
Abstract
Heterobasidion root rot causes severe problems in the coniferous dominated forests of Northern hemisphere by decreasing timber value, reducing tree growth and making trees prone to other disturbance agents such as wind and bark beetles. According to current practices an infected stand is protected by treating fresh stumps at final cutting and generating the stand with tree species other than the host to reduce the current infection levels. However, prevailing guidelines do not provide instructions how to manage the neighbouring stands around the already infected stand. In contrast to earlier studies, we expand the analysis to an entity (a landscape) consisting of both infected and uninfected stands so that stand establishment of uninfected area is taken into account, too. The objective of the study is to optimize stand establishment of the uninfected area so that the revenue losses are simultaneously minimized when the average carbon storage is maximized within the whole landscape (infected + uninfected). Our results demonstrate the complexity of Heterobasidion root rot management: the optimum stand establishment strategy differs depending on the geographic location and interest rate (i.e., yield on capital). Further, the results implicate that for successful fight against Heterobasidion root rot the magnitude of infected area as well as the severity of the infection need to be taken into account.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Pathogenic fungi of the Heterobasidion annosum (Fr.) Bref. complex infecting coniferous trees cause severe problems for forestry throughout the Northern Hemisphere. In Europe, disease is caused by species H. annosum s.s., H. parviporum, H. abietinum, and H. irregulare, of which the first two are the most significant. H. annosum s.s. is a generalist considering host species and can cause symptoms on many coniferous and deciduous species, whereas H. parviporum is specialized mainly on Norway spruce (Picea abies (L.) Karst.). It has been estimated that Heterobasidion root rot has caused approximately €790 million annual economic losses in the European Union, mainly due to growth reduction and degradation of wood. Alone in Sweden and Finland the annual economic losses due to Heterobasidion root rot are valued to be €120 million (Woodward et al. 1998). These costs do not include costs of controlling and diagnosing the disease nor the cost of reduced resistance of the infected stands to storm damages (Gonthier and Thor 2013). The fungus initiates primary infections to previously healthy stands via airborne basidiospores which need freshly cut stumps or wounded trees for successful infection. From infected trees or stumps the fungus spreads vegetatively with mycelium to adjacent trees via root contacts and connections at a rate of up to 200 cm per year causing the secondary infections within the already infected stand (Asiegbu et al. 2008; Garbelotto and Gonthier 2013). Warming climate will enhance the activity of Heterobasidion root rot (Müller et al. 2015), potentially increasing the economic losses in the future.
The economically most important host of Heterobasidion root rot in coniferous-dominated northern European forests is Norway spruce. In Norway spruce the decay develops mainly in the heartwood of the stem and the decay column can proceed high in the stem without noticeable reduction in tree vigour (Vollbrecht and Agestam 1995). This decreases the value of timber and timber-sized trees must be used as lower value pulp or energy wood. During pulp production, the decay decreases the amount of fibres and also more chemicals are needed in the bleaching process (Mäkelä et al. 1998). In addition to these direct losses Heterobasidion root rot also reduces the growth of the tress causing indirect losses (reviewed in Stenlid and Redfern 1998; Oliva et al. 2010). The trees respond to fungal infection by allocating carbon reserves where needed to protect water and energy transport within the roots and stem, e.g., in Norway spruce formation of a specific reaction zone and at the border of heartwood and sapwood (Bendz-Hellgren and Stenlid 1995). Further, Heterobasidion root rot creates also other indirect losses by increasing time consumption of harvesting due to extra bucking and sorting of decayed wood (Honkaniemi et al. 2019).
Once a stand is infected, Heterobasidion root rot is impossible to eradicate (Müller et al. 2018). According to current practices uninfected, high primary infection risk areas can be protected from novel spore infections by treating freshly cut stumps with either urea or biological control agent Phlebiopsis gigantea (Fr.) Jülich (Rishbeth 1963; Garbelotto and Gonthier 2013). To reduce secondary infections, the stand can be established with less susceptible tree species to reduce the current infection levels over time (Garbelotto and Gonthier 2013). Thus, current practices do not explicitly consider how the neighbouring stands, let alone a landscape should be managed against Heterobasidion root rot. Further, recent literature on Heterobasidion root rot either focuses on the stand level (e.g., Aza et al. 2021) or on comparing rot-infested spruce stands to healthy pine stands in economic terms (Aza et al. 2022). The diversification of forest landscapes creates resilience to many different disturbances (Jactel et al. 2017) and a similar approach could be tested also with root rot to reduce the potential for future infections.
