
Abstract
Context
Widespread globally, roads impact the distribution of wildlife by influencing habitat use and avoidance patterns near roadways and disrupting movement across them. Wildlife responses to roads are known to vary across species; however within species, the response to roads may depend on the season or the individual’s behavioral state.
Objectives
We assess the movement behavior and space use of the most widespread large herbivore in Patagonia, the guanaco (Lama guanicoe). We estimated the preference or avoidance to paved or unpaved roads (the proximity effect) and the preference or avoidance to traverse them (the crossing effect).
Methods
Using GPS collar data, we combined Hidden Markov Models with an integrated step selection analysis to segment guanaco movement trajectories into individual behaviors and test for differences in road effects on movement.
Results
We found that guanacos display distinct movement responses to different types of roads depending on their behavioral state. Guanacos select for proximity to paved roads while foraging, but against them when traveling. Yet, guanacos select for unpaved roads when traveling. Despite the selection for proximity to paved roads, guanacos avoid crossing them, irrespective of their behavioral state.
Conclusion
Our findings offer significant implications for guanaco distribution and management across Patagonia. The selection for roads strongly influences the distribution of guanacos, which could concentrate grazing in some areas while freeing others. Despite potential benefits such as increased vegetation near roadsides, increased association with roads while foraging may result in an ecological trap. Finally, the strong aversion to crossing paved roads raises concerns about habitat loss and connectivity.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Animal movement is driven by the spatiotemporal distribution of resources as individuals search for food, acquire mates, and seek safety (Nathan 2008). However, anthropogenic changes, such as habitat loss, energy development, and climate change, are rapidly creating novel landscape conditions that impact animal’s resources (Chambers et al. 2022; Pike et al. 2023). In response to these changes, animals may need to adapt and adjust their behavior and movement patterns to survive. Human activities can also create both physical barriers, like fences and dams, as well as perceived barriers due to fear of human persecution. Barriers that impede movement can lead to a diminished capacity to track essential resources, thereby resulting in individual fitness consequences, a decline in population connectivity, and a reduced flow of nutrients across an ecosystem (Tanner and Perry 2007; Holderegger and Di Giulio 2010; Bauer and Hoye 2014). Therefore, it is crucial to understand how human activities reshape the resource landscape while also recognizing their potential to restrict animal movement.
Spanning over 21 million kilometers, roads are amongst the most widespread anthropogenic features on landscapes around the world (Meijer et al. 2018). The population impacts of roads on wildlife, like road-related mortality, are well documented. Vehicles cause 7% of mammal, 13% of reptile, and 4% of amphibian mortality worldwide (Hill et al. 2019). As a dominant feature on the landscape, roads can also influence wildlife behavior through habitat modification that drastically changes conditions like light and soil characteristics impacting animal foraging, breeding, and predator avoidance strategies (Teixeira et al. 2020; Hill et al. 2021). These changes to the resource landscape can lead to alterations in habitat selection through road avoidance or attraction. The proximate causes of road avoidance varies from species to species as some individuals may be avoiding increased noise, differences in habitat quality, or even changes to the microclimate (Ortega and Capen 1999; McClure et al. 2013). Conversely, other species may select for habitat near roads due to increasing plant productivity, refuge from predation, thermoregulation, or the facilitation of movement (Hill et al. 2021). Roads can also impact habitat selection by impeding movement as some species avoid crossing them (Brehme et al. 2013; Aiello et al. 2023). Responses to roads can vary across species, but also within the same species. Individual decisions to associate with or avoid roads may be influenced by seasonal variations or their current behavioral state (resting, foraging, or traveling), potentially reflecting the influence of distinct proximate mechanisms underlying these movement patterns. Some individuals might be attracted to roads when traveling due to a reduction in energy costs, but that same individual might avoid roads when resting due to the risk of vehicle collisions (Abrahms et al. 2015; Buxton et al. 2020). Roads undoubtedly impact wildlife, however these impacts may be more dynamic than previously appreciated.
Guanacos (Lama guanicoe) are the most widespread large herbivore in Patagonia, but land-use change, overhunting, and competition with livestock drove their populations to approximately 3–7% of their original abundance by the early twentieth century (Baldi et al. 2016). Roads are a potential threat to remaining guanaco populations. Previous research suggests that guanacos are negatively affected by roads both directly, via vehicle collisions and roadside fence entanglements, and indirectly, as roads provide hunters easy access to guanacos (Rey et al. 2012; Radovani et al. 2014). Still, studies looking at the effects of roads on guanaco abundance and distribution have found mixed results (Cappa et al. 2017, 2019; Schroeder et al. 2018). In La Payunia Reserve, unpaved roads were found to have no effect on guanaco abundance (Schroeder et al. 2018). However, guanaco responses to roads may differ based on the road type. Unpaved dirt roads have lower traffic volume and speed, and potentially offer easier travel for guanacos. However, this easier travel may come at a cost of increased accessibility for poachers and hunters. In contrast, paved roads, while posing risks from high-speed traffic, can significantly alter the physical terrain, potentially facilitating water accumulation and plant growth that could attract guanacos. One study in northern Argentina found a greater abundance of guanaco dung piles near unpaved roads than paved roads, potentially indicating that paved roads negatively affect guanaco distribution (Cappa et al. 2019). Still, it is unclear how both paved and unpaved roads impact guanaco behavior, whether they attract or repel guanacos, and how they may act as barriers to movement. Understanding this behavior is particularly important as guanaco population estimates almost entirely rely on road-based surveys. The attraction or avoidance of roads can introduce substantial bias, jeopardizing the accuracy of these estimates.
