
Abstract
Background
Antimicrobial peptides (AMPs) are a diverse class of molecules that represent a vital part of innate immunity. AMPs are evolutionarily conserved molecules that exhibit structural and functional diversity. They provide a possible solution to the antibiotic-resistance crisis.
Main text
These small cationic peptides can target bacteria, fungi, and viruses, as well as cancer cells. Their unique action mechanisms, rare antibiotic-resistant variants, broad-spectrum activity, low toxicity, and high specificity encourage pharmaceutical industries to conduct clinical trials to develop them as therapeutic drugs. The rapid development of computer-assisted strategies accelerated the identification of AMPs. The Antimicrobial Peptide Database (APD) so far contains 3324 AMPs from different sources. In addition to their applications in different fields, some AMPs demonstrated the potential to combat COVID-19, and hinder viral infectivity in diverse ways.
Conclusions
This review provides a brief history of AMPs and their features, including classification, evolution, sources and mechanisms of action, biosynthesis pathway, and identification techniques. Furthermore, their different applications, challenges to clinical applications, and their potential use against COVID-19 are presented.
Similar content being viewed by others

Background
Antimicrobial peptides (AMPs) are a diverse class of molecules found in nature, and expressed in all organisms, including mammals [1]. They are indispensable components of the innate immunity, and have a broad spectrum of activity against bacteria, fungi, yeasts, viruses, as well as cancer cells [2]. Generally, AMPs are relatively short in length [10–100 amino acid (aa) residues], cationic (positively charged), and amphiphilic (hydrophobic and hydrophilic) [3]. AMPs are encoded in the genome as prepropeptides, where the signal peptide occurs at the N-terminus, and the antimicrobial peptide domain occurs at the C-terminus [4]. By post-translational modification, the mature (active) peptide is generated through proteolytic cleavage [5].
Misuse and long-term use of conventional antibiotics have led to the growing problem of bacterial drug resistance [6], which stimulated interest in the development of AMPs as the next generation anti-infectives. The key advantages of AMPs over traditional antibiotics are the immunomodulatory activities and bactericidal action of most AMPs [5], making them significant compounds in the development of novel therapeutics [7]. AMPs have several biological activities, such as wound healing [8], antiviral [9], anti-biofilm [1], anti-tumor activities [10]. Increasing evidence shows that AMPs can inhibit SARS-CoV-2 (COVID-19), paving the way for their prospective use as therapeutic drugs [11]. Apart from their prospective therapeutic applications, AMPs have different applications in various fields, such as the food industry [12], aquaculture [13], agriculture [14], and animal husbandry [15].
This review provides insight into the history, sources, classes, evolution, mode of action, biosynthesis pathway, prediction, and the different applications of AMPs. In addition, the research progress on AMPs against COVID-19 is discussed.
History of antimicrobial peptides
In 1922, Alexander Fleming succeeded in identifying the first AMP named Lysozyme. However, the discovery of lysozyme was overshadowed in 1928, when Fleming discovered penicillin [16]. Florey, Chain, and Fleming brought the potential of penicillin in medical use to fruition, and shared the 1945 Nobel Prize in Medicine. In the 1940s and at the beginning of the golden age of antibiotics, the interest in the therapeutic prospective of natural AMPs was lost. In the 1960s, the interest in AMPs as host defense molecules was awakened in the 1960s due to the rise of multidrug-resistant microbial pathogens [6]. In 1981, α-helical AMPs named cecropins -from Cecropia silk moth- was characterized, followed by magainin from African clawed frog Xenopus laevis, in 1987 [17, 18]. In the 1990s, the field of antimicrobial peptides expanded rapidly, reporting over 300 peptides [19]. AMPs have since been broadly identified and characterized in approximately all organisms. Currently, more than 3324 AMPs have been deposited in the Antimicrobial Peptide Database [20].
Classification of antimicrobial peptides
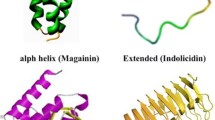
Based on their secondary structure, AMPs are divided into four main classes: α-helix, β-Sheet, α/β, and extended/random-coil peptides (Fig. 1). α-helix and β-Sheet are the most abundant classes in nature [3]. α-helical peptides contain plenty of helix-stabilizing residues, such as lysine, alanine, and leucine, and lack cysteine residues. The structure of α-helical peptides is lost in solution, but acquires an amphipathic helical structure in contact with a biological membrane. Frog magainins belong to this class [21]. On the other hand, β-Sheet peptides contain two to ten cysteine residues that form one to five interchain disulfide bonds. This bonding interaction allows peptides to adopt the β-sheet conformation. Unlike α-helix peptides, the structure of β-sheet peptides remains stable in aqueous solution [22]. Mammalian defensins belong to β-sheet peptides. Protegrins represent the mixed α/β class, as their secondary structure contains both α-helix and β-Sheet. The fourth class, namely extended/random-coil peptides, comprises a small part of the AMPs, and contains a high content of certain amino acids, such as arginine, tryptophan, proline, or histidine [19]. They also have greatly variable secondary structures [23]. Members of this class include the arginine and proline-rich porcine, histidine-rich human histatins, and tryptophan-rich bovine indolicidin [24].

Different classes of AMPs
Sources of antimicrobial peptides
The Antimicrobial Peptide Database (APD) comprises 3324 peptides from all kingdoms: archaea (5), animals (2446), bacteria (391), protists (8), plants (364), and fungi (22), besides some synthetic peptides [20].
Bacteria
Some AMPs are produced by bacteria to maintain the control of the population, and combat other microorganisms competing for space and nutrients in their environment. AMPs produced by bacteria are called bacteriocins. Colicins and Microcins are the two classes of bacteriocins produced by Gram-negative bacteria, while lantibiotics are produced by Gram-positive bacteria [25].
Plants
Twelve families of AMPs are produced from plants. For example, thionins are synthesized in seeds, flowers, and leaves, whereas defensins are synthesized in seeds [25].
Insects
Insects produce a larger range of AMPs than any other taxonomic group [26]. AMPs are synthesized in the insect fat bodies, blood cells, hemolymph, and salivary glands. Cecropins, defensins, drasomycin, and thanatin are examples of insect AMPs [27].
Marine invertebrates
Marine invertebrates lack acquired immune response; hence, AMPs are vital for their survival. Several AMPs are detected in marine invertebrates, such as discodermins, halicylindramides, aurelins, and hedistins [28].
Amphibians
The glands of amphibian skin are rich sources of antimicrobial peptides. The skin of frogs and toads produces numerous AMPs, such as citropins, bombinins, plasticins, and magainins [29].
Birds
For avian species, three major classes of AMPs are identified: cathelicidins, β-defensins, and liver-expressed antimicrobial peptide-2 [30].
Fish
In fish, AMPs are defensive weapons against devastating diseases [30]. Piscidin, hepcidin, dicentracin, and NK lysine are examples of fish AMPs [31, 32].
Mammals
Cathelicidins and defensins are the two principal AMPs families In mammalian species [33].