Here we focus on managing Norway spruce stands and assume the stands to be infected with the more common Heterobasidion parviporum solely infecting Norway spruce. More specifically this study addresses whether there is an optimum stand establishment strategy with alternative tree species for the uninfected forest area when the proportion (%) of infected forests increases assuming we continue with spruce management in the infected area. In other words, we accepted that the infected forest area will gradually increase despite the stump treatment, and that area is left for spruce management while the diminishing uninfected forest area is optimized by stand establishment with alternative tree species. The rationale of this study is to find out (a) how much timber revenues is lost with the optimal management (compared to scenario without any infection) and (b) can carbon sequestration be enhanced by optimum stand establishment strategy? Optimization involves simultaneously minimizing timber revenue losses and maximizing average carbon storage within the whole forest area (henceforth denoted as landscape), consisting of infected and uninfected forest areas. Then, how do the primary (spore density) and secondary infection rates (number of infected trees in the previous generation) within the infected forest area affect to the optimal solution for the whole landscape?
Materials and methods
Study regions and stands
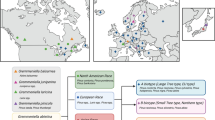
To include temperature gradient into analyses three locations where root rot frequently occur were chosen: Salo, Lahti and Viitasaari (Fig. 1). In each location the site type was identical, nutrient-rich site on mineral soil (corresponding Myrtillus type in the Finnish classification system, see Tonteri et al. 1990) favouring Norway spruce management. The temperature sum (cumulative sum of temperatures during the growing season with a threshold of 5℃) of the southernmost location, Salo is 1297degree days, d.d. while in the northernmost, Viitasaari the temperature sum corresponds to 1106 d.d (in Lahti 1251 d.d.). Temperature sum affects both tree and root rot growth rates in the models. Temperature sum affects directly the annual height and diameter increment of trees (e.g., Henttonen et al. 2017), as well as modifies the rate of decay growth in roots and stem parts leading to faster expansion of infected clusters (Honkaniemi et al. 2017). The length of the growing season in Finland varies considerably due to temperature, ranging from 170 to 220 days in southernmost Finland where Salo and Lahti are located, and 140 to 180 days in central Finland where Viitasaari is located (Venäläinen et al. 2005).

Three locations (Salo, Lahti and Viitasaari) of the analyses
Stand projections
Uninfected stand projections were generated with Motti stand simulator, a stand-level decision-support tool for assessing the effects of forest management on stand dynamics (Salminen et al. 2005; Hynynen et al. 2015). The core of Motti is a stand simulation module consisting of stand-level and individual-tree level models, both based on an empirical-statistical modelling approach (e.g., Hynynen et al. 2015). In brief, natural regeneration and early growth models are based on stand-level modelling, while for mature trees (dominant height > 7 m), predictions are based on sample trees each representing a certain number of trees and simulated with individual tree models (for technical details on the models, see Juutinen et al. 2018 Suppl. Data). The Motti stand simulator has been widely applied at stand level (e.g., Honkaniemi et al. 2014; Ahtikoski and Hökkä 2019). Uninfected stands were simulated as starting from a bare land and managed according to silvicultural guidelines (Äijälä et al. 2014). Therefore, the simulations corresponded to practical forestry. The simulations were made for pure spruce, pure pine and pure birch stands, as well as a mixture of spruce and birch. The resulting management regimes are presented in Table 1.
