
Abstract
Context
Landscape habitat amount is known to increase biodiversity, while the effects of habitat fragmentation are still debated. It has been suggested that negative fragmentation effects may occur with a time lag, which could explain inconsistent results. However, there is so far no empirical support for this idea.
Objectives
We evaluated whether habitat amount and fragmentation at the landscape scale affect the species density of deadwood-dwelling lichens, and whether these effects occur with a time lag.
Methods
We surveyed deadwood-dwelling lichens in woodland key habitats in two regions in northern Sweden, and modelled their species density as a function of past (1960s) and present (2010s) habitat amount (old forest area) and fragmentation (edge density) in the surrounding landscapes.
Results
Present habitat amount generally had weak positive effects on lichen species density. Positive effects of the past habitat amount were stronger, indicating a time lag in habitat amount effects. Habitat fragmentation effects were generally weak and similar whether fragmentation was measured in the past or the present landscapes, indicating no time lag in fragmentation effects.
Conclusions
We found a time lag effect of habitat amount, but not fragmentation. This result is not consistent with suggestions that time lags explain the mixed observations of fragmentation effects. Time-lag effects of habitat amount suggest that the studied lichen communities face an extinction debt. Conservation should therefore prioritize increasing the amount of old forest, for example by creating forest reserves, to maintain the current lichen diversity. More generally, our results imply that studies examining only the present habitat amount risk under-estimating its importance.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Habitat loss is one of the main causes of the ongoing rapid decline in biodiversity around the globe (Tilman et al. 2017; Díaz et al. 2019). Habitat loss is often associated with habitat fragmentation, i.e. the breaking of continuous habitat into smaller patches. While the negative effects of habitat loss on biodiversity are clear, the effects of fragmentation per se (i.e. habitat fragmentation independent of habitat amount) are still debated (Miller-Rushing et al. 2019). Fragmentation is often considered to have negative effects on biodiversity (e.g. Haddad et al. 2015; Fletcher et al. 2018; Betts et al. 2019), but recent reviews have found that after the effects of habitat amount have been accounted for, fragmentation effects are mainly insignificant and, when they occur, often positive (Fahrig 2017; Watling et al. 2020). Several authors have proposed explanations for this inconsistency (e.g. Ewers and Didham 2006). One suggested reason is confounding temporal effects, specifically time lags in species responses to landscape change (e.g. Miller-Rushing et al. 2019).
Time lags are known to occur in response to habitat loss: often, species do not disappear immediately after habitat in the surrounding landscape is lost, but can persist in the remaining habitat for decades or even centuries until finally going extinct, a so-called extinction debt (Tilman et al. 1994; Kuussaari et al. 2009). Similarly, after habitat restoration, there is often a time lag before species begin to colonize the newly created habitats (e.g. Watts et al. 2020). Therefore, especially in landscapes that have recently undergone changes in habitat amount, the species communities may reflect the past rather than present landscape composition. Studies that examine only the present landscape composition and fail to account for potential time lags can therefore risk underestimating the effects of habitat amount. Time lags have been found to occur in various ecosystems and species groups (Kuussaari et al. 2009). Species traits, most importantly longevity and dispersal capacity, can affect the probability of a time lag, as well as its length. Time lags are more likely for longer-lived species with longer generation times, since these are typically slow to respond to changes in the surrounding landscape (e.g. Watts et al. 2020). Regarding dispersal capacity, poor dispersers typically respond to landscape changes more slowly than good dispersers and are therefore suggested to be more likely to show time lags (e.g. Lira et al. 2019).
While the existence of time lags is well known for habitat loss, it is unclear whether there are also time lags in response to habitat fragmentation per se. Time lags in fragmentation effects have been suggested as one explanation for the inconsistent results of fragmentation effects. Specifically, some suggest that negative fragmentation effects would occur with a time lag, and therefore studies that focus only on the present landscape configuration would fail to detect these negative fragmentation effects (Soulé and Simberloff 1986; Ewers and Didham 2006; Miller-Rushing et al. 2019). However, there is so far no empirical support for this idea. Comparisons across studies find that negative fragmentation effects on species richness are not more common in studies where the pattern of habitat has been in place for longer (Fahrig 2020; Riva and Fahrig 2023), suggesting the absence of time lags. To our knowledge, only two empirical studies have tested for time lag effects of fragmentation per se by comparing current ecological responses to fragmentation per se in present vs. past landscapes. Semper-Pascual et al. (2021) studied birds and mammals in the Argentinian Chaco and found lag responses to habitat loss, but when effects of habitat loss were statistically controlled, they found no lag response to habitat fragmentation per se. Similarly, Herrero-Jáuregui et al. (2022), also working in the Argentinian Chaco, observed time lags for the effects of habitat amount, but not for fragmentation per se on tree species diversity, community composition, and community traits. Similar studies from other regions and taxa are required to determine whether the lack of time lag effects of habitat fragmentation per se observed in these studies is a general pattern.
Here we examine the effects of habitat amount and fragmentation per se, and the possible time lags in these effects, on deadwood-dwelling lichens in old forests in northern Sweden. The amount of suitable habitat in the surrounding landscape is known to have a positive effect on lichen species richness (e.g. Paltto et al. 2006; Randlane et al. 2017; Kärvemo et al. 2021mäläinen et al. 2023). This is because more habitat in the surrounding landscape means both a larger species pool due to the species-area relationship, and higher colonization rates, which lead to higher local species richness (Fahrig 2013). In comparison, the effects of fragmentation per se are less well known. Fragmentation may affect forest-dwelling lichens through edge effects: increased light levels and higher wind speeds near forest edges can have negative impacts on lichens, although the response is species-specific (e.g. Kivistö and Kuusinen 2000; Hilmo and Holien 2002; Esseen 2019) and can vary among different forest types. Moreover, fragmentation can affect habitat connectivity and thereby lichen dispersal: in more fragmented landscapes, distances between habitat patches are typically shorter, which may facilitate dispersal between patches (Galán-Acedo et al. 2024). Since several forest-dwelling lichens are assumed to be dispersal-limited (e.g. Ellis 2012), they should benefit from increased connectivity. However, empirical evidence on fragmentation effects on lichens is lacking. While several studies have examined the effects of habitat connectivity (e.g. Johansson et al. 2013), habitat patch size, and isolation (e.g. Esseen 2019), only one study has examined how landscape-scale fragmentation affects lichen species richness (Ellis and Coppins 2007). In that study, species richness was determined mainly by habitat amount, while the effects of fragmentation were statistically insignificant once the effect of habitat amount was accounted for.
