
Abstract
Context
The abandonment of traditional practices has transformed agro-pastoral systems, leading to a more frequent occurrence of passive rewilding of Mediterranean landscapes. Reconstructing ecosystem responses to climate under different grazing conditions (i.e., wild, and domestic ungulates) is important to understand the future of these ecosystems.
Objectives
Here we study the different roles of domestic and wild herbivory in defining the climate-vegetation interaction. Specifically, we evaluated (1) the effect of climate on primary productivity at the landscape scale and (2) the long-term trends of vegetation biomass in response to passive rewilding or maintenance of traditional grazing systems.
Methods
This study was carried out in South-eastern Spain. We used satellite images to generate NDVI time series that proxy primary productivity and vegetation biomass. We combined the NDVI and climate data from two key landscapes: one with wild ungulates and another predominantly with domestic ungulates.
Results
We detected a secondary succession process in areas with only wild ungulates. In domestic herbivory areas, vegetation biomass remained constant throughout time (30 years). In domestic herbivory areas temperature and seasonal precipitation affected primary productivity. In areas with only wild herbivory, primary productivity was mainly driven by annual precipitation, and it was less dependent on seasonal precipitation.
Conclusion
These results highlight the distinctive roles of herbivores in defining Mediterranean landscapes' adaptability to climate, through passive rewilding or traditional livestock use. Maintaining both ecosystems can enhance landscape heterogeneity and ecological sustainability in a context of climatic changes.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Changes in the foraging behaviour of different ungulate species and grazing intensity are key biotic factors that drive functional dynamics of plant communities, particularly in ecosystems where vegetation and ungulates have coevolved due to a long history of trophic interaction (Ellison 1960; Milchunas and Lauenroth 1993; Hobbs 1996; Augustine and McNaughton 1998). For instance, ungulate species composition and grazing intensity control temporal and spatial patterns of primary productivity (Frank et al. 2016; Velamazán et al. 2023), enhance nitrogen and carbon cycling (Frank et al. 1994; Hu et al. 2016), modify the nutrient content of growing plants (McNaughton 1985; Doughty 2017) and ultimately determine plant community composition and functional traits (Niu et al. 2016; Hernández et al. 2019). Because ungulates can drive all these ecological processes, we can expect that different grazing patterns can modulate the landscape-scale response of vegetation to resource availability and environmental conditions.
Previous studies have highlighted that grazing intensity is the main driver of long-term vegetation changes in humid regions, where annual and inter-annual precipitation variation is low (Vetter 2005). Moreover, the combined effect of low herbivore pressure and rising temperatures increase shrub areas in cold and dry areas, such as tundra biomes (Olofsson and Post 2018). In arid and semi-arid ecosystems, some authors have highlighted the importance of stochastic abiotic factors, such as droughts, as a significant driver of ecosystem functioning, especially affecting vegetation in overgrazed areas (Sullivan and Rohde 2002). Together, these studies suggest an interrelated influence of grazing and climate to determine landscape structure, composition, and functioning. These studies also highlight the importance of investigating how different grazing contexts (e.g., livestock, wild herbivores) affect vegetation responses to a variable climatic condition at different spatial and temporal scales.
Historically, domestic or wild ungulates are important drivers of ecological succession and ecosystem heterogeneity (Perevolotsky and Seligman 1998; San Miguel-Ayanz et al. 2010). In the past, livestock has replaced wild ungulates and occupied the most accessible grasslands due to human economic activities, while wildlife was displaced to more inaccessible ones (San Miguel-Ayanz et al. 2010; Giguet-Covex et al. 2014). Grasslands are one of the world's major ecosystems, covering about one-third of the earth's land surface (Bengtsson et al. 2019). However, in the past years, the abandonment of traditional practices and the climatic changes have been transforming agro-pastoral systems, a dynamic process that is still ongoing (Bowen et al. 2007). In fact, between 1990 and 2010, traditional grazing livestock in Europe decreased by 25% (Navarro and Pereira 2015). The progressive abandonment of these traditional practices is causing the decline of cultural landscapes (García-Ruiz et al. 2020) and allowing a consecutive process of passive rewilding (Nogués-Bravo et al. 2016; Pettorelli et al. 2019), that is a passive recolonization of wild herbivores (Speed et al. 2019). Understanding how ecosystems respond to landscape changes after land abandonment is a complex issue that requires local level information on land use change, climate and ungulates (MacDonald et al. 2000). Grazing pressure of wild and domestic ungulate species on Mediterranean landscapes differ because wild grazing contexts usually show a lower number of individuals and higher number of species when compared with livestock production. More important, the foraging behaviour of wild species is driven by a complex pattern of intra-specific species interaction and by a full dependence of the species on the ecosystem (Dupke et al. 2017).