In the case of Heterobasidion root rot-infected stands, the disease dynamics were simulated with Hmodel (Honkaniemi et al. 2014, 2019) and the stand dynamics with Motti. Hmodel is a mechanistic model based on the biology of the fungal agent causing the disease. Hmodel creates a spatially explicit stand based on the sample trees created in Motti stand simulator. The main input variables to define the Heterobasidion spp. spread potential in the simulated tree generation are primary infection pressure (i.e., spore density) and the secondary infection pressure (i.e. the number of infected stumps in previous stand). The applied stump treatment affects only by reducing primary spore infections. During the simulations, the secondary root rot infections spread within the stand and Hmodel explicitly simulates the decay development within stems (see Honkaniemi et al. 2014) causing growth losses, tree mortality and decreasing timber quality (Honkaniemi et al. 2014, 2019). Ten simulations were executed for each scenario combination due to stochasticity in the model (Honkaniemi et al. 2014). In this study we applied two alternative scenarios of primary and secondary infection pressure:1) combination of 200 spores m− 2 hour− 1 (primary infection) and 10% infected stems per hectare in the previous tree generation (secondary) and 2) 800 spores m − 2 hour− 1 and 30% infected stems per hectare in the previous tree generation. The former combination (1) is referred later as mild and the latter (2) as severe root rot infection (see Honkaniemi et al. 2019). The quality of stump treatment was assumed to be good (for classification of stump treatment, see Honkaniemi et al. 2019). The simulated management regimes of infected forest area are also presented in Table 1.
Financial and carbon assessments
According to the stand projections associated with each stand establishment option the bare land value, BLV was assessed by the following. The bare land value, BLV (interchangeably land expectation value, LEV) represents the present value of net revenues from growing an infinite series identical timber stand (Chang 2020). BLV enables theoretically solid financial comparison between management regimes regardless of their time horizons (Amacher et al. 2009). Maximizing the bare land value yields the discounted economic surplus over an infinite time horizon (Faustmann 1849). Let\({Z_{{t_i}}}\)denote standing volume (m3 ha− 1) before the ith thinning at age ti, i > 0,…,T (t0 and tT denote the beginning and the end of rotation, respectively), k denotes timber assortments (k = 1,…,K) and pk the stumpage price (€ m− 3) of each timber assortment. Let b be the discount factor s.t. b = 1/(1 + r) where r is the interest rate in real terms (here 2%). An interest rate describes an annual yield over capital invested and discounting exerts a crucial effect on the valuation, particularly with long time periods (e.g., Price 2018). The removal (in m3) of each timber assortment k in ith thinning is denoted by hki. The removal is a function of stand state, thinning intensity, and timing. Thinning intensity (removal relative to the growing stock) in ith thinning is gi. Using these notations bare land value, BLV (€ ha− 1) is calculated according to:
where \(sc_{{s{t_l}}}^{n}\) is the cost (€ ha− 1) of a silvicultural action s at stand age tl, € ha− 1 (Note that i ≠ l and M < T). Having calculated the BLVs the next step was to assess the average carbon storage, Ca for each stand establishment option.
Carbon storage was calculated for living and dead biomass with models incorporated into Motti. Living biomass was estimated by biomass compartments (stems, branches, needles/leaves, stumps and roots) using biomass models of Repola (2008, 2009). Dead biomass consists of natural mortality and logging residues, and the prediction of mortality is based on individual-tree survival model, individual-tree model for age-related mortality and stand-level model for self-thinning (Hynynen et al. 2014). Then, decomposition of above-ground dead biomass was estimated using models of Mäkinen et al. (2006). For each stand establishment option an annual average carbon storage Ca was estimated.
Optimization
Finally, the landscape-level optimization (nonlinear) problem describing the Heterobasidion root rot-infected and uninfected forest area is formulated. For simplicity we assume a 100 hectare landscape consisting of 100 individual stands, each 1 hectare. A root rot-infected hectare is denoted by di and uninfected (i.e. healthy) by dh so that \(\sum _{i}^{I}{d}_{i}\)\(+\sum _{h}^{H}{d}_{h}=\)100. For the sake of division between uninfected and infected forest area the decreased BLV and Ca in the infected area are denoted by BLVde and Cade, respectively. Further, for the whole 100 hectare landscape the best financial outcome is to establish all stands with spruce, which corresponds to a situation with zero infection, \(\sum _{h=1}^{100}{d}_{h}{BLV}_{spruce}\). Now the optimization problem can be presented by:
, where BLVz (Cz) refers to bare land value (average carbon storage) associated with a stand establishment option z, z corresponding to spruce, pine, birch or mixed-species forest stand. Note that \({BLV}_{spruce}^{de}\) < \({BLV}_{spruce}\) due to infection of H. annosum. Equation (2) minimizes the loss of BLV compared to the situation without any infection in the whole landscape (100 hectares) while Eq. (4) maximizes the average carbon storage for the landscape when there already exists an infected area \(\sum _{i}^{I}{d}_{i}\). The rationale is to find out the mimimum financial loss corresponding to maximum carbon storage – such an optimization problem resembles of a dual approach in optimization theory (see, e.g.Pavoni et al. 2018) which often requires nonsmooth optimization approaches (e.g. Bagirov et al. 2014; Montonen et al. 2019). As a sideproduct the optimum solution [Eqs. (2), (3) and (4)] provides a stand establishment strategy to be applied in practical forestry. Had we minimized only the BLV loss or maximized the carbon storage separately this would have been trivial since the “optimum solution” associated with the healthy forest area would have solely consisted of individual stands, which provided the highest BLV or carbon storage, respectively. Indeed, this would have not even required any optimization – just picking up the best performers of BLV and carbon storage separately would have resulted in the “solution”. Instead, both objectives (min BLV loss and max carbon storage) were continuous variables indicating that technically this was not a mixed-integer nonlinear, nor a bilevel optimization problem (cf. Colson et al. 2007). However, it is the dual nature of the optimization problem which creates the challenge here.