Evolution and diversification of antimicrobial peptides
Hosts and pathogens live in a never-ending struggle, with pathogens constantly escaping the host’s immune response, while the immune system of the host advances its barriers against pathogens. This arms race results in the rapid evolution of immune system-related genes [34]. On the molecular level, this combat leads to the evolutionary response, represented in mutations. As a result, natural selection may act vigorously on immune-related genes such as AMPs, which subsequently show high levels of genetic diversity [35]. Over the past years, some families of AMPs from all species have showed a high and variable level of sequence diversity. Increasing evidence shows that all organisms present a particular armory of various AMP families. This variability can play a critical role in shaping the pathogenicity of microbes [35].
On the genomic level, positive and negative selection forces can figure evolution. In the positive selection, new alleles present a great strength to an individual, and exacerbate by time, resulting in the substitution of ancestral allele in the population. Contrariwise, in the negative selection, new mutations decrease the strength, and head towards disappearing from the population [36]. The most recognized AMPs diversification mechanisms are represented in gene copy number variation, gene duplication, recombination, and allelic polymorphisms. The degree to which selection forces genetic variation within AMP genes depends on several factors, such as the density, diversity, and virulence of pathogens, in addition to the action timescale of selection, effective population size, and environmental variables [37].
The different families of defensins are the best examples of evolving AMPs, since they are age-old conserved peptides that exhibit diversification patterns, indicating a common evolutionary origin [38]. It was suggested that the first of the vertebrate defensin families is the β-defensins. The phylogenetic analysis of defensins in both vertebrates and invertebrates revealed that the relationship between invertebrate defensins, Cysteine-stabilized α-helix/β-sheet motif (CSαβ), and vertebrate defensins, β-defensins, was closer than that between α- defensins and β-defensins [38]. In addition, β-defensins are present in fish, birds, and reptiles, which are phylogenetically distant groups [39]. Interestingly, CSαβ defensin peptides display high homology among plants, fungi, and invertebrates. These findings indicate that they may share a common genetic origin [40].
Action mechanism of antimicrobial peptides
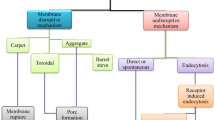
The action mechanism of AMPs depends on various physicochemical properties, such as the sequence of amino acids, structure-especially secondary structure, charge, and amphipathic property [3]. AMPs have different mechanisms by which they kill target microbes (Fig. 2) [41]. AMPs mainly interfere with the cytoplasmic membrane of the target, and may bind essential molecules in the living cells, leading to the inhibition of DNA, RNA, or protein synthesis, as well as the inhibition of certain enzymes and cell wall synthesis, in addition to the activation of autolysin, and distortion of cytoplasmic membrane (Fig. 2A) [42]. AMPs cause bacterial death through multiple and harmonizing actions, referred to as a multi-hit mechanism [7]. This strategy boosts the efficiency of AMPs to evade resistance development.

Mechanism of action of AMPs; inhibition of vital cell processes (a), direct pore information (b)
AMPs bind to bacterial membranes through three models [42] (Fig. 2B). The first is the barrel-stave model in which peptides are vertically positioned between the membrane, and attached forming an ion channel. Pardaxin and alamethicin act through this mechanism [43]. The second is the toroidal pore wormhole model in which pore formation does not originate from peptide–peptide interactions, rather from the peptide-induced curvature in the lipid bilayer; the pore is generated by both the peptide and the phospholipid head groups [44]. Many AMPs act in the toroidal model, such as protegrin-1, melittin, and magainin-2. The toroidal model shows bilayer disturbance, but remains intact in the barrel-stave model [45]. The third is the carpet model where AMPs are likely to adsorb to the membrane. Once they reach a particular concentration, they apply detergent-like effect that breaks up the membrane by forming micelles. Leucine-Leucine 37 (LL-37) and cecropin adopt the carpet model mechanism [46].
Furthermore, the cytoplasmic membrane of mammalian cells is rich in the zwitterionic phospholipids, phosphatidylethanolamine, sphingomyelin, and phosphatidylcholine, providing a membrane with a neutral net charge, while the acidic phospholipids, such as cardiolipin and phosphatidylglycerol, form the bacterial cell membranes [47]. This difference between mammalian and microbial membrane protects mammalian cells against AMPs (Fig. 3). Contrary to microbes, mammalian cell membranes have a high cholesterol content, which reduces the activity of AMPs [47]. Besides the direct bactericidal action, several AMPs have complex immunomodulatory activities, which include the induction of chemokines and cytokines, pro/anti-inflammatory activity, wound healing, angiogenesis, direct chemotaxis, apoptotic activity, and adjuvant activity [7].

Comparison between interaction of AMP with plasma membrane of animal and bacteria
Biosynthesis pathway of antimicrobial peptides
AMPs are produced in nature either by ribosomal translation of mRNA, or by nonribosomal peptide synthesis. Nonribosomally synthesized peptides are chiefly produced by bacteria [48], whereas ribosomally synthesized AMPs (the most common case in eukaryotes) are genetically encoded, and expressed as inactive precursors (prepropeptides) (Fig. 4). Various AMP genes such as α- and β-defensins are clustered at a single chromosomal locus, and can be co-expressed. The prepropeptide comprises a signal peptide and a pro-domain. The pro-domain keeps the mature peptide inactive until it is required. Therefore, the propeptide is anionic, while the mature peptide is generally cationic. The prodomain can have several biological functions, including intracellular trafficking, inhibition of mature peptide activity, or the correct folding of the C-terminus. After secretion into the extracellular space, the mature peptide is proteolytically released [48]. The pro-domain mostly occurs in the N-terminus, except for plants and some fish species, in which it occurs in the C-terminus. Usually, the signal peptide is more conserved than the mature peptide. The variability of mature peptide is a result of adaptation to challenges by pathogens. The majority of the gene-encoded AMPs undergoes post-translational modifications that are important to their function and structure [42]. Currently, there are more than 15 types of these modifications, including glycosylation, and C-terminal and N-terminal capping (amidation acetylation, and pyroglutamic acid formation), as well as disulfide-bridge formation, hydroxylation, phosphorylation, halogenation, etc. [48].

Schematic representation of precursor structure of AMPs
Cathelicidins and defensins families
Cathelicidins and defensins represent the largest cationic families of AMPs. They characterize the most important part of the immune system in humans and farm animals [49]. Cathelicidins are a family of cationic AMPs widely found in mammals. This family comprises more than 30 members. While there is only one cathelicidin gene in humans and mice, there are many in other mammals [50]. LL-37 is the only member of the cathelicidin expressed in humans [42]. The genes responsible for the synthesis of cathelicidins are about 2 kb, and comprise four-exons and three introns. The first exon covers the signal peptide (part pre-) (29–30 aa). The second and third exons encode the cathelin domain (part pro-) (99–114 aa). The mature peptide (12–100 aa) is encoded by the fourth exon. Cathelicidin peptides are produced by dendritic cells, lymphocytes, NK cells macrophages, epithelial cells, neutrophils, and keratinocytes [51]. The propeptides are stored as inactive precursors, so that when the mature peptide is required, it is cleaved by neutrophil elastase to be released. The signal peptide and cathelin domain are highly conserved among species and different peptides, whereas the mature peptide shows significant heterogeneity. In some cathelicidin peptides, the C-terminal regions are α-helical, β-hairpin, or proline/arginine-rich [51].
