
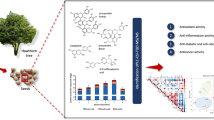
Abstract
Jack bean (JB), Canavalia ensiformis (L.) DC, is a commonly cultivated legume in Indonesia. It is rich in protein, which can be hydrolyzed, making it potentially a good source of bioactive peptides. Intestinal inflammation is associated with several diseases, and the production of interleukin-8 (IL-8) in intestinal epithelial cells induced by tumor necrosis factor (TNF)-α has an important role in inflammatory reaction. The present study investigated the anti-inflammatory effects of peptides generated from enzymatic hydrolysis of JB protein on human intestinal Caco-2BBe cells. Additionally, in silico approaches were used to identify potential bioactive peptides. JB protein hydrolysate (JBPH) prepared using pepsin and pancreatin reduced the IL-8 expression at protein and mRNA levels in Caco-2BBe cells stimulated with TNF-α. Immunoblot analysis showed that the JBPH reduced the TNF-α-induced phosphorylation of c-Jun-NH(2)-terminal kinase, nuclear factor kappa B (NF-κB), and p38 proteins. Anti-inflammatory activity was observed in the 30% acetonitrile fraction of JBPH separated on a Sep-Pak C18 column. An ultrafiltration method revealed that relatively small peptides (< 3 kDa) had a potent inhibitory effect on the IL-8 production. Purification of the peptides by reversed-phase and anion-exchange high performance chromatography produced three peptide fractions with anti-inflammatory activities. A combination of mass spectrometry analysis and in silico approaches identified the potential anti-inflammatory peptides. Peptides derived from JB protein reduces the TNF-α-induced inflammatory response in Caco-2BBe cells via NF-κB and mitogen-activated protein kinase signaling pathways. Our results may lead to a novel therapeutic approach to promote intestinal health.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Jack bean (JB, Canavalia ensiformis (L.) DC) is a protein-rich legume commonly cultivated in Indonesia. It is highly productive at harvest and can be grown on marginal land with low nutrients. JB can be a good source of essential amino acids [1] and a promising protein alternative to soybeans, which are widely utilized worldwide. Despite its nutritional advantage, the presence of anti-nutritional substances such as tannin, hydrogen cyanide, and phytate limits the consumption of JB [2, 3]. Jack bean protein is mainly composed by Canavalin as major storage protein of the jack bean and belongs to the classical vicilin fraction. Additionally, Concanavalin A and B is renowned as lectin presented in jack bean [4]. A growing body of evidence suggested jack bean protein showed beneficial physiological function, including anti-oxidative and anti-cholesterol [4, 5]. Another study suggested that fermented jack bean (jack bean tempeh) exhibited angiostensin-converting enzyme (ACE) inhibitory effect [6]. JB protein is rich in hydrophobic amino acids such as leucine, valine, proline, and alanine [7]. Previous studies show that most bioactive peptides that exert beneficial physiological effects on human health are rich in the hydrophobic amino acids. These peptides penetrate into the intestinal mucosa and interact with target molecules to show biological effect [8]. Indeed, it has been reported that protein hydrolysates derived from JB protein exhibit various biological activities such as radical scavenging activity, ferric reducing power, and angiotensin-converting enzyme inhibitory activity [9, 10]. A variety of proteins in jack bean provide a material basis for diverse peptides after digestion. Thus, the JB protein can be a promising source of bioactive peptides. Accumulating evidence demonstrates that intestinal inflammation is involved in both intestinal and non-intestinal diseases [11]. Although intestinal inflammation is induced by a wide range of stimuli, tumor necrosis factor (TNF)-α expressed by activated immune cells such as macrophages and dendritic cells upregulates the production of other pro-inflammatory cytokines in various cells [12]. Interleukin-8 (IL-8) is produced in intestinal epithelial cells in response to TNF-α and plays a critical role in recruiting neutrophils during inflammatory conditions [13]. The excessive neutrophil activation contributes to tissue damage and the pathogenesis of inflammatory diseases. Accordingly, the suppression of IL-8 production by dietary peptides may be an effective approach to promote intestinal health [14, 15].
