
Abstract
Background and aims
Decline in tree species is a complex phenomenon involving multiple factors, among edaphic conditions are assumed to play an important role as factor of predisposition of forests to this process. In this regard, scarce information exists on the effects of the internal variability of the soil with depth on the predisposition to decline, an aspect that requires further evaluation.
Methods
Characterization of the internal variability of soil was carried out at 20 sites (10 with evidence of decline and 10 with no signs of decline) and the results analyzed to determine their role in modulating the effect of drought, which is the main cause of the observed decline in Aleppo pine stands in the Comunidad Valenciana (Spain).
Results
The soil properties found to be the most explanatory were those associated with soil quality in terms of available space for root exploration, which is vital for nutrition and, above all, water uptake. Episodes of decline are associated with stands where soils have a shallow effective depth due to a low degree of profile development or through marked textural anisotropy because of particularly clayey horizons that cause abrupt changes in permeability and aeration.
Conclusion
The internal variability of the soil, closely linked to the degree of pedogenetic development, is identified as a factor that plays an important role in predisposing the vegetation to the effects of drought.
Similar content being viewed by others


Introduction
The phenomenon of decline and death of tree species, common in natural systems (Franklin et al. 1987), is particularly complex as it involves a varied set of commonly interdependent factors (Manion 1991; Ceilsa and Donaubauer 1994). All of the factors involved interact with each other, driving multiple mechanisms that operate at differing intensities across the landscape. These mechanisms are not mutually exclusive and act in an interdependent (or compensatory) way, favoring the appearance of complex spatio-temporal patterns of decline that show a marked stochastic character (Fensham and Holman 1999; Das et al. 2008).
According to the waterfall model of Manion (1991), the decline process begins with predisposing factors, which weaken individual resilience and increase susceptibility to the damaging defects of other factors that lead to tree morbidity, considered as either initiating or contributing factors (Manion and Lachance 1992; Ciesla and Donaubauer 1994).
Drought stress (whether accompanied or not by high temperatures) is a key factor in understanding the dynamics of many terrestrial ecosystems throughout the world (Dobbertin et al. 2005; Gitlin et al. 2006; Miao et al. 2009; Anderegg et al. 2012). An unusually severe decrease in rainfall can lead to high mortality in forest stands that are particularly vulnerable to the effects of drought. Even within the ecological optimum of their geographic distribution, a severe lack of rainfall can lead to trees being affected in areas where conditions predispose them to possible decline. In the Mediterranean area, this is the most common initiating factor in the decline of forest stands (Peñuelas et al. 2001; Gea-Izquierdo et al. 2014; Sánchez-Salguero et al. 2012; Camarero et al. 2015).
At forest-stand scale, certain site characteristics interact with density-dependent processes, such as competition or facilitation, defining the degree of predisposition to processes that lead to decline (Suarez et al. 2004; Koepke et al. 2010; Hosseini et al. 2017; Brown et al. 2018). For example, high densities frequently lead to mortality as a result of exacerbated competition for water and / or high activity of biotic agents (Lloret et al. 2004; Greenwood and Weisberg 2008).
Regarding drought, the combination of stand age, density and structure along with the specific site conditions (physical environment) produce complex spatial patterns of predisposition to decline, which are very difficult to predict (Fensham and Holman 1999; Das et al. 2008). The consequence is that forest decline episodes frequently manifest diffuse and irregular spatial boundaries (fronts) as well as highly variable degrees of mortality (Dobbertin et al. 2005: Miao et al. 2009; Klos et al. 2009).
Spatial patterns of drought-induced mortality are closely related to spatial and temporal variability in the conditions of the physical environment that determine water availability. Although climate determines water availability at larger scales, the physiographic characteristics (such as slope, exposure and position within the landscape) along with soil status (stoniness, texture and depth of the soil) act as local controllers of water availability for vegetation (Brown et al. 2018; Preisler et al. 2019). The role of these factors is especially relevant in the summer months in regions with Mediterranean climate, when water lost through evapotranspiration continuously exceeds precipitation (Prieto-Recio et al. 2015).
