly ML, Tobler MW, Sonderegger DL, Beier P (2017) Spatial and temporal response of wildlife to recreational activities in the San Francisco Bay ecoregion. Biol Conserv 207:117–126. https://doi.org/10.1016/j.biocon.2016.11.003
Riber AB (2006) Habitat use and behaviour of European hedgehog Erinaceus europaeus in a Danish rural area. Acta Theriol 51:363–371. https://doi.org/10.1007/BF03195183
Robinson RA (2005) BirdFacts: profiles of birds occurring in Britain and Ireland (BTO Research Report 407). BTO, Thetford. Available from: https://www.bto.org/birdfacts
Rowcliffe JM (2022) Overview of the activity package. https://cran.r-project.org/web/packages/activity/activity.pdf. Accessed 1 Jul 2022
Rowcliffe JM, Carbone C (2008) Surveys using camera traps: are we looking to a brighter future? Anim Conserv 11:185–186. https://doi.org/10.1111/j.1469-1795.2008.00180.x
Rowcliffe JM, Kays R, Kranstauber B, Carbone C, Jansen PA (2014) Quantifying levels of animal activity using camera trap data. Methods Ecol Evol 5(11):1170–1179. https://doi.org/10.1111/2041-210x.12278
Sidorovich VE, Sidorovich AA, Izotova IV (2006) Variations in the diet and population density of the red fox Vulpes vulpes in the mixed woodlands of northern Belarus. Mamm Biol 71(2):74–89. https://doi.org/10.1016/j.mambio.2005.12.001
Sollmann R, Mohamed A, Samejima H, Wilting A (2013) Risky business or simple solution – Relative abundance indices from camera-trapping. Biol Cons 159:40–412. https://doi.org/10.1016/j.biocon.2012.12.025
Soulsbury CD, White PC (2015) Human-wildlife interactions in urban areas: a review of conflicts, benefits and opportunities. Wildlife Res 42(7):541–553. https://doi.org/10.1071/WR14229
Steenweg R, Whittington J, Hebblewhite M, Forshner A, Johnston B, Petersen D, Shepherd B, Lukacs PM (2016) Camera-based occupancy monitoring at large scales: Power to detect trends in grizzly bears across the Canadian Rockies. Biol l Conserv 201:192–200. https://doi.org/10.1016/j.biocon.2016.06.020
Storch I, Leidenberger C (2003) Tourism, mountain huts and distribution of corvids in the Bavarian Alps, Germany. Wildlife Biol 9:301–308. https://doi.org/10.2981/wlb.2003.018
Taylor K, Anderson P, Taylor R, Longden K, Fisher P (2005) Dogs, Access and Nature Conservation. English Nature Research Reports, Number 649, 157. https://publications.naturalengland.org.uk/publication/65013. Accessed 19 Jan 2021
Triguero-Mas M, Dadvand P, Cirach M, Martínez D, Medina A, Mompart A, Basagaña X, Gražulevičienė G, Nieuwenhuijsen MJ (2015) Natural outdoor environments and mental and physical health: Relationships and mechanisms. Environ Int 77(1):35–41. https://doi.org/10.1016/j.envint.2015.01.012
van der Zande AN, Vos P (1984) Impact of a semi-experimental increase in recreation intensity on the densities of birds in groves and hedges on a lake shore in The Netherlands. Biol Cons 30:237–259. https://doi.org/10.1016/0006-3207(84)90086-7
Vazquez C, Rowcliffe JM, Spoelstra K, Jansen PA (2019) Comparing diel activity patterns of wildlife across latitudes and seasons: time transformation using day length. Methods Ecol Evol 10:2057–2066. https://doi.org/10.1111/2041-210X.13290
Waggershauser CN, Taberlet P, Coissac E, Kortland K, Hambly C, Lambin X (2022) Interspecific coprophagia by wild red foxes: DNA metabarcoding reveals a potentially widespread form of commensalism among animals. Ecol Evol 12:e9029. https://doi.org/10.1002/ece3.9029
Welbourne DJ, Claridge AW, Paull DJ, Lambert A (2016) How do passive infrared triggered camera traps operate and why does it matter? Breaking down common misconceptions. Remote Sens Ecol Conserv 2(2):77–83. https://doi.org/10.1002/rse2.20
Wilson E, Wembrige D (2018) The State of British Hedgehogs 2018. People’s Trust for Endangered Species: London, UK [online] Available at: https://www.hedgehogstreet.org/wp-content/uploads/2018/02/SoBH-2018_final.pdf. [Accessed 11 Aug 2022]
Acknowledgements
Staff from the Zoological Society of London, City of London and volunteers from Heath Hands set up and managed the camera traps used in the data collection.
Funding
The data collection using camera traps was majorly funded by the City of London, British Hedgehog Preservation Society, the People's Trust for Endangered Species, and the Pauline Meredith Charitable Trust. CC and MR are funded by Research England.
Author information
Authors and Affiliations
Contributions
Jeff Waage, Chris Carbone, Rachael Beasley, and Marcus Rowcliffe contributed to the study conception and design. Data collection was performed by Chris Carbone and Adrian Brooker. Analysis was performed by Jeff Waage and Rachael Beasley, and advised by Marcus Rowcliffe. The map of Hampstead Heath was made by Adrian Brooker. The first draft of the manuscript was written by Rachael Beasley and Jeff Waage. All authors commented and edited previous versions of the manuscript. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval/Consent to participate/Consent for publication
Signs were placed around Hampstead Heath informing visitors that camera traps were in use. No effort was or will be made to identify visitors recorded by the camera traps.
Conflicts of interest
The authors have no conflicts of interest to declare.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Beasley, R., Carbone, C., Brooker, A. et al. Investigating the impacts of humans and dogs on the spatial and temporal activity of wildlife in urban woodlands. Urban Ecosyst 26, 1843–1852 (2023). https://doi.org/10.1007/s11252-023-01414-z
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11252-023-01414-z