The present study aimed to produce anti-inflammatory peptides from JB protein by enzymatic digestion using pepsin and pancreatin. To investigate the anti-inflammatory effect of peptides from jack bean protein hydrolysate (JBPH) and their underlying mechanisms, a well-known human intestinal cell model, Caco-2Bbe, stimulated with TNF-α was used in the study. In addition, for the identification of potential peptides responsible for the anti-inflammatory activity, an in silico approach was used.
Materials and methods
The material and methods section is presented as supplementary material.
Results and Discussion
Effect of JBPH on Viability of Caco-2BBe Cells
Because dietary peptides should be non-toxic to the intestinal cells, the effect of JBPH on the viability of Caco-2BBe cells was investigated. The treatment of cells with JBPH at concentrations up to 2500 µg/mL for 24 h did not influence the viability of Caco-2BBe cells (Fig. 1).

Effect of JBPH on viability of Caco-2BBe cells. Cells were incubated with JBPH for 24 h and the cell viability was evaluated using a commercially available kit. Values are presented as the mean ± standard error of mean (n = 5)
Anti-inflammatory Effect of JBPH
The production of IL-8 in intestinal epithelial cells induces the neutrophil recruitment to inflammatory site and is closely involved in the pathogenesis of various intestinal diseases [16]. Figure 2a shows that TNF-α treatment substantially increased IL-8 production in intestinal Caco-2BBe cells compared to that in control cells, indicating an inflammatory response. Pre-treatment of cells with JBPH reduced the TNF-α-induced IL-8 production in a more or less dose-dependent manner. The results showed that JBPH at 1000, 1500 and 2500 µg/mL potently suppressed the IL-8 production, indicated by the observation that the IL-8 levels with these treatments were not significantly different from the control cells. Therefore, the cells incubated with 1000 and 1500 µg/mL JBPH were subjected to qRT-PCR analysis of the IL-8 mRNA expression. Consistent with the protein levels of IL-8, JBPH reduced the IL-8 mRNA expression in a dose-dependent manner in the TNF-α-treated Caco-2BBe cells (Fig. 2b).

Effect of JBPH on IL-8 protein and mRNA expression in Caco-2BBe cells. Caco-2BBe cells were incubated with and without JBPH in the presence and absence of TNF-⍺. The IL-8 protein production was determined by ELISA (a). The IL-8 mRNA expression was determined by qRT-PCR analysis (b). Values are presented as the mean ± SEM (n = 5). Means not marked by a common letter are significantly different as determined using the Tukey–Kramer’s post-hoc test (P < 0.05)
Cellular Mechanism of Anti-inflammatory Effect of JBPH
It has been reported that TNF-α transcriptionally upregulates the IL-8 expression through various signaling pathways such as NF-κB and MAPK [17]. Immunoblot analysis (Fig. 3a-c) showed that the stimulation of Caco-2BBe cells with TNF-α upregulated the phosphorylation of p65, p38, and JNK, indicating cellular activations. Pre-treatment with JBPH suppressed the phosphorylation of p65, p38 and JNK proteins caused by TNF-α stimulation in a dose-dependent manner. These results suggest that JBPH suppresses inflammatory signaling induced by TNF-α to decrease the IL-8 transcription. The regulation of NF-κB, p38,and JNK pathways seems to be a central event in the JBPH-mediated suppression of IL-8 production.