Despite the important influence of non-climatic conditions on water availability and therefore on the spatial patterns of tree decline, few studies have specifically evaluated this influence (Galiano et al. 2010). In the scarce research addressing this issue, the focus tends not to be on physiographic factors but rather, these are dealt with as secondary or complementary aspects, with the exception being the effect of altitude, which has been analyzed in several studies (Guarin and Taylor 2005; Candel-Pérez et al. 2012).
Much less attention has been directed towards evaluating the relationship between site soil conditions and decline, despite clear indications that site-scale soil characteristics play an important role in the predisposition of vegetation to decline (La Manna and Rajchenberg 2004; Dobbertin et al. 2005; Prieto-Recio et al. 2015). By modulating water availability in periods of maximum water deficit, soil is assumed to be an especially important component in the mechanisms that regulate the effects of drought at local scale (Lloret et al. 2004; Galiano et al. 2012; Peterman et al. 2012). Empirical evidence for this notion is particularly scarce, and studies documenting drought-induced decline rarely include detailed sampling of soil conditions at sites where mortality is observed, which is essential to capture small-scale variability (at stand and tree level) and thus relate soil conditions to the individual health status of the trees or the stand as a whole (Brunner et al. 2009; Galiano et al. 2010; Brown et al. 2018).
Assuming that the availability of water to tree roots depends largely on the volume of soil that can be explored and its porosity, both attributes are nuanced by properties such as stoniness, which increases the infiltration capacity of the soil but reduces its effective volume (Hlaváčiková et al. 2016; Preisler et al. 2019), the textural class, understood as a combination of percentages of sand, silt and clay that determine the state and behavior of water and air in edaphic volumes (Gitlin et al. 2006; Colins and Bras 2007; Koepke et al. 2010; Soil Survey Division Staff 2017) and the content of organic matter, as a structure-creating agent, this being an important determinant of porosity (Six et al. 2004).
These properties are not homogeneous throughout the entire depth of the soil, so the soil profiles show different degrees of internal variability that delimit spaces with different aptitudes for the storage and transmission of water and for the exploration of root systems (Suarez et al. 2004). Consequently, the spatial variability in this markedly anisotropic behavior with respect to depth, closely linked to the degree of pedological evolution (differentiation of genetic horizons), is considered to be transcendent in the predisposition to decline. Based on this hypothesis, the main objective of this study is to evaluate the role of intrinsic soil variability in forest decline processes, focusing in particular on determining the effect that the variability of edaphic properties along the profile can have on the predisposition of forest stands to suffer episodes of decline.
Materials and methods
The study was undertaken in a set of Pinus halepensis Mill forest stands in the Comunidad Valenciana (Spain), a region which is particularly characteristic of the Mediterranean area of the Iberian Peninsula. These plant communities are a common element in the configuration of the current landscape, presenting a rich variety in terms of origin, maturity conditions and vegetative vigor (Del Río et al. 2009). Given the highly frugal and drought-resistant nature of the species, it was routinely used in reforestations undertaken in the second half of the 20th century, often on marginal land as regards physical environmental requirements (García de la Serrana et al. 2015; Gómez-Sanz 2019). In recent years, episodes of notable decline have been observed in the woodlands of this circum-Mediterranean species, which have caused its complete disappearance in the most extreme cases (Allen et al. 2010).
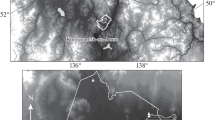
In this biogeographic context, a total number of 10 study locations were selected: two in Castellón province, six in Valencia and two in Alicante (Fig. 1). Their UTM coordinates and general site characteristics are included in Table 1. Elevation ranges from almost sea level (Almenara, Castellón) to more than 750 m in Jalance (Valencia), which obviously affects the variability of climatic conditions at each of the analyzed locations.

Spatial distribution of study locations
Based on the hypothesis that the internal variability of soil characteristics can have a notable influence on the decline process, a paired sampling was proposed in each location. The aim was to eliminate the effects of climatic variability as well as stand origin and management, thus allowing the small-scale variability of site conditions to be determined. Consequently, taking the notable presence of tree crowns with evident signs of recent death (crowns that still retain almost all of the completely withered leaves)as the selection criterion, two different situations were established: (1) stand with a satisfactory physiological state, where more than 85% of the trees had crowns that showed no clear signs of decline (site with a label ending in “N”); (2) stand geographically and physiographically close to the latter, with a similar origin, but in which evidence of recent death was found in more than 15% of the trees (site with a label ending in “D”). The spatial pattern of mortality was relatively heterogeneous in the sites with decline and their immediate surroundings. Some sites presented isolated dead trees in the same physiographic situation, while in others the dead trees appeared in relatively contiguous groups in similar physiographic situations. In the most severe cases, mortality affected all, or almost all, of the cover previously present at the site.
