
Abstract
Aspergillus fumigatus is a saprophytic fungal pathogen that causes opportunistic infections in animals and humans. Azole resistance has been reported globally in human A. fumigatus isolates, but the prevalence of resistance in isolates from animals is largely unknown. A retrospective resistance surveillance study was performed using a collection of clinical A. fumigatus isolates from various animal species collected between 2015 and 2020. Agar-based azole resistance screening of all isolates was followed by in vitro antifungal susceptibility testing and cyp51A gene sequencing of the azole-resistant isolates. Over the 5 year period 16 (11.3%) of 142 A. fumigatus culture-positive animals harbored an azole-resistant isolate. Resistant isolates were found in birds (15%; 2/13), cats (21%; 6/28), dogs (8%; 6/75) and free-ranging harbor porpoise (33%; 2/6). Azole-resistance was cyp51A mediated in all isolates: 81.3% (T-67G/)TR34/L98H, 12.5% TR46/Y121F/T289A. In one azole-resistant A. fumigatus isolate a combination of C(-70)T/F46Y/C(intron7)T/C(intron66)T/M172V/E427K single-nucleotide polymorphisms in the cyp51A gene was found. Of the animals with an azole-resistant isolate and known azole exposure status 71.4% (10/14) were azole naive. Azole resistance in A. fumigatus isolates from animals in the Netherlands is present and predominantly cyp51A TR-mediated, supporting an environmental route of resistance selection. Our data supports the need to include veterinary isolates in resistance surveillance programs. Veterinarians should consider azole resistance as a reason for therapy failure when treating aspergillosis and consider resistance testing of relevant isolates.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
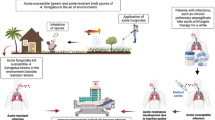
Aspergillus fumigatus is a saprophytic fungal pathogen that causes opportunistic infections in animals and man, and can be isolated from soil [1, 2]. The inhalation of A. fumigatus conidia is the principal route of infection leading to pulmonary aspergillosis [2]. Hypersensitivity diseases such as allergic bronchopulmonary aspergillosis and severe asthma with fungal sensitization are common in humans. Invasive infections are less frequently seen and immunocompromised hosts are most at risk to develop such an infection [1, 3]. In both domesticated and wild animals A. fumigatus infection can have a variety of clinical manifestations, such as infection of the air sacs in birds, sino-nasal aspergillosis in dogs and cats, guttural pouch infection in horses and mycotic pneumoniae in ruminants and cetaceans [1, 4,5,6]. Similar to humans, immunocompromised animals, including animals kept in immuno-compromising conditions, e.g. poor husbandry, are more at risk to develop aspergillosis [1].
The treatment of choice for aspergillosis in human and veterinary medicine are azole antifungal drugs [1, 7], which inhibit ergosterol biosynthesis by binding the cytochrome P450 enzyme sterol 14α-demethylase (Cyp51) resulting in inhibition of fungal growth by cell membrane disruption [8, 9]. From the year 1998 onwards azole resistance has emerged in A. fumigatus [10, 11] and occurs worldwide with varying prevalence [12]. The ARTEMIS global surveillance study showed 5.8% azole resistance in A. fumigatus isolates [13] and a mainly European international surveillance network reported a prevalence of 3.2% [14]. The prevalence of azole-resistant A. fumigatus in humans in the Netherlands is 0.8–14.7% with TR34/L98H being the dominant resistance mechanism present in 69–94% of resistant isolates [10, 15,16,17]. Differences in the study period, isolate selection and geographical location will account for some of the difference in reported prevalence. However, it is clear that there is an increasing trend in resistance frequency [10, 16, 17]. Resistance has been demonstrated in clinical isolates from both azole treated and azole naive human patients [15]. The use of azole fungicides in agriculture is considered to lead to resistance selection in environmental A. fumigatus isolates [18,19,20]. In the Netherlands as many as 64.3% of patients with azole-resistant Aspergillus spp. were azole naive, supporting a plausible role of environmental resistance selection in human resistant Aspergillus disease [15].
Several azole resistance mechanisms have been described in Aspergillus spp. and for A. fumigatus the most important one is decreased azole affinity through amino acid substitutions within the Cyp51A target protein. These alterations are the result of single nucleotide polymorphisms (SNPs) in the encoding cyp51A gene [8, 9]. The most common SNPs associated with patient treatment with azole drugs are G54, G138, M220 and G448. In case of environmental exposure SNPs are accompanied by tandem repeats (TR) of base pairs in the promotor region of the cyp51A gene, leading to increased gene expression, with predominantly TR34/L98H and TR46/Y121F/T289A being found [21].
Few studies have been conducted to evaluate azole resistance in A. fumigatus isolates from animal infections. Most studies focus on birds as A. fumigatus is an important pathogen of the avian respiratory tract [22, 23]. Triazole resistance frequencies of 0–21% in poultry [24,25,26,27] and 0–17.1% in other birds [28,29,30,31,32,33] have been reported. Studies on A. fumigatus isolates from dogs and cats are very limited and only one triazole resistant Australian canine isolate is described so far [34]. This isolate harbored the non-synonymous F46Y and E427K mutations in the cyp51A gene [34], which are associated with reduced triazole susceptibility [35, 36].