Lichens are a taxon for which we might expect time-lag effects. They are long-lived and sessile, and can persist on their substrate (e.g. dead wood) for a long time after changes in the surrounding landscape take place. Previous studies have observed time lag responses to habitat amount of lichens on old oaks (Ranius et al. 2008; Johansson et al. 2013), in old beech forests (Paltto et al. 2006), and in old aspen forests (Ellis and Coppins 2007). However, previous studies have not evaluated time lags in response to habitat fragmentation per se; they either focused on habitat amount only (e.g. Paltto et al. 2006) or used landscape measures that combined habitat amount with habitat configuration (e.g. Johansson et al. 2013). Thus, it is unclear whether there are time-lag effects of fragmentation per se on lichens.
We study these questions by surveying deadwood-dwelling lichens in old-growth forests in two regions in northern Sweden. Using old forest as a proxy for lichen habitat, we quantify the habitat amount and fragmentation per se in the landscapes surrounding survey sites in two time periods, close to the time of the survey (2010) and ca. 60 years before the survey, in the 1960s. If the past habitat amount or fragmentation explains lichen species density better than the present, it would indicate a time lag. We expect to observe time lags because the amount of old forest (i.e. lichen habitat) has decreased greatly in northern Sweden since the early 20th century, when modern forestry with extensive clear-cutting became common (e.g. Kivinen et al. 2012; Svensson et al. 2019). Because extensive clear-cutting started several decades later in the northern region than the southern region, we expected that time lags should be more likely in the northern region, where lichen assemblages have had less time to react to the loss of old forest. Therefore, we examine time lags separately for the two regions.
Finally, we consider lichen traits that have been suggested to influence their sensitivity to habitat amount and fragmentation, and the probability of time lags. These are degree of habitat specialization and dispersal capacity. Time lags are suggested to be more likely for species with narrower habitat niches (e.g. Kuussaari et al. 2009; Sverdrup-Thygeson et al. 2014; Lira et al. 2019) and for species with poor dispersal capacity (e.g. Lira et al. 2019), although the exact mechanisms that should cause these differences are unclear. We assess the importance of niche width by comparing red-listed and non-red-listed lichen species; red-listed species are assumed to have narrower niches. Specifically, many of the red-listed deadwood-dwelling lichens require old, hard dead wood (Santaniello et al. 2017; Larsson Ekström et al. 2023) while the non-red-listed species can typically use a wider variety of dead wood substrates. In addition, we assess the importance of dispersal capacity by comparing lichen species that disperse via vegetative propagules and those that disperse via spores; species dispersing via vegetative propagules are assumed to have more limited dispersal (e.g. Ellis 2012).
Specifically, we aim to answer the following questions:
-
1.
Do landscape habitat amount and fragmentation per se affect lichen species density, and are there time lags in these effects, i.e. does the past landscape structure explain species density better than the present?
-
2.
Are red-listed species and vegetatively-dispersing species more likely than non-red-listed species and spore-dispersing species (respectively) to show responses to habitat amount and fragmentation per se and time lags in those responses?
Method
Study sites and data collection
We surveyed deadwood-dwelling lichens and associated, non-lichenized fungi (hereafter collectively referred to as “lichens”) in two regions in northern Sweden (Fig. 1). The northern region was located in the northern boreal vegetation zone (Ahti et al. 1968) in the county of Norrbotten, and the southern region in the middle boreal vegetation zone in the counties of Västerbotten, Västernorrland, and Jämtland. Both regions are dominated by coniferous forests, with about 15–20% non-forested area consisting mainly of open mires and lakes, with some smaller settlements and agricultural fields. A majority of the forests are managed by rotation forestry (i.e., clear-cutting). In northern Sweden, clear-cutting became the dominant management method in the beginning of the 20th century: before that, the forests were managed mainly by selective harvesting (e.g. Esseen et al. 1997; Östlund et al. 1997). The use of clear-cutting first began in the southern part of the country, from which it expanded northwards. Therefore, we expected the southern region to have a longer history of intensive, clear-cutting-based forestry than the northern region. This expectation was supported by data from the Swedish National Forest Inventories (NFI) conducted during the 1920s and 1950s (Swedish NFI 1925, 1955). These data showed that both in the 1920s and 1950s, the southern region had a lower proportion of old forests (> 120 years) and clear-cuts than the northern region (21% and 49% of the total productive forest area in southern and northern region, respectively). In addition, the proportion of young forests (< 20 years) was higher in the southern (13%) than the northern region (8%).

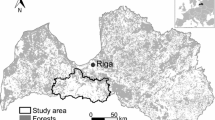
Map showing the locations in Sweden of the study sites where deadwood-dwelling lichens were surveyed
We surveyed deadwood-dwelling lichens in 2020 in 63 forest stands (33 in the northern and 30 in the southern region). All of the stands were old, unmanaged forests dominated by Scots pine (Pinus sylvestris L.). The stands were either woodland key habitats (hereafter WKHs) or forest reserves. WKHs are forest stands that are not formally protected, but classified as important for forest biodiversity and rarely harvested. The WKHs selected for this study were located on land owned by the forest companies Sveaskog AB and SCA, and had been identified in field inventories carried out by these companies. Permissions for the lichen surveys were obtained from the companies and, in the case of forest reserves, from the county administration boards of Västerbotten and Västernorrland.
To examine the effects of habitat amount and fragmentation on lichen species density, we chose stands of varying sizes (1.7 to 101.7 ha, median 11.2 ha), located in landscapes with different amounts of old forest (> 100 years old; used as a proxy of habitat amount, see “Landscape variables”). The forests in the landscapes were dominated by Scots pine: > 50% of the tree volume in the landscape within 1 km from the study stands was Scots pine. The minimum distance between any two stands was 5 km (median distance between two nearest neighbors 6.5 km, maximum 29.8 km).
In each stand, we surveyed deadwood-dwelling lichens from 16 dead Scots pine trees, including eight standing dead trees and eight fallen trees. The trees were selected by choosing four locations within each stand: the first location was chosen randomly, and the other three so that the distance between the locations was approximately similar (ca. 100 m) in all stands, despite variation in stand size. We then surveyed the two standing dead pines and two fallen dead pines closest to each location. Only decorticated dead trees that were not yet overgrown by vascular plants and bryophytes were chosen for the survey. We recorded the diameter, height, and decay stage (on a 5-point scale) of each surveyed tree. For each tree, a standardized area of 0.62 m2 was surveyed for lichens. This area was located within the lowest 2 m of each tree. This was the height to which standing dead trees could be surveyed without climbing them and, for comparability, the fallen trees were surveyed in a similar way. Thus, a standardized dead wood area of 9.92 m2 was surveyed in each stand. We recorded only lichen species considered as obligately lignicolous (following Spribille et al. 2008). When necessary, samples of lichen specimens were collected for laboratory identification using microscopy and chemical spot tests. Nomenclature of lichens followed Jääskeläinen et al. (2015).