In the field work undertaken at each of the sites, a representative point of dominant physiographic and canopy conditions was selected, followed by a detailed ecological inventory. For this task, a circular plot of 500 m2 was established, in which dasometric and botanical information was recorded. Furthermore, a soil profile study was carried out, opening the corresponding pit, identifying and describing different soil horizons, and taking a representative sample from each of them for laboratory analyses. The presence of non-sampleable stoniness (NSS) was estimated visually in percentage by volume (FAO 2006).
Soil samples were air dried and passed through a 2 mm sieve, determining the percentage by weight of the material retained in the sieve (coarse elements, CE). The USDA textural fractions (sands, SAN; silts, SIL; clays, CLA) were obtained by the Bouyoucos method, and the organic matter (OM) content was determined by volumetry redox (Walkey-Black method). In addition, the pH in water (WPH) was measured by potenciometry in a soil:solution ratio of 1:2.5, and the presence of total carbonates (CT) was obtained using the Bernard calcimetry method. The soil profile characterizations in inventoried plots are summarized in Table 2.
In turn, genetic horizons were identified for each profile, thus allowing us to approximate the degree of evolution and assess changes in physical conditions (textural and structural) derived from the presence of markedly clayey and/or strongly cemented horizons or of relatively continuous hard rock. Given its significance in the amount of edaphic space that can be explored by root systems, the depth at which a notable change was observed in textural conditions between genetic horizons was established as a new study variable, termed “depth easily explored” (EED). To determine the latter variable, the criterion used was the change in USDA textural class in the underlying genetic horizon such that the textural conditions lead to a decrease in the conditions of permeability, aeration and water retention capacity due to an increase in the presence of fine particles (Soil Survey Division Staff 2017). Each value obtained was corroborated through confirmation of marked differences in biological activity (especially degree of root abundance) observed between the two genetic horizons in which the change in textural class is identified.
Finally, all analyzed profiles were classified, using the classification system proposed by FAO-UNESCO (IUSS Working Group 2014), for which different diagnostic elements were previously identified in accordance with criteria established by this taxonomic system.
The information obtained from the analyzed samples was statistically treated as paired observations. Hence, Normality in distribution of differences between sites with and without decline was verified (Kolmogorov-Smirnov test) and a Student’s t-test was performed for paired data. The significance level for hypothesis testing of means equality in paired differences was set at 0.05.
Results
The set of soils evaluated presents a range of internal variability at profile level that allowed the proposed objectives to be addressed. The depths of the profiles vary sufficiently, although 80% present more or less continuous hard rock at a depth of less than 1 m (leptic conditions according to WRB (IUSS Working Group 2014).
In general, there is a notable presence of non-sampleable stoniness and coarse elements, with 80% of the profiles having less than 40% fine earth (skeletic or hyperskeletic features for WRB (IUSS Working Group 2014).
As regards texture, equilibrated texture horizons dominate (loam and sandy clay loam), making up around 50% of the total. There are also horizons of coarse textures (sandy loam and loamy sand), accounting for approximately 30%, and of slightly imbalanced fine textures (clay loam and silt loam), and even fine textures (clay). The profiles are mostly homogeneous in terms of texture, although some present a marked heterogeneity, which leads to sudden changes in permeability and aeration conditions within the profile (abruptic qualifier of WRB (IUSS Working Group 2014).
From a biochemical perspective, organic matter content is moderate-low, and dominant soil reaction is either strongly or extremely basic, especially in sites that do not show decline. Furthermore, the majority of sites where decline is observed present a scarcity or absence of carbonates (non-calcareous material). No significant differences were observed among the mean values of the textural and biochemical variables studied for each profile (Fig. 2) according to the decline factor (all paired t test gave a p-value greater than 0.05).