The aim of this retrospective surveillance study was to determine the azole resistance frequency, resistance phenotype and genotype in a collection of clinical A. fumigatus isolates from various animal species collected over a period of 5 years.
Materials and Methods
Isolates
Between January 2015 and October 2020 all A. fumigatus isolates cultured from clinical samples submitted to the Veterinary Microbiological Diagnostic Center (VMDC) of the Faculty of Veterinary Medicine, Utrecht University, The Netherlands were stored, yielding 142 isolates. The VMDC receives samples from various animal species submitted by Dutch veterinarians and by the Pathology Department and Dutch Wildlife Health Centre (both part of Faculty of Veterinary Medicine). Samples submitted by the latter two were taken post mortem (n = 15). Samples were cultured on Sabouraud dextrose agar and malt extract agar (Biotrading, Mijdrecht, the Netherlands). Species identification was based on macroscopic and microscopic morphologies. If A. fumigatus colonies were present one random colony was subcultured on Sabouraud dextrose agar and stored in the biobank. Only one isolate was included for each animal. Information regarding geographical origin, i.e. owner postal code or location where the animal was found for wildlife, age, sex, clinical signs, prior azole treatment, therapy and clinical response were obtained from the submission form when provided. For all samples with an azole-resistant isolate submitting veterinarians/pathologists were contacted to obtain missing information.
Antifungal Susceptibility Testing
The stored isolates were retrieved from the VMDC biobank and cultured on Sabouraud dextrose agar at 30 °C until sporulation occurred. According to the manufacturer’s instructions a 2 McFarland spore suspension was prepared in distillated water and a single drop (about 25 µl) was used to inoculate a commercial agar-based screening system to screen for azole resistance (VIPcheck™, https://www.vipcheck.nl). The system consists of a 4-well plate with 4 mg/l itraconazole (ITC), 2 mg/l voriconazole (VRC), 0.5 mg/l posaconazole (POS) and a growth control [37,38,39]. Plates were incubated at 37 °C for 48 h and checked for growth on day one and two of incubation. Antifungal susceptibility testing (AST) according to the EUCAST broth microdilution reference method (E.Def 9.3.2) was performed for isolates showing (minimal) growth on ≥ 1 azole-containing well. Minimal Inhibitory Concentrations (MICs) were determined for amphotericin B (AMB) and triazoles ITC, VRC, POS and isavuconazole (ISC). In addition MICs were determined for the imidazole miconazole (MCZ) for the aforementioned isolates and an additional random selection of four to five wildtype A. fumigatus veterinary isolates per year. In absence of veterinary breakpoints, EUCAST breakpoints for humans were used [40] for interpretation of MICs. Clinical breakpoints classify isolates as drug susceptible or resistant. For triazoles area of technical uncertainty (ATU) is defined for drug-bug combinations where there is uncertainty regarding the classification as susceptible or resistant. Clinical breakpoints were available for AMB (S ≤ /R > = 1/1), ITC (S ≤ /R > = 1/1, ATU = 2), VRC (S ≤ /R > = 1/1, ATU = 2), POS (S ≤ /R > = 0.125/0.25, ATU = 0.25) and ISC (S ≤ /R > = 1/2, ATU = 2). Since there are currently no breakpoints available for MCZ, the epidemiological cutoff (ECOFF) was determined based on the wildtype MIC distribution [41]. Isolates were defined as azole-resistant if they were resistant to one or more triazole drug.
Cyp51A Gene Sequencing
From isolates with an azole-resistant phenotype the cyp51A gene and promoter region were subsequently sequenced as described before [42].
Statistical Analysis
Statistical analysis was performed using R version 4.2.3 [43]. For each year the azole-resistant proportion of isolates was calculated. A pairwise comparison of proportions using the Fisher’s exact test was used to determine whether differences between proportions were statistically significant (p ≤ 0.05). Figures were created using postal codes and publicly available data using datawrapper (https://www.datawrapper.de).
Results
Isolates were cultured from different animal species and sites of infection (Table 1), originating from both wildlife (bird, hare and harbor porpoise) and kept animals (bird, cat, dairy cow, dog, gazelle, horse). The number of collected isolates per year, results of resistance screening and resistance frequencies are summarized in Table 2. Resistance screening identified 16 isolates showing growth on azole-containing wells. MIC-testing confirmed azole resistance in all 16 isolates, with a pan-azole-resistant phenotype in 15 (94%) isolates. One isolate (#12; cat, nose) showed multi-azole resistance for VRC, POS and ISC, but was susceptible to ITC. Azole resistance was cyp51A mediated in all sixteen isolates. TR34/L98H was the most prevalent resistance genotype with 81.3% (13/16); two isolates within this group harbored an additional T-67G substitution in the cyp51A promoter region. TR46/Y121F/T289A was found in 2 isolates (12.5%). In one isolate (#16; harbor porpoise, lung) a combination of C(-70)T/F46Y/C(intron7)T/C(intron66)T/M172V/E427K single-nucleotide polymorphisms in the cyp51A gene was found. The geographical origin of isolates and resistance genotype is shown in Fig. 1. Isolate distribution reflects population density [44] and no foci are seen.