In this study, we integrate multiple animal movement analyses to evaluate guanaco habitat use and movement behavior near both paved and unpaved roads (i.e., proximity effect), and to understand how roads influence movement across roads (i.e., crossing effect). The proximity effect is the probability of using habitat as a function of distance to the road, whereas the crossing effect assesses the permeability of the road. Further, because guanaco behavioral responses may be mediated by their behavioral state or by seasonality, we analyzed the effects of roads separately by season (spring–summer vs. autumn–winter) and behavioral state (resting, foraging, and traveling). We hypothesized that roads will impact the habitat selection of guanacos, but only during specific behavioral states. Specifically, we predict that guanacos would select for roads while foraging due to greater primary productivity all year round, but selection for roads will be stronger for paved roads in the spring and summer months when there is less water availability for plant growth. In addition, we predicted that guanacos would avoid paved roads when traveling due to high traffic volume and select unpaved roads, as they may facilitate less energetically demanding movement. Finally, again due to the variation in traffic volume, we predict that guanacos would avoid crossing paved roads but would readily traverse unpaved roads. We do not expect these patterns to change seasonally.
Methods
Study area
Established in 2004, Monte León National Park (MLNP) is located on the east coast of Argentina within the Santa Cruz province, covering 61,000 hectares and 30 km of coastline (Fig. 1). The park is characterized by coastal steppe with both shrubland and grassland ecosystems. Black shrub (Junellia tridens) and pine-scented daisy (Lepidophyllum cupressiforme), dominate the shrublands, while thatching grass (Festuca pallescens), salt grass (Puccinellia sp.), bentgrass (Agrostis sp.), and coast tussock-grass (Poa atropidiformis) dominate the grasslands (Oliva et al. 2006). Average annual precipitation is 255 mm, characterized by rainfall during the autumn–winter months (April-September) and a dry season in spring–summer (October–March) (Paruelo et al. 1998). Average temperatures are 4.7 ℃ during the autumn–winter months and 13.8 ℃ during the spring–summer months. Therefore, we split our data into two distinct periods, the autumn–winter months (April-September) with lower temperature and more precipitation and the spring–summer months (October–March) with higher temperatures and lower precipitation.

Map of Monte León National Park (MLNP) and location of the highway (Ruta 3) and unpaved roads within the park. Inset map includes Argentina with a black dot representing the location of MLNP
Some guanaco populations are beginning to recover in Patagonia due to the creation of protected areas and the abandonment of ranches in the region (Carmanchahi and Lichtenstein 2022). MLNP is one key area where guanaco populations are thought to be recovering and is one of the few places in their range without hunting and competition with livestock (Walker and Novaro 2010). The park is bordered by a major, heavily used state highway, Ruta 3, that separates the park from neighboring ranches. Near MLNP, guanacos are often observed both on and alongside the paved highway.
GPS collar data
From 2019 to 2022 we monitored 29 adult guanacos (14 males and 15 females) during the spring–summer season and 28 (14 males and 14 females) during the autumn–winter season in Monte León National Park in Southern Argentina following all National Park regulations and protocols (Supplementary Table 1). Each GPS collar (LiteTrack Iridium 420, Lotek, Ontario, Canada) was programmed to capture one fix every two hours. For all subsequent analyses, we included GPS locations from collars that recorded data for a minimum of four months between 2019 and 2022. We treated each year of monitoring for individual guanacos as separate individuals (referred to as a guanaco year) to account for changing habitat conditions between years of study (Prokopenko et al. 2017). Multiple guanacos were monitored in more than 1 year of the study resulting in 58 guanaco-years for the spring–summer season and 57 guanaco-years during the autumn–winter season. Fieldwork was conducted under permit IF-2019–111378017-APN-DRPA#APNAC and subsequent renewals issues from the National Park Administration of Argentina (Administración de Parques Nacionales), and UC Berkeley IACUC Protocol # AUP-2019–10-12628.
Behavioral segmentation
We fit Hidden Markov Models (McClintock and Michelot 2018) to segment guanaco trajectories into distinct behavioral states, based on modeled step lengths and turning angles. Step lengths were modeled using a gamma distribution, while turning angles were modeled using a Von Mises distribution (Langrock et al. 2012). For the initial parameter values, we defined three behavioral states: a resting state characterized by short step lengths (gamma distribution with a mean of 20 m and a standard deviation of 20 m) and uniform turning angles (Von Mises distribution with a mean of π and a concentration of 0), a foraging state with medium-sized step lengths (gamma distribution with a mean of 350 m and a standard deviation of 350 m) and uniform turning angles (Von Mises distribution with a mean of π and a concentration of 0), and a traveling state with large step lengths (gamma distribution with a mean of 1,500 m and a standard deviation of 1,500 m) and directed movement (Von Mises distribution with a mean of 0 and a concentration of 1.5). We then assigned each step to the most probable behavioral state using the Viterbi algorithm based on the results of the HMM. This analysis was conducted using the momentumHMM package (McClintock and Michelot 2018) in the R programming language.