Because global temperature is expected to increase in the coming years (Tebaldi and Knutti 2010), the combined changes in climate and land uses can affect landscape functioning and grasslands ecosystem services (Maestre et al. 2022) which can increase uncertainty as to whether the vegetation will persist in its original regime or shift to new community composition and functioning (Kaarlejärvi et al. 2015). Hence, we need to deepen our knowledge on reconstructing ecosystem responses to climate under different conditions of predominant grazing interactions (domestic or wild herbivores) and at large spatial and temporal scales. This scientific framework allows a retrospective perception of the ecological processes of these systems that can be extrapolated to future conditions and thus to better understand how Mediterranean landscapes function under different management scenarios.
Determining long-term ecological processes related to the interaction between climate, herbivory and vegetation at the landscape scale is difficult because quantitative measurements of vegetation are needed, and these data are often not available. However, in the last decade, analyses of remote sensing data have progressed enormously, providing different tools to evaluate landscape changes and trophic interactions at different spatial and temporal scales (Barbosa et al. 2020; Velamazán et al. 2023). There is a diverse collection of publicly accessible images of reflectance data from around the globe with unprecedented spatial, temporal, and radiometric resolutions (Babcock et al. 2021). All this information makes it possible to revisit fundamental ecological questions about the ecosystem functioning of different natural systems in a more holistic manner. For example, remote sensing data offer a unique opportunity to evaluate very long relationships between climate, vegetation, and herbivory over large regions, based on vegetation indices and, particularly, the Normalised Difference Vegetation Index (NDVI) (Pettorelli et al. 2011). Here, we have used images obtained from several Landsat and MODIS satellites to reconstruct vegetation indices time series as a proxy for temporal tendencies and oscillation of primary productivity and biomass.
Using a Bayesian approach to combine long-term vegetation greenness and climate data from two key landscapes, we studied the ecological roles of wild and domestic ungulates in determining the vegetation responses to climate and the long-term trends of semi-arid grassland vegetation. The studied landscapes show high similarities in both vegetation type and long-term climate conditions, albeit their contrasting managing conditions: one dominated by wild ungulates and the other dominated by domestic ungulates. Using as reference these contrasting grazing conditions, we ask: (1) How do long-term climate conditions affect landscape-scale primary productivity in each studied grazing context? (2) How do rewilding or the maintenance of traditional grazing systems determine the temporal tendencies of ecosystem primary productivity? We expect that areas with a long history of transhumant grazing practice will result in a sustainable production system with grassland ecosystem with high temporal stability, while areas where grazing has been limited and reduced over the years, will trend to a secondary succession towards an increase of shrublands. We also expect that both systems will follow the seasonality pattern typical in the Mediterranean ecosystems (Mooney et al. 1974). However, we expect that the most intensified grazed areas follow closely the seasonality pattern, and they are more controlled by climatic factors as they are more dependent on the water balance (Padilla and Pugnaire 2007). These results can easily be extended to other scenarios of change in traditional activities and land uses.