Geographical origin of A. fumigatus isolates and their resistance genotype. Created by datawrapper (https://www.datawrapper.de)
MICs for MCZ ranged from 2 to 8 mg/l (median 4 mg/l) for wildtype isolates (n = 29), resulting in a MCZ ECOFF of ≤ 8 mg/l. MCZ showed no activity against TR34-mediated resistant A. fumigatus isolates (MIC > 16 mg/l), whereas the MCZ MIC of two isolates harboring TR46/Y121F/T289A was 8 mg/l. The isolate harboring C(-70)T/F46Y/C(intron7)T/C(intron66)T/M172V/E427K showed a MCZ MIC of 16 mg/l. Azole resistance frequencies were elevated in the years 2017 and 2020, without a trend over the years.
Resistant isolates were found in birds, cats, dogs, and harbor porpoise. Characteristics of the sample population for these animal species and prior azole treatment status are shown in Table 3. For prior azole treatment status only the information provided in the submission form was used. Additional information obtained for samples with an azole-resistant isolate by contact with the submitting veterinarian/pathologist was not taken into account here to avoid bias. When stratified for animal species overall resistance frequencies were 15% (2/13) for birds, 21% (6/28) for cats, 8% (6/75) for dogs and 33% (2/6) for harbor porpoises, without significant differences (p > 0.05) between these proportions. Both birds with an azole-resistant isolate were captive birds, with two wildlife and nine captive birds in the group with azole-susceptible isolates. Overall and within species there were no significant differences (p > 0.05) in proportions of animals treated with azole antifungal drugs when comparing the azole-susceptible to the azole-resistant group.
Azole-resistant isolates were cultured from the respiratory tract (n = 9) and the external ear canal (n = 7). Information on prior azole exposure, treatment and clinical outcome was acquired for fourteen of sixteen animals harboring an azole-resistant A. fumigatus isolate (Table 4); four animals (28.6%) received azole treatment within 1 week to 4 months prior to sampling and ten (71.4%) were azole naive. Prior treatment consisted of one to multiple courses with a duration of one to several weeks with MCZ or ITC. Six of the fourteen animals did not receive subsequent antifungal therapy, one was treated with ketoconazole, two with itraconazole, in one animal therapy information was unavailable and in four animals not applicable as samples were taken at pathological examination. Information on clinical outcome reported recovery for three of fourteen animals, four animals had died prior to sampling and no follow up information was available for seven animals. Death was attributable to aspergillosis in two of the four animals sampled at pathological examination (#6, #9) and for two animals this was unclear from the information provided by the pathologist.
Discussion
In this study of A. fumigatus isolates from animals in the Netherlands an overall azole resistance prevalence of 11.3% was found. A Dutch national surveillance program is in place to monitor the prevalence of azole resistance in A. fumigatus in the human population. In this surveillance program five University Medical Centers and five teaching hospitals screen clinical A. fumigatus isolates using VIPcheck™, which corresponds to the methods used in our study. Human surveillance shows overall resistance rates of 10.7%, 12.9%, 14.7%, 10.5%, 9.1% and 8.2% for the years 2015–2020 respectively [46,47,48,49,50,51]. In addition to this surveillance program several studies have investigated the prevalence of azole resistance in clinical A. fumigatus isolates from the Dutch human population. One study from a tertiary care hospital found an overall patient azole-resistance prevalence of 5.3% in 2,051 patients in 1994–2016. Patient resistance frequency increased from 0% in 1997 to 9.5% in 2016 [17]. Two studies on isolates from multiple University Medical Centers found an overall patient azole-resistance prevalence of 5.3% in 1,192 patients in 2007–2009 [15] and 11% in 4,496 patients in 2013–2018, with annual resistance frequencies increasing from 7.6 to 14.7% [16]. Our study period and prevalence of 11.3% is in line with these findings from the Dutch human population. However, in our study no screening method was used at initial culture and only one random A. fumigatus isolate was stored. As mixed (azole-susceptible and azole-resistant) genotypes may be present in clinical cultures, azole-resistant colonies may have been missed [52] and true prevalence might be higher. In our study prevalence of resistance varied widely between the years and no trend was visible, which may be due to the limited number of A. fumigatus isolates per year.
Few studies have been conducted to evaluate azole resistance in A. fumigatus isolates from animals. We found a resistance frequency of 15% (2/13) in wild life and captive birds. Previous studies found azole-resistance in A. fumigatus isolates from four of 54 birds (7.4%) from Belgium and the Netherlands [29] and only azole susceptible isolates from nine birds from Japan [28]. It is difficult to interpret the outcomes of these studies, because sample background information is missing. A recent study reported azole resistance in 17.1% of clinical (6/35) and 11.8% (8/68) of environmental A. fumigatus isolates from Humboldt Penguins and other birds and their surroundings from 2017 to 2022 in a Belgian Zoo [32]. In this study sequencing of the cyp51A gene revealed the TR34/L98H mutation in all environmental isolates and TR34/L98H (2/6), F46Y/M172V/E427K (1/6) and the absence of known resistance mutations (3/6) in clinical isolates. Genotyping showed an interesting cluster of azole-resistant isolates from the same time point encompassing environmental strains and one clinical strain from a Humboldt pinguin indicating environment to bird transmission. Resistance frequency and mutations are in line with the results of our study and support the environment to animal transmission. Another study investigated 159 cases of aspergillosis in captive birds from Germany in 2015–2018 and found one (0.6%) azole-resistant A. fumigatus isolate [30]. In this study samples were collected from all avian aspergillosis cases seen at the University Clinic during the study period. In our study results are possibly biased as it is likely that samples were more often submitted from animals with severe disease and/or not responding to antifungal therapy, thus leading to a higher resistance frequency. Such differences in sample selection can lead to variation in azole-resistance prevalence.