Nonetheless, defensins are cationic peptides with a molecular mass of 3.5–4.5 kDa, containing six cysteine residues. These residues form three disulfide bridges [48]. They are divided into three classes: α-defensins, β-defensins, and θ-defensins, according to the disulfide pairings between their six conserved cysteine residues. Various AMP genes, such as α- and β-defensins, are clustered at a single chromosomal locus, and can be co-expressed. α- and β-defensins occur in the granules of neutrophils, macrophages, NK cells, epithelial tissues, skin, and in many body fluids. The triple-stranded antiparallel β-sheet structure of α-and β-defensins is stabilized by disulfide and hydrogen bonds [42]. θ-defensins are novel AMPs with a circular structure. They have been isolated from rhesus monkey neutrophils [52]. The majority of genes encoding mammalian defensins consists of only two exons; the first exon encodes the signal peptide, and the second encodes the propeptide and mature peptide [53]. The pro-peptides of α-defensins are larger than those of β-defensins. Human neutrophil defensin is processed by signal peptidase at position 19, in order to release a propeptide that is subjected to further processing by a proteolytic enzyme, trypsin in human, and metalloproteinase-7 in mouse, to release the mature peptide [42].
Identification and prediction of antimicrobial peptides
The prediction and design of effective AMPs have significant roles in meeting the demand for novel efficient antimicrobial therapies. In the past, tissue homogenization was the first step in the identification of novel AMPs, followed by peptide extraction. Chromatographic techniques were used to isolate the crude peptide. Animals were sometimes exposed to bacterial infection or electric shocks to stimulate AMP production [48]. Afterwards, assay-guided fractionation was used to isolate potential AMPs, and the sequence was determined using special techniques, including mass spectrometry and Edman degradation. Despite its success, this approach has disadvantages, as it is time-consuming and produces low yields [48].
The rapid development and cost reductions of the next-generation sequencing technologies (NGS), in combination with efficient solid-phase synthesis techniques, enabled functional testing with no need for polypeptide isolation, nor the search for precious sequence data buried in the genome [48]. A huge amount of data, including protein, RNA, and DNA, have been generated using NGS techniques, by which peptides with antimicrobial activity could be found. Identifying AMPs is only possible through the development of computer-assisted strategies that can automatically estimate a great amount of data, and identify candidates to antimicrobial peptides before their biological evaluation in the wet lab [54]. In recent years, many computational methods have been developed to accelerate the process of antimicrobial-drug prediction and design, by providing a rational basis for candidate selection [55]. Computational research has focused on the recognition of AMPs as a means for determining which features relate to activity. Machine learning algorithms (MLA) represent the main method for training guide sequence-based classifiers to discriminate AMPs from non-AMPs [54]. Machine learning algorithms include, for example, random forest (RF), discriminant analysis (DA), support vector machine (SVM), and artificial neural network (ANN) [56]. The composition of the feature set is the critical factor for a successful prediction method, and should catch the key and accurate modules in the sequence to discriminate the real positives from negatives [55].
Applications of antimicrobial peptides
Besides their prospective therapeutic applications, AMPs have different applications in various fields, such as the food industry, aquaculture, agriculture, and animal husbandry.
Antimicrobial peptides as natural bio-preservative
Chemicals used in food preservation (e.g., nitrites and sulfur dioxide) may have adverse effects on the food’s nutritional value, and on human health [57]. Antimicrobial peptides have recently been used as safe food preservatives [58]. Nisin, commercially known as nisaplin™, is effective against cheese contaminating bacteria (i.e., Staphylococcus aureus and Lactobacillus spp.) [59]. It is also used for the preservation of yogurt, canned vegetables, and juice. Pediocin PA-1, marketed as Alta™ 2341, is effective against Listeria monocytogenes that spoils meat [12].
Antimicrobial peptides in aquaculture
In aquaculture, the use of AMPs is a solution to troubles caused by the widespread use of antibiotics [60]. Jia et al. [61] observed that the daily administration of an amidated form of pleurocidin to Coho Salmon, infected with Vibrio anguillarum, reduced the accumulated mortalities. Antiviral and antifungal effects were also detected when using synthetic peptides [. Moreover, oral administration of epinecidin-1 enhanced the survival of zebrafish and grouper, infected with Vibrio vulnificus and Streptococcus agalactiae [13].
Antimicrobial peptides in pearl farming industry
In the pearl farming industry, a bacterial infection is considered a major problem because it leads to nucleus rejections and oyster mortality. Antibiotics are used to reduce postoperative mortality, and increase pearl quality. Nevertheless, such antibiotics pose an environmental and public health issues [62]. Lately, AMPs have been applied in the pearl industry. Simon-Colin et al. [62] used tachyplesin combined with exopolysaccharides, as filming agents, to reduce oyster post-operative mortality, and increase pearl quality.
Antimicrobial peptides in plants
Transformation with gene-encoding AMPs is currently a promising approach to develop transgenic crops resistant to pathogens, thus reducing the use of harmful pesticides in agriculture [14]. Zainal et al. [63] suggested that defensins expression of chili (Capsicum annuum) in tomatoes improved the resistance of tomatoes to Fusarium sp. Giacomelli et al. [64] revealed that the presence of small defensin-like sequence genes in the genome of the grapevine could inhibit the conidial germination of Botrytis cinerea in the fruit.
Antimicrobial peptides as feed additives
Recently, AMPs have been used as feed additives. Liu et al. [65] recommended AMPs as feed additives for goats in commercial farms, due to their beneficial effects on growth performance, rumen morphology of juvenile goats, and ruminal fermentation function. Peng et al. [66] demonstrated that dietary supplementation with crude recombinant porcine β-defensin 2 had useful effects on the growth and intestinal morphology of weaned piglets, reducing a number of potential pathogens in the caecum, and the incidence of post-weaning diarrhea. Chen et al. [67] revealed that dietary supplementation with AMP improved egg production of hens during the late laying period.
Applications in animal husbandry
AMPs have the ability to overcome difficulties associated with conventional antibiotics, and enhance the quality of animal production [15]. Kerr et al. [68] reported that the expression of a bioactive variant of lysostaphin in the mammary glands of mice suppressed Staphylococcus aureus, the major infectious mastitis pathogen. Donovan et al. [69] succeeded in creating transgenic dairy cows that expressed lysostaphin in their mammary epithelium, as a step to prevent and cure of mastitis.
Therapeutic applications of antimicrobial peptides
AMPs appear to be promising therapeutic drugs for different skin and soft tissue infections. They present a broad spectrum of antimicrobial activity, wound-healing promoting activities, such as angiogenesis, and induction of cell migration and proliferation, in addition to immune-modulatory activity [70]. For example, cathelicidin LL-37 has been used as a local treatment for leg ulcers [71]. A set of AMPs was approved by the food and drug administration (FDA) for clinical use. For example, Omiganan has been established as a local treatment for catheter infections [72]. Daptomycin (approved in 2003), Omiganan (approved in 2014), telavancin (approved in 2014), and dalbavancin (approved in 2009) are used for injection against complicated skin, and skin structure infections caused by different Gram-positive bacterial infections [8]. AMPs also have the potential to prevent sexually transmitted infections. For example, LL-37 has an antimicrobial effect against Chlamydia trachomatis, Candida albicans, and human immunodeficiency virus (HIV) [73]. AMPs could be used as novel contraceptive microbicides. For example, VRP is a small synthetic peptide that can arrest sperm motility without damaging the vaginal epithelial cells [74].