We measured the amount of dead wood in the study stands using a line intersect method. We placed four 75 m long transects in each stand, starting from each of the four lichen survey locations, and measured dead wood along these transects. In some cases, the length or number of the transects had to be adjusted due to the size or shape of the stand, but the total length was always the same, i.e. 300 m. For fallen dead wood, we measured the diameter of each fallen tree that crossed the transect and calculated their total volume according to the formula by Marshall et al. (2000). For standing dead wood, we included all trees located within 10 m from the transects (i.e. within four 20 × 75 m rectangles). We measured the diameter of each standing tree, and used the formula by Laasasenaho (1982) to calculate their volume. In addition, we recorded the tree species and decay stage of each dead tree.
Landscape variables
We calculated habitat amount and fragmentation in the landscapes surrounding the study stands using ArcMap 10.8 (Esri Inc. 2021). We defined habitat as old forest (> 100 years), since our previous study found it to explain the species density of deadwood-dwelling lichens better than other tested variables (Hämäläinen et al. 2023). To examine the potential time-lag in lichen responses to changes in the landscape, we calculated the habitat amount and fragmentation in two periods: at the time of the lichen survey (present landscape) and in the 1960s (ca. 60 years before the lichen survey; past landscape). We calculated all landscape variables at 11 different scales: 500 m, 1 km, 2 km, 3 km etc., up to 10 km from the centers of the study stands.
In the present landscapes, we defined habitat as forest over 100 years old. This could include both protected forests as well as old forests on managed forest land. Independent of the protection status, the old forests had most likely never been clear-cut. In addition, they typically contain more dead wood than younger forests (e.g. Fridman and Walheim 2000) and can therefore function as a proxy for deadwood-dwelling lichen habitat (Hämäläinen et al. 2023). Data on forest age were obtained from the SLU Forest Map, which is produced from a combination of satellite data and field data from the Swedish National Forest Inventory. These data were from the year 2010 (the most recent data available for the studied regions).
To quantify the area and fragmentation of old forest in the 1960s, we used old aerial photos that are freely available from the National Land Survey (Lantmäteriet). The aerial photos have been scanned and projected into an orthogonal map projection, and are thus ready to be used in map analyses. In the southern region, the photos were taken from 1958 to 1965, and in the northern region from 1958 to 1960. To calculate the area of old forest, we first excluded all non-forested areas, such as water and agricultural fields. We did this by first using modern land cover data (from the Swedish National Land Cover Database, data from 2018) to divide the landscape into forested and non-forested areas. We then compared the land cover data with the old aerial photos, and manually corrected any areas that had changed from forested to non-forested or vice versa (such changes were relatively few in the studied landscapes). This way, we were able to construct a map of the extent of forested areas in the 1960s. After this, we manually digitized all clear-cuts and young forests from the aerial photos and excluded them to calculate the area of old forest.
In both present and 1960s landscapes, we calculated habitat amount as the proportion of the landscape consisting of old forests. In the present landscape, we used two measures of habitat fragmentation: the number of old forest patches, and edge density calculated as the length of old forest edges divided by landscape area (m/ha). This measurement does not consider the width of the edges. We also measured the size of the focal habitat patch where the lichens were surveyed, i.e. the size of the WKH or forest reserve. In the 1960s landscapes we used only edge density as our measure of fragmentation, as these landscapes contained much more old forest, and so individual forest patches were generally not distinguishable. In addition, we calculated the total forest area (both managed and unmanaged, of all ages) in the landscapes to account for possible differences in matrix quality. We assumed that non-forested areas were lower quality matrix than young forests because non-forested areas would contain no dead wood at all, whereas young forests might contain small amounts of dead wood that could function as lichen habitat, at least for some of the studied species. The total forest area in the present landscapes was obtained from the Swedish National Land Cover Database, and the total forest area in the 1960s landscapes from the combination of land cover data and aerial photos described above. In the 1960s landscapes, the forests in the matrix (i.e. those forests that were not classified as old) were all clear-cuts or relatively young forests (ca. < 20 years). In the present landscapes, the age of the forests in the matrix varied. This may affect matrix quality and fragmentation effects: specifically, edge effects can be expected to be stronger when an old forest stand is surrounded by clear-cut or young forest (ca. < 20 years) than when it is surrounded by mature managed forest. Therefore, we calculated the proportions of < 20 years old and 20–100 years old forest in the studied landscapes (Table S1).
Statistical analyses
All statistical analyses were conducted with R 4.3.1 (R Core Team 2023). We ran all analyses separately for the total species density of deadwood-dependent lichens and for four different groups of deadwood-dependent lichens: red-listed species, non-red-listed species, spore-dispersing species, and vegetatively-dispersing species (i.e. species dispersing mainly by isidia, soredia, or thallus fragments). Species that frequently disperse by both spores and vegetative propagules were included in the spore-dispersing group, as we assumed that the maximum dispersal distance (by spores) would determine their responses to landscape change. The red-listed species were those assessed as threatened or near-threatened in the latest Swedish Red List (Swedish Species Information Centre 2015). The species groups are presented in Table S2. Lichen species density refers to the number of species observed across the four sample sites in each stand, i.e. across all surveyed dead wood (9.92 m2) in each stand.
We constructed species accumulation curves to examine whether the sample size of 16 trees per study stand was sufficient to capture the deadwood-dwelling lichen species present at the stands. We did this separately for each study stand, plotting the number of observed lichen species as a function of the number of surveyed trees, using the package iNEXT (Hsieh et al. 2016).
To choose the best scale for examining the effects of the landscape variables, we calculated Pearson’s correlation coefficients between lichen species density and (i) the amount of habitat (% old forest) and (ii) fragmentation of old forest (edge density or number of patches), in the landscapes at each of the 11 scales (500 m – 10 km radius). The scales at which the correlations were strongest for each of the two predictors were then used in the following analyses, i.e. we chose the best scale separately for habitat amount and fragmentation. In addition, we chose these scales separately for each of the abovementioned species groups, and for the present and past habitat amount and fragmentation.
We used generalized linear models (GLMs) to examine the effects of habitat amount and fragmentation on lichen species density in the study stands (N = 63). We constructed separate models for each of the lichen groups and for the present and past landscapes. In addition, we constructed two different models for the present landscapes, using two different measures of fragmentation (edge density or number of patches). Thus, we ran three different models for each species group, 15 models in total. In all of the models, we used lichen species density (one of the five species groups) in the study stands as a response variable. The predictor variables are given in Table 1. All continuous predictor variables were standardized (mean = 0, SD = 1) prior to analyses to enable comparison of model coefficients. Especially at the larger scales in present landscape, habitat amount and edge density were correlated (Tables S3-4). However, we do not consider this problematic, because it should not cause bias in the model coefficients (Smith et al. 2009; Morrissey and Ruxton 2018). We included study region as a categorical variable (north/south). In addition, we included interaction terms between region and habitat amount, as well as between region and fragmentation.