Little is known of the prevalence of azole-resistant A. fumigatus in companion animals. In the Netherlands triazole resistance was not observed in 27 isolates from eight dogs with sino-nasal aspergillosis in 2010–2012 [53]. A study on 50 isolates originating from Australia, United States and Belgium collected between 1988 and 2014 from 46 dogs and four cats with sino-nasal aspergillosis showed one triazole resistant Australian canine isolate from 1992 [34]. Resistance frequencies for dogs (8%) and cats (21%) are much higher in our study. This might be due to a higher number of isolates, more recent study period and the possible submission bias in our study as suggested above. However, both studies might be prone to a similar bias as isolates were retrieved from a culture collection of a clinical research group on infectious diseases [34] or included dogs from a veterinary referral hospital only [53].
Studies investigating aspergillosis in harbor porpoises are scarce [5, 6] and none have investigated azole-resistance in A. fumigatus isolates. In our study two of the six A. fumigatus isolates from stranded, free-ranging harbor porpoises were azole-resistant. Both animals were deceased at stranding and suffered from fungal pneumonia, which has been described in harbor porpoise [5, 6].
Overall VIPcheck™ resistance screening accurately predicted azole-resistance in isolates showing (minimal) growth on ≥ 1 azole-containing wells, so specificity for this screening method is high. Isolates that only grew in the control well were not further tested, therefore sensitivity cannot be evaluated with our data. It is unlikely that many azole resistant isolates were missed, as studies evaluating the VIPcheck™ system have reported an excellent sensitivity and specificity of 98–99% and 93–99%, respectively [37, 54]. However, isolate number 15 underlines the importance of including minimal growth as a positive result, as it showed only minimal growth for ITC and VRC, while MIC-testing showed resistance to ITC, VRC and POS. This is in line with previous findings [37, 54] and underscores the necessity of subsequent MIC-testing for accurate antifungal susceptibility profiling.
Although triazole resistance has been broadly investigated in A. fumigatus, there is a paucity of in vitro susceptibility information on imidazole drugs. We included MCZ in our current study because imidazole drugs, e.g. MCZ, enilconazole, clotrimazole and ketoconazole, are licensed for use in animals [55] and some imidazole drugs are used for treatment of sino-nasal aspergillosis in dogs and cats [56, 57]. In human medicine MCZ is not considered an appropriate drug for treatment of Aspergillus diseases, due to various reasons including relatively high MIC-values [58]. We indeed found relatively high MICs in our wildtype A. fumigatus isolates with an ECOFF of ≤ 8 mg/l. This MIC distribution is very similar to that observed in an early study investigating the in vitro activity of MCZ [59]. MCZ showed no in vitro activity against TR34/L98H, whereas the MICs of two TR46/Y121F/T289A were at the ECOFF concentration. Studies with an imidazole fungicide, imazalil, indicated that the presence of a F495I SNP was associated with imidazole resistance [60]. Indeed MCZ showed no activity against two clinical A. fumigatus isolates harboring TR34/L98H/S297T/F495I [61]. However, our data suggest that MCZ also shows no activity against TR34/L98H sensu stricto isolates. Likely various factors including molecule structure and chemical characteristics and resistance mutation variations determine the resistance phenotype, underscoring the complexity of resistance in A. fumigatus.
The distribution of resistance genotypes was similar to that observed in the national resistance surveillance program, with dominance of TR34/L98H. In humans, in host resistance selection during triazole therapy is characterized by single resistance mutations mainly in the cyp51A gene. Such resistance mechanisms were not observed in veterinary isolates, possibly pointing towards a low risk of resistance selection during antifungal therapy. Prolonged azole therapy is less common in veterinary medicine and (to our knowledge) was not administered to animals in our study. However, often no information on previous azole therapy was provided and only actively obtained for samples with an azole-resistant isolate. It is therefore difficult to accurately assess the effect of previous azole exposure on our collection of isolates. In our study 71.4% of animals with an azole-resistant A. fumigatus isolate and known azole exposure status were azole naive, which is in line with the 64.3% previously found in the Dutch human population (2007–2009) [15] and indicates an environmental route of resistance development.
Resistance mutations (T67G/)TR34/L98H and TR46/Y121F/T289A are associated with environmental resistance selection [16, 21] and accounted for all but one (94%) mutations in our collection from animals. The mutations in the C(-70)T/F46Y/C(intron7)T/C(intron66)T/M172V/E427K isolate recovered from a free ranging harbor porpoise, have been described for both human clinical and environmental strains [35, 62]. This suggests that, similar to humans, the environmental resistance route is dominant in veterinary medicine and that veterinary azole use is an unlikely hotspot for resistance selection [19, 20]. Recent genomic studies confirm environment-to-patient transmission of azole-resistant A. fumigatus, with closely-related genomes of environmental and human TR34/L98H isolates [63]. Studies investigating the genomic epidemiology of resistance in animal isolates are lacking but will be required to confirm environment-to-animal transmission.