Mean values per profile of the textural and biochemical variables in the profiles with and without decline
The same is not the case for the variables related to the volume of available soil, that is, for the total depth of the soil (TD) and for the easily explored depth (EED) by the roots within the profile (Fig. 3). The former shows no significant differences between the profiles at sites with decline and those without (p-value of 0.385 for a paired t statistic of 0.91; n = 10), while the latter does show statistically significant paired differences. The mean difference between EED pairs is 22.5 cm (standard deviation 25.74 cm), with a p-value for the paired t test of 0.022 (t statistic = 2.76; n = 10). Therefore, this variable is the only one of those analyzed that explains the differences between sites with and without decline.

Variation of the total depth of the profile and the easily explored depth for the profiles at sites with decline and without decline
Regarding the degree of development, there is a wide variation among the profiles, from scarcely-developed young soils, without intermediate horizons and which usually lay directly over consolidated bedrock (profile type “A - R”), to soils with intermediate horizons of alteration (profile type “A - Bw - C - R”) or with highly evolved profiles, in which there is a genetic horizon of accumulation, with a notable presence of clay (profile type “A - Bt - C - R”).
Finally, Fig. 4 presents a classification of soils evaluated according to the FAO-UNESCO WRB (IUSS Working Group 2014) and its relation with decline. The soils in which it is possible to identify an argic diagnostic horizon are associated in all cases with sites where decline is observed.

Relationship between general soil condition according to the FAO-UNESCO WRB categories (IUSS Working Group 2014) and observed decline (final letter of the site label indicates whether it shows decline, letter D, or not, letter N)
Discussion
The results indicate that of the set of soil characteristics, the depth of soil most easily explored by roots stands out as a property that can predispose trees to suffering decline. This variable affects the role of the soil in decoupling water availability from the purely climatic component associated with the rainfall pattern, a key quality to vegetation survival (Nadal-Sala et al. 2017), especially in the Mediterranean area.
Development of root system consumes a large part of primary production (McDowell et al. 2008), and therefore requires a compromise between developing and maintaining a larger root system and the additional water it can provide (Jackson et al. 2000). Consequently, those sites with deep well developed soils will host individuals with a more extensive, hydraulically balanced root system, which implies an advantage in water-limited environments over other individuals growing on shallow soils and therefore with less developed root systems (Nadal-Sala et al. 2017).
In shallow or poorly developed soils (Leptosols in WRB (IUSS Working Group 2014), the volume explored by roots systems is small, and this is frequently accompanied by low water retention capacity (these soils tend to be particularly stony, showing skeletic, or even hyperskeletic, conditions). On these edaphic conditions, trees cannot develop an adequate root system, which makes them especially vulnerable to drought as a trigger of decline (Schenk and Jackson 2005; Preisler et al. 2019). This is the case at sites where mortality is present in Jalance and Jijona-Alicante.
In more developed soils, the depth most easily explored is determined by the textural variability according to the depth of the profile, which is closely related to the degree of soil evolution. The pedogenetic development of profiles that include genetic horizons with highly differentiated textural behavior implies a transcendent heterogeneity which conditions the development and distribution of root systems.
In this regard, water in an “argic” diagnostic horizon (IUSS Working Group 2014) is less available to vegetation because it is more strongly retained by hygroscopic and capillary forces, which are of higher intensity. Hence, the presence of this kind of horizon reduces the volume of soil most easily explored by roots in search of water and nutrients, causing root development to be limited almost entirely to the horizons above it in the profile (closest to the surface). This situation is particularly evident in sites with decline, and especially in those soils exhibiting textural changes that can be classified as abrupt in their profiles (WRB abruptic qualifier (IUSS Working Group 2014). If, in addition, the surface horizon has a texture that does not promote water retention (sandy and/or low content in fine earth), as occurs in Vilamarxant, Almenara and Gilet-Sagunto, the predisposition to decline is considerably accentuated.