Effect of JBPH on phosphorylation of NF-κB and MAPK signaling molecules. Caco-2BBe cells were incubated with and without JBPH in the presence and absence of TNF-α. Phosphorylation of p65 (a), p38 (b), and JNK (c) was determined by immunoblot analysis. Values are presented as the mean ± SEM (n = 5). Means not marked by a common letter are significantly different as determined using the Tukey–Kramer’s post-hoc test (P < 0.05)
TNF-α is known to activate these cellular signaling pathways through TNF receptors, resulting in the IL-8 expression in intestinal epithelial cells. Upon NF-κB activation and translocation into the nucleus, it binds to the promoter region of the IL-8 gene, thereby initiating the transcription of IL-8 mRNA. JNK and p38 also activate the downstream transcription factors such as AP-1, c-Jun, ATF-2, and CREB, which induce the transcription of IL-8 [18, 19]. Previous studies demonstrated that peptides derived from egg-white and a dipeptide derived from ovotransferrin effectively suppressed the activation of NF-κB and MAPK pathways [20, 21]. Accordingly, our results suggest that the reduced activation (phosphorylation) of NF-κB, p38, and JNK by JBPH results in the downregulation of IL-8 transcription in the cells.
Anti-inflammatory Effect of Peptides Fractionated from JBPH
Fractionation of JBPH was performed in a sequential step to isolate potent anti-inflammatory peptides. First, JBPH was subjected to a Sep-Pak C18 cartridge using a three-step acetonitrile gradient elution (15, 30, and 45%). Based on the BCA assay, the recovery ratio of the peptide in the 15, 30, and 45% acetonitrile fractions were 17.9, 52.7, and 15.3%, respectively. However, when JBPH was eluted at higher acetonitrile concentration (65%), the recovery ratio of peptide was very low (1.44%). Therefore, the three fractions from the lower acetonitrile concentrations were administered to Caco-2BBe cells at concentration of 1000 µg/mL 6 h before TNF-α stimulation.
Among the three fractions, suppression of IL-8 production by the 30% acetonitrile fraction was roughly comparable to that of the original JBPH (Fig. 4a). Next, the 30% acetonitrile fraction was further separated using ultrafiltration with 3 kDa MWCO. Based on the BCA assay, 14 and 86% peptides from the 30% acetonitrile fraction were distributed in the filtrate (< 3 kDa) and retentate (> 3 kDa) fractions, respectively.
Interestingly, both fractions suppressed the IL-8 production in the TNF-α-stimulated Caco-2 BBe cells in a similar manner (Fig. 4b); peptides in the filtrate fraction with a relatively low protein concentration (142 µg/mL) exhibited IL-8 suppression similar to those by the retentate fraction (858 µg/mL) and the original sample. This suggested that the peptides in the filtrate fraction also had potent anti-inflammatory activity.

Effect of JBPH on IL-8 protein levels in Caco-2BBe cells. Caco-2BBe cells were incubated with and without different JBPH in the presence and absence of TNF-α. The IL-8 protein production was determined by ELISA. The three fractions of JBPH were prepared by a step-gradient separation on Sep-Pak C18 cartridges (a); and JBPH were separated by ultrafiltration with 3 kDa molecular weight cut-off (b). Values are presented as the mean ± SEM (n = 5). Means not marked by a common letter are significantly different as determined using the Tukey–Kramer’s post-hoc test (P < 0.05)
Accordingly, the filtrate fraction was further separated using RP-HPLC. As shown in Fig. 5a, four fractions (F1–F4) were collected based on the retention time, of which anti-inflammatory activity was tested in fractions F2 and F3 in Caco-2BBe cells since the peptides were mostly distributed in these two fractions according to the BCA assay. Both the fractions suppressed the TNF-α-induced IL-8 production (Fig. 5b).