Due to the limited number of animals with an azole-resistant A. fumigatus isolate and paucity of clinical information it is impossible to make general statements regarding clinical implications of azole resistance for the animals in our study. However, the results of our study have implications for veterinary microbiological diagnostic laboratories and veterinarians. An expert panel made recommendations for resistance management depending on the resistance prevalence [64]. In an area with a local environmental resistance rate of < 5% screening for azole resistance was recommended for patients with insufficient clinical response to azole therapy, 5% was recommended as threshold for routine resistance screening of A. fumigatus isolates and 10% for reconsidering the use of azole monotherapy as primary treatment for patients with aspergillosis [21, 64]. In our study an overall azole resistance prevalence of 11.3% was found in clinical A. fumigatus isolates from animal infections. Therefore veterinarians should be aware of azole therapy failure due to resistance and diagnostic laboratories should consider offering screening of A. fumigatus isolates for azole resistance. Methods to be considered are agar-based resistance screening followed by MIC-testing if growth on azole-containing wells is observed. Where phenotypic screening takes 24–48 h, PCR directly on clinical samples can potentially detect cyp51A mediated resistance much quicker, provided sensitivity and coverage of azole resistance markers of the assay are sufficient, but non-cyp51A mediated resistance will not be detected [65]. Finally, it would be preferable to include imidazole drugs in in vitro susceptibility tests as they are used for treatment. However, clinical breakpoints will need to be established to interpret MIC values. Reconsidering the use of azole therapy as primary treatment for animals is impaired by lack of alternatives. In the European Union only imidazole and triazole antifungal drugs are registered for therapy in animals [55]. Fortunately, off-label AMB treatment has been described [1] and offers an alternative treatment option in animals with azole-resistant Aspergillus diseases.
Our study shows that in the Netherlands azole resistance is present in clinical A. fumigatus isolates from animals and at similar frequency as found in humans. We therefore advise veterinarians to be aware of azole resistance as cause of aspergillosis treatment failure and consider resistance screening of relevant isolates. The predominantly cyp51A TR-mediated resistance mutations support an environmental route of resistance selection, and warrants further studies from a One Health perspective.
References
Seyedmousavi S, et al. Aspergillus and aspergilloses in wild and domestic animals: a global health concern with parallels to human disease. Med Mycol. 2015;53(8):765–97. https://doi.org/10.1093/mmy/myv067.
Kwon-Chung KJ, Sugui JA. Aspergillus fumigatus—what makes the species a ubiquitous human fungal pathogen? PLoS Pathog. 2013;9(12): e1003743. https://doi.org/10.1371/JOURNAL.PPAT.1003743.
Buil JB, Meijer EFJ, Denning DW, Verweij PE, Meis JF. Burden of serious fungal infections in the Netherlands. Mycoses. 2020;63(6):625–31. https://doi.org/10.1111/MYC.13089.
Tell LA. Aspergillosis in mammals and birds: Impact on veterinary medicine. Med Mycol. 2005. https://doi.org/10.1080/13693780400020089.
Kapetanou A, et al. Mycotic infections in free-ranging harbor porpoises (Phocoena phocoena). Front Mar Sci. 2020. https://doi.org/10.3389/FMARS.2020.00344/FULL.
van Elk CE, et al. Clinical, pathological, and laboratory diagnoses of diseases of harbour porpoises (Phocoena phocoena), live stranded on the Dutch and adjacent coasts from 2003 to 2016. Vet Res. 2019. https://doi.org/10.1186/S13567-019-0706-3.
Patterson TF, et al. Practice guidelines for the diagnosis and management of Aspergillosis: 2016 update by the infectious diseases society of America. Clin Infect Dis. 2016;63(4):e1–60. https://doi.org/10.1093/CID/CIW326.
Pérez-Cantero A, López-Fernández L, Guarro J, Capilla J. Azole resistance mechanisms in Aspergillus: update and recent advances. Int J Antimicrob Agents. 2020. https://doi.org/10.1016/j.ijantimicag.2019.09.011.
Parker JE, Warrilow AGS, Price CL, Mullins JGL, Kelly DE, Kelly SL. Resistance to antifungals that target CYP51. J Chem Biol. 2014;7(4):143–61. https://doi.org/10.1007/S12154-014-0121-1.
Snelders E, et al. Emergence of azole resistance in Aspergillus fumigatus and spread of a single resistance mechanism. PLoS Med. 2008;5(11):1629–37. https://doi.org/10.1371/JOURNAL.PMED.0050219.
Verweij PE, te Dorsthorst DTA, Rijs AJMM, de Vries-Hospers HG, Meis JFGM. Nationwide survey of in vitro activities of itraconazole and voriconazole against clinical Aspergillus fumigatus isolates cultured between 1945 and 1998. J Clin Microbiol. 2002;40(7):2648–50. https://doi.org/10.1128/JCM.40.7.2648-2650.2002.
Sharpe AR, Lagrou K, Meis JF, Chowdhary A, Lockhart SR, Verweij PE. Triazole resistance surveillance in Aspergillus fumigatus. Med Mycol. 2018;56(suppl_1):S83–92. https://doi.org/10.1093/MMY/MYX144.