We ran the GLMs using a Poisson distribution and log link. In some cases, the data were found to be under-dispersed, and to correct for this, we ran the models with a COM-Poisson distribution that can handle both under- and over-dispersed data (Brooks et al. 2019). We used COM-Poisson in all models for vegetatively-dispersing species, and in the model for the total species density using past landscape variables. The Poisson GLMs were run using the package lme4 (Bates et al. 2015) and the COM-Poisson GLMs with the package glmmTMB (Brooks et al. 2017). We assessed model fit by plotting residuals using the package DHARMa (Hartig 2022). In addition, we tested for residual spatial autocorrelation by calculating Moran’s I and by constructing variograms on the residuals using the package gstat (Gräler et al. 2016). No evidence of spatial autocorrelation was found in any of the models. Finally, we plotted model results using packages coefplot (Lander 2022), ggplot2 (Wickham 2016), and ggeffects (Lüdecke 2018).
Results
We found 28 species of deadwood-dependent lichens: 25 in the northern and 27 in the southern region (Table S2). One species was only found in the northern region, and three species only in the southern region. The mean species density was 14.5 species per 9.92 m2 dead wood, i.e. the area of dead wood surveyed per study stand. Eight of the observed species were red-listed (23 of the 28 species had been assessed for a red list classification), and six dispersed predominantly by vegetative propagules (isidia, soredia, or thallus fragments). According to the species accumulation curves, almost all species present in the study stands were detected across the 16 surveyed trees per stand (Figure S1).
The best scales for analyzing the landscape variables differed among variables and lichen groups, and between the present and past landscapes (Fig. S2). The best scale for analyzing habitat amount in the present landscape was 500 m for red-listed species, 1 km for species dispersing by vegetative propagules, and 6 km for total species density, non-redlisted species, and spore-dispersing species. The best scale for analyzing edge density in the present landscape was 500 m for total species density and for red-listed species, 1 km for vegetatively dispersing species, 6 km for non-red-listed species, and 7 km for spore-dispersing species. The best scale for analyzing number of patches was 2 km for vegetatively dispersing species, and 9 km for the other groups. The best scale for analyzing habitat amount in the past landscape was 3 km for all of the studied species groups except for non-redlisted species, for which it was 4 km. For analyzing edge density in the past landscapes, the best scales were 500 m for non-red-listed and spore-dispersing species, 2 km for red-listed species, 3 km for the total species density, and 10 km for vegetatively dispersing species. The means and ranges of the landscape variables at each scale are given in Tables S5-6. At the scales used in the models, correlations between the landscape variables were generally low to moderate (Tables S3-4). An exception was a high correlation between habitat amount (% of old forest) and edge density in the present landscapes at scales of 1 km or larger (r > 0.7).
Two different measures were used to quantify habitat fragmentation in the present landscapes: edge density and number of patches. These yielded similar results in the GLMs. Thus, we present here the results of GLMs in which edge density was used, as these are comparable with the GLMs for past landscapes, which used edge density. Results of the GLMs run with the number of patches are presented in Figures S5–9.
Do landscape habitat amount and fragmentation per se affect total lichen species density, and are there time lags in these effects?
For habitat amount, there was an indication of a time lag in the southern region. The total species density was positively related to past habitat amount (i.e. measured in the 1960’s), while there was no evident relationship to current habitat amount (Fig. 2).The relationship between habitat fragmentation and total species density was very weak for fragmentation measured in both time periods (Fig. 2), suggesting no lag effect.

The effects of landscape and stand variables on the total species density of deadwood-dependent lichens. On the left, (a) estimated model coefficients (+ 95% CI); on the right, the predicted lichen species density (+ 95% CI), in (b) the northern and (c) the southern regions, as a function of % old forest in the surrounding landscape and as a function of edge density (m/ha of old forest edge in the surrounding landscape). Separate models were constructed for the present landscape (2010, orange) and the past landscape (1960, green). In the present landscapes, old forest area was measured within 6 km and edge density within 500 m of the centers of the study stands. In the past landscapes, old forest area and edge density were both measured within 3 km
Are red-listed species and vegetatively-dispersing species more likely than non-redlisted species and spore-dispersing species (respectively) to show responses to habitat amount and fragmentation, and time lags in those responses?
None of the past or present landscape variables were found to affect species density of non-redlisted species (Fig. S3). In contrast, in the southern region, present habitat amount had a very weak positive effect on species density of red-listed lichens, while past habitat amount had a strong positive effect (Fig. 3). This indicates a time lag in effects of habitat amount on red-listed species in the south. Effects of fragmentation on red-listed species were slightly positive in the south, with no suggestion of a time lag (Fig. 3).

The effects of landscape and stand variables on the species density of red-listed lichens. On the left, (a) estimated model coefficients (+ 95% CI); on the right, the predicted lichen species density (+ 95% CI), in (b) the northern and (c) the southern regions, as a function of % old forest and edge density in the surrounding landscape. In the present landscapes, old forest area and edge density were measured at 500 m of the centers of the study stands. In the past landscapes, old forest area was measured within 3 km and edge density within 2 km. See Fig. 2 for further explanations
There was no evidence that any of the past or present landscape variables affected spore-dispersing lichens (Fig. S4). In contrast, in the north, vegetatively-dispersing lichen species density was positively related to both past and present habitat amount. In the south there was a time lag in the effect of habitat amount on vegetatively-dispersing lichen density, with a weak negative effect of present habitat amount and a positive effect of past habitat amount (Fig. 4). The effect of fragmentation on vegetatively-dispersing lichens was negative in the north and positive in the south, with no indication of time lags (Fig. 4).

The effects of landscape and stand variables on the species density of vegetatively dispersing lichen species. On the left, (a) estimated model coefficients (+ 95% CI); on the right, the predicted lichen species density (+ 95% CI), in (b) the northern and (c) the southern regions, as a function of % old forest and edge density in the surrounding landscape. In the present landscape, old forest area and edge density were measured within 1 km of the centers of the study stands. In the past landscapes, old forest area was measured within 3 km and edge density within 10 km. See Fig. 2 for further explanations
Discussion
The effects of landscape habitat amount on the species density of deadwood-dwelling lichens occurred with a time lag, while there was no time lag in the effects of fragmentation per se. This latter result is not consistent with suggestions that lag effects explain the variation in observed responses to fragmentation per se. Time-lagged responses to habitat amount were found for red-listed and vegetatively dispersing lichen species, but not for non-red-listed and spore-dispersing species.