Environmental covariates
Roads for our analysis were both downloaded from publicly available datasets and hand digitized. Ruta Nacional 3 (“Ruta 3”) is a major transportation corridor for Argentina that stretches from Buenos Aires to Tierra del Fuego. Geospatial data for Ruta 3 was downloaded from The World Bank (https://datacatalog.worldbank.org/search/dataset/0038248). For the unpaved roads, we hand digitized roads using satellite imagery and supplementary information from Administración de Parques Nacionales. The roads were categorized into two classes: highways, which are major roads characterized by high speeds and heavy traffic volume, and unpaved roads, comprising unpaved and gravel roads that traverse the park with minimal usage. Traffic for both Ruta 3 and the unpaved roads changes from season to season, with traffic increasing during the spring–summer months. Still, as a major shipping route, Ruta 3 remains heavily trafficked throughout the year. Further, while tourist visitation to the park increases during the summer, visitation is relatively low. We included two additional environmental covariates in our habitat selection model (see below), the terrain ruggedness index (TRI) and the Normalized Difference Vegetation Index (NDVI), in our modeling framework. To calculate a terrain ruggedness index (TRI), we downloaded NASA’s STRM 30 m Digital Elevation Model from Google Earth Engine and calculated TRI from the terrain function in R. Finally, using the rgee and reticulate package in R, we matched each GPS location with the closest 30 m spatial resolution Landsat 8 image (every 16 days) and calculated the Normalized Difference Vegetation Index.
Roadside fencing
Roadside fences are commonly deployed worldwide along high-traffic, high-speed roadways to prevent wildlife collisions and demarcate borders or private property. In some cases, these fences may exacerbate the barrier effect of large roadways (McInturff et al. 2020). Assessing the individual effects of each feature becomes challenging due to the close proximity of these fences to the road (Jones et al. 2022). Further, at a 2-h fix rate, the barrier effect observed is at a coarser scale and may miss occasions at which individuals quickly cross back and forth between barriers. At MLNP, our GPS data revealed that after crossing the roadside fence, guanacos crossed back over the fence without crossing the road 65% of the time. In our study, we treat Ruta 3 and the roadside fence as a unified barrier.
Habitat selection
We used a series of integrated step selection analyses (iSSA) to compare the GPS relocations of guanacos (i.e., used locations) with available locations to estimate relative probability of use for each behavioral state (Avgar et al. 2016; Picardi et al. 2021). To evaluate habitat selection, we divided the data based on behavioral state (resting, foraging, and traveling) and season (spring–summer and autumn–winter). We generated 10 random steps for each used step by randomly selecting the step length and turning angles from a gamma and von Mises distribution (Avgar et al. 2016). The gamma and von Mises distributions were generated separately for each individual and behavioral state, and parameterized based on the empirical distribution of used step lengths and turning angles of that individual. For each model, we included consistent core environmental covariates expected to influence guanaco habitat selection regardless of road effects (Prokopenko et al. 2017; Londe et al. 2022). Our selection of environmental variables was informed by previous research on guanaco habitat selection, which emphasized the importance of ruggedness and forage availability (Verta 2022). Both forage availability (here estimated by NDVI) and terrain ruggedness have been found to be important predictors of guanaco movement and distribution in several studies (Marino and Baldi 2008; Pedrana et al. 2010; Flores et al. 2012; Smith et al. 2020; Verta 2022). For NDVI, we also included a quadratic term as individuals may be selecting for more intermediate values of forage (Verta 2022). These covariates were estimated at the end of each individual step. In all models we also included the step length, the logarithm of step length, and the cosine of the turning angles as covariates to account for the movement process (Avgar et al. 2016). Lastly, COVID-19 led to widespread restrictions in vehicular traffic in Argentina from April 2020 until November 2020, so we included an interaction term for the months with restricted traffic due to COVID-19 on our proximity and crossing variables.
To model selection of proximity to each road type, we included a variable for the distance to a highway and unpaved road at the end of a random or observed step. This variable assumes that if a road influences selection, individuals will, on average, select steps that end either closer to or farther away from the road compared to what would be expected randomly. For each distance variable, we used a decay function, with ɑ set to 1000 m, to account for a decreased behavioral response further from the road (Carpenter et al. 2010). Further, we separately modeled a crossing effect for both road types. We created a crossing covariate by tracing movement paths from each point (used and available) to the last point. We separated models containing road proximity covariates and road crossing covariates, as their simultaneous evaluation within a single model is discouraged due to collinearity (Prokopenko et al. 2017; Jones et al. 2022; Londe et al. 2022). Prior to modeling by season and state, we employed model selection with Akaike’s information criteria (AIC) evaluation for all covariate combinations, finding that the full model with all environmental and road variables yielded the lowest AIC score (for details see Supplementary Table 2). Therefore, we included all covariates for subsequent modeling by season and behavior.
We built mixed conditional logistic regression models for each model by state and season, with strata for each pairing of used and available locations and guanaco year as a random effect. All covariates were scaled and centered. We defined each model using the R package glmmTMB to allow for each guanaco year as a random effect (Brooks et al. 2017). We evaluated the support for each fixed effect by examining the 95% confidence intervals, and we interpreted variables with confidence intervals that did not overlap zero as significant. We also calculated the Variance Inflation Factor for each variable to ensure an acceptable level of collinearity (VIF < 4.0 for all predictors in each model). Lastly, we evaluated the performance of each model by calculating the continuous Boyce index which compares the predicted values with expected values across the study area (Hirzel et al. 2006). Values for the continuous Boyce index range from 0 to 1 with values closer to 1 indicating better model performance.