Lockhart SR, Frade JP, Etienne KA, Pfaller MA, Diekema DJ, Balajee SA. Azole resistance in Aspergillus fumigatus isolates from the ARTEMIS global surveillance study is primarily due to the TR/L98H mutation in the cyp51A gene. Antimicrob Agents Chemother. 2011;55(9):4465–8. https://doi.org/10.1128/AAC.00185-11.
van der Linden JWM, et al. Prospective multicenter international surveillance of azole resistance in Aspergillus fumigatus. Emerg Infect Dis. 2015;21(6):1041–4. https://doi.org/10.3201/EID2106.140717.
van der Linden JWM, et al. Clinical implications of azole resistance in Aspergillus fumigatus, The Netherlands, 2007–2009. Emerg Infect Dis. 2011;17(10):1846–54. https://doi.org/10.3201/EID1710.110226.
Lestrade PPA, et al. Paradoxal trends in azole-resistant Aspergillus fumigatus in a national multicenter surveillance program, the Netherlands, 2013–2018. Emerg Infect Dis. 2020;26(7):1447–55. https://doi.org/10.3201/EID2607.200088.
Buil JB, Snelders E, Denardi LB, Melchers WJG, Verweij PE. Trends in azole resistance in Aspergillus fumigatus, the Netherlands, 1994–2016. Emerg Infect Dis. 2019;25(1):176–8. https://doi.org/10.3201/EID2501.171925.
Snelders E, Huis In’t Veld RAG, Rijs AJMM, Kema GHJ, Melchers WJG, Verweij PE. Possible environmental origin of resistance of Aspergillus fumigatus to medical triazoles. Appl Environ Microbiol. 2009;75(12):4053–7. https://doi.org/10.1128/AEM.00231-09.
Schoustra SE, et al. Environmental hotspots for azole resistance selection of Aspergillus fumigatus, the Netherlands. Emerg Infect Dis. 2019;25(7):1347–53. https://doi.org/10.3201/EID2507.181625.
Doughty KJ, Sierotzki H, Semar M, Goertz A. Selection and amplification of fungicide resistance in Aspergillus fumigatus in relation to DMI fungicide use in agronomic settings: hotspots versus coldspots. Microorganisms. 2021. https://doi.org/10.3390/microorganisms9122439.
Wiederhold NP, Verweij PE Aspergillus fumigatus and pan-azole resistance: who should be concerned?. Current opinion in infectious diseases. 2020;33(4), NLM (Medline); 290–297. https://doi.org/10.1097/QCO.0000000000000662.
Melo AM, Stevens DA, Tell LA, Veríssimo C, Sabino R, Xavier MO. Aspergillosis, avian species and the one health perspective: the possible importance of birds in azole resistance. Microorganisms. 2020;8(12):1–22. https://doi.org/10.3390/MICROORGANISMS8122037.
Arné P, Risco-Castillo V, Jouvion G, Le Barzic C, Guillot J. Aspergillosis in wild birds. J Fungi (Basel). 2021. https://doi.org/10.3390/jof7030241.
Nawrot U, Wieliczko A, Włodarczyk K, Kurzyk E, Brillowska-Dąbrowska A. Low frequency of itraconazole resistance found among Aspergillus fumigatus originating from poultry farms in Southwest Poland. J Mycol Med. 2019;29(1):24–7. https://doi.org/10.1016/J.MYCMED.2018.12.005.
Wang DY, et al. Mutations in the Cyp51A gene and susceptibility to itraconazole in Aspergillus fumigatus isolated from avian farms in France and China. Poult Sci. 2014;93(1):12–5. https://doi.org/10.3382/PS.2013-03541.
Ziółkowska G, Tokarzewski S, Nowakiewicz A. Drug resistance of Aspergillus fumigatus strains isolated from flocks of domestic geese in Poland. Poult Sci. 2014;93(5):1106–12. https://doi.org/10.3382/PS.2013-03702.
Spanamberg A, et al. Antifungal susceptibility profile of Aspergillus fumigatus isolates from avian lungs. Pesqui Vet Bras. 2020;40(2):102–6. https://doi.org/10.1590/1678-5150-PVB-6297.
Okabayashi K, et al. Antifungal activity of itraconazole and voriconazole against clinical isolates obtained from animals with mycoses. Jpn J Med Mycol. 2009;50(2):91–4. https://doi.org/10.3314/jjmm.50.091.
Beernaert LA, et al. Avian Aspergillus fumigatus strains resistant to both itraconazole and voriconazole. Antimicrob Agents Chemother. 2009;53(5):2199–201. https://doi.org/10.1128/AAC.01492-08.
Barber AE, Scheufen S, Walther G, Kurzai O, Schmidt V. Low rate of azole resistance in cases of avian aspergillosis in Germany. Med Mycol. 2020;58(8):1187–90. https://doi.org/10.1093/MMY/MYAA045.
Sabino R, et al. Molecular identification of clinical and environmental avian Aspergillus isolates. Arch Microbiol. 2019;201(2):253–7. https://doi.org/10.1007/S00203-019-01618-Y/FIGURES/1.
Debergh H, et al. Pulmonary Aspergillosis in Humboldt Penguins-susceptibility patterns and molecular epidemiology of clinical and environmental Aspergillus fumigatus isolates from a Belgian Zoo, 2017–2022. Antibiotics (Basel). 2023. https://doi.org/10.3390/antibiotics12030584.