Time lags
In accordance with our hypothesis, we observed a time lag for the effect of habitat amount. The past habitat amount had a stronger positive effect on lichen species density than the present habitat amount, especially for red-listed lichens. This has been observed before for lichens on living trees (e.g. Ellis and Coppins 2007; Ranius et al. 2008; Johansson et al. 2013). On the contrary, the effects of fragmentation did not occur with a time lag; the present fragmentation had minor effects on lichen species density, while we did not observe any effect of past fragmentation. Thus, in accordance with previous empirical studies (Semper-Pascual et al. 2021; Herrero-Jáuregui et al. 2022), we did not find support for the hypothesis that negative fragmentation effects should occur with a time lag (e.g. Soulé and Simberloff 1986). This does not support the notion that the varying findings regarding the effects of fragmentation are explained by the occurrence of time lags.
Our results suggest that deadwood-dwelling lichens, especially red-listed ones, occurring in the studied WKHs and nature reserves face an extinction debt, as the amount of old forest (i.e. lichen habitat) has decreased significantly in recent decades in the studied regions. For example, Svensson et al. (2019) estimated that between the 1970s and 2014, the proportion of old forest that had never been clear-cut decreased from 75 to 38% in inland areas of northern Sweden. Decreases of similar magnitude occurred in our study landscapes between the 1960s and 2010 (Tables S5-6). Given the observed time lag for habitat amount, the effects of these decreases on lichen species density may not have been realized yet. Extinction debts of lichens in WKHs have also been suggested by Berglund and Jonsson (2005), who compared WKHs in landscapes with different times since habitat removal and found indications of an extinction debt for lichens occurring on living trees, but not for deadwood-dwelling fungi. Our results suggest that even deadwood-dwelling lichen species face an extinction debt. Therefore, the WKHs and reserves in landscapes with low old forest cover will not be able to support their current lichen diversity into the future, unless the amount of old forest in the surrounding landscapes is increased.
We were able to assess the past landscape structure approximately 60 years before the lichen surveys. It is possible that the time lags might be even longer than this. For example, Johansson et al. (2013) observed a 180 year lag for lichens on old oaks, while Ellis and Coppins (2007) found a ca. 120 year lag for lichens in aspen forests. However, these studies focused on lichens on living trees, which are typically a more long-lived substrate than dead wood. Oaks, for example, can function as lichen habitat for several hundred years (Drobyshev and Niklasson 2010), and although dead wood of Scots pine can take around 200 years to decay in northern boreal forests (e.g. Shorohova and Kapitsa 2015), fallen dead wood is typically overgrown by ground flora and therefore becomes unsuitable for most deadwood-dwelling lichens much earlier than this. Thus, we might expect shorter time lags for lichens occurring on dead wood substrates than for those occurring on living trees. In addition to the longevity of the substrate itself, lichen species dynamics might be different on old living trees than on dead wood. Stochastic extinctions of lichens on living trees are typically very rare and the lichens tend to remain until tree death (e.g. Johansson et al. 2012). Lichen dynamics on dead wood are less studied, but there are indications that stochastic extinctions may be relatively frequent (Caruso et al. 2010). If this is the case, it would also contribute to shorter time lags for species occurring on dead wood in comparison to those on living trees.
We had expected to find stronger evidence for time lags in the more northern region than in the more southern region, but we found the opposite. We have no clear explanation for this finding; the decrease in old forest area was relatively similar in the two regions, as was the lichen species composition. Overall, the effects of the present landscape were also less pronounced in the northern region, which could imply that the definition of habitat (old forest) was less accurate in this region, but we have no other evidence supporting that suggestion. Another potential explanation is that in the northern region, dead wood amounts on the studied stands were generally higher, thus, the landscape structure might in general be less important in this region, if the local habitat amount was sufficiently high to ensure the presence of most lichen species.
Effects of habitat amount, fragmentation, and stand-scale variables
The present amount of habitat, i.e. old forest, in the landscape had in most cases a positive effect on the species density of deadwood-dwelling lichens. However, these effects were rarely significant, and typically quite weak in comparison to previous studies that have found lichen diversity to increase with landscape habitat amount in boreal (Kärvemo et al. 2021; Hämäläinen et al. 2023) and temperate forests (Paltto et al. 2006). Moreover, in some cases, the present amount of habitat in the southern study region had a negative (though weak) effect on lichen species density. These weak and even negative effects are most likely due to the time lag, i.e. the species density is determined by the past rather than present landscape habitat amount. This implies more generally that studies that examine only present landscape structure risk under-estimating the importance of habitat amount, particularly if the studied species are prone to time lags. In our study system, this would especially be the case for red-listed species; the amount of habitat in the landscapes had a strong effect on them, but this effect would have been missed if only the habitat amount in the present landscapes had been examined.
The effects of present fragmentation were weak and in most cases positive: an exception was vegetatively-dispersing species in the southern region. The pine-dominated old forests that we studied had relatively open stand structure, and edge effects in the form of altered light availability or microclimate are therefore likely moderate. The effects of fragmentation per se have rarely been studied for lichens or other cryptogams, but the few existing studies have found negligible effects of fragmentation (Ellis and Coppins 2007; Sundberg 2013) once habitat amount is accounted for. This is in accordance with our results and with studies of other taxa (Fahrig 2017). In our study landscapes, the matrix consisted mainly of managed forests. In the past landscapes, these forests were all young (ca. < 20 years), but in the present landscapes the ages of the forests varied (Table S1). This could cause variation in the edge contrast between the old forests and the surrounding matrix, which might weaken observed edge effects. Edge effects can be assumed to be weaker when an old forest stand is surrounded by mature managed forest than when it is surrounded by clear-cut or young forest. However, this should only be an issue in the present landscapes, because all managed forests in the past landscapes were young. This implies that if variation in edge contrast led to weaker edge effects, we should observe a stronger effects of past than present fragmentation, which we did not find. Moreover, we should observe stronger effects of present fragmentation in the southern study region, where the proportion of young (< 20 years) forest was higher. Again, we did not find such a difference, which implies that variation in edge contrasts does not notably affect our results. Nevertheless, we note that our results may not be generalizable to landscapes where the matrix is more hostile, e.g. agricultural fields instead of the managed forests. Future studies are thus needed to examine whether our results apply to other types of landscapes. In addition, we note that habitat amount and fragmentation (edge density) were correlated in the studied landscapes, especially at the larger scales. This may decrease the likelihood of obtaining statistically significant results, but should not cause a bias in the model coefficients (Smith et al. 2009; Morrissey and Ruxton 2018). In addition, we obtained similar results when measuring fragmentation as patch density (Fig. S5-S9), which was not strongly correlated with habitat amount.