Results
The average number of relocations varied across seasons with each guanaco having on average 1,982 (SD = 1,131) observed steps in the spring–summer season and 1,971 (SD = 1,267) observed steps in the autumn–winter season. Estimated mean step length (SD) was 10 (7) m for the resting state, 266 (231) m for the foraging state, and 1,012 (811) m for the traveling state (Supplementary Material Figs. 1, 3 and 5). Turning angle distribution (concentration parameter) had an estimated mean of 3.11 (0.34) for the resting state, 2.95 (0.02) for the foraging state, and 0.04 (0.87) for the traveling state (Supplementary Material Figs. 2, 4, and 6 l). Overall, using the Viterbi Algorithm, guanacos were resting 28% of the time, foraging 48% of the time, and traveling 24% of the time. During the daytime guanacos were resting 1% of the time, foraging 60% of the time, and traveling 39% of the time. However, during the nighttime, guanacos were resting 55% of the time, foraging 36% of the time, and traveling 9% of the time. During the spring–summer period, guanacos were resting 21% of the time, foraging 56% of the time, and traveling 23% of the time. Similarly, during the winter-autumn period, guanacos were resting 36% of the time, foraging 42% of the time, and traveling 22% of the time.
During the spring–summer season, guanacos had 4,961 (8%) and 41,389 (67%) observed steps within 1 km of the highway and unpaved roads, respectively (Fig. 2). During this time, guanacos crossed the highway a total of 101 times and unpaved roads 5,899 times. During the autumn–winter season, guanacos had 6,424 (10%) and 31,358 (59%) observed steps within 1 km of the highway and unpaved roads, respectively. During this time, guanacos crossed the highway a total of 51 times and unpaved roads 3,345 times. Proximity to the highway was not contingent on the time of day, with 56.1% of the observed steps within 1 km of the highway occurring during the day and 43.9% at night. Likewise, proximity to unpaved roads was not dependent on the time of day, with 54.6% of the observed steps within 1 km of unpaved roads occurring during the day and 45.4% at night. There were 2,497 GPS points within one average step length (245 m) of the highway, and of all those points, 2,401 points (96%) were on the parkside. There was considerable individual variation as the maximum number of highway crossings observed by an individual was 35, while the minimum was 0, and the median was 2. Six individuals never crossed the highway. Of the 152 times guanacos crossed the highway, they crossed back within 24 h 88% of the time (n = 133).

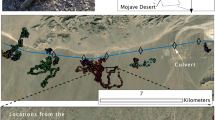
Guanacos foraging along the highway (Ruta 3). The map includes all GPS locations within 1 km of the highway east and every GPS location west of the highway. Parque Nacional Monte León, Santa Cruz Province, Argentina
Model parameter estimates for the proximity model differed by behavioral states and season (Fig. 3). The interaction term between the COVID-19 months and the proximity and crossing variables was not significant in all models and was therefore removed (p > 0.05 in all cases). In a resting state, guanacos only selected for proximity to unpaved roads during the autumn–winter season (autumn:winter 95% CI 0.01—0.04). In a foraging state, guanacos selected for proximity to highways in both seasons (spring:summer 95% CI -0.06—-0.04 and autumn:winter 95% CI -0.03—-0.01), but only selected for unpaved during the spring–summer season (spring:summer 95% CI -0.03—-0.01). However, in a traveling state, guanacos selected against proximity to highways in both seasons (spring:summer 95% CI: 0.04—0.09 and autumn:winter 95% CI: 0.20—0.70), and for proximity to unpaved roads (spring:summer 95% CI -0.08—-0.03 and autumn:winter 95% CI -0.04—-0.01).

Spring–summer (A) and autumn–winter (B) proximity model parameter estimates by separate behavioral state models. Parque Nacional Monte León, Santa Cruz Province, Argentina
Model parameter estimates for the crossing model also differed by behavioral state and season (Fig. 4). Due to the short step lengths of the resting state, only 1 available step crossed the highway. Therefore, the crossing model for the resting state during both seasons failed to converge. During the foraging state, guanacos selected against crossing the highway during both seasons (spring:summer 95% CI -1.55—-0.99 and autumn:winter 95% CI -2.20—-1.42) and unpaved roads during the spring–summer season (95% CI -0.12—-0.04). Similarly during a traveling state, guanacos selected against crossing the highway during both seasons (spring:summer 95% CI -2.10—-1.51 and autumn:winter 95% CI -2.70—-1.98), but showed no selection for or against crossing unpaved roads.

Spring–summer (A) and autumn–winter (B) crossing model parameter estimates by separate behavioral state models. Parque Nacional Monte León, Santa Cruz Province, Argentina
Finally, each model had a relatively high predictive performance (Hirzel et al. 2006). The continuous Boyce index values were all higher than 0.70 (Supplementary Table 3).