Separation of JBPH with RP-HPLC and its effect on IL-8 protein in Caco-2BBe cells. The chromatogram of JBPH by RP-HPLC was shown. The dashed line indicated the acetonitrile gradient for elution (a). Caco-2BBe cells were incubated with and without jack bean protein hydrolysates (JBPH) RP-HPLC fractions in the presence and absence of TNF-α. The IL-8 protein production was determined by ELISA (b). Values are presented as the mean ± SEM (n = 5). *P < 0.05 compared to the control (+ TNF-⍺) as determined using Dunnet’s post-hoc test
The result also revealed that the suppression by F3 was comparable to the original filtrate fraction. Furthermore, the peptides in F3 may be more hydrophobic than those in F2, as they were eluted later in the RP-HPLC. Considering that hydrophobicity is an important feature in the biological activity of peptides [22, 23], F3 was subjected to separation using anion exchange chromatography. As shown in Fig. 6a, three peaks designated as P1, P2, and P3 were collected, and the anti-inflammatory activities and cytotoxicity were examined in Caco-2BBe cells. All three peaks (P1, P2, and P3) suppressed the IL-8 production induced by TNF-α without any cytotoxicity (Fig. 6b and Fig. S1). Our results also suggest that different hydrophobic peptides with relatively small molecular weights (< 3 kDa) possibly have a role in the regulation of inflammatory response in Caco-2BBe cells. We found that the anti-inflammatory peptides isolated from JBPH are of low molecular weight (< 3 kDa) with hydrophobic nature.

Separation of JBPH with anion exchange HPLC and its effect on IL-8 protein in Caco-2BBe cells. The chromatogram of JBPH by anion exchange HPLC was shown. The dashed line indicated the NaCl gradient for elution (a). Caco-2Bbe cells were incubated with and without jack bean protein hydrolysates (JBPH) anion exchange HPLC fractions in the presence and absence of TNF-α. The IL-8 protein production was determined by ELISA (b). Values are presented as the mean ± SEM (n = 5). *P < 0.05 compared to the negative control (without TNF-⍺) as determined using Dunnet’s post-hoc test
Screening of bioactive peptides in previous studies suggested that peptide with molecular weight below 3 kDa exhibited greater bioactivities such as anti-inflammatory, anti-oxidative, and anti-microbial activities compared to larger peptides [24,25,26]. This is not unexpected, since small peptides may be easily taken up by cells and thus exhibit bioactivity. The C18 reverse phase column enables the separation of peptides based on the hydrophobic interaction of peptides between the elution buffer and the stationary phase [27]. Peptides isolated from JBPH are likely to have high hydrophobicity because they eluted at acetonitrile concentrations between 68 and 82% and were collected as the F3 fraction. Similar to our study, it has been reported that plant-derived bioactive peptides with high hydrophobic properties effectively reduce the inflammatory response by inducing membrane depolarization and disrupting the inflammatory cascade pathway due to their high affinity to cell membranes in jejunal epithelial cell line [8].
Identification of Potential Anti-inflammatory Peptides
To identify the amino acid sequences of peptides, the fractions P1, P2, and P3 were subjected to the de novo peptide sequencing with mass spectrometry, which identified 267, 72, and 64 peptides in fractions P1, P2, and P3, respectively. Using a cutoff average local confidence (ALC) of ≥ 85% on a de novo peptide sequencing that represented the spectrum quality, we obtained 77, 31, and 19 peptides from P1, P2, and P3, respectively (Table S2, S3, and S4). In National Center for Biotechnology Information (NCBI) database, only five proteins, Canavalin, Concanavalin-A, Concanavalin-B, Alpha-mannosidase, and Urease, were found as parent proteins of these peptides. Using the in silico approach with PeptideRanker score, BIOPEP peptide database, and PreTP-EL, 23 peptides were identified as the anti-inflammatory peptides (Table 1). Specifically, the BIOPEP, PreTP-EL, and PeptideRanker identified 6, 8, and 9 peptides with amino acid lengths ranging from 4 to 13. BIOPEP is a database of peptides that show biological activity and has become a standard tool for screening and identifying the potential bioactive peptides [28]. BIOPEP can also predict the potential bioactivity of the parent peptides and their cleavage. In this study, BIOPEP has identified three peptides in JBPH, DDYFPYADGRNA, YWGQREDGL, and ANPRHLDAGGKA, which possibly release the anti-inflammatory peptides, PY, YW, and ANP, respectively. We also determined that the parent proteins of DDYFPYADGRNA and YWGQREDGL are α-mannosidase and Concanavalin B, respectively. PreTP-EL is a novel therapeutic peptide prediction method using ensemble learning that combines individual predictors through a genetic algorithm. It provides information on whether the peptide sequences entered into the query are anti-inflammatory peptides or not [29]. In this study, PreTP-EL has identified eight peptides as potential anti-inflammatory peptides, including LLNPDNNQNLRLL with the highest score (Table S5). Although Canavalin has been found to be the parent protein of the FLSSTKRLPSYL, other seven peptides are still orphan. PeptideRanker is a bioactive peptide prediction server based on a novel N-to-1 neural network and used in many bioactive peptide studies [30, 31]. It predicted three peptides with high scores from the P1, P2, and P3 fractions, indicating that they may have anti-inflammatory properties. The peptides include WTGF, FGLM, RLFG, FVDF, FDYL, HFDY, DLDF, FEEF and YEGLDF.