Furthermore, AMPs with anti-tumor activity, called anticancer peptides (ACPs), are new drugs that may overcome the problems associated with tumor resistance to conventional chemotherapy. In recent years, the number of natural AMPs that have antitumor activity has increased [10]. Several AMPs target tumor cells through binding the negatively charged phospholipids, i.e. phosphatidylserines (PS), localized in the outer leaflet of plasma membranes [75]. The outer leaflet of the plasma membrane of normal human cells contains neutral phospholipids, i.e. phosphatidylcholines and sphingomyelins, while the inner leaflet contains negatively charged PS. Nevertheless, this condition is inverted in cancer, owing to oxidative stress, acidity, thrombin, and inflammatory cytokines [10]. The high fluidity of cancer cells, due to the decreased levels of cholesterol, facilitates AMP-induced apoptosis [76]. Numerous ACPs have lately been discovered. For example, Cecropin B exhibits anticancer activity against lung and stomach cancer, and leukemia cells [1]. Bacteriocin was found to have cytotoxic activity against various human cancer cells [77]. Qin et al. [78] revealed that the novel modified peptide of cecropin B, CB1a showed an apoptosis activity in carcinoma cells.
Besides the direct administration of AMPs, several agents induce the expression of AMPs by the body to improve immune responses [7]. Vitamin D3 was shown to increase the gene expression of cathelicidin [79]. Recently, it has been suggested that the combination of AMPs with traditional antibiotics increases their efficiency [80].
Challenges to clinical applications
Limitations on the clinical applications of AMPs are imposed by the susceptibility to proteolytic degradation, toxicity, specificity, immunogenicity concerns, rapid clearance from the kidney and liver, hemolytic activity, and delivery issues [81, 82]. For instance, whereas Indolicidin has a broad spectrum of antimicrobial activity, it exhibits hemolytic activity that limits its clinical application [3]. Furthermore, several of these cationic AMPs were reported to be more or less toxic to human cells [83]. Immunogenicity is also a serious obstacle in peptide drug development, although AMPs are small molecules with little to no immunogenicity. As a result, improving the safety of AMPs is a critical concern in drug research and development [3].
Different strategies have been explored to obtain safer AMPs as a potential drug candidate [73]. The formulation and chemical modification of the peptides are among the most commonly used strategies. Natural AMPs can be modified chemically by glycosylation [84], non-natural amino acids incorporation [85], acetylation, lipidation, and cyclization [86]. Kumar et al. [87] reported an increase in proteolysis stability of AMPs when positively-charged arginine residue was substituted with charged non-natural amino acids (e.g. L-ornithine and L-homoarginine). Wang et al. [88] utilized a peptidomimetics strategy, in which polymers with a modified backbonewere used to prevent proteolysis, and increase stability. Diverse delivery systems, such as surfactant/lipid-self assembly systems, inorganic materials, and polymers, were used to improve the half-life, stability, and toxicity of AMPs [89]. Lately, nanotechnology has been used as a convenient system to control the release and the delivery of AMPs [90].
Potential use against COVID-19
Coronaviruses (CoVs) are a family of viruses that has the ability to infect various animals, and cause respiratory illness in humans. Severe acute respiratory syndrome coronavirus (SARS-CoV), and Middle East respiratory syndrome coronavirus (MERS-CoV) are examples of extremely pathogenic coronaviruses that emerged in humans, causing fatal respiratory illness [91]. COVID-19 is an infectious disease detected in China in 2019, and its causative agent is SARS-CoV-2. It has resulted in global deaths that exceeded 5.5 million since its detection (https://coronavirus.jhu.edu/map.html). Considering the acute crisis of COVID-19 pandemic, there is an urgent need for developing effective antiviral therapeutics for the prevention and treatment of COVID-19.
CoVs are enveloped viruses with single-stranded ribonucleic acid (ssRNA). Their genome is considered the largest RNA virus (26 to 32 kb). The 3ʹ terminus of SARS-CoV-2 encodes envelope (E), spike (S) glycoproteins, nucleocapsid (N) proteins, and membrane (M) glycoproteins (Fig. 5) [92, 93]. The viral infection in humans is initiated by the binding of receptor-binding domain (RBD) of spike protein to human angiotensin-converting enzyme 2 (hACE2) receptor. It is mainly expressed on the epithelial cells of lung; thus, the main target of SARS-CoV-2 is the lung [94]. In addition to the genes that encode structural proteins, certain regions at the genome encode non-structural proteins, such as coronavirus’ main protease (3CLpro), and papain-like protease (PLpro), and the viral proteins required for replication [95]. Therefore, the targets of antiviral drugs are generally the blockage of host cell surface receptors, virolysis (direct interaction with virus), inhibition of viral replication, and the inhibition of viral fusion to host cells [96].

Schematic figure of coronavirus structure
Despite the partial protection conferred by the present vaccines against viruses prior to exposure, antiviral drugs are the first line of defense when individuals are already infected. However, a specific antiviral with high efficiency against SARS-CoV-2 is not yet available [97]. Among the 3324 discovered or synthesized AMPs, 190 peptides have antiviral activity, and called antiviral peptides (AVPs) [9]. Since AMPs have inhibitory effects on the known proteins of the SARS-CoV-2 virus, their usage is a significantly promising solution for the treatment of COVID-19 [98].
The lectin-like human defensins-5 peptide (HD5) was reported to have high-affinity binding with ACE2, protecting host cells from viral infection [94]. Bakovic et al. [99] suggested that brilacidin (a de novo designed synthetic AMP) had a potent antiviral activity in vitro against SARS-CoV-2. Its proposed mechanisms include preventing viral binding to cells, and enhancing the integrity of the viral membrane. Using in silico molecular docking studies, it was found that several AVPs could prevent COVID-19. Fakih [100] confirmed that dermaseptin-S9 peptide could prohibit the attachment of the spike protein of SARS-CoV-2 to the surface of the ACE-2 receptor. Liscano et al. [101] reported that AMPs might be selective for viral proteins, concluding that caerin 1.6 and caerin 1.10 (Amphibian AMPs) had the ability to interact with Sgp, yet had low affinity for ACE2 proteins. More recently, Bhattacharya et al. [102] unraveled that the binding affinity of the natural food preservative peptide, Nisin, to hACE2 was higher than that of the spike protein of SARS-CoV-2. Furthermore, using molecular docking and molecular dynamics simulations, Balmeh et al. [98] revealed that glycocin F and lactococcine G were the best bio-AMPs to block RdRp, 3CL, S, and N proteins of SARS-CoV-2, with minimal side effects. It was found that the antibacterial peptide DP7 had a potential effect on inhibiting SARS-CoV-2 infection, through SARS-CoV-2 S protein pseudovirus infection of ACE2-293 T cells, SARS-CoV-2 S protein-mediated cell–cell fusion, and the inhibition of SARS-CoV-2-3CLpro enzyme activity [103]. These findings reported that AVPs could be effective in treating COVID-19 infection. Nevertheless, experimental and preclinical studies are necessary to assay their therapeutic effects.