Melo AM, et al. Aspergillosis in free-ranging Magellanic penguins. Med Mycol. 2023. https://doi.org/10.1093/mmy/myad008.
Talbot JJ, Kidd SE, Martin P, Beatty JA, Barrs VR. Azole resistance in canine and feline isolates of Aspergillus fumigatus. Comp Immunol Microbiol Infect Dis. 2015;42:37–41. https://doi.org/10.1016/j.cimid.2015.08.002.
Garcia-Rubio R, et al. Insight into the significance of Aspergillus fumigatus cyp51A polymorphisms. Antimicrob Agents Chemother. 2018. https://doi.org/10.1128/AAC.00241-18.
Chowdhary A, Sharma C, Meis JF. Azole-resistant aspergillosis: epidemiology, molecular mechanisms, and treatment. J Infect Dis. 2017;216(suppl_3):S436–44. https://doi.org/10.1093/INFDIS/JIX210.
Buil JB, et al. Single-center evaluation of an agar-based screening for azole resistance in Aspergillus fumigatus by using VIPcheck. Antimicrob Agents Chemother. 2017;61(12):e01250-e1317. https://doi.org/10.1128/AAC.01250-17.
Arendrup MC, Verweij PE, Mouton JW, Lagrou K, Meletiadis J. Multicentre validation of 4-well azole agar plates as a screening method for detection of clinically relevant azole-resistant Aspergillus fumigatus. J Antimicrob Chemother. 2017;72(12):3325–33. https://doi.org/10.1093/JAC/DKX319.
Guinea J, et al. How to: EUCAST recommendations on the screening procedure E.Def 10.1 for the detection of azole resistance in Aspergillus fumigatus isolates using four-well azole-containing agar plates. Clin Microbiol Infect. 2019;25(6):681–7. https://doi.org/10.1016/j.cmi.2018.09.008.
European Committee on Antimicrobial Susceptibility Testing. Breakpoint tables for interpretation of MICs for antifungal agents. 2020. Accessed 21 Jan 2022 [Online]. Available: https://www.eucast.org/astoffungi/clinicalbreakpointsforantifungals/.
Kahlmeter G, et al. European harmonization of MIC breakpoints for antimicrobial susceptibility testing of bacteria. J Antimicrob Chemother. 2003;52(2):145–8. https://doi.org/10.1093/JAC/DKG312.
Mellado E, Diaz-Guerra TM, Cuenca-Estrella M, Rodriguez-Tudela JL. Identification of two different 14-alpha sterol demethylase-related genes (cyp51A and cyp51B) in Aspergillus fumigatus and other Aspergillus species. J Clin Microbiol. 2001;39(7):2431–8. https://doi.org/10.1128/JCM.39.7.2431-2438.2001.
R Core Team. R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. 2023. [Online]. Available: https://www.r-project.org/.
CBS, PBL, RIVM, and WUR. Bevolkingsgroei, 2015–2020 (indicator 2102, versie 07, 20 oktober 2020) | Compendium voor de Leefomgeving. Centraal Bureau voor de Statistiek (CBS), Den Haag; PBL Planbureau voor de Leefomgeving, Den Haag; RIVM Rijksinstituut voor Volksgezondheid en Milieu, Bilthoven; en Wageningen University and Research, Wageningen. Accessed 15 Dec 2022. [Online]. Available: https://www.clo.nl/indicatoren/nl2102-bevolkingsgroei-nederland-.
Arendrup MC, et al. How to interpret MICs of antifungal compounds according to the revised clinical breakpoints v. 10.0 European committee on antimicrobial susceptibility testing (EUCAST). Clin Microbiol Infect. 2020;26(11):1464–72. https://doi.org/10.1016/J.CMI.2020.06.007.
Dutch Foundation of the Working Party on Antibiotic Policy and National Institute for Public Health and the Environment of the Netherlands. NethMap 2021 Consumption of antimicrobial agents and antimicrobial resistance among medically important bacteria in the Netherlands. 2021. Accessed 07 Feb 2023 [Online]. Available: https://swab.nl/nl/nethmap.
Dutch Foundation of the Working Party on Antibiotic Policy and National Institute for Public Health and the Environment of the Netherlands. NethMap 2020 Consumption of antimicrobial agents and antimicrobial resistance among medically important bacteria in the Netherlands. 2020. Accessed 07 Feb 2023 [Online]. Available: https://swab.nl/nl/nethmap.
Dutch Foundation of the Working Party on Antibiotic Policy and National Institute for Public Health and the Environment of the Netherlands. NethMap 2019 consumption of antimicrobial agents and antimicrobial resistance among medically important bacteria in the Netherlands. 2019. Accessed 07 Feb 2023 [Online]. Available: https://swab.nl/nl/nethmap.
Dutch Foundation of the Working Party on Antibiotic Policy and National Institute for Public Health and the Environment of the Netherlands. NethMap 2018 consumption of antimicrobial agents and antimicrobial resistance among medically important bacteria in the Netherlands. 2018. Accessed 07 Feb 2023. [Online]. Available: https://swab.nl/nl/nethmap.