Methods
Study area
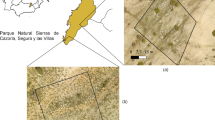
This study was conducted at two sites in the Sierras de Cazorla, Segura, and Las Villas Natural Park (Cazorla Natural Park) with an area of 2143 km2, one of Europe’s largest protected areas located in south-eastern Spain. The climate is Mediterranean, with annual rainfall between 300 and 1600 mm and an average temperature of 12–16 °C (Aguilera-Alcalá et al. 2022). We focused our study on two areas of this mountainous region: Los Campos de Hernán Pelea (CHP) and El Calar de Juana (CJ), located approximately 15 km apart (Fig. 1). Both study areas have similar topographic characteristics with altitudes ranging between 1600 and 1700 m, and similar climate conditions (see Supplemental Material from Fig. S.1. to Fig. S.2.). Annual rainfall in CHP is 670 mm, with a maximum of 1065 mm. The average temperature is 14.65 °C, with maximum temperatures up to 37 °C and minimum temperatures down to − 6 °C. CJ has an average annual rainfall of 630 mm, with a maximum of 958 mm. The average temperature in CJ is 14 °C, and it can fluctuate between a maximum of 36 °C and a minimum of − 7 °C. In both study areas, maximum rainfall occurs in December and minimum rainfall in July and August, coinciding with maximum temperatures.

Map of the study areas, El Calar de Juana (CJ; wild herbivory area) and Los Campos de Hernán Pelea (CHP; transhumant livestock herbivory area) in the Sierras de Cazorla, Segura, and Las Villas Natural Park (Spain)
Both CHP and CJ are situated within an area characterised by the same geomorphological, geological, and edaphic characteristics. Soils of both study localities are calcareous in origin and are diagnostic of scarce development generally lacking horizon development. The dominant soils are Calcaric Regosols and Leptosols (IUSS Working Group WRB 2015), typically located on fragmented bedrock. Continuous grazing practices over the years have induced modifications of the soil in both areas, with this effect being greater in CHP, due to its grazing history. These changes have resulted in distinct characteristics in these areas, mainly shaped by the presence of herbaceous vegetation adapted to grazing.
Both study areas are covered by forests, meadows, and shrublands, although the tree cover is very limited in both study areas. Most of the territory is covered by grassland, dotted with scrubland. Grasslands are dominated by Festuca hystrix, often accompanied by species like Poa ligulata, Koeleria vallesiana, and small woody plants from genera Thymus sp., Helianthemum sp., Arenaria sp. and others. Additionally, some areas feature grasslands dominated by Poa bulbosa. Subnitrophilous grasslands rich in endemic species appear in well-illuminated clearings within discontinuous shrublands and perennial grasslands of variable coverage, often accompanied by species from genera such as Limonium sp., Filago sp., Linaria sp., among others. Shrublands are characterized by low growth due to wind and snow exposure, dominated by Juniperus sabina, along with species like Prunus prostrata, Berberis vulgaris, Daphne oleoides, and others, including hawthorns like Erinacea anthyllis and Vella spinosa. Forests are scarce and disperse. They are primarily Pinus nigra subsp. salzmanii, with accompanying undergrowth like Juniperus phoenicea, J. sabina, and J. communis. Holm oak forests of Quercus rotundifolia appear stunted at higher altitudes due to colder temperatures, with undergrowth species including Lonicera arborea, Crataegus monogyna, and Sorbus aria (Valle Tendero et al. 2004; Benavente 2008; Gómez-Mercado 2011). In the past, scrub encroachment in the study area was partially managed through controlled fires. However, this practice was prohibited in 1986, when the Sierras de Cazorla, Segura, and Las Villas Natural Park was declared.