Discussion
Roads are among the most prominent anthropogenic features on the planet, yet their impact on wildlife can be difficult to discern because it can vary by species, behavioral state, and season. Our findings indicate that the guanaco, the most widespread large herbivore in Patagonia, responds differently to different types of roads based on their behavioral state. When foraging, guanacos selected strongly for highways. However, when traveling, guanacos selected for unpaved roads and against highways. Despite strong selection for highways when foraging, guanacos consistently avoided crossing them. The aversion to crossing highways was observed regardless of their behavioral state. Combined, the strong selection of proximity to highways while foraging and the avoidance of crossing them, drastically impact guanaco behavior. This pattern could have substantial effects on guanaco distributions throughout Patagonia, with potential implications for the conservation and management of this species.
We found that guanacos selected for roads, particularly while they are foraging. Guanacos may be foraging near roads to capitalize on the facilitation of foraging opportunities provided by the presence of roads. In addition, the selection of intermediate values of NDVI while foraging further supports these findings. In fact, previous research also found guanaco to prefer intermediate values of vegetation cover (Verta 2022). Roads can have significant impacts on vegetation patterns and productivity, and these effects may be more pronounced in semi-arid and arid ecosystems. In these environments, where water is typically a limiting factor for plant growth, the runoff water from roads can enhance vegetation productivity, resulting in patches of more abundant and diverse vegetation along road edges (Dean et al. 2019). Plant communities alongside roads may also include more introduced species that green up faster than the surrounding landscape (Roever et al. 2008). Additionally, the concentration of total dissolved nitrogen can be higher near roads. Therefore, changes to vegetation productivity and quality may result in increased selection of habitat near roads. In the semi-arid saltbrush steppe of Australia, kangaroos are attracted to roads during drought years when resources are scarce (Lee et al. 2004; Klöcker et al. 2006). Guanacos in Patagonia, however, appear to be consistently attracted to the highway while foraging throughout the year. Further, some species may associate with roads to avoid predation (Hill et al. 2021). Across their range, pumas, the main predator of guanacos at MLNP, are thought to mostly avoid high-use, paved roads (Dickson et al. 2005; Caruso et al. 2015). Despite potential anti-predator benefits, the scarcity of guanaco sightings near roadsides at night suggests road association is unlikely to be driven primarily by predator avoidance. Ultimately, the proximate mechanism for this behavior is still not well understood and future work should test for differences in nutrient availability or minerals in plants along the road to better understand if differences in forage availability results in increased selection of roads.
Although guanacos forage near highways, they consistently avoid crossing them, indicating that highways serve as significant barriers to their movement. Ruta 3, the highway bordering MLNP, is a heavily trafficked transportation and shipping route frequented by freight trucks. Throughout the length of the study, we observed very few instances in which guanacos crossed the highway. However, around MLNP and other areas with high densities of guanacos, it is very common to see individuals walking on the highway. So, given our 2-h fix rate, it is likely that we missed events in which guanacos quickly crossed the highway, but then crossed back. Still, guanaco locations within an average step length of the highway were concentrated in the parkside (96%), suggesting minimal crossing events. It is widely known that roads can be barriers to movement for wildlife, hence the proliferation of road crossing structures in recent years (Smith et al. 2015; Sijtsma et al. 2020). However, considering the strong preference for roads while foraging, the extent of the barrier effect on guanaco movement is surprising. The ultimate explanation for this barrier effect is still not fully understood. Factors such as high traffic volume and roadside fencing are both commonly cited reasons for road avoidance and likely deter some individual guanacos from crossing (Jacobson et al. 2016). However, if individuals are crossing short spatiotemporal scales but returning to the park side of the road, it could indicate lower vegetation quality in ranchlands or the avoidance of private lands due to fear of persecution as potential drivers of this behavior. In fact, previous studies, for example, have found that guanacos have increased flight behavior where hunting pressure is high (Donadio and Buskirk 2006). A barrier effect that limits guanaco space use is a major concern, so further unpacking whether the driver of this barrier is traffic, roadside fencing, forage availability in neighboring ranches, or other human influences may be critical for future guanaco conservation efforts (Puig et al. 1997; Schroeder et al. 2014).
The road-induced impacts on guanaco behavior requires further investigation to mitigate potential management and conservation challenges. First, while the attraction of guanacos to roads can benefit some individuals, it can also result in an ecological trap, leading to detrimental consequences for individuals and populations. For other species, increased association with roads can lead to increases in vehicle collisions, poaching, exposure to heavy metals, noise pollution, and artificial light (Frangini et al. 2022). Second, increased association with roads may contribute to both a public perception of guanaco overabundance and skewed population estimates by managers, potentially leading to the mismanagement of this species. Throughout Patagonia, sheep ranchers are increasingly concerned about the recovery of some guanaco populations due to potential competition with livestock, and many believe that current populations are overabundant, beyond carrying capacity, and contributing to a decline in the profitability of livestock husbandry (Flores et al. 2023). In response to concerns about declining productivity, the Santa Cruz province implemented a resolution in 2023 that increased the daily guanaco harvest quota from 1 to 2 animals per hunter (El Consejo Agrario Provincial de Santa Cruz 2023). Regular sightings of guanacos along the roadside may be influencing the public perceptions of these animals, fueling a bad reputation of this camelid in the region. In addition to local perceptions of guanaco overabundance, the attraction to roads may also be contributing to inaccurate population estimates. Due to their cost-effectiveness and ability to be easily repeated, ground transect surveys conducted along both paved and unpaved roads are the main methodology used for estimating guanaco population densities (Baldi et al. 2001; Travaini et al. 2007, 2015; Pedrana et al. 2010; Marino et al. 2014; Zubillaga et al. 2018). Even in MLNP, population estimates for guanacos are conducted from roads. However, a major assumption of population estimates from road surveys is that the impact of roads on the target species distribution is negligible. Therefore, it’s possible that population estimates for guanacos in MLNP and other regions where guanacos are drawn to roads may be inflated. An overestimation of the guanaco population can have direct impacts on harvest and other wildlife policies, so park managers and researchers should explore other options to measure their densities (e.g. drones, or ground surveys away from roads) to ensure accurate assessments of guanaco populations and informed wildlife management policies.