Technically, the optimization problem was solved by using a Solver application of Excel spreadsheet (Microsoft 365 Apps for enterpise). However, the Solver was incorporated with a subroutine (coded with Microsoft Visual Basic for Applications, ver 7.1 ) which maximized the average carbon storage, i.e. Equation (3) by iterating the value in a loop with a threshold (convergence) set to 0.5% value change. At the same time the Solver mimimized the BLV loss, Eq. (2). The 0.5% convergence value (criterion) is considered to be accurate enough for this kind of optimization problem – however, slightly smaller (Pohjola and Valsta 2007) and larger values (Pukkala et al. 2010) can be found in the literature. Finally, to restrict the computing time we decided to set \(\sum _{i}^{I}{d}_{i}\) to 10, 20,… ,60 and correspondingly \(\sum _{h}^{H}{d}_{h}\) to 90, 80,…,40. In practice this meant that a solution was sought (through optimization) for each combination of infected and uninfected forest area, starting with the combination of 10 (infected) and 90 (uninfected) hectares and ending with 60 (infected) and 40 (uninfected) hectares.
Financial data
The stumpage prices and per unit silvicultural costs used were based on the time series of 10-yr period covering years 2011–2020. This was the most recent time series available for both the prices and costs (http://statdb.luke.fi ). The nominal values (both stumpage prices and silvicultural costs) were converted to real prices through deflation (applying the cost-of-living index of Statistics Finland 2020; http://www.stat.fi/til/khi/index_en.html) after which arithmetic averages were calculated. The averages stumpage prices, silvicultural costs and stump treatment costs are presented in Table 2. The stumpage prices for decayed timber were for low-cost pulp (< 50% decay at stump level) 75% of the pulpwood price and the energywood (> 50% of decay at stump level) 50% of the pulpwood price.
Sensitivity analysis
The effect of interest rate on optimal solution (Eqs. 2–4) was tested in Salo with 2% and 4% interest rates when a severe root rot infection was assumed.
Results
The results distinctively demonstrate three findings. First, the optimal stand establishment strategy (i.e., how the uninfected forest area would be regenerated to increase tree species diversity and reduce root rot risk) varies between geographic locations (Fig. 2a, b). For instance, with 10% infected forest area (90% uninfected) the optimal stand establishment indicates that 89% of the uninfected forest area should be regenerated with birch in Salo while in Lahti 32% should be regenerated with pine, when the root rot infection is mild (Fig. 2a). Second, with increasing infected forest area, the optimal establishment strategy changes: with 50% infected area the corresponding percentages would be 53% and 45% of uninfected area, respectively (Fig. 2a). Third, with severe root rot infection the optimal establishment strategy is different compared to mild infection. When the infected forest area is 30% (70% uninfected) and the root rot infection mild, the optimal establishment strategy for Salo would be to regenerate 23% with birch and 77% with spruce while with severe root rot infection the percentages would be 61% and 39%, respectively (Fig. 2a, b). Finally, pine dominates in the northernmost Viitasaari (see Fig. 1) regardless of the severity of root rot infection and the proportion of infected forest area (Fig. 2a, b).