Dutch Foundation of the Working Party on Antibiotic Policy and National Institute for Public Health and the Environment of the Netherlands. NethMap 2017 consumption of antimicrobial agents and antimicrobial resistance among medically important bacteria in the Netherlands. 2017. Accessed 07 Feb 2023 [Online]. Available: https://swab.nl/nl/nethmap.
Dutch Foundation of the Working Party on Antibiotic Policy and National Institute for Public Health and the Environment of the Netherlands. NethMap 2016 consumption of antimicrobial agents and antimicrobial resistance among medically important bacteria in The Netherlands in 2015. 2016. Accessed 07 Feb 2023 [Online]. Available: https://swab.nl/nl/nethmap.
van Leer-Buter C, Takes RP, Hebeda KM, Melchers WJ, Verweij PE. Aspergillosis–and a misleading sensitivity result. Lancet. 2007;370(9581):102. https://doi.org/10.1016/S0140-6736(07)61055-1.
Valdes ID, et al. Comparative genotyping and phenotyping of Aspergillus fumigatus isolates from humans, dogs and the environment. BMC Microbiol. 2018. https://doi.org/10.1186/s12866-018-1244-2.
Arendrup MC, Verweij PE, Mouton JW, Lagrou K, Meletiadis J. Multicentre validation of 4-well azole agar plates as a screening method for detection of clinically relevant azole-resistant Aspergillus fumigatus. J Antimicrob Chemother. 2018;73(8):2274. https://doi.org/10.1093/JAC/DKY218.
European Medicines Agency. Union product database. Accessed 02 Dec 2022. [Online]. Available https://medicines.health.europa.eu/veterinary/nl.
Sharman MJ, Mansfield CS. Sinonasal aspergillosis in dogs: a review. J Small Anim Pract. 2012;53(8):434–44. https://doi.org/10.1111/j.1748-5827.2012.01245.x.
Hartmann K, et al. Aspergillosis in cats: ABCD guidelines on prevention and management. J Feline Med Surg. 2013;15(7):605–10. https://doi.org/10.1177/1098612X13489223.
Wingard JR, Vaughan WP, Braine HG, Merz WG, Saral R. Prevention of fungal sepsis in patients with prolonged neutropenia: a randomized, double-blind, placebo-controlled trial of intravenous miconazole. Am J Med. 1987;83(6):1103–10. https://doi.org/10.1016/0002-9343(87)90949-1.
Dixon DM, Wagner GE, Shadomy S, Shadomy HJ. In vitro comparison of the antifungal activities of R34,000, miconazole and amphotericin B. Chemotherapy. 1978;24(6):364–7. https://doi.org/10.1159/000237809.
Chen Y, et al. Elevated MIC values of imidazole drugs against Aspergillus fumigatus Isolates with TR34/L98H/S297T/F495I Mutation. Antimicrob Agents Chemother. 2018. https://doi.org/10.1128/AAC.01549-17.
Pontes L, et al. Aspergillus fumigatus clinical isolates carrying CYP51A with TR34/L98H/S297T/F495I substitutions detected after four-year retrospective azole resistance screening in Brazil. Antimicrob Agents Chemother. 2020. https://doi.org/10.1128/AAC.02059-19.
Fraaije B, Atkins S, Hanley S, Macdonald A, Lucas J. The multi-fungicide resistance status of Aspergillus fumigatus populations in arable soils and the wider European environment. Front Microbiol. 2020;11: 599233. https://doi.org/10.3389/FMICB.2020.599233/FULL.
Rhodes J, et al. Population genomics confirms acquisition of drug-resistant Aspergillus fumigatus infection by humans from the environment. Nat Microbiol. 2022;7(5):663–74. https://doi.org/10.1038/S41564-022-01091-2.
Verweij PE, et al. International expert opinion on the management of infection caused by azole-resistant Aspergillus fumigatus. Drug Resist Updat. 2015;21–22:30–40. https://doi.org/10.1016/J.DRUP.2015.08.001.
Buil JB, Zoll J, Verweij PE, Melchers WJG. Molecular detection of azole-resistant Aspergillus fumigatus in clinical samples. Front Microbiol. 2018. https://doi.org/10.3389/FMICB.2018.00515.
Acknowledgements
The authors are grateful to Dr. J. Houbraken for his assistance in confirming the identification of some of the A. fumigatus isolates.
Funding
The authors declare that no funds, grants, or other support were received during the preparation of this manuscript.
Author information
Authors and Affiliations
Contributions
Paul Verweij, Jaap Wagenaar, Els Broens, Jochem Buil and Marloes van Dijk contributed to the study conception and design. Material preparation, data collection and analysis were performed by Marloes van Dijk, Marlou Tehupeiory-Kooreman and Marian Broekhuizen. The first draft of the manuscript was written by Marloes van Dijk and all authors commented on previous versions of the manuscript. All authors read and approved the final manuscript.
Corresponding author
Additional information
Handling Editor: Ferry Hage.
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
van Dijk, M.A.M., Buil, J.B., Tehupeiory-Kooreman, M. et al. Azole Resistance in Veterinary Clinical Aspergillus fumigatus Isolates in the Netherlands. Mycopathologia 189, 50 (2024). https://doi.org/10.1007/s11046-024-00850-5
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s11046-024-00850-5