The opposite occurs in the case of soils with relatively isotropic profiles as regards their textural behavior with depth. These soils, that have a lower degree of pedogenetic development, may present genetic horizons of alteration (“cambic” type according to WRB (IUSS Working Group 2014), although this does not imply significant textural changes affecting the availability of water. Although effective depth may be limited by the presence of continuous hard rock at medium depths (WRB leptic qualifier (IUSS Working Group 2014), the soil volume is sufficient, especially if it is of a suitable textural class (not coarse unbalanced, nor coarse or very stony) providing a water retention capacity capable of sufficiently mitigating some levels of water stress, thus reducing or buffering the physiological dysfunctions that lead to decline.
Other soil characteristics have not proven to be as decisive. Differences in biological (organic matter content) and chemical (soil reaction and presence of carbonates) properties do not appear to be related to the observed response. Although acidic or neutral reaction soils are associated with sites showing decline in all the cases analyzed, according to their autoecology (Gandullo and Sánchez 1994), Aleppo pine is tolerant to these acidity values (pH in water), so the effect on predisposition to decline is not sufficiently clear. However, the metabolic response to these chemical conditions could predispose the trees growing under these conditions to the synergistic effect of other factors that contribute to decline.
At Guadassuar, Lliria and Orihuela, the soils in sites with decline are similar to those in sites where decline is not present. In this case, the observed decline cannot be attributed to differences in intrinsic edaphic variability at site level, so the causes must be sought among other predisposing factors, such as: (a) the consequences of different disturbances which generate physiological dysfunctions (for example, soil compaction due to recreational use in Guadassuar); (b) the situation in locally drier landscape positions (Suarez et al. 2004) in the case of Lliria; (c) the exacerbated marginal climatic conditions (Gómez-Sanz 2019), as in the case of Orihuela. In this latter location, situated in a climatic environment which is clearly inadequate for the development of stable populations of the species, the good soil conditions (deep, texturally homogeneous profile) favor excessive extension of the root system in exacerbated non-drought years, making it hydraulically unviable when a relatively intense drought episode occurs (Martínez-Vilalta and Piñol 2002; Collins and Bras 2007) as was the case in 2014 in the studied area (García de la Serrana et al. 2015).
The decline of trees is a complex process in which all the factors involved interact with each other, driving multiple mechanisms that operate with different intensities across the landscape. Among these factors is the orientation of the site to the incident solar radiation, generating greater water stress in the Northern Hemisphere in sites with a marked southern orientation, especially in summer, when the demand for water is greater. Although empirical support is scarce and often contradictory, it is assumed that orientation is also a pedogenetic factor (Schaetzl and Anderson 2005), which enhances some processes that favor textural anisotropy on south-facing slopes. Consequently, the aspect of the slope and the easily explored depth may show highly correlated. The internal variability of soil quality in terms of its ability to store water and make it available to the vegetation efficiently is incorporated as a mechanism that synergistically contributes to the global effect attributable to orientation as a predisposition factor to decline.
Conclusions
Within a synoptic region prone to drought, specific episodes of decline are significantly associated with stands where the soils have a smaller easily explored volume, due to a low degree of development of the edaphic profile or the presence of a marked textural anisotropy in the soil, caused by the development of particularly clayey horizons, which cause transcendent changes in the soil’s capacity to offer water to the vegetation in a manner decoupled from the precipitation regime.
The conditions of internal variability of the soil, closely linked to its degree of pedogenetic development, have thus been identified as an important predisposing factor that modulates the effects of drought as a triggering factor for the decline of Aleppo pine stands in the Mediterranean biogeographic area.
The notion that the climate in the Mediterranean basin will evolve towards warmer and more arid scenarios is constantly garnering greater acceptance (Allen et al. 2010). The consequences of this predicted climate change on tree stands in general, and Aleppo pine stands in particular, will be highly nuanced by specific site conditions, especially those relating to the internal variability of soil conditions, giving rise to highly heterogeneous patterns of decline at local scale. The planning and implementation of adaptive management models should not, where possible, ignore this reality.
Data availability
The datasets generated during and/or analysed during the current study are available from the corresponding author on reasonable request.