The main difference between CJ and CHP is the presence of domestic livestock. Wild ungulates such as deer (Cervus elaphus), fallow deer (Dama dama), mouflon (Ovis orientalis), and Iberian ibex (Capra pyrenaica) (Martínez Martínez 2002) can access both study areas. CJ is located in the Cazorla Reserve Area, where livestock access was restricted since 1986 and only wild ungulates can access. CHP is a communal grazing plateau, where transhumance has been the main practice for decades. Transhumance consists of the seasonal migration of livestock to optimize the exploitation of pastures. Livestock moves between summer pastures in highlands at northern latitudes and winter pastures in lowlands at more southern latitudes (Oteros-Rozas et al. 2012). Due to this regimen, there are two seasons in CHP, related to the presence (from June to November) or absence of livestock (from December to May). Most herds (> 90%) are composed of sheep, but there are also goats, cattle, and horses (Aguilera-Alcalá et al. 2022). Wild ungulate’s average yearly density in CHP is around 3.9 individuals per km2 (field data), and between 6 and 13 individuals per km2 in CJ according to the information provided by the Junta de Andalucía of the hunting censuses carried out in the Reserva Andaluza de Caza de Cazorla (R.A.C. Cazorla). However, in CHP from June to November, the livestock adds to the wild ungulates density between 131 and 163 animals per km2 according to the census provided by Agrupación de Defensa Sanitaria Ganadera Los Campos (ADSG). In addition, transhumant livestock is very controlled by shepards and, during half of the year, these animals are not present in the area as part of a strategy to optimise grassland utilisation at the landscape scale. Consequently, the grazing intensity in these two areas is quite different due to the dissimilarities in herbivory density and management regimes, as described by other authors (Rebollo and Gómez-Sal 2004). Therefore, CHP and CJ are two ideal study zones for determining landscape changes due to the different land use management and evaluating the consequences of maintenance or abandonment of traditional farming practices.
Datasets
Primary productivity and biomass
We used the Normalized Difference Vegetation Index (NDVI) to detect seasonal and interannual changes in primary productivity and biomass (Huete et al. 2002; Hamel et al. 2009), especially when vegetation cover is low (Gamon et al. 1995; Xu et al. 2012). NDVI has also proved to be useful to evaluate long-term interactions between ungulates and vegetation (Pettorelli et al. 2011), including the effect of grazing on spatial and temporal changes of primary productivity (Navalgund et al. 2007; Barbosa et al. 2020; Velamazán et al. 2023). To build a complete and consistent NDVI time series, we used a Bayesian model to combine the mean NDVI data for each area and the satellite images from several sensors: Landsat 5 ETM (L5), Landsat 7 ETM + (L7), Landsat 8 OLI (L8), and MODIS from the NASA Land Processes Distributed Active Archive Center (LP DAAC) (https://lpdaac.usgs.gov/). One of the major advantages of the Bayesian learning procedure is the possibility of introducing the information of one model in another as a previous or as subjective information to the model. We used this methodology as there is not always a useful image for each month due to the presence of clouds, shadows, snow, or the idiosyncrasies of the satellites. Although all these satellites have the same 16-day orbital repeat they do not all pass at the same instant, so while one may have passed in cloudy weather, the next may have passed in an open sky. Besides, the satellites were launched in different years, so they do not all cover the same time interval. We selected images for each sensor that had a minimum of 70% good pixels (no clouds, shadows, snow, or water) within the study area. We choose only areas of pastures and grasslands by masking pixels without tree cover fraction using Copernicus Global Land Cover Layers (Buchhorn et al. 2020). To process the satellite data, we used the platform Google Earth Engine (GEE) (Gorelick et al. 2017).
Climate data
We obtained precipitation and temperature data from ERA5, the latest climate reanalysis produced by the ECMWF (European Centre for Medium-Range Weather Forecasts) available at monthly time resolution from Copernicus Climate Data Store (cds.climate.copernicus.eu) which combines model data with observations from across the world. We obtained data from 1990 to 2019, from the 2 m average, minimum, and maximum monthly air temperature, as well as monthly total precipitation. We also used precipitation data to obtain cumulative precipitation for one, three, and twelve months. We did not include maximum and minimum temperatures in the model as they are strongly correlated with the average temperature (> 0.80) and the best models were those that included the average temperature. To process the climate data, we used the platform Google Earth Engine (GEE) (Gorelick et al. 2017).
Statistical analysis
Building the NDVI time series
To compare the relative importance of climate variables on primary productivity between the two study areas (CHP and CJ), we built two models, one for each of them. We used a multivariate autoregressive state-space model (MARSS) developed by Hinrichsen and Holmes (2009) because of its capability to infer the underlying trends from several observations without the need for prior variance estimates or complete observation datasets (Ward et al. 2010). We used this approach through the MARSS R-package (Holmes et al. 2012) to fill gaps in the NDVI time series and study its relationship with the climatic variables. The MARSS models include a process (1) model and an observation model (2).