Data availability
Data will be made available at Dryad upon acceptance of the manuscript.
References
Abrahms B, Jordan NR, Golabek KA et al (2015) Lessons from integrating behavior and resource selection: activity-specific responses of African wild dogs to roads. Anim Conserv 19:247–255. https://doi.org/10.1111/acv.12235
Aiello CM, Galloway NL, Prentice PR et al (2023) Movement models and simulation reveal highway impacts and mitigation opportunities for a metapopulation-distributed species. Landsc Ecol 38:1085–1103. https://doi.org/10.1007/s10980-023-01600-6
Avgar T, Potts JR, Lewis MA, Boyce MS (2016) Integrated step selection analysis: bridging the gap between resource selection and animal movement. Methods Ecol Evol 7:619–630. https://doi.org/10.1111/2041-210X.12528
Baldi R, Albon S, Elston D (2001) Guanacos and sheep: Evidence for continuing competition in arid Patagonia. Oecologia 129:561–570. https://doi.org/10.1007/s004420100770
Baldi RB, Acebes P, Cuéllar E, Funes M, Hoces D, Puig S, Franklin WL (2016) Lama guanicoe. The IUCN Red List of Threatened Species 2016 e.T11186A18540211
Bauer S, Hoye BJ (2014) Migratory Animals Couple Biodiversity and Ecosystem Functioning Worldwide. Science 344:1242552. https://doi.org/10.1126/science.1242552
Brehme CS, Tracey JA, Mcclenaghan LR, Fisher RN (2013) Permeability of Roads to Movement of Scrubland Lizards and Small Mammals. Conserv Biol 27:710–720. https://doi.org/10.1111/cobi.12081
Brooks ME, Kristensen K, van Benthem KJ et al (2017) glmmTMB Balances Speed and Flexibility Among Packages for Zero-inflated Generalized Linear Mixed Modeling. R J 9:378–400
Buxton RT, Mckenna MF, Brown E et al (2020) Varying behavioral responses of wildlife to motorcycle traffic. Glob Ecol Conserv 21:e00844. https://doi.org/10.1016/j.gecco.2019.e00844
Cappa F, Campos V, Giannoni S, Andino N (2017) The effects of poaching and habitat structure on anti-predator behavioral strategies: A guanaco population in a high cold desert as case study. PLoS ONE 12:1–11. https://doi.org/10.1371/journal.pone.0184018
Cappa FM, Borghi CE, Giannoni SM (2019) How roads affect the spatial use of the guanaco in a south american protected area: Human connectivity vs animal welfare. Diversity 11:1–10. https://doi.org/10.3390/D11070110
Carmanchahi P, Lichtenstein G (2022) Guanacos and people in Patagonia: a social-ecological approach to a relationship of conflicts and opportunities. Springer Nature. https://doi.org/10.1007/978-3-031-06656-6
Carpenter J, Aldridge C, Boyce MS (2010) Sage-Grouse Habitat Selection During Winter in Alberta. J Wildl Manag 74:1806–1814
Caruso N, Guerisoli M, Luengos Vidal EM et al (2015) Modeling the ecological niche of an endangered population of Puma concolor: First application of the GNESFA method to an elusive carnivore. Ecol Model 297:11–19. https://doi.org/10.1016/j.ecolmodel.2014.11.004
Chambers S, Villarreal ML, Duane O et al (2022) Conflict of energies: spatially modeling mule deer caloric expenditure in response to oil and gas development. Landsc Ecol 37:2947–2961. https://doi.org/10.1007/s10980-022-01521-w
Dean WRJ, Seymour CL, Joseph GS, Foord SH (2019) A review of the impacts of roads on wildlife in semi-arid regions. Diversity 11:1–19. https://doi.org/10.3390/D11050081
Dickson BG, Jenness JS, Beier P (2005) Influence of Vegetation, Topography, and Roads on Cougar Movement in Southern California. J Wildl Manag 69:264–276. https://doi.org/10.2193/0022-541X(2005)069%3c0264:IOVTAR%3e2.0.CO;2
Donadio E, Buskirk SW (2006) Flight behavior in guanacos and vicuñas in areas with and without poaching in western Argentina. Biol Conserv 127:139–145. https://doi.org/10.1016/j.biocon.2005.08.004
El Consejo Agrario Provincial de Santa Cruz (2023) Temporada de Caza 2023 in Santa Cruz, Argentina. Resolution 113, Article 2
Flores CE, Cingolani AM, von Müller A, Barri FR (2012) Habitat selection by reintroduced guanacos (Lama guanicoe) in a heterogeneous mountain rangeland of central Argentina. Rangel J 34:439–445. https://doi.org/10.1071/RJ12040
Flores C, Lichtenstein G, Schiavini A (2023) Human-wildlife conflicts in Patagonia: Ranchers’ perceptions of guanaco Lama guanicoe abundance. Oryx 1–11. https://doi.org/10.1017/S0030605322001508
Frangini L, Sterrer U, Franchini M et al (2022) Stay home, stay safe? High habitat suitability and environmental connectivity increases road mortality in a colonizing mesocarnivore. Landsc Ecol 37:2343–2361. https://doi.org/10.1007/s10980-022-01491-z