References
Allen CD, Macalady AK, Chenchouni H, Bachelet D, McDowell N, Vennetier M, Kitzberger T, Rigling A, Breshears DD, Hogg EH, Gonzalez P, Fensham R, Zhang Z, Castro J, Demidova N, Lim J-H, Allard G, Running SW, Semerci A, Cobb N (2010) A global overview of drought and heat-induced tree mortality reveals emerging climate change risks for forests. For Ecol Manage 259:660–684. https://doi.org/10.1016/j.foreco.2009.09.001
Anderegg WR, Kane JM, Anderegg LD (2012) Consequences of widespread tree mortality triggered by drought and temperature stress. Nat Clim Chang 3:30–36. https://doi.org/10.1038/nclimate1635
Brown N, Vanguelova E, Pamell S, Broadmeadow S (2018) Predisposition of forests to biotic disturbance: Predicting the distribution of Acute Oak decline using environmental factors. Ecol Manage 407:145–154. https://doi.org/10.1016/j.foreco.2017.10.054
Brunner I, Graf-Pannatier E, Frey B, Rigling A, Landolt W, Dobbertin M (2009) Morphological and physiological responses of scots pine fine roots to water supply in a climatic dry area in Switzerland. Tree Physiol 29:542–550. https://doi.org/10.1093/treephys/tpn046
Camarero JJ, Gazol A, Sangüesa-Barreda G, Oliva J, Vicente-Serrano SM (2015) To die or not to die: early warnings of tree dieback in response to a severe drought. J Ecol 103:44–57. https://doi.org/10.1111/1365-2745.12295
Candel-Pérez D, Linares JC, Viñegla B, Lucas-Borja ME (2012) Assessing climate-growth relationships under contrasting stands of co-occurring Iberian pines along an altitudinal gradient. Ecol Manage 274:48–57. https://doi.org/10.1016/j.foreco.2012.02.010
Ciesla WM, Donaubauer ME (1994) Decline and dieback of trees and forests: a global overview. FAO, Rome
Collins DBG, Bras RL (2007) Plant rooting strategies in water-limited ecosystems. Water Resour Res 43:W06407. https://doi.org/10.1029/2006WR005541
Das AJ, Battles JJ, van Mantgem PJ, Stephenson NL (2008) Spatial elements of mortality risk in old-growth forests. Ecology 89:1744–1756. https://doi.org/10.1890/07-0524.1
Del Río M, Calama R, Montero G (2009) Selvicultura de Pinus halepensis Mill. In: Serrada R, Montero G, Reque J (eds) Compendio de selvicultura aplicada en España. INIA-FUCOVASA, Madrid, pp 289–313
Dobbertin M, Mayer P, Wohlgemuth T, Feldmeyer-Christe E, Graf U, Zimmermann NE, Rigling A (2005) The decline of Pinus sylvestris L. forests in the Swiss Rhone Valley—a result of drought stress? Phyton. Ann Rei Botanicae 45:153–156. https://www.dora.lib4ri.ch/wsl/islandora/object/wsl%3A2672
FAO (2006) Guidelines for soil descriptions, 4th edn. FAO, Rome
Fensham RJ, Holman JE (1999) Temporal and spatial patterns in drought-related tree dieback in Australian savanna. J Appl Ecol 36:1035–1050. https://doi.org/10.1046/j.1365-2664.1999.00460.x
Franklin JF, Shugart HH, Harmon ME (1987) Tree death as an ecological process. Bioscience 27:259–288. http://www.jstor.org/stable/1310665
Galiano L, Martínez-Vilalta J, Lloret F (2010) Drought-induced multifactor decline of scots pine in the pyrenees and potential vegetation change by the expansion of co-occurring oak species. Ecosystems 13(7):978–991. https://doi.org/10.1007/s10021-010-9368-8
Galiano L, Martínez-Vilalta J, Sabaté S, Lloret F (2012) Determinants of drought effects on crown condition and their relationship with depletion of carbon reserves in a Mediterranean Holm oak forest. Tree Physiol 32:478–489. https://doi.org/10.1093/treephys/tps025
Gandullo JM, Sánchez O (1994) Estaciones ecológicas De Los pinares españoles. ICONA, Madrid
García de la Serrana R, Vilagrosa A, Alloza JA (2015) Pine mortality in southeast Spain after an extreme dry and warm year: interactions among drought stress, carbohydrates and bark beetle attack. Trees. https://doi.org/10.1007/s00468-015-1261-9