Conclusion
Since the discovery of lysozyme in 1922, more than 3324 AMPs have been recognized in most organisms. AMPs are evolutionarily conserved molecules. These small cationic peptides exhibit structural and functional diversity, and have the ability to target bacteria, viruses, fungi, and parasites, as well as cancer cells. The accelerating growth of antimicrobial resistance has deepened the need for the discovery of novel antimicrobial agents. Some limitations are still imposed on pharmaceutical development, due to immunogenicity, proteolytic degradation, and toxicity. Recently, different strategies have been explored, including chemical modifications, and modern techniques, such as drug delivery, in order to obtain safer AMPs as potential therapeutic agents. Increasing evidence shows that AMPs can inhibit SARS-CoV-2 (COVID-19), paving the way for their prospective use as therapeutic drugs.
References
Seyfi R, Abarghooi F, Tahereh K, Soheila E, Eyvazi MS, Babaeipour V, Tarhriz, V (2020) Antimicrobial peptides (AMPs): roles, functions and mechanism of action. Int J Pept Res Ther 26:1451–1463. https://doi.org/10.1007/s10989-019-09946-9
Huan Y, Kong Q, Mou H, Yi H (2020) Antimicrobial peptides: classification, design, application and research progress in multiple fields. Front Microbiol 11:582779. https://doi.org/10.3389/fmicb.2020.582779
Lei J, Sun L, Huang S, Zhu C, Li P, He J, Mackey V, Coy DH, He Q (2019) The antimicrobial peptides and their potential clinical applications. Am J Transl Res 11(7):3919–3931
Shi Y, Li C, Wang M, Chen Z, Luo Y, Xia XS, Song Y, Sun Y, Zhang AM (2020) Cathelicidin-DM is an antimicrobial peptide from duttaphrynus melanostictus and has wound-healing therapeutic potential. ACS Omega 5(16):9301–9310. https://doi.org/10.1021/acsomega.0c00189
Zhang LJ, Gallo RL (2016) Antimicrobial peptides. Curr Biol 26(1):R14–R19. https://doi.org/10.1016/j.cub.2015.11.017
Muller C (2022) Antibiotics and antimicrobials resistance: mechanisms and new strategies to fight resistant bacteria. Antibiotics 11(3):400. https://doi.org/10.3390/antibiotics11030400.PMID:35326863
MahlapuuHAkanssonRingstadBjörn MJLC (2016) Antimicrobial peptides: an emergingcategory of therapeutic agents. Front Cell Infect Microbiol. https://doi.org/10.3389/fcimb.2016.00194
Chen CH, Lu TK (2020) Development and challenges of antimicrobial peptides for therapeutic applications. Antibiotics 9(1):24. https://doi.org/10.3390/antibiotics9010024
Wang G (2020) The antimicrobial peptide database provides a platform for decoding the design principles of naturally occurring antimicrobial peptides. Protein Sci 29(1):8–18. https://doi.org/10.1002/pro.3702
Tornesello AL, Borrelli A, Buonaguro L, Buonaguro FM, Tornesello ML (2020) Antimicrobial peptides as anticancer agents: functional properties and biological activities. Molecules 25(12):2850. https://doi.org/10.3390/molecules25122850
Mousavi Maleki MS, Rostamian M, Madanchi H (2021) Antimicrobial peptides and other peptide-like therapeutics as promising candidates to combat SARS-CoV-2. Expert Rev Anti Infect Ther 12:1–13. https://doi.org/10.1080/14787210.2021.1912593
Khorshidian N, Khanniri E, Mohammadi M, Mortazavian AM, Yousefi M (2021) Antibacterial activity of pediocin and pediocin-producing bacteria against Listeria monocytogenes in meat products. Front Microbiol 12:709959. https://doi.org/10.3389/fmicb.2021.709959
Chee PY, Mang M, Lau ES, Tan LT-H, He Y-W, Lee W-L, Pusparajah P, Chan K-G, Lee L-H, Goh B-H (2019) Epinecidin-1, an antimicrobial peptide derived from grouper (Epinephelus coioides): pharmacological activities and applications. Front Microbiol 10:2631. https://doi.org/10.3389/fmicb.2019.02631
Meneguetti BT, Machado LD, Oshiro KG, Nogueira ML, Carvalho CM, Franco OL (2017) Antimicrobial peptides from fruits and their potential use as biotechnological tools-a review and outlook. Front Microbiol 7:2136. https://doi.org/10.3389/fmicb.2016.02136
Kumar R, Ali SA, Singh SK, Bhushan V, Mathur M, Jamwal S, Mohanty AK, Kaushik JK, Kumar S (2020) Antimicrobial peptides in farm animals: an updated review on its diversity, function, modes of action and therapeutic prospects. Vet Sci 7(4):206. https://doi.org/10.3390/vetsci7040206
Fleming A (1929) On the antibacterial action of cultures of a Penicillium, with special reference to their use in the isolation of B. influenzæ. Br J Exp Pathol 10(3):226–236
Steiner H, Hultmark D, Engstrom A, Bennich H, Boman HG (1981) Sequence and specific city of two anti-bacterial proteins involved in insect immunity. Nature 292:246–248
Zasloff M (1987) Magainins, a class of antimicrobial peptides from Xenopusskin: isolation, characterization of two active forms, and partial cDNA sequence of a precursor. Proc Natl Acad Sci USA 84:5449–5453
Bahar AA, Ren D (2013) Antimicrobial peptides. Pharmaceuticals 6:1543–1575. https://doi.org/10.3390/ph6121543
APD. Antimicrobial Peptide Database (APD; http://aps.unmc.edu/AP, accessed on 6 January, 2022).