In our case, the multivariate hidden process is given by Eq. 1, where the response variable \({{\varvec{x}}}_{t}\) is the NDVI at time t. The \({\varvec{B}}\) matrix indicates the relation of the states over time. The \({\varvec{u}}\) vector indicates the underlying stochastic growth rates. To determine the effect of climatic covariates the matrix \({{\varvec{c}}}_{t}\) was included in Eq. 1. This matrix consists of data about monthly precipitation, cumulative precipitation over the previous 3 and 12 months, and monthly average temperature. To ease comparisons between the effect of each covariate, this matrix was standardised (mean of 0 and standard deviation of 1). The matrix \({\varvec{C}}\) about how covariates interact with the response variable is set as unconstrained so that the effect of each covariate is flexible when fitting the model. The process error matrix at time t, \({{\varvec{w}}}_{t}\), has a multivariate normal distribution with a mean of zero and a covariance matrix \({\varvec{Q}}\).
The observation model (Eq. 2) is defined by the matrix \({{\varvec{y}}}_{t}\) composed of the NDVI time series obtained from the satellite images: L5, L7, L8, and MODIS. The \({{\varvec{y}}}_{t}\) matrix was standardised (mean of 0 and standard deviation of 1). The \({\varvec{Z}}\) matrix defines how the observations relate to the true response variable \({{\varvec{x}}}_{t}\). We assumed that each observation has the same trajectory since the same area is observed using different sensors. Consequently, matrix \({\varvec{Z}}\) was set as a 4 × 1 matrix of ones. The vector \({\varvec{a}}\) specifies the bias between the observations and the true response variable and it is set as an n × 1 vector of biases, the first of which is set equal to 0. \({{\varvec{v}}}_{t}\) specifies the observational errors of serially uncorrelated errors with a mean zero and a \({\varvec{R}}\) covariance matrix. We considered models that have the same level of process-error variance or that present covariance between the process errors, as we assume that they all contemplate constant observation errors.
We obtained the hidden state-space prediction from 500 iterations to generate estimates and fill in the missing NDVI data with a 95% credible interval (CI). The model has also been used to complete the gaps for the temporal series to solve the lack of information that occurs due to clouds. We choose the model with the lowest AICc (Hurvich and Tsai 1989) (see Table S.2 and S.3 in the Supplemental Material). We also graphically analysed the distribution of MARSS smoothing residuals, which are the model residuals using the expected value of \({x}_{t}\) conditioned on all the data, t = 1 to T (Holmes 2014). We used the distribution of residuals to analyse how well the models fit the data (Holmes 2014) (see Fig. S.4 to Fig. S.11 in the Supplemental Material). We tested that the model one-step-ahead residuals (innovations) have no temporal trend, and they fall within the 95% CI. We also tested that the Cholesky standardized model smoothation residuals from the observed data and the states have no temporal trend. We performed a residuals normality test to check the correlation between observed residuals and expected residuals under normality.
NDVI long-term trends
To determine whether changes in vegetation biomass occurred due to the different herbivory regimes, we used the resulting data from the MARSS model (see Fig. S.3 in the Supplementary Material) to perform a linear regression model between time and NDVI. This procedure was performed with a Hamiltonian Monte Carlo sampler using the R package brms (Bürkner 2021). To fit both models, we specify four chains of 1000 iterations, half of which are used for warm-up. Model diagnostics indicate an adequate convergence of the chains. We tested chain convergence by trace plots and the Gelman-Rubin statistic (Rhat < 1.1) (Gelman and Hill 2006). Bulk and tail effective sample sizes of the bulk and tail were greater than 400 for all parameters following Vehtari et al. 2019 recommendations. We graphically tested posterior predictive controls by comparing the observed outcome variable and the simulated data sets of the posterior predictive distribution. We considered that there is strong evidence of an effect when the 95% credible interval (CI) from the posterior distribution does not overlap 0.