Gea-Izquierdo G, Viguera B, Cabrera M, Cañellas I (2014) Drought induced decline could portend widespread pine mortality at the xeric ecotone in managed mediterranean pine-oak woodlands. Ecol Manage 320:70–82. https://doi.org/10.1016/j.foreco.2014.02.025
Gitlin AR, Sthultz CM, Bowker MA, Stumpf S, Paxton KL, Kennedy K, Muñoz A, Bailey JK, Whitham JK (2006) Mortality gradients within and among dominant plant populations as barometers of ecosystems change during extreme drought. Conserv Biol 20(5):1477–1486. https://doi.org/10.1111/j.1523-1739.2006.00424.x
Gómez-Sanz V (2019) Site- scale ecological marginality: evaluation model and application to a case study. Ecol Model 408:108739. https://doi.org/10.1016/j.ecolmodel.2019.108739
Greenwood DL, Weisberg PJ (2008) Density-dependent tree mortality in pinyon–juniper woodlands. Ecol Manage 255:2129–2137. https://doi.org/10.1016/j.foreco.2007.12.048
Guarin A, Taylor AH (2005) Drought triggered tree mortality in mixed conifer forests in Yosemite National Park, California, USA. Ecol Manage 218:229–244. https://doi.org/10.1016/j.foreco.2005.07.014
Hlaváčiková H, Novák V, Šimůnek J (2016) The effects of rock fragment shapes and positions on modeled hydraulic condcutivities of stony soils. Geoderma 281:39–48. https://doi.org/10.1016/j.geoderma.2016.06.034
Hosseini A, Hosseini SM, Linares JC (2017) Site factors and stand conditions associated with Persian oak decline in Zagros mountain forests. Forest Syst 26(3):e014, 13 pages. https://doi.org/10.5424/fs/2017263-11298
IUSS Working Group (2014) International soil classification system for naming soils and creating legends for soil maps. World reference base for Soil resources 2014. World Soil Resources Reports Nº 106. FAO, Rome
Jackson RB, Sperry JS, Dawson TE (2000) Root water uptake and transport: using physiological processes in global predictions. Trends Plant Sci 5(11):482–488. https://doi.org/10.1016/S1360-1385(00)01766-0
Klos RJ, Wang GG, Bauerle WL, Rieck JR (2009) Drought impact on forest growth and mortality in the southeast USA: an analysis using Forest Health and Monitoring data. Ecol Appl 19:699–708. https://doi.org/10.1890/08-0330.1
Koepke DF, Kolh TE, Adams HD (2010) Variation in woody plant mortality and dieback form severe drought among soils, plant groups, and species within a northern Arizona ecotone. Oecologia 163:1079–1090. https://doi.org/10.1007/s00442-010-1671-8
La Manna L, Rajchenberg M (2004) Soil properties and Austrocedrus chilensis forest decline in Central Patagonia, Argentina. Plant Soil 263:29–41. https://doi.org/10.1023/B:PLSO.0000047723.86797.13
Lloret F, Siscart D, Dalmases C (2004) Canopy recovery after drought dieback in holm-oak Mediterranean forests of Catalonia (NE Spain). Glob Chang Biol 10:2092–2099. https://doi.org/10.1111/j.1365-2486.2004.00870.x
Manion PD (1991) Tree disease concepts, 2nd edn. Prentice-Hall Inc, Upper Saddle River, NJ
Manion PD, Lachance D (1992) Forest decline concepts. APS Press, St Paul
Martínez-Vilalta J, Piñol J (2002) Drought-induced mortality and hydraulic architecture in pine populations of the NE Iberian Peninsula. Ecol Manage 161:247–256. https://doi.org/10.1016/S0378-1127(01)00495-9
McDowell N, Pockman WT, Allen CD, Breshears DD, Cobb N, Kolb T, Sperry J, West A, Williams D, Yepez EA (2008) Mechanisms of plant survival and mortality during drought: why do some plants survive while others succumb to drought? New Phytol 178:719–739. https://doi.org/10.1111/j.1469-8137.2008.02436.x
Miao SL, Zou CB, Breshears DD (2009) Vegetation responses to extreme hydrological events: sequence matters. Am Nat 173:113–118. https://doi.org/10.1086/593307
Nadal-Sala D, Sabaté S, Gracia C (2017) Importancia relativa de la profundidad del suelo para la resiliencia de Los bosques de pino carrasco (Pinus Halenpesis Mill) frente al incremento de aridez debido al cambio climático. Ecosistemas 26(2):18–26. https://doi.org/10.7818/ECOS.2017.26-2.03