Bechinger B, Juhl DW, Glattard E, Aisenbrey C (2020) Revealing the mechanisms of synergistic action of two magainin antimicrobial peptides. Front Med Technol 2:615494. https://doi.org/10.3389/fmedt.2020.615494
Zhang SK, Song J, Gong F, Li SB, Chang HY, Xie HM, Gao HW, Tan YX, Ji SP (2016) Design of an α-helical antimicrobial peptide with improved cell-selective and potent anti-biofilm activity. Sci Rep 6:27394. https://doi.org/10.1038/srep27394
Sarkar T, Chetia M, Chatterjee S (2021) Antimicrobial peptides and proteins: from nature’s reservoir to the laboratory and beyond. Front Chem 9:691532. https://doi.org/10.3389/fchem.2021.691532
Shahbaaz M, Al-Samghan AS, Malik A, Afaq S, Alwabli AS, Ahmad I, Hussien MA, Zubair M, Fatima F, Begum S, Alzahrani O, Tarique M (2020) Functional and structural analysis of predicted proteins obtained from Homo sapiens minisatellite 33.15-tagged transcript pAKT-45 variants. BioMed Res Int. https://doi.org/10.1155/2020/2562950
Simons A, Alhanout K, Duval RE (2020) Bacteriocins, antimicrobial peptides from bacterial origin: overview of their biology and their impact against multidrug-resistant bacteria. Microorganisms 8(5):639. https://doi.org/10.3390/microorganisms8050639
Mylonakis E, Podsiadlowski L, Muhammed M, Vilcinskas A (2016) Diversity, evolution and medical applications of insect antimicrobial peptides. Philos Trans R 371(1695):20150290. https://doi.org/10.1098/rstb.2015.0290
Moretta A, Scieuzo C, Petrone AM, Salvia R, Manniello MD, Franco A, Lucchetti D, Vassallo A, Vogel H, Sgambato A, Falabella P (2021) Antimicrobial peptides: a new hope in biomedical and pharmaceutical fields. Front Cell Infect Microbiol 14(11):668632. https://doi.org/10.3389/fcimb.2021.668632
Ponnappan N, Budagavi DP, Yadav BK, Chugh A (2015) Membrane-active peptides from marine organisms–antimicrobials, cell-penetrating peptides and peptide toxins: applications and prospects. Probiotics Antimicrob Proteins 7(1):75–89. https://doi.org/10.1007/s12602-014-9182-2
König E, Bininda-Emonds OR, Shaw C (2015) The diversity and evolution of anuran skin peptides. Peptides 63:96–117. https://doi.org/10.1016/j.peptides.2014.11.003
Nazeer N, Uribe-Diaz S, Rodriguez-Lecompte JC, Ahmed M (2021) Antimicrobial peptides as an alternative to relieve antimicrobial growth promoters in poultry. Br Poult Sci 62(5):672–685. https://doi.org/10.1080/00071668.2021.1919993
Mahrous KF, Mabrouk DM, Aboelenin MM, El-Kader HAAM, Gaafar AY, Younes AM, Mahmoud MA, Khalil WKB, Hassanane MS (2020) Molecular characterization and immunohistochemical localization of tilapia piscidin 3 in response to Aeromonas hydrophila infection in nile tilapia. J Pept Sci 26(11):e3280. https://doi.org/10.1002/psc.3280
Mahrous KF, Mabrouk DM, Aboelenin MM, Kader HAAE, Hassanane MS (2021) Identification and characterization of antimicrobial peptide genes in Clarias gariepinus and Chelon ramada. Jordan J Biol Sci 14(1):51–64
Zhu S, Gao B (2017) Positive selection in cathelicidin host defense peptides: adaptation to exogenous pathogens or endogenous receptors? Heredity 118(5):453–465. https://doi.org/10.1038/hdy.2016.117
Hanson MA, Hamilton PT, Perlman SJ (2016) Immune genes and divergent antimicrobial peptides in flies of the subgenus Drosophila. BMC Evol Biol 16(1):228. https://doi.org/10.1186/s12862-016-0805-y
Schmitt P, Rosa RD, Destoumieux-Garzón D (2016) An intimate link between antimicrobial peptide sequence diversity and binding to essential components of bacterial membranes. Biochimica et Biophysica Acta. https://doi.org/10.1016/j.bbamem.2015.10.011
Vy HMT, Won Y, Kim Y (2017) Multiple modes of positive selection shaping the patterns of incomplete selective sweeps over African populations of Drosophila melanogaster. Mol Biol Evol 34:2792–2807. https://doi.org/10.1093/molbev/msx207
Chapman JR, Hill T, Unckless RL (2019) Balancing selection drives the maintenance of genetic variation in Drosophila antimicrobial peptides. Genome Biol Evol 11(9):2691–2701. https://doi.org/10.1093/gbe/evz191
Zhu S, Gao b (2012) Evolutionary origin of β-defensins. Dev Comp Immunol. https://doi.org/10.1016/j.dci.2012.02.011
Machado LR, Ottolini B (2015) An evolutionary history of defensins: a role for copy number variation in maximizing host innate and adaptive immune responses. Front Immunol 6:115. https://doi.org/10.3389/fimmu.2015.00115
Dias Rde O, Franco OL (2015) Cysteine-stabilized αβ defensins: from a common fold to antibacterial activity. Peptides 72:64–72. https://doi.org/10.1016/j.peptides.2015.04.017
Wang Q, Yang S, Liu J, Terecskei K, Ábrahám E, Gombár A, Wang T (2017) Host-secreted antimicrobial peptide enforcessymbiotic selectivity in Medicago truncatula. Proc Natl AcadSci USA 114:6854–6859
Zhao BR, Zheng Y, Gao J, Wang XW (2020) Maturation of an antimicrobial peptide inhibits Aeromonas hydrophila infection in crayfish. J Immunol 204(3):487–497. https://doi.org/10.4049/jimmunol.1900688
Harder J, Schroder JM (2016) Antimicrobial peptides. Springer, Cham
Gaspar D, Castanho MA (2016) Anticancer peptides: prospective innovation in cancer therapy. In: Epand R (ed) Host defense peptides and their potential as therapeutic agents. Springer, Cham
Di Somma A, Avitabile C, Cirillo A, Moretta A, Merlino A, Paduano L, Duilio A (1864) Romanelli A (2020) The antimicrobial peptide Temporin L impairs E. coli cell division by interacting with FtsZ and the divisome complex. Biochim Biophys Acta Gen Subj. https://doi.org/10.1016/j.bbagen.2020.129606
Geitani R, Moubareck CA, Xu Z, Karam Sarkis D, Touqui L (2020) Expression and roles of antimicrobial peptides in innate defense of airway mucosa: potential implication in cystic fibrosis. Front Immunol 11:1198. https://doi.org/10.3389/fimmu.2020.01198
Greco I, Molchanova N, Holmedal E, Jenssen H, Hummel BD, Watts JL, Håkansson J, Hansen PR, Svenson J (2020) Correlation between hemolytic activity, cytotoxicity and systemic in vivo toxicity of synthetic antimicrobial peptides. Sci Rep 10(1):13206. https://doi.org/10.1038/s41598-020-69995-9
Roncevic T, Puizina J, Tossi A (2019) Antimicrobial peptides as anti-infective agents in pre-post-antibiotic era? Int J Mol Sci 20(22):5713. https://doi.org/10.3390/ijms20225713