Results
We compiled images from four satellites (L5, L7, L8 and MODIS) to reconstruct the NDVI time series (see Table S.1 in the Supplemental Material). We used 883 and 838 NDVI records to build the CHP model (transhumant livestock herbivory) and the CJ model (wild herbivory), respectively (Table S1). Thus, we obtained NDVI values and CI for the 1990–2019 period (see Fig. S3, S4 and Fig. S8 in the Supplemental Material). We choose the best models based on the lowest AICc for both study areas (Table S1 and S2), in which we assumed that all satellite records have the same hidden trajectory. The best Bayesian model structure (lowest AICc) used to retrieve the monthly NDVI indicates that imagery from different satellites has a non-constrained relation between states, non-constant growth rate, independent process errors, similar observation variance and no interactions (see Tables S2 and S3 in the Supplemental Material). Using the Bayesian model with the lowest AICc, we predicted the one-step-ahead (innovations) residuals to check the model fit. Residuals fall within the CI, they have no temporal trend (see Fig. S5 and S10 in the Supplemental Material) and follow the normality test (see Fig. S7 and S11 in the Supplemental Material), indicating the model fits the data.
In the CJ model (wild herbivores dominance) (Fig. 2), precipitation accumulated in the previous year was the only covariate that has a strong statistical effect on primary productivity (mean = 0.19, 95% CI [0.05, 0.033]). In contrast, in the CHP model (transhumant livestock dominance) (Fig. 2), the accumulated precipitation during the previous three months showed a strong positive statistical effect on primary productivity (0.29 [0.06, 0.51]). We found strong statistical support for the temperature effect on primary productivity in the CHP model (− 0.11 [− 0.27, 0.05]). In CJ, however, monthly temperature showed no strong statistical effect. These results suggest that primary productivity in CJ depends more on annual cumulative rainfall whereas primary productivity in CHP is more closely influenced by seasonal variations in climatic conditions.

Effect size and 95% credible intervals (C.I.) of the different covariates on the Normalized Difference Vegetation Index (NDVI) for the 1990–2019 period in, a: El Calar de Juana (CJ; wild herbivory area); b: Los Campos de Hernán Pelea (CHP; transhumant livestock herbivory area). All values are standardized to the same scale and range
As expected, primary productivity in both study areas showed a similar seasonal pattern. That is, both study zones have two productivity maxima, in spring and autumn, and two productivity minima, in winter and summer, which agrees with a typical Mediterranean climate seasonality (Fig. 3, Fig. S1, Fig S2). However, we also found significant differences between the primary productivity in most of the monthly values (Fig. 3). Furthermore, the maximum and minimum productivity values occur at different periods of the year for each study area. While in areas with only wild herbivory the maximum and minimum primary productivity occurs in May and September respectively, in the transhumant livestock area, maximum and minimum primary productivity occurs in June and August respectively. Therefore, the period of low primary productivity is longer in areas dominated by transhumant livestock than in areas with only wild herbivory. Interestingly, the annual NDVI oscillations (i.e., the difference between maxima and minima) are greater in CHP than in CJ. This pattern coincides with the results shown in Fig. 2, where the effect of seasonal precipitation is higher in CHP given that the intra-annual variability of biomass in CHP fluctuates more and more rapidly than in CJ.

Median monthly NDVI values obtained from the MARSS model of El Calar de Juana (CJ; wild herbivory area) in colour pink and Los Campos de Hernán Pelea (CHP; transhumant livestock herbivory area) in colour green. The NDVI value has been standardized to the same scale and range. The statistical significance is noted with the following codes: 0.001 '***', 0.01 '**' 0.05 '*'
Concerning the long-term trend of vegetation biomass, we found that CJ and CHP differ from each other. Specifically, we found that the vegetation biomass in CJ increased between 1990 and 2019 (2.32 × 10–3 [1.56 × 10–3, 3.08 × 10–3]). Whereas, in CHP, the mean growth rate is centred in zero (5.87 × 10–5 [-7.83 × 10–4, 8.68 × 10–4]), indicating a stable temporal pattern (Fig. 4). This shows that whereas in CJ the vegetation increases biomass throughout time, in CHP vegetation biomass has maintained stable. These results may indicate that while grazing and land use dynamics have been maintained over time in CHP, in CJ the dynamics between vegetation biomass, ungulates and climate have changed and resulted in a periodic increase in vegetation biomass, suggesting a tendency of shrub encroachment.