a. Optimal stand establishment strategy (regeneration with different tree species) for the uninfected area in Salo, Lahti and Viitasaari with increasing infected area (from 10 to 60% of 100 hectares) assuming a mild root rot infection with 200 spores per m2 hour− 1 and 10% infected stumps in the previous generation. Interest rate 2% applied in optimization (see Eq. 2). b. Optimal stand establishment strategy for the uninfected area in Salo, Lahti and Viitasaari with increasing infected forest area (from 10–60%) assuming a severe root rot infection with 800 spores per m2 hour− 1 and 30% infected stumps in the previous generation. Interest rate 2% applied
As anticipated the revenue losses associated with maximum average carbon storage slightly increase with increasing infected area (Table 3). Further, in Viitasaari with 10% infected forest area there were actually no revenue losses (Table 3) – this is due to the fact that pine dominates both financially and carbon-wise in Viitasaari (see Table 1).
Sensitivity analysis revealed that interest rate had a distinctive impact on the results (Fig. 3). Namely, with higher interest rate (4%) the proportion of birch in optimal establishment strategy was considerably higher than with 2% (Fig. 3).

Effect of interest rate (2% and 4%) on optimal stand establishment strategy in Salo with increasing infected forest area according to severe root rot infection, %. Infected forest area ranges from 10–60%
Discussion
So far financial analyses on Heterobasidion root rot have focused on the stand level (see Honkaniemi et al. 2019; Aza et al. 2021) while the disease dynamics evidently suggest that landscape-level assessment would be desirable to better capture the characteristics of the disease. Here, we have expanded the existing literature to an analysis over a landscape consisting of multiple stands and different tree species analysing the optimal management practices for financial performance and carbon sequestration.
In this study we introduced a framework in which a landscape was divided into uninfected (healthy) and infected (Heterobasidion root rot) areas, and the goal was to optimize stand establishment through minimizing the revenue losses and maximizing the average carbon storage within the whole landscape (infected + uninfected area). Stand establishment here relates to regeneration with different tree species in the healthy forest area with increasing proportion of the infected area. This kind of framework with simultaneous minimizing and maximising objectives resembles a dual approach in optimization theory with high complexity and nonlinear features (Pavoni et al. 2018).
Prior to concluding a few issues need to be raised. First, this study was primarily based on simulations on tree growth. In general simulation is a powerful methodology but it has several faults (see Koivisto 2017) of which the interpretation pitfall might be relevant here. In brief, we have first produced stand projections by simulations to describe stand structure and dynamics of tree growth, and then applied the simulations as input to the optimization. In such a procedure there is a chance which involves losing critical distance to the initial simulation results, which might complicate the interpretation of the results. However, we consider that the distance in this study (from initial simulations to optimization) is short enough not to create active problems in interpretation of the optimization results. Then we simplified the set-up by assuming only one site type - in practice a forested landscape (such as 100 hectares here) usually consists of different site types on both mineral soils and peatlands. Finally, the analyses focused only on carbon sequestration and timber revenues without addressing other management goals such as biodiversity and recreation known to be important, too.
The results distinctively demonstrated the complexity of Heterobasidion root rot management, even within a simplified setting. In terms of forest management there is no “one size fits all” recipe. Instead the best stand establishment strategy depends strongly on the geographic location through temperature sum, proportion of infected forest area (%) and even on the severity of infection (expressed as spore density and number of infected stumps in previous tree generation). For instance, with identical proportion of infected forest area (%) and identical severity of infection there is a totally different optimal stand establishment strategy (within the uninfected forest area) in Salo, Lahti and Viitasaari – even though the three locations and their temperature sums are not so far apart from each other (max. distance between the locations 330 km). This calls for more precise forest management adapted for each location. The optimal stand establishment strategy also changes nonlinearly with increasing proportion of infected forest area, which complicates the interpretation of suitable forest management even further. In a nutshell, however, one could say that in Viitasaari (the most northern location) pine is dominated in optimal stand establishment mainly due to the fact that pine is better acclimatized for prevailing climatic conditions there than other tree species while in Salo (the most southern) the same applies for spruce (this can be depicted in Table 1 by comparing cutting removals and average carbon storage between tree species).