Peel E, Cheng Y, Djordjevic JT, O’Meally D, Thomas M, Kuhn M, Sorrell TC, Huston WM, Belov K (2021) Koala cathelicidin PhciCath5 has antimicrobial activity, including against Chlamydia pecorum. PLoS ONE 16(4):e0249658. https://doi.org/10.1371/journal.pone.0249658
Silva T, Gomes MS (2017) Immuno-stimulatory peptides as a potential adjunct therapy against intra-macrophagic pathogens. Molecules 22(8):1297. https://doi.org/10.3390/molecules22081297
Scheenstra MR, van Harten RM, Veldhuizen EJA, Haagsman HP, Coorens M (2020) Cathelicidins modulate TLR-activation and inflammation. Front Immunol 11:1137. https://doi.org/10.3389/fimmu.2020.01137
Schaal JB, Maretzky T, Tran DQ, Tran PA, Tongaonkar P, Blobel CP, Ouellette AJ, Selsted ME (2018) Macrocyclic θ-defensins suppress tumor necrosis factor-α (TNF-α) shedding by inhibition of TNF-α-converting enzyme. J Biol Chem 293(8):2725–2734. https://doi.org/10.1074/jbc.RA117.000793
Meade KG, O’Farrelly C (2019) β-defensins: farming the microbiome for homeostasis and health. Front Immunol 9:3072. https://doi.org/10.3389/fimmu.2018.03072
Beltran JA, Rio JD, Brizuela CA (2020) An automatic representation of peptides for effective antimicrobial activity classification. Comput Struct Biotechnol J 18:455–463. https://doi.org/10.1016/j.csbj.2020.02.002
Bhadra P, Yan J, Li J (2018) AmPEP: Sequence-based prediction of antimicrobial peptides using distribution patterns of amino acid properties and random forest. Sci Rep 8:1697. https://doi.org/10.1038/s41598-018-19752-w
Han T, JiangD ZQ, Wang L, Yin K (2018) Comparison of random forest, artificial neural networks and support vector machine for intelligent diagnosis of rotating machinery. Trans Inst Meas Control 40(8):2681–2693. https://doi.org/10.1177/0142331217708242
Rai M, Pandit R, Gaikwad S, o¨vics G, (2016) Antimicrobial peptides as natural bio-preservative to enhance the shelf-life of food. J Food Sci Technol 53:3381–3394. https://doi.org/10.1007/s13197-016-2318-5
Wang S, Zeng X, Yang Q, Qiao S (2016) Antimicrobial peptides as potential alternatives to antibiotics in food animal industry. Int J Mol Sci 17(603):1–12
Upendra RS, Khandelwal P, Jana K, Ajay Kumar N, Gayathri Devi M, Stephaney ML (2017) Bacteriocin production from indigenous strains of lactic acid bacteria isolated from selected fermented food sources. Int J Pharma Res Health Sci 4(1):982–990
Okocha RC, Olatoye IO, Adedeji OB (2018) Food safety impacts of antimicrobial use and their residues in aquaculture. Public Health Rev 39:1. https://doi.org/10.1186/s40985-018-0099-2
Jia X, Patrzykat A, Devlin RH, Ackerman PA, Iwama GK, Hancock REW (2000) Antimicrobial peptides protect Coho salmon from Vibrio Anguillarum infections. Appl Environ Microbiol 66:1928–1932. https://doi.org/10.1128/AEM.66.5.1928-1932.2000
Simon-Colin C, Gueguen Y, Bachere E (2015) Use of natural antimicrobial peptides and bacterial biopolymers for cultured pearl production. Mar Drugs 13(6):3732–3744. https://doi.org/10.3390/md13063732
Zainal Z, Marouf E, Ismail I, Fei CK (2009) Expression of the Capsicuumannum chili defensin gene in transgenic tomatoes confers enhanced resistance to fungal pathogens. Am J Plant Physiol 4:70–79. https://doi.org/10.3923/ajpp.2009.70.79
Giacomelli L, Nanni V, Lenzi L, Zhuang J, Dalla Serra M, Banfield MJ, Town CD, Silverstein KA, Baraldi E, Moser C (2012) Identification and characterization of the defensin-like gene family of grapevine. Mol Plant Microbe Interact 25(8):1118–1131. https://doi.org/10.1094/MPMI-12-11-0323
Liu Q, Yao S, Chen Y, Gao S, Yang Y, Deng J, Ren Z, Shen L, Cui H, Hu Y, Ma X, Yu S (2017) Use of antimicrobial peptides as a feed additive for juvenile goats. Sci Rep 7:12254. https://doi.org/10.1038/s41598-017-12394-4
Peng Z, Wang A, Xie L, Song W, Wang J, Yin Z, Zhou D, Li F (2016) Use of recombinant porcine β-defensin 2 as a medicated feed additive for weaned piglets. Sci Rep 6:26790. https://doi.org/10.1038/srep26790
Chen X, Zhan Y, Ma W, Zhu Y, Wang Z (2020) Effects of Antimicrobial peptides on egg production, egg quality and caecal microbiota of hens during the late laying period. Anim Sci J. https://doi.org/10.1111/asj.13387
Kerr DE, Plaut K, Bramley AJ, Williamson CM, Lax AJ, Moore K, Wells KD, Wall RJ (2001) Lysostaphin expression in mammary glands confers protection against staphylococcal infection in transgenic mice. Nat Biotechnol 19:66–70. https://doi.org/10.1038/83540
Donovan DM, Kerr DE, Wall RJ (2005) Engineering disease resistant cattle. Transgenic Res 14:563–567. https://doi.org/10.1007/s11248-005-0670-8
Hilchie AL, Hoskin DW (2019) Power coombs MR. anticancer activities of natural and synthetic peptides. Adv Exp Med Biol 1117:131–147. https://doi.org/10.1007/978-981-13-3588-4_9
Krejner A, Litwiniuk M, Grzela T (2017) LL-37 but Not 25-hydroxy-vitamin D serum level correlates with healing of venous leg ulcers. Arch Immunol Ther Exp 5:455–461. https://doi.org/10.1007/s00005-016-0423-9
Mahlapuu M, Björn K, Ekblom J (2020) Antimicrobial peptides as therapeutic agents: opportunities and challenges. Crit Rev Biotechnol 40(7):978–992. https://doi.org/10.1080/07388551.2020.1796576
Madanchi H, Sardari S (2019) The role of antimicrobial peptides in the prevention of sexually transmitted infection (STI). Womens Health 8(2):192–194. https://doi.org/10.15406/mojwh.2019.08.00234
Ghosh P, Bhoumik A, Saha S, Mukherjee S, Azmi S, Ghosh JK, Dungdung SR (2018) Spermicidal efficacy of VRP, a synthetic cationic antimicrobial peptide, inducing apoptosis and membrane disruption. J Cell Physiol 233(2):1041–1050. https://doi.org/10.1002/jcp.25958
Wodlej C, Riedl S, Rinner B, Leber R, Drechsler C, Voelker DR, Choi JY, Lohner K, Zweytick D (2019) Interaction of two antitumor peptides with membrane lipids—influence of phosphatidylserine and cholesterol on specificity for melanoma cells. PLoS ONE 14(1):e0211187
Beloribi-Djefaflia S, Vasseur S, Guillaumond F (2016) Lipid metabolic reprogramming in cancer cells. Oncogenesis 5:e189. https://doi.org/10.1038/oncsis.2015.49