In addition to landscape variables, the focal patch area, i.e. the area of the studied WKH or reserve, and the amount of dead wood within the stand had positive effects on lichen species density in the southern region, even when the landscape habitat amount was taken into account. This is in contrast with the habitat amount hypothesis (Fahrig 2013), which predicts that the focal patch area should not affect species density within a sample area, when the landscape habitat amount has been accounted for. Furthermore, it is in contrast with a previous study that did not observe any effects of local dead wood amount on lichen species density in old pine forests (Hämäläinen et al. 2023). Our result implies that larger patches of old forest can maintain higher local species density, at least in some regions. In the northern region, neither focal patch area nor dead wood amount affected species density, which may again be due to the generally higher dead wood amounts in the northern region, potentially sufficient to support the occurrence of most species even in smaller patches.
Functional traits
We found a time lag in the effects of habitat amount on red-listed species. Their species density was not affected by the present habitat amount, but increased with the past habitat amount. On the contrary, the density of non-red-listed species was not affected by present or past habitat amount. This is in accordance with the prediction that time lags would be more likely for habitat specialists (Kuussaari et al. 2009; Lira et al. 2019), which the red-listed species are expected to be. We assume that the observed stronger effect of past habitat amount on red-listed than non-red-listed species was not due to a difference in the time lag itself, but simply to stronger effects of landscape habitat amount in general on red-listed species. Red-listed lichen species are typically more specialized to old forest habitats (Nirhamo et al. 2023), while non-red-listed species are more likely to occur even in the matrix (young forests) if suitable dead-wood substrates are available. Therefore, the amount of old forest will have a stronger effect on red-listed species than non-red-listed species. This has also been observed for other deadwood-dwelling taxa, e.g. saproxylic fungi (e.g. Nordén et al. 2013; Sverdrup-Thygeson et al. 2014).
When comparing lichens with contrasting dispersal modes, we found that the density of vegetatively-dispersing species was affected by both present and past landscape structure, and there was an indication of a time lag for habitat amount. In contrast, spore-dispersing species were not significantly affected by the landscape structure, and there were no time lags. Vegetative propagules are larger than spores, and vegetatively-dispersing species are therefore assumed to be poorer dispersers: in modelling studies, lichens with larger propagules have been found to have smaller dispersal ranges (Ruete et al. 2014) and lower colonization rates (Johansson et al. 2012) than species with smaller propagules. Vegetatively-dispersing lichens have also been found to be more affected by the landscape habitat amount than spore-dispersing lichens (Ellis and Coppins 2007; Hedenås and Ericson 2008). Moreover, poor dispersers have been suggested to be more likely to show time lags (Lira et al. 2019). However, it is not clear whether our result supports this suggestion. Instead, as discussed above for non-red-listed species, the apparent lack of time lag effects on spore-dispersing species may actually be due to generally weak effects of landscape structure on them.
Conclusions
We found that the effects of landscape habitat amount on lichen species density occurred with a time lag, while the effects of fragmentation did not. Instead, any fragmentation effects were temporary and disappeared within the studied 60-year period. This is not consistent with suggestions that time lags are the reason for mixed effects of habitat fragmentation on biodiversity. If our results can be generalized, they imply that studies that examine only the present landscapes may underestimate the effects of habitat amount In our study, the models that included only present landscape variables did not find as strong positive relationship between habitat amount and lichen species density as the models with past landscape variables. Most notably, the strong positive effect of landscape habitat amount on the density of red-listed lichen species in the south would have gone unnoticed if only responses to the present habitat amount had been analyzed.
Our results imply that the deadwood-dwelling lichens in the studied WKHs and nature reserves face an extinction debt, as has been suggested also for lichens on living trees (Berglund and Jonsson 2005). Several studies have questioned the capacity of the WKHs and reserves to maintain species diversity over long timescales (e.g. Aune et al. 2005; Jönsson et al. 2017), and our results support this concern. Thus, to maintain the lichen diversity that the WKHs and reserves currently host, the amount of old forest in the surrounding landscapes should be increased, for example by creating new forest reserves. This is especially important for WKHs and reserves with currently little old forest in their surrounding landscapes. Our results further suggest that habitat fragmentation – in both present and past landscapes – has only minor effects on lichen species density. Lichen conservation should therefore prioritize maintaining or increasing habitat amount instead of minimizing fragmentation.
Data availability
Should this manuscript be accepted, we will deposit the data in a publicly available repository (e.g. in Dryad, www.datadryad.org).
References
Ahti T, Hämet-Ahti L, Jalas J (1968) Vegetation zones and their sections in northwestern Europe. Ann Bot Fenn 5:169–211
Aune K, Jonsson BG, Moen J (2005) Isolation and edge effects among woodland key habitats in Sweden: is forest policy promoting fragmentation? Biol Conserv 124:89–95. https://doi.org/10.1016/j.biocon.2005.01.015
Bates D, Maechler M, Bolker B, Walker S (2015) Fitting linear mixed-effects models using lme4. J Stat Softw 67:1–48. https://doi.org/10.18637/jss.v067.i01
Berglund H, Jonsson BG (2005) Verifying an extinction debt among lichens and fungi in northern Swedish boreal forests. Conserv Biol 19:338–348. https://doi.org/10.1111/j.1523-1739.2005.00550.x
Betts M, Wolf C, Pfeifer M et al (2019) Extinction filters mediate the global effects of habitat fragmentation on animals. Science 366:1236–1239. https://doi.org/10.1126/science.aax9387