Further, our results highlight the importance of interest rate – optimal establishment strategy is distinctively dependent on the interest rate applied (Fig. 3). There is a logical explanation for this: in the optimization problem bare land value, BLV is calculated according to the interest rate (Eq. (2) resulting in different solutions for different interest rates. For instance, the difference in BLV calculated with 2% and with 4% is less for birch than other tree species in absolute values (€ ha− 1) while the average carbon storage is insensitive to interest rate (i.e., intact). This leads to favor birch in stand establishment with 4% interest rate. The intriguing question would be: which interest rate is relevant for such analyses? According to Price (2018) suggested interest rates in UK, Norway and France fluctuated between 2% and 4% when the time horizon of the analyses falls between 30 and 200 years. This study operated with time horizon from 58 to 70 years (Table 1) applying interest rate 2% and 4%.
In conclusion, our work shows that simultaneous optimization of revenues and carbon sequestration for a forest landscape partially infected by Heterobasidion root rot is possible. Our results add to the growing literature base with the importance of landscape-scale forest management and planning to mitigate spatio-temporally correlated forest disturbances instead of focusing on the management of individual stands. However, the landscape-scale dynamics are highly complex, and the results difficult to be reduced to simple, universally applicable guidelines of forest management.
Data availability
All authors confirm that all data and materials as well as software application or custom code support their published claims and comply with field standards. Further, all data are located in corresponding author’s OneDrive, and they can easily be accessed upon request.
References
Ahtikoski A, Hökkä H (2019) Intensive forest management – does it financially pay off on drained peatlands? Can J for Res 49:1101–1111. https://doi.org/10.1139/cjfr-2019-0007
Äijälä O, Koistinen A, Sved J, Vanhatalo K, Väisänen P (2014) Metsänhoidon suositukset [Silvicultural recommendations]. The Forestry Development Centre Tapio. Helsinkin, Finland. (In Finnish).
Amacher GS, Ollikainen M, Koskela E (2009) Economics of forest resources. The MIT, Cambridge, p 448
Asiegbu FO, Adomas A, Stenlid J (2008) Conifer root and butt rot caused by Heterobasidion Annosum (Fr.) Bref.s.l. Mol Plant Pathol 6:395–409. https://doi.org/10.1111/j.1364-3703.2005.00295.x
Aza A, Kangas A, Gobakken T, Kallio AMI (2021) Effect of root and but rot uncertainty on optimal harvest schedules and expected incomes at the stand level. Ann Sci 78:70. https://doi.org/10.1007/s13595-021-01072-1
Aza A, Kallio AMI, Pukkala T, Hietala A, Gobakken T, Astrup R (2022) Species selection in areas subjected to risk of root and butt rot: applying Precision forestry in Norway. Silva Fenn 56 no 3 article 10732. https://doi.org/10.14214/sf.10732
Bagirov A, Karmitsa N, Mäkelä MM (2014) Comparing different nonsmooth minimization methods and software. Optim Methods Softw 27:131–153. https://doi.org/10.1080/10556788.2010.526116
Bendz-Hellgren M, Stenlid J (1995) Long-term reduction in the diameter growth of butt rot affected Norway spruce, Picea abies. For Ecol Manage 74:239–243. https://doi.org/10.1016/0378-1127(95)03530-N
Chang SJ (2020) Twenty-one years after the publication of the generalized Faustmann formula. For Policy Econ 118(2020):102238. https://doi.org/10.1016/j.forpol.2020.102238