However, the anti-inflammatory activity of JBPH was not examined in in vivo using animal models in the present study. Murine models of experimental colitis induced by dextran sulfate sodium and 2,4,6-trinitrobenzene sulfonic acid have been used to study the anti-inflammatory activities of dietary factors [32]. It is important for the bioactive peptides to be resistant to digestive enzymes such as pepsin, trypsin, and chymotrypsin. It is very likely that the JBPH is resistant to the digestive enzyme and reaches the site of inflammation in intact forms in the intestine, because was prepared by hydrolysis using pepsin and pancreatin. The anti-inflammatory activity of JBPH using in vivo models will be investigated in future studies.
Conclusion
Hydrolysis of JB protein using pepsin and pancreatin generates potential anti-inflammatory peptides. The peptides reduce the IL-8 expression by suppressing the NF-κB and MAPK signaling pathways in the TNF-α-stimulated Caco-2BBe cells. In addition, the in silico approaches revealed the sequences of potential anti-inflammatory peptides. Taken together, our findings may contribute to the development of nutraceutical, especially from underutilized plant-based protein hydrolysates, to promote intestinal health. However, further studies are needed to investigate the biological functions of these peptides in in vitro and in vivo studies.
Data Availability
No datasets were generated or analysed during the current study.
Abbreviations
- AU:
-
Arbitrary unit
- ELISA:
-
Enzyme-linked immunosorbent assay
- IL-8:
-
Interleukin-8
- JBPH:
-
Jack bean protein hydrolysates
- JNK:
-
c-Jun-NH(2)-terminal kinase
- MAPK:
-
Mitogen-activated protein kinase
- MS:
-
Mass spectrometry
- NF-κB:
-
Nuclear factor kappa B
- qRT-PCR:
-
Real-time reversed transcription polymerase chain reaction
- RP-HPLC:
-
Reversed-Phase High-Performance Liquid Chromatography
- SEM:
-
Standard error of mean
- TNF-α:
-
Tumor necrosis factor-α
References
Sridhar KR, Seena S (2006) Nutritional and antinutritional significance of four unconventional legumes of the genus Canavalia - A comparative study. Food Chem 99(2):267–288
Ngatchic T, Yanou Nn, Oben J, Mbofung C, Yanou Nicolas N (2013) Protein quality and antigrowth effect of protein isolate of Mucuna (Mucuna Pruriens) and Canavalia (Canavalia ensiformis) seeds. Sch Acad J Biosci 1(5):183–191
Betancur-Ancona D, Gallegos-Tintoré S, Delgado-Herrera A et al (2008) Some physicochemical and antinutritional properties of raw flours and protein isolates from Mucuna pruriens (velvet bean) and Canavalia ensiformis (Jack bean). Int J Food Sci Technol 43(5):816–823
Marfo EK, Wallace P, Timpo G, Simpson BK (1990) Cholesterol lowering effect of Jackbean (Canavalia Ensiformis) seed protein. Gen Pharmac Vol. 21
Yusuf D, Kholifaturrohmah R, Nurcholis M, Setiarto RHB, Anggadhania L, Sulistiani (2023) Potential of White Jack Bean (Canavalia ensiformis L. DC) Kefir as a Microencapsulated antioxidant. Prev Nutr Food Sci 28(4):453–462
Puspitojati E, Indrati R, Cahyanto MN, Marsono Y (2019) Formation of ACE-inhibitory peptides during fermentation of jack bean tempe inoculated by usar Hibiscus tiliaceus leaves starter. IOP Conf Ser: Earth Environ Sci 292:012022