The long-term trend of the Normalized Difference Vegetation Index (NDVI) of the MARSS model time series of a.1: El Calar de Juana (CJ; wild herbivory area) and b.1: Los Campos de Hernán Pelea. Posterior distribution of the monthly NDVI increment built from a Bayesian linear model in a.2: El Calar de Juana and b.2: Los Campos de Hernán Pelea (CHP; transhumant livestock herbivory area). All values are standardized to the same scale and range
Discussion
Here, we highlight how ungulate herbivory in different grazing regimes (i.e., wild ungulates and transhumant livestock production) differently affects temporal trends of primary productivity and modulate the vegetation responses to climatic factors in semi-arid mountain areas. Where grazing intensity has decreased over time and only wild herbivores are present, we found that landscape-scale vegetation biomass has increased over the years. However, vegetation biomass remained stable throughout the study period (30 years) in areas where transhumant use was maintained. We also observed differences in the relative effect of climatic conditions and seasonality between areas with domestic and wild herbivory regimes. While temperature and seasonal precipitation affected the temporal dynamics of primary productivity in areas with transhumant livestock herbivory, we found that the variability of primary productivity mostly depended on annual precipitation in areas with only wild herbivory. These results highlight the importance of herbivores in defining how Mediterranean landscapes function and indicate how passive rewilding or livestock transhumant practices can affect the ecosystem's adaptability to climate conditions.
Climate and vegetation dynamics
We found that primary productivity dynamics in each study zone have fundamental differences in their responses to climate, albeit their similarity in geomorphological, geological, and edaphic characteristics, climatic conditions, and vegetation type. The long history of grazing, especially transhumance in CHP, has influenced the ecosystem, not only affecting its composition, but also shaping the structure of grassland soils and vegetation types. Both areas follow a seasonality pattern that characterises Mediterranean environments (Mooney et al. 1974). However, only in areas where livestock grazing remains, seasonal precipitation shows a positive effect on primary productivity and monthly temperature presents a negative effect. Indeed, grasslands react more immediately to climate effects than shrublands (Knapp and Smith 2001; Sala et al. 2012). Grassland species in semi-arid ecosystems present traits that support drought tolerance, although grasses usually rely on the upper layers of the soil profile. Therefore, unlike scrublands, they are highly determined by the amount of precipitation of the wet season (Knapp and Smith 2001; Sala et al. 2012) and the negative indirect effect of temperature from increased soil drought associated with higher evapotranspiration (Dulamsuren et al. 2013).
Although climate seasonality affects both grasslands and shrublands, the intensity of these effects can vary depending on vegetation type or herbivory intensity (Patton et al. 2007). In domestic pastoral areas, temporal oscillations in primary productivity occur earlier at the beginning of spring than in areas with only wild herbivory, which results in a month-to-month lag between wild in contrast to domestic grazing areas. Besides, the ecosystem primary productivity difference between spring and summer is higher in areas where livestock use has been maintained than in areas with only wild herbivory. To make optimal exploitation of existing resources, transhumant livestock moves to the mountainous zones in a period that matches the seasonal peaks of pasture productivity (Oteros-Rozas et al. 2012). Livestock consumes vegetation biomass at the time of maximum growth and before the dry season when drought events are common. This dynamic has resulted in highly adapted grasslands composed of plant species adapted to grazing that also benefit from the enrichment of available nutrients through the deposition of manure and urine (Bailey et al. 1996). Thus, the effect of seasonal precipitation and seasonal grazing is likely to be a major determinant factor of the intra-annual primary productivity variation in domestic herbivory areas, resulting in an adapted and sustainable transhumance-based grazing system.