Ankaiah D, Palanichamy E, Antonyraj CB, Ayyanna R, Perumal V, Ahamed SIB, Arul V (2018) Cloning, overexpression, purification of bacteriocin enterocin-B and structural analysis, interaction determination of enterocin-A, B against pathogenic bacteria and human cancer cells. Int J Biol Macromol 116:502–512. https://doi.org/10.1016/j.ijbiomac.2018.05.002
Qin Y, Malik A, Afaq S, Alwabli AS (2019) From antimicrobial to anticancer peptides: the transformation of peptides. Recent Pat Anti-cancer Drug Discov 14:70–84
Vargas Buonfiglio LG, Cano M, Pezzulo AA (2017) Effect of vitamin D3 on the antimicrobial activity of human airway surface liquid: preliminary results of a randomised placebo-controlled double-blind trial. BMJ Open Respir Res 4(1):e000211. https://doi.org/10.1136/bmjresp-2017-000211
Zharkova MS, Orlov DS, Golubeva OY, Chakchir OB, Eliseev IE, Grinchuk TM, Shamova OV (2019) Application of ntimicrobial peptides of the innate immune system in combination with conventional antibiotics—a novel way to combat antibiotic resistance? Front Cell Infect Microbiol 9:128. https://doi.org/10.3389/fcimb.2019.00128
Borah A, Chakraborty BDS (2020) A crosstalk on antimicrobial peptides. Int J Pept Res Ther. https://doi.org/10.1007/s10989-020-10075-x
Divyashree M, Mani MK, Reddy D, Kumavath R, Ghosh P, Azevedo V, Barh D (2020) Clinical applications of antimicrobial peptides (AMPs): where do we stand now? Protein Pept Lett 27(2):120–134. https://doi.org/10.2174/0929866526666190925152957
Lei J, Sun L, Huang S, Zhu C, Li P, He J, Mackey V, Coy DH, He Q (2019) The antimicrobial peptides and their potential clinical applications. Am J Transl Res 11(7):3919–3931
Moradi SV, Hussein WM, Varamini P, Simerska P, Toth I (2016) Glycosylation, an effective synthetic strategy to improve the bioavailability of therapeutic peptides. Chem Sci 7(4):2492–2500. https://doi.org/10.1039/c5sc04392a
Erak M, Bellmann-Sickert K, Els-Heindl S, Beck-Sickinger AG (2018) Peptide chemistry toolbox-transforming natural peptides into peptide therapeutics. Bioorg Med Chem 26(10):2759–2765. https://doi.org/10.1016/j.bmc.2018.01.012
Kowalczyk R, Harris PW et al (2017) Peptide lipidation–a synthetic strategy to afford peptide based therapeutics. In: Care A, Bergquist PL (eds) Sunna A: peptides and peptide-based biomaterials and their biomedical applications. Springer, New York, pp 185–227
Kumar P, Kizhakkedathu JN, Straus SK (2018) Antimicrobial peptides: diversity, mechanism of action and strategies to improve the activity and biocompatibility in vivo. Biomolecules 8(1):4. https://doi.org/10.3390/biom8010004
Wang J, Dou X, Song J (2019) Antimicrobial peptides: promising alternatives in the post feeding antibiotic era. Med Res Rev 39(3):831–859. https://doi.org/10.1002/med.21542
Makowski M, Silva ÍC, Pais do Amaral C, Gonçalves S, Santos NC, (2019) Advances in lipid and metal nanoparticles for antimicrobial peptide delivery. Pharmaceutics 11(11):588. https://doi.org/10.3390/pharmaceutics11110588
Biswaro LS, da Costa Sousa MG, Rezende TMB, Dias SC, Franco OL (2018) Antimicrobial peptides and nanotechnology. Recent Adv Chall Front Microbiol 8(9):855. https://doi.org/10.3389/fmicb.2018.00855
Hu B, Guo H, Zhou P, Shi ZL (2021) Characteristics of SARS-CoV-2 and COVID-19. Nat Rev Microbiol 19(3):141–154. https://doi.org/10.1038/s41579-020-00459-7
Schoeman D, Fielding BC (2019) Coronavirus envelope protein: current knowledge. Virol J 16:69. https://doi.org/10.1186/s12985-019-1182-0
Yu F, Du L, Ojcius DM, Pan C, Jiang S (2020) Measures for diagnosing and treating infections by a novel coronavirus responsible for a pneumonia outbreak originating in Wuhan. China Microbes Infect 22(2):74–79. https://doi.org/10.1016/j.micinf.2020.01.003
Wang C, Wang S, Li D, Zhao X, Han S, Wang T (2020) Lectin-like Intestinal Defensin Inhibits 2019-nCoV spike bind ing to ACE2. Biorxiv. https://doi.org/10.1101/2020.03.29.013490
Cui J, Li F, Shi ZL (2019) Origin and evolution of pathogenic coronaviruses. Nat Rev Microbiol 17(3):181–192. https://doi.org/10.1038/s41579-018-0118-9
Mustafa S, Balkhy H, Gabere MN (2018) Current treatment options and the role of peptides as potential therapeutic components for Middle East Respiratory Syndrome (MERS): a review. J Infect Public Health 11(1):9–17. https://doi.org/10.1016/j.jiph.2017.08.009
Tonk M, R ̊užekD, Vilcinskas A, (2021) Compelling evidence for the activity of antiviral peptidesagainst SARS-CoV-2. Viruses 13:912. https://doi.org/10.3390/v13050912
Balmeh N, Mahmoudi S, Fard NA (2021) Manipulated bio antimicrobial peptides from probiotic bacteria as proposed drugs for COVID-19 disease. Inform Med Unlocked 23:100515. https://doi.org/10.1016/j.imu.2021.100515
Bakovic A, Risner K, Bhalla N, Alem F, Chang TL, Weston WK, Harness JA, Narayanan A (2021) Brilacidin demonstrates inhibition of SARS-CoV-2 in cell culture. Viruses 13(2):271. https://doi.org/10.3390/v13020271
Fakih TM (2020) Dermaseptin-based antiviral peptides to prevent COVID-19 through in silico molecular docking studies against SARS-CoV-2 spike protein. Pharm Sci Res 7(4):8. https://doi.org/10.7454/psr.v7i4.1079
Liscano Y, Oñate-Garzón J, Ocampo-Ibáñez ID (2020) In silico discovery of antimicrobial peptides as an alternative to control SARS-CoV-2. Molecules 25(23):5535. https://doi.org/10.3390/molecules25235535
Bhattacharya R, Gupta AM, Mitra S, Mandal S, Biswas SR (2021) A natural food preservative peptide nisin can interact with the SARS-CoV-2 spike protein receptor human ACE2. Virology. 552:107–111. https://doi.org/10.1016/j.virol.2020.10.002
Zhang R, Jiang X, Qiao J, Wang Z, Tong A, Yang J, Yang S, Yang L (2021) Antimicrobial peptide DP7 with potential activity against SARS coronavirus infections. Signal Transduct Target Ther 6(1):140. https://doi.org/10.1038/s41392-021-00551-1
Funding
Open access funding provided by The Science, Technology & Innovation Funding Authority (STDF) in cooperation with The Egyptian Knowledge Bank (EKB). The authors declare that no fund was received during the preparation of this manuscript.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Mabrouk, D.M. Antimicrobial peptides: features, applications and the potential use against covid-19. Mol Biol Rep 49, 10039–10050 (2022). https://doi.org/10.1007/s11033-022-07572-1
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11033-022-07572-1