Kanetro B, Riyanto M, Pujimulyani D, Huda N (2021) Improvement of Functional properties of Jack Bean (Canavalia ensiformis) Flour by Germination and its relation to amino acids Profile. Curr Res Nutr 9(3):812–822
Liu W, Chen X, Li H, Zhang J, An J, Liu X (2022) Anti-inflammatory function of plant-derived bioactive peptides: a review. Foods MDPI 11
Wijatniko BD, Murdiati A (2019) Antioxidant activity of bioactive peptides derived from the hydrolysates of jack bean (Canavalia ensiformis (L.) DC.) Protein isolate. AIP Conf Proc 2099:020028. https://doi.org/10.1063/1.5098433
Puspitojati E, Cahyanto MN, Marsono Y, Indrati R (2023) Jack Bean (Canavalia ensiformis) Tempeh: ACE-Inhibitory peptide formation during absorption in the small intestine. Food Technol Biotechnol 61(1):64–72
Ahluwalia B, Moraes L, Magnusson MK, Öhman L (2018) Immunopathogenesis of inflammatory bowel disease and mechanisms of biological therapies. Scand J Gastroenterol 53:79–89
Lee SH, Kwon Jeun, Cho M, La (2018) Immunological pathogenesis of inflammatory bowel disease. Intestinal Res 16:26–42
Cotton JA, Platnich JM, Muruve DA, Jijon HB, Buret AG, Beck PL (2016) Interleukin-8 in gastrointestinal inflammation and malignancy: induction and clinical consequences. Int J Interferon Cytokine Mediat Res 8:13–34
La Manna S, Di Natale C, Florio D, Marasco D (2018) Peptides as therapeutic agents for inflammatory-related diseases. Int J Mol Sci 19(9)
Zhu W, Ren L, Zhang L, Qiao Q, Farooq MZ, Xu Q (2020) The Potential of Food Protein-Derived Bioactive Peptides against Chronic Intestinal Inflammation, Mediators of Inflammation, vol. 2020. https://doi.org/10.1155/2020/6817156
Kang L, Fang X, Song YH, He ZX, Wang ZJ, Wang SL et al (2022) Neutrophil–epithelial crosstalk during intestinal inflammation. CMGH 14:1257–1267
Ruder B, Atreya R, Becker C (2019) Tumour necrosis factor alpha in intestinal homeostasis and gut related diseases. Int J Mol Sci 20(8)
Hoesel B, Schmid JA (2013) The complexity of NF-κB signaling in inflammation and cancer. Mol Cancer 12(86). https://doi.org/10.1186/1476-4598-12-86
Sabio G, Davis RJ (2014) TNF and MAP kinase signalling pathways. Semin Immunol 26(3):237–245
Zhang M, Zhao Y, Yao Y, Xu M, Du H, Wu N et al (2019) Isolation and identification of peptides from simulated gastrointestinal digestion of preserved egg white and their anti-inflammatory activity in TNF-α-induced Caco-2 cells. J Nutr Biochem 63:44–53
Wang X, Zhao Y, Yao Y, Xu M, Du H, Zhang M et al (2017) Anti-inflammatory activity of di-peptides derived from ovotransferrin by simulated peptide-cut in TNF-α-induced Caco-2 cells. J Funct Foods 37:424–432
He R, Liu M, Zou Z, Wang M, Wang Z, Ju X et al (2022) Anti-inflammatory activity of peptides derived from millet bran in vitro and in vivo. Food Funct 13(4):1881–1889
Grancieri M, Martino HSD, de Mejia EG (2021) Protein digests and pure peptides from Chia seed prevented adipogenesis and inflammation by inhibiting pparγ and nf-κb pathways in 3t3l-1 adipocytes. Nutrients 13(1):1–23
Peng L, Kong X, Wang Z, Ai-lati A, Ji Z, Mao J (2021) Baijiu vinasse as a new source of bioactive peptides with antioxidant and anti-inflammatory activity. Food Chem 339 (2021) 128159