Hill JE, De Vault TL, Belant JL (2021) A review of ecological factors promoting road use by mammals. Mammal Rev 51:214–227. https://doi.org/10.1111/mam.12222
Hill JE, Devault TL, Belant JL (2019) Cause ‐ specific mortality of the world ’ s terrestrial vertebrates. 1–10. https://doi.org/10.1111/geb.12881
Hirzel AH, Le Lay G, Helfer V et al (2006) Evaluating the ability of habitat suitability models to predict species presences. Ecol Model 199:142–152. https://doi.org/10.1016/j.ecolmodel.2006.05.017
Holderegger R, Di Giulio M (2010) The genetic effects of roads: A review of empirical evidence. Basic Appl Ecol 11:522–531. https://doi.org/10.1016/j.baae.2010.06.006
Jacobson SL, Bliss-Ketchum LL, de Rivera CE, Smith WP (2016) A behavior-based framework for assessing barrier effects to wildlife from vehicle traffic volume. Ecosphere 7:e01345. https://doi.org/10.1002/ecs2.1345
Jones PF, Jakes AF, Vegter SE, Verhage MS (2022) Is it the road or the fence? Influence of linear anthropogenic features on the movement and distribution of a partially migratory ungulate. Mov Ecol 10:1–13. https://doi.org/10.1186/s40462-022-00336-3
Klöcker U, Croft DB, Ramp D (2006) Frequency and causes of kangaroo–vehicle collisions on an Australian outback highway. Wildl Res 33:5–15. https://doi.org/10.1071/WR04066
Langrock R, King R, Matthiopoulos J et al (2012) Flexible and practical modeling of animal telemetry data: hidden Markov models and extensions. Ecology 93:2336–2342. https://doi.org/10.1890/11-2241.1
Lee E, Klöcker U, Croft DB, Ramp D (2004) Kangaroo-vehicle collisions in Australia’s sheep rangelands, during and following drought periods. Aust Mammal 26:215–226. https://doi.org/10.1071/am04215
Londe DW, Elmore RD, Davis CA et al (2022) Why did the chicken not cross the road? Anthropogenic development influences the movement of a grassland bird. Ecol Appl 32:1–17. https://doi.org/10.1002/eap.2543
Marino A, Baldi R (2008) Vigilance patterns of territorial guanacos (Lama guanicoe): The role of reproductive interests and predation risk. Ethology 114:413–423. https://doi.org/10.1111/j.1439-0310.2008.01485_1.x
Marino A, Pascual M, Baldi R (2014) Ecological drivers of guanaco recruitment: Variable carrying capacity and density dependence. Oecologia 175:1189–1200. https://doi.org/10.1007/s00442-014-2965-z
McClintock BT, Michelot T (2018) momentuHMM: R package for generalized hidden Markov models of animal movement. Methods Ecol Evol 9:1518–1530. https://doi.org/10.1111/2041-210X.12995
McClure CJW, Ware HE, Carlisle J et al (2013) An experimental investigation into the effects of traffic noise on distributions of birds: avoiding the phantom road. Proc R Soc B Biol Sci 280:20132290. https://doi.org/10.1098/rspb.2013.2290
McInturff A, Xu W, Wilkinson CE et al (2020) Fence Ecology: Frameworks for Understanding the Ecological Effects of Fences. Bioscience 70:971–985. https://doi.org/10.1093/biosci/biaa103
Meijer JR, Huijbregts MAJ, Schotten KCGJ, Schipper AM (2018) Global patterns of current and future road infrastructure. Environ Res Lett 13. https://doi.org/10.1088/1748-9326/aabd42
Nathan R (2008) An emerging movement ecology paradigm. PNAS 105:19050–19051
Oliva G, Escobar J, Siffredi G, et al (2006) Monitoring Patagonian Rangelands: The MARAS System. Monitoring Science and Technology Symposium. U.S. Dept. Agriculture, Forest Service, Denver, CO
Ortega YK, Capen DE (1999) Effects of Forest Roads on Habitat Quality for Ovenbirds in a Forested Landscape. Auk 116:937–946. https://doi.org/10.2307/4089673
Paruelo JM, Beltran A, Jobbagy E et al (1998) The climate of Patagonia: General patterns and controls on biotic processes. Ecol Austral 8:85–101
Pedrana J, Bustamante J, Travaini A, Rodríguez A (2010) Factors influencing guanaco distribution in southern Argentine Patagonia and implications for its sustainable use. Biodivers Conserv 19:3499–3512. https://doi.org/10.1007/s10531-010-9910-1
Picardi S, Mathews S, Coates P, Kolar J (2021) Behavioural state-dependent habitat selection and implications for animal translocations. https://doi.org/10.1111/1365-2664.14080
Pike KN, Blake S, Gordon IJ et al (2023) Navigating agricultural landscapes: responses of critically endangered giant tortoises to farmland vegetation and infrastructure. Landsc Ecol 38:501–516. https://doi.org/10.1007/s10980-022-01566-x
Prokopenko CM, Boyce MS, Avgar T (2017) Characterizing wildlife behavioural responses to roads using integrated step selection analysis. J Appl Ecol 54:470–479. https://doi.org/10.1111/1365-2664.12768