Colson B, Marcotte P, Savard G (2007) An overview of bilevel optimization. Ann Oper Res 153:235–256. https://doi.org/10.1007/s10479-007-0176-2
Faustmann M (1849) Berechnung Des Werthes, Welchen Waldboden, sowie noch nicht haubare Holzbestände für die Waldwirthschaft besitzen.[Calculation of the value which forest land and immature stands possess for forestry]. Allgemeine Forst- und Jagd-Zeitung 25:441–455
Garbelotto M, Gonthier P (2013) Biology, epidemiology and control of Hererobasidion species worldwide. Annu Rev Phytopathol 51:39–59. https://doi.org/10.1146/annurev-phyto-082712-102225
Gonthier P, Thor M (2013) Annosus root and but rots. In: Gonthier P, Nicolotti G (eds) Infectious forest diseases. CABI International, Wallingford, pp 128–158
Henttonen HM, Nöjd P, Mäkinen H (2017) Environment-induced growth changes in the Finnish forests during 1971–2010 – an analysis based on National Forest Invetory. For Ecol Manage 386:22–36. https://doi.org/10.1016/j.foreco.2016.11.044
Honkaniemi J, Ojansuu R, Piri T, Kasanen R, Lehtonen M, Salminen H, Kalliokoski T, Makinen H (2014) Hmodel, a Heterobasidion annosum model for even-aged Norway spruce stands. Can J for Res 44(7):796–809. https://doi.org/10.1139/cjfr-2014-0011
Honkaniemi J, Piri T, Lehtonen M, Siipilehto J, Heikkinen J, Ojansuu R (2017) Modelling the mechanisms behind the key epidemiological processes of the conifer pathogen Heterobasidion Annosum. Fungal Ecol 25:29–40. https://doi.org/10.1016/j.funeco.2016.10.007
Honkaniemi J, Ahtikoski A, Piri T (2019) Financial incentives to perform stump treatment against Heterobasidion root rot in Norway spruce dominated forests, the case of Finland. For Policy Econ 105:1–9. https://doi.org/10.1016/j.forpol.2019.05.015
Hynynen J, Salminen H, Ahtikoski A, Huuskonen S, Ojansuu R, Siipilehto J, Lehtonen M, Eerikäinen K (2015) Long-term impacts of forest management on biomass supply and forest resource development: a scenario analysis for Finland. Eur J Res 134:415–431. https://doi.org/10.1007/s10342-014-0860-0
Hynynen J, Salminen H, Huuskonen S, Ahtikoski A, Ojansuu R, Siipilehto J Lehtonen Rummukainen M, Kojola A, Eerikäinen S (2014) K Scenario analysis for the biomass supply potential and the future development of Finnish forest resources. Working Papers of the Finnish Forest Research Institute 302. Available at http://www.metla.fi/julkaisut/workingpapers/2014/mwp302.htm
Jactel H, Bauhus J, Boberg J et al (2017) Tree diversity drives Forest stand resistance to natural disturbances. Curr Forestry Rep 3:223–243. https://doi.org/10.1007/s40725-017-0064-1
Juutinen A, Ahtikoski A, Lehtonen M, Mäkipää R, Ollikainen M (2018) The impact of a short-term carbon payment scheme on forest management. For Policy Econ 90:115–127. https://doi.org/10.1016/j.forpol.2018.02.005
Kärhä K, Koivusalo V, Palander T, Ronkanen M (2018) Treatment of Picea abies and Pinus sylvestris stumps with Urea and Phlebiopsis gigantea for control of Heterobasidion. Forests 2018, 9(3), 139. https://doi.org/10.3390/f9030139
Koivisto M (2017) Pitfalls in modelling and simulation. 6th Internat. Young Scientists Conference in HPC and simulation, YSC 2017, 1–3 Nov. 2017, Kotka, Finland. Procedia Computer Science 119: 8–15. https://doi.org/10.1016/j.procs.2017.11.154
Mäkelä M, Lipponen K, Sainio M (1998) Tyvilahoa sisältävän kuusen määrä, laatu ja käyttömahdollisuudet sellun raaka-aineena. [Butt rot in spruce: quality, amount and potential as a raw material for cellulose]. Metsäteho Report 50 (In Finnish).