Brooks ME, Kristensen K, van Benthem KJ et al (2017) glmmTMB balances speed and flexibility among packages for zero-inflated generalized linear mixed modeling. R J 9:378–400. https://doi.org/10.32614/RJ-2017-066
Brooks ME, Kristensen K, Darrigo MR et al (2019) Statistical modeling of patterns in annual reproductive rates. Ecology 100:e02706. https://doi.org/10.1002/ecy.2706
Caruso A, Thor G, Snäll T (2010) Colonization–extinction dynamics of epixylic lichens along a decay gradient in a dynamic landscape. Oikos 119:1947–1953. https://doi.org/10.1111/j.1600-0706.2010.18713.x
R Core Team (2023) R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. https://www.R-project.org/
Díaz S, Settele J, Brondízio ES et al (2019) Pervasive human-driven decline of life on Earth points to the need for transformative change. Science 366:eaax3100. https://doi.org/10.1126/science.aax3100
Drobyshev I, Niklasson M (2010) How old are the largest southern Swedish oaks? A dendrochronological analysis. Ecol Bull 155–164
Ellis CJ (2012) Lichen epiphyte diversity: a species, community and trait-based review. Perspect Plant Ecol Evol Syst 14:131–152. https://doi.org/10.1016/j.ppees.2011.10.001
Ellis CJ, Coppins BJ (2007) 19th century woodland structure controls stand-scale epiphyte diversity in present-day Scotland. Divers Distrib 13:84–91. https://doi.org/10.1111/j.1366-9516.2006.00310.x
Esri Inc (2021) ArcGIS Desktop: Release 10. Redlands, CA: Environmental Systems Research Institute
Esseen P-A (2019) Strong influence of landscape structure on hair lichens in boreal forest canopies. Can J Res 49:994–1003. https://doi.org/10.1139/cjfr-2019-0100
Esseen P-A, Ehnström B, Ericson L, Sjöberg K (1997) Boreal forests. Ecol Bull 46:16–47
Ewers RM, Didham RK (2006) Confounding factors in the detection of species responses to habitat fragmentation. Biol Rev 81:117–142. https://doi.org/10.1017/S1464793105006949
Fahrig L (2013) Rethinking patch size and isolation effects: the habitat amount hypothesis. J Biogeogr 40:1649–1663. https://doi.org/10.1111/jbi.12130
Fahrig L (2017) Ecological responses to habitat fragmentation per se. Annu Rev Ecol Evol Syst 48:1–23. https://doi.org/10.1146/annurev-ecolsys-110316-022612
Fahrig L (2020) Why do several small patches hold more species than few large patches? Glob Ecol Biogeogr 29:615–628. https://doi.org/10.1111/geb.13059
Fletcher RJ, Didham RK, Banks-Leite C et al (2018) Is habitat fragmentation good for biodiversity? Biol Conserv 226:9–15. https://doi.org/10.1016/j.biocon.2018.07.022
Fridman J, Walheim M (2000) Amount, structure, and dynamics of dead wood on managed forestland in Sweden. Ecol Manag 131:23–36. https://doi.org/10.1016/S0378-1127(99)00208-X
Galán-Acedo C, Fahrig L, Riva F, Schulz T (2024) Positive effects of fragmentation per se on the most iconic metapopulation. Conserv Lett e13017. https://doi.org/10.1111/conl.13017
Gräler B, Pebesma E, Heuvelink G (2016) Spatio-temporal interpolation using gstat. R J 8:204–218
Haddad NM, Brudvig LA, Clobert J et al (2015) Habitat fragmentation and its lasting impact on Earth’s ecosystems. Sci Adv 1:e1500052. https://doi.org/10.1126/sciadv.1500052
Hämäläinen A, Fahrig L, Strengbom J, Ranius T (2023) Effective management for deadwood-dependent lichen diversity requires landscape-scale habitat protection. J Appl Ecol 60:1597–1606. https://doi.org/10.1111/1365-2664.14429
Hartig F (2022) DHARMa: residual diagnostics for hierarchical (multi-level/mixed) regression models. R Package Version 0.4.6
Hedenås H, Ericson L (2008) Species occurrences at stand level cannot be understood without considering the landscape context: cyanolichens on aspen in boreal Sweden. Biol Conserv 141:710–718. https://doi.org/10.1016/j.biocon.2007.12.019
Herrero-Jáuregui C, Camba Sans G, Andries DM et al (2022) Past and present effects of habitat amount and fragmentation per se on plant species richness, composition and traits in a deforestation hotspot. Biol Conserv 276:109815. https://doi.org/10.1016/j.biocon.2022.109815
Hilmo O, Holien H (2002) Epiphytic lichen response to the edge environment in a boreal Picea abies forest in central Norway. Bryologist 105:48–56. https://doi.org/10.1639/0007-2745(2002)105[0048:ELRTTE]2.0.CO;2
Hsieh TC, Ma KH, Chao A (2016) iNEXT: an R package for rarefaction and extrapolation of species diversity (Hill numbers). Methods Ecol Evol 12:1451–1456. https://doi.org/10.1111/2041-210X.12613
Jääskeläinen K, Pykälä J, Vitikainen O, Ahti T (2015) Suomen jäkälien luettelo. In: Stenroos S, Velmala S, Pykälä J, Ahti T (eds) Suomen rupijäkälät. Norrlinia 28:1–454
Johansson V, Ranius T, Snäll T (2012) Epiphyte metapopulation dynamics are explained by species traits, connectivity, and patch dynamics. Ecology 93:235–241. https://doi.org/10.1890/11-0760.1
Johansson V, Snäll T, Ranius T (2013) Estimates of connectivity reveal non-equilibrium epiphyte occurrence patterns almost 180 years after habitat decline. Oecologia 172:607–615. https://doi.org/10.1007/s00442-012-2509-3
Jönsson MT, Ruete A, Kellner O et al (2017) Will forest conservation areas protect functionally important diversity of fungi and lichens over time? Biodivers Conserv 26:2547–2567. https://doi.org/10.1007/s10531-015-1035-0
Kärvemo S, Jönsson M, Hekkala A-M et al (2021) Multi-taxon conservation in northern forest hot-spots: the role of forest characteristics and spatial scales. Landsc Ecol 989–1002. https://doi.org/10.1007/s10980-021-01205-x
Kivinen S, Berg A, Moen J et al (2012) Forest fragmentation and landscape transformation in a reindeer husbandry area in Sweden. Environ Manage 49:295–304. https://doi.org/10.1007/s00267-011-9788-z
Kivistö L, Kuusinen M (2000) Edge effects on the epiphytic lichen flora of Picea abies in middle boreal Finland. Lichenologist 32:387–398. https://doi.org/10.1006/lich.2000.0282
Kuussaari M, Bommarco R, Heikkinen RK et al (2009) Extinction debt: a challenge for biodiversity conservation. Trends Ecol Evol 24:564–571. https://doi.org/10.1016/j.tree.2009.04.011
Laasasenaho J (1982) Taper curve and volume functions for pine, spruce and birch. Comm Inst Fenn 108:1–74
Lander P (2022) Coefplot: plots coefficients from fitted models. R Package Version 1.2.8
Larsson Ekström A, Sjögren J, Djupström LB, Thor G, Löfroth T (2023) Reinventory of permanent plots show that kelo lichens face an extinction debt. Biol Cons 288:110363. https://doi.org/10.1016/j.biocon.2023.110363
Lira PK, de Souza Leite M, Metzger JP (2019) Temporal lag in ecological responses to landscape change: where are we now? Curr Landsc Ecol Rep 4:70–82. https://doi.org/10.1007/s40823-019-00040-w