Puig S, Videla F, Cona MI (1997) Diet and abundance of the guanaco (Lama guanicoeMüller 1776) in four habitats of northern Patagonia, Argentina. J Arid Environ 36:343–357. https://doi.org/10.1006/jare.1996.0186
Radovani NI, Funes MC, Walker RS et al (2014) Guanaco Lama guanicoe numbers plummet in an area subject to poaching from oil-exploration trails in Patagonia. Oryx 754:42–50. https://doi.org/10.1017/S0030605312001226
Rey A, Novaro AJ, Guichón ML (2012) Guanaco (Lama guanicoe) mortality by entanglement in wire fences. J Nat Conserv 20:280–283. https://doi.org/10.1016/j.jnc.2012.05.004
Roever CL, Boyce MS, Stenhouse GB (2008) Grizzly bears and forestry: II: Grizzly bear habitat selection and conflicts with road placement. For Ecol Manag 256:1262–1269. https://doi.org/10.1016/j.foreco.2008.06.006
Schroeder NM, Matteucci SD, Moreno PG et al (2014) Spatial and Seasonal Dynamic of Abundance and Distribution of Guanaco and Livestock: Insights from Using Density Surface and Null Models. PLoS ONE 9:e85960. https://doi.org/10.1371/journal.pone.0085960
Schroeder NM, González A, Wisdom M, et al (2018) Roads have no effect on guanaco habitat selection at a Patagonian site with limited poaching. Glob Ecol Conserv 14. https://doi.org/10.1016/j.gecco.2018.e00394
Sijtsma FJ, van der Veen E, van Hinsberg A et al (2020) Ecological impact and cost-effectiveness of wildlife crossings in a highly fragmented landscape: a multi-method approach. Landsc Ecol 35:1701–1720. https://doi.org/10.1007/s10980-020-01047-z
Smith DJ, van der Ree R, Rosell C (2015) Wildlife Crossing Structures. In: Handbook of Road Ecology. John Wiley & Sons, Ltd, pp 172–183 https://doi.org/10.1002/9781118568170.ch21
Smith JA, Donadio E, Bidder OR, et al (2020) Where and when to hunt? Decomposing predation success of an ambush carnivore. Ecology. https://doi.org/10.1002/ecy.3172
Tanner D, Perry J (2007) Road effects on abundance and fitness of Galápagos lava lizards (Microlophus albemarlensis). J Environ Manage 85:270–278. https://doi.org/10.1016/j.jenvman.2006.08.022
Teixeira FZ, Rytwinski T, Fahrig L (2020) Inference in road ecology research: what we know versus what we think we know. Biol Lett 16. https://doi.org/10.1098/rsbl.2020.0140
Travaini A, Bustamante J, Rodríguez A et al (2007) An integrated framework to map animal distributions in large and remote regions. Divers Distrib 13:289–298. https://doi.org/10.1111/j.1472-4642.2007.00338.x
Travaini A, Zapata SC, Bustamante J, et al (2015) Guanaco abundance and monitoring in southern patagonia: Distance sampling reveals substantially greater numbers than previously reported. Zool Stud 54. https://doi.org/10.1186/s40555-014-0097-0
Verta G (2022) Multi-scale habitat selection by a large herbivore in a protected Patagonian landscape. University of California, Berkeley
Walker S, Novaro A (2010) The World’s Southernmost Pumas in Patagonia and the Southern Andes. In: Cougar: Ecology and Conservation 91–102. The University of Chicago Press
Zubillaga M, Skewes O, Soto N, Rabinovich JE (2018) How density-dependence and climate affect guanaco population dynamics. Ecol Model 385:189–196. https://doi.org/10.1016/j.ecolmodel.2018.07.010
Acknowledgements
We thank Administracion de Parques Nacionales (APN) and APN staff at MLNP for their gracious support and assistance. We also thank Conservacion Patagonica Association Civil for logistical and financial support. Finally, we would like to thank Matias Chambon, Emilia Perucca, Rocio Asueta, and Carolina Chiara for field assistance and Dr. Justin Brashares and Dr. Christopher Schell for feedback on the manuscript. The research was conducted under permit # DRPA 162 issued by APN, Argentina. Funding was provided by Prince Albert II of Monaco Foundation and University of California, Berkeley.
Funding
Funding was provided by Prince Albert II of Monaco Foundation and University of California, Berkeley.
Author information
Authors and Affiliations
Contributions
M.W.S. conceived the idea, designed the methodology, led the analysis, and wrote the manuscript. P.A.E.A. and E.D. led the collection of data. E.D. and A.D.M. obtained the grants for the research. All authors contributed to the editing and approval of the final manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors have no relevant financial or non-financial interests to disclose.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Serota, M.W., Alarcón, P.A.E., Donadio, E. et al. Behavioral state-dependent selection of roads by guanacos. Landsc Ecol 39, 110 (2024). https://doi.org/10.1007/s10980-024-01909-w
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s10980-024-01909-w