Peñuelas J, Lloret F, Montoya R (2001) Severe drought effects on mediterranean woody flora in Spain. Sci 47(2):214–218. https://doi.org/10.1093/forestscience/47.2.214
Peterman W, Waring RH, Seager T, Pollock WL (2012) Soil properties affect pinyon pine – juniper response to drought. Ecohydrology 6(3):455–463. https://doi.org/10.1002/eco.1284
Preisler Y, Tatarinov F, Grünzweig JM, Bert D, Ogée J, Wingate L, Rotenberg E, Rohatyn S, Her N, Moshe I, Klein T, Yakir D (2019) Mortality versus survival in drought-affected Aleppo pine forest depends on the extent of rock cover and soil stoniness. Funct Ecol 00:1–12. https://doi.org/10.1111/1365-2435.13302
Prieto-Recio C, Martín-García J, Bravo F, Diez JJ (2015) Unravelling the associations between climate, soil properties and forest management in Pinus pinaster decline in the Iberian Peninsula. Ecol Manage 356:74–83. https://doi.org/10.1016/j.foreco.2015.07.033
Sánchez-Salguero R, Navarro-Cerrillo RM, Camarero JJ, Fernández-Cancio A (2012) Selective drought-induced decline of pine species in southeastern Spain. Clim Change 113(3–4):767–785. https://doi.org/10.1007/s10584-011-0372-6
Schaetzl R, Anderson S (2005) Soil: Genesis and Geomorphology. Cambridge University Press. https://doi.org/10.1017/CBO9780511815560
Schenk HJ, Jackson RB (2005) Mapping the global distribution of deep roots in relation to climate and soil characteristics. Geoderma 126:129–140. https://doi.org/10.1016/j.geoderma.2004.11.018
Six J, Bossuyt H, Degryze S, Denef K (2004) A history of research on the link between (micro)aggregates, soil biota, and soil organic matter dynamic. Soil till Res 79:7–31. https://doi.org/10.1016/j.still.2004.03.008
Soil Survey Division Staff (2017) Soil Survey Manual. United States Department of Agriculture Handbook No. 18. https://www.nrcs.usda.gov/sites/default/files/2022-09/The-Soil-Survey-Manual.pdf. Accesed 15 Jan 2024
Suarez ML, Ghermandi L, Kitzberger T (2004) Factors predisposing episodic drought-induced tree mortality in Nothofagus: site, climatic sensitivity and growth trends. J Ecol 92:954–966. https://doi.org/10.1111/j.1365-2745.2004.00941.x
Acknowledgements
We would like to thank Emilio González and Eduardo Péres Laorga, technical staff of the “Servicio de Ordenación y Gestión Forestal de la Consellería D’Infraestructures, Territori I Medi Ambient de la Generalitat Valenciana”, for making the study possible, along with the staff and service agents who carry out Forest Health related functions in VAERSA and in the Consellería for their help in facilitating the field work.
Funding
Open Access funding provided thanks to the CRUE-CSIC agreement with Springer Nature. The authors declare that no funds, grants, or other support were received during the preparation of this manuscript.
Author information
Authors and Affiliations
Contributions
The study conception and design were carried out by Valentín Gómez-Sanz and Rafael Serrada Hierro. All authors performed in data collection and analysis. The draft of the manuscript was written by Valentín Gómez-Sanz and all authors provided comments and suggestions on it and approved manuscript final version.
Corresponding author
Ethics declarations
Competing Interests
The authors have no relevant financial or non-financial interests to disclose.
Additional information
Responsible Editor: Timothy J. Fahey.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Gómez-Sanz, V., Gastón, A., García-Viñas, J.I. et al. Site-scale soil conditions influencing the decline of Aleppo pine stands in Mediterranean Spanish woodland. Plant Soil (2024). https://doi.org/10.1007/s11104-024-06638-2
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s11104-024-06638-2