Lüdecke D (2018) Ggeffects: tidy data frames of marginal effects from regression models. J Open Source Softw 3:772. https://doi.org/10.21105/joss.00772
Marshall P, Davis G, LeMay VM (2000) Using line intersect sampling for coarse woody debris. Technical Report TR-003, Research Section, Vancouver Forest Region, BCMOF
Miller-Rushing AJ, Primack RB, Devictor V et al (2019) How does habitat fragmentation affect biodiversity? A controversial question at the core of conservation biology. Biol Conserv 232:271–273. https://doi.org/10.1016/j.biocon.2018.12.029
Morrissey MB, Ruxton GD (2018) Multiple regression is not multiple regressions: the meaning of multiple regression and the Non-problem of Collinearity. Philos Theory Pract Biol 10. https://doi.org/10.3998/ptpbio.16039257.0010.003
Nirhamo A, Pykälä J, Jääskeläinen K, Kouki J (2023) Habitat associations of red-listed epiphytic lichens in Finland. Silva Fenn 57. https://doi.org/10.14214/sf.22019
Nordén J, Penttilä R, Siitonen J et al (2013) Specialist species of wood-inhabiting fungi struggle while generalists thrive in fragmented boreal forests. J Ecol 101:701–712. https://doi.org/10.1111/1365-2745.12085
Östlund L, Zackrisson O, Axelsson A-L (1997) The history and transformation of a scandinavian boreal forest landscape since the 19th century. Can J Res 27:1198–1206. https://doi.org/10.1139/x97-070
Paltto H, Nordén B, Götmark F, Franc N (2006) At which spatial and temporal scales does landscape context affect local density of Red Data Book and Indicator species? Biol Conserv 133:442–454. https://doi.org/10.1016/j.biocon.2006.07.006
Randlane T, Tullus T, Saag A et al (2017) Diversity of lichens and bryophytes in hybrid aspen plantations in Estonia depends on landscape structure. Can J Res 47:1202–1214. https://doi.org/10.1139/cjfr-2017-0080
Ranius T, Eliasson P, Johansson P (2008) Large-scale occurrence patterns of red-listed lichens and fungi on old oaks are influenced both by current and historical habitat density. Biodivers Conserv 17:2371–2381. https://doi.org/10.1007/s10531-008-9387-3
Riva F, Fahrig L (2023) Landscape-scale habitat fragmentation is positively related to biodiversity, despite patch-scale ecosystem decay. Ecol Lett 26:268–277. https://doi.org/10.1111/ele.14145
Ruete A, Fritz Ö, Snäll T (2014) A model for non-equilibrium metapopulation dynamics utilizing data on species occupancy, patch ages and landscape history. J Ecol 102:678–689. https://doi.org/10.1111/1365-2745.12229
Santaniello F, Djupström LB, Ranius T, Weslien J, Rudolphi J, Thor G (2017) Large proportion of wood dependent lichens in boreal pine forest are confined to old hard wood. Biodivers Conserv 26:1295–1310. https://doi.org/10.1007/s10531017-1301-4
Semper-Pascual A, Burton C, Baumann M et al (2021) How do habitat amount and habitat fragmentation drive time-delayed responses of biodiversity to land-use change? Proc R Soc B Biol Sci 288. https://doi.org/10.1098/rspb.2020.2466
Shorohova E, Kapitsa E (2015) Stand and landscape scale variability in the amount and diversity of coarse woody debris in primeval European boreal forests. Ecol Manag 356:273–284. https://doi.org/10.1016/j.foreco.2015.07.005
Smith AC, Koper N, Francis CM, Fahrig L (2009) Confronting collinearity: comparing methods for disentangling the effects of habitat loss and fragmentation. Landsc Ecol 24:1271–1285. https://doi.org/10.1007/s10980-009-9383-3
Soulé ME, Simberloff D (1986) What do genetics and ecology tell us about the design of nature reserves? Biol Conserv 35:19–40. https://doi.org/10.1016/0006-3207(86)90025-X
Spribille T, Thor G, Bunnell FL et al (2008) Lichens on dead wood: species-substrate relationships in the epiphytic lichen floras of the Pacific Northwest and Fennoscandia. Ecography 31:741–750. https://doi.org/10.1111/j.1600-0587.2008.05503.x
Sundberg S (2013) Spore rain in relation to regional sources and beyond. Ecography 36:364–373. https://doi.org/10.1111/j.1600-0587.2012.07664.x
Svensson J, Andersson J, Sandström P et al (2019) Landscape trajectory of natural boreal forest loss as an impediment to green infrastructure. Conserv Biol 33:152–163. https://doi.org/10.1111/cobi.13148
Sverdrup-Thygeson A, Gustafsson L, Kouki J (2014) Spatial and temporal scales relevant for conservation of dead-wood associated species: current status and perspectives. Biodivers Conserv 23:513–535. https://doi.org/10.1007/s10531-014-0628-3
Swedish National Forest Inventory (1925, 1955). Unpublished data. The Department of Forest Resource Management. Swedish University of Agricultural Sciences, Umeå. http://www.slu.se/nfi
Swedish Species Information Centre (2015) Red-listed species in Sweden 2015. Swedish Species Information Centre, SLU, Uppsala
Tilman D, May RM, Lehman CL, Nowak MA (1994) Habitat destruction and the extinction debt. Nature 371:65. https://doi.org/10.1038/371065a0
Tilman D, Clark M, Williams DR et al (2017) Future threats to biodiversity and pathways to their prevention. Nature 546:73–81. https://doi.org/10.1038/nature22900
Watling JI, Arroyo-Rodríguez V, Pfeifer M et al (2020) Support for the habitat amount hypothesis from a global synthesis of species density studies. Ecol Lett 23:674–681. https://doi.org/10.1111/ele.13471
Watts K, Whytock RC, Park KJ et al (2020) Ecological time lags and the journey towards conservation success. Nat Ecol Evol 4:304–311. https://doi.org/10.1038/s41559-019-1087-8
Wickham H (2016) ggplot2: elegant graphics for data analysis. Springer-Verlag, New York. https://ggplot2.tidyverse.org
Acknowledgements
We thank Anna-Lena Axelsson for providing the data from the Swedish National Forest Inventory and Edith Bremer for help with the field inventories. The study was funded by the Swedish research council Formas (grant n. 2019-01559 to AH).
Funding
Open access funding provided by Swedish University of Agricultural Sciences. The study was funded by the Swedish research council Formas (grant n. 2019 − 01559 to AH).
Author information
Authors and Affiliations
Contributions
Both authors contributed to the study conception and design as well as to the writing of the manuscript. Aino Hämäläinen conducted the field inventories and data analyses, and wrote the first draft of the manuscript. Both authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Hämäläinen, A., Fahrig, L. Time-lag effects of habitat loss, but not fragmentation, on deadwood-dwelling lichens. Landsc Ecol 39, 111 (2024). https://doi.org/10.1007/s10980-024-01910-3
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s10980-024-01910-3