J Ashokbhai A, Basaiawmoit B, Das S, Sakure A, Maurya R, Bishnoi M et al (2022) Antioxidative, antimicrobial and anti-inflammatory activities and release of ultra-filtered antioxidative and antimicrobial peptides during fermentation of sheep milk: In-vitro, in-silico and molecular interaction studies. Food Biosci 47(2022):101666
Hu X, Yang X, Wu Q, Li L, Wu Y, Chen S et al (2019) Purification and identification of antioxidant peptides from Schizochytrium Limacinum hydrolysates by consecutive chromatography and electrospray ionization-mass spectrometry. Molecules 24(16)
Cavaliere C, Capriotti AL, La Barbera G, Montone CM, Piovesana S, Laganà A (2018) Liquid chromatographic strategies for separation of bioactive compounds in food matrices. Molecules 23(12)
Minkiewicz P, Iwaniak A, Darewicz M (2019) BIOPEP-UWM database of bioactive peptides: current opportunities. Int J Mol Sci 20(23)
Guo Y, Yan K, Lv H, Liu B (2021) PreTP-EL: prediction of therapeutic peptides based on ensemble learning. Brief Bioinform 22(6)
Chirinos R, Villasante-Bravo N, Aguilar-Galvez A, Figueroa-Merma A, Carpentier S, Pedreschi R et al (2022) Antioxidant, antihypertensive and antidiabetic potential of peptidic fractions obtained from tarwi (Lupinus mutabilis) protein hydrolysate and identification of promising multifunctional bioactive peptides. Int J Food Sci Technol 57(11):7402–7411
Zhang B, Xu Y, Zhao C, Zhang Y, Lv H, Ji X et al (2022) Protective effects of bioactive peptides in foxtail millet protein hydrolysates against experimental colitis in mice. Food Funct 13(5):2594–2605
Eichele DD, Kharbanda KK (2017) Dextran sodium sulfate colitis murine model: an indispensable tool for advancing our understanding of inflammatory bowel diseases pathogenesis. World J Gastroenterol 23(33):6016–6029
Acknowledgements
This work was supported in part by the Natural Science Center for Basic Research and Development (NBARD-00057). We would like to thank Editage (www.editage.jp) for their English language editing services.
Funding
Open Access funding provided by Hiroshima University.
Author information
Authors and Affiliations
Contributions
B.D.W.: Conceptualization, Data curation, Investigation, Methodology, Formal analysis, Writing-original draft, Visualization. Y.Y.: Supervision, Validation. H.M.: Supervision, Validation, Resources. T.S.: Conceptualization, Data curation, Writing – review & editing, Funding acquisition, Supervision, Validation, Resources.
Corresponding author
Ethics declarations
Competing Interests
The authors declare no competing interests.
Conflict of interest
The authors declare they have no conflicts of interest to this work.
Ethics Approval
Not applicable.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic Supplementary Material
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Wijatniko, B.D., Yamamoto, Y., Hirayama, M. et al. Identification and Molecular Mechanism of Anti-inflammatory Peptides Isolated from Jack Bean Protein Hydrolysates: in vitro Studies with Human Intestinal Caco-2BBe Cells. Plant Foods Hum Nutr (2024). https://doi.org/10.1007/s11130-024-01201-x
Accepted:
Published:
DOI: https://doi.org/10.1007/s11130-024-01201-x