Mäkinen H, Hynynen J, Siitonen J, Sievänen R (2006) Predicting the decomposition of Scots pine Norway spruce, and birch stems in Finland. Ecol Appl 16(5): 1865–1879. https://www.jstor.org/stable/40061757
Montonen O, Ranta T, Mäkelä MM (2019) Planning the schedule for the disposal of the spent nuclear fuel with interactive multiobjective optimization. Algorithms 12(12):252. https://doi.org/10.3390/a12120252
Müller MM, Hamberg L, Kuuskeri J, laPorta N, Pavlov L, Korhonen K (2015) Respiration rate determinations suggest Heterobasidion parviporum subpopulations have potential to adapt global warming. For Pathol 45:515–524. https://doi.org/10.1111/efp.12203
Müller MM, Henttonen HM, Penttilä R, Kulju M, Helo T, Kaitera J (2018) Distribution of Heterobasidion butt rot in northern Finland. For Ecol Manage 425:85–91. https://doi.org/10.1016/j.foreco.2018.05.047
Oliva J, Thor M, Stenlid J (2010) Reaction zone and periodic increment decrease in Picea abies trees infected by Heterobasidion Annosum s.l. For Ecol Manage 260:692–698. https://doi.org/10.1016/j.foreco.2010.05.024
Pavoni N, Sleet C, Messner M (2018) The dual approach to recursive optimization: theory and examples. Econometrica 86:133–172. https://doi.org/10.3982/ECTA11905
Pohjola J, Valsta L (2007) Carbon credits and management of scots pine and Norway spruce stands in Finland. For Policy Econ 9:789–798. https://doi.org/10.1016/j.forpol.2006.03.012
Price C (2018) Declining discount rate and the social cost of carbon: forestry consequences. J for Econ 3:39–45. https://doi.org/10.1016/j.jfe.2017.05.003
Pukkala T, Lähde E, Laiho O (2010) Optimizing the structure and management of uneven-sized stands of Finland. Forestry 83:129–142. https://doi.org/10.1093/forestry/cpp037
Repola J (2008) Biomass equations for birch in Finland. Silva Fenn 42:605–624. https://doi.org/10.14214/sf.236
Repola J (2009) Biomass equations for scots pine and Norway spruce in Finland. Silva Fenn 43:625–647. https://doi.org/10.14214/sf.184
Rishbeth J (1963) Stump protection against Fomes Annosus. Ann Appl Biol 52(1):63–77. https://doi.org/10.1111/j.1744-7348.1963.tb03728.x
Salminen H, Lehtonen M, Hynynen J (2005) Reusing legacy FORTRAN in the MOTTI growth and yield simulator. Comput Electron Agr 49:103–113. https://doi.org/10.1016/j.compag.2005.02.005
Stenlid J, Redfern DB (1998) Spread within the Tree and Stand. In: Woodward S, Stenlid J, Karjalainen R, Hu ̈ttermann A (eds.) Heterobasidion annosum: Biology, Ecology, Impact and Control. CABInternational, Wallingford, New York, pp. 125–141
Tonteri T, Hotanen JP, Kuusipalo J (1990) The Finnish forest site type approach: ordination and classification studies of mesic forest sites in southern Finland. Vegetatio 87:85–98. https://doi.org/10.1007/BF00045658
Venäläinen A, Tuomenvirta H, Pirinen P, Drebs A (2005) A basic Finnish climate data set 1961–2000 - description and illustrations. Finnish Meteorological Institute, Helsinki, Finland, Reports 2005:5
Vollbrecht G, Agestam E (1995) Modelling incidence of root rot in Picea abies plantations in southern Sweden. Scand J for Res 10:74–81. https://doi.org/10.1080/02827589509382870
Woodward S, Stenlid J, Karjalainen R, Hüttermann A (1998) Heterobasidion annosum: biology, ecology, impact and control. CAB International, Wallingford, UK. ISBN-0-85199-275-7
Acknowledgements
We would like to thank Specialist Esko Oksa for generating an excellent map (Fig. 1). A math student Veeti Ahtikoski at the University of Turku is also appreciated for conducting majority of the optimizations. This research was supported by the TyviTuho project funded by the Ministry of Agriculture and Forestry of Finland through the Catch the Carbon Research and Innovation program (funding decision VN/5206/2021).
Funding
Open access funding provided by Natural Resources Institute Finland.
Author information
Authors and Affiliations
Contributions
AA Methodology, Conceptualization, Formal analysis, Investigation, Writing—Original Draft, Writing-Editing. JH Investigation, Conceptualization, Methodology, Data curation, Writing- Final Review and Editing. EH Conceptualization, Methodology, Data curation, Writing- Final Review and Editing, Project administration. MP Resources, Conceptualization, Investigation, Writing-Final Review and Editing.
Corresponding author
Ethics declarations
Competing interests
There are no competing interests related to this manuscript. The authors have no relevant financial or non-financial interests to disclose. The authors have no financial or proprietary interests in any material discussed in this article.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Ahtikoski, A., Honkaniemi, J., Holmström, E. et al. Optimal species composition for stand establishment in root-rot infected forest areas. New Forests (2024). https://doi.org/10.1007/s11056-024-10046-w
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s11056-024-10046-w