In areas with only wild herbivory, where grazing intensity is much lower than it was 30 years ago, only the precipitation accumulated in the year has a positive effect on the primary productivity. The landscape has undergone a process of shrub encroachment, and the area covered by shrubs has now increased. The vegetation of areas with only wild herbivory has deeper root systems, allowing them to access deeper and less variable water supplies (Padilla and Pugnaire 2007). Besides, vegetation cover is denser than in grasslands and prevents soil moisture loss to a greater extent (Miranda et al. 2011). This makes primary productivity in areas with only wild herbivory less dependent on seasonal precipitation, although it still depends on annual precipitation.
Based on our results, we expect that changes in climate variability and the unpredictable occurrence of extreme weather events can interact with grazing to impact the sustainability of these transhumance-based grazing system. The stability and maintenance of the ecosystem processes of these landscapes depend on matching the seasonal movement of livestock with seasonal patterns. If precipitation becomes increasingly unpredictable and variable, this well-adapted grassland ecosystem would be destabilised because transhumant movements would not coincide with the ideal vegetation growing season. Moreover, if the effects of rising temperatures and precipitation anomalies are manifested through negative impacts on the water balance, this semi-arid grassland might be one of the most affected ecosystems by global climatic changes (Mowll et al. 2015). However, climate warming may have less effect on the persistence of trees or shrubs (Brown and Archer 1999; Dulamsuren et al. 2013) as they are less dependent on the water available on the upper layers of the soil profile.
Cultural landscapes and passive rewilding
In traditional grassland areas (transhumant), we observed a stable ecosystem functioning (i.e., greenness oscillation and tendency) over the 30-year studied period, indicating that it is a stable landscape and suggesting no sign of land degradation. Semi-arid mountain grasslands have a very long history of co-evolution between plant species and ungulates, as they have been used by native ungulates and for livestock feeding for many years. Vegetation in domestic pastoral areas is in constant renewal due to their use for livestock feeding and tends to show high productivity and nutrient content for ungulates (McNaughton 1979; Austrheim et al. 2014; Jarque-Bascuñana et al. 2022; Castillo-Garcia et al. 2022). Moreover, the main reason these ecosystems have been maintained over time is related to the ability of this traditional pastoral system to exploit apparently unproductive areas for livestock feed, playing a key role in food security (O’Mara 2012) and ecological sustainability. Grasslands maintained by human management where the grazing intensity has been adequate can result in very stable systems (Molinillo et al. 1997) and can be described as mature ecosystems (Rebollo and Gómez-Sal 2004). In Mediterranean environments with strong inter-annual variability, transhumant herbivores overlap productivity peaks and herbivore loads can be very high for short periods. The vegetation adapts to this scenario and the soil is enriched in structure, herbaceous perennials, and below-ground biomass (Gómez Sal 2017). Therefore, the studied transhumant system (i.e. CHP) presents livestock production using cultural and non-intensive practices (García-Ruiz et al. 2020), suggesting an environmentally sustainable management context.
In areas where only wild ungulates can access the vegetation, biomass has increased over time. In these areas, traditional land-use practices (transhumance) have been limited. This suggests that the landscape, after livestock exclusion, is undergoing secondary succession towards shrub encroachment because of the reduction in grazing pressure during the passive rewilding process (Nogués-Bravo et al. 2016; Pettorelli et al. 2019). We note that the increase in vegetation biomass can be detected in a relatively short period, which agrees with other studies on secondary succession (Molinillo et al. 1997; García-Ruiz et al. 2020; Baumann et al. 2020). Vegetation biomass shows no signs of temporal stabilisation in CJ, indicating that it is still in transition towards a different stage of the ecological succession. However, in Mediterranean mountains areas, landscape changes are slow (Pueyo and Beguería 2007; Bonet and Pausas 2012; Rey Benayas et al. 2015; Peña-Angulo et al. 2019) and these areas might remain as they are for decades.
The process of rewilding from land abandonment is a highly controversial issue in the scientific community. For instance, there are open questions on whether it is better to manage landscapes with low human activity, thus improving heterogeneity in landscape structure and configuration, or whether it is better to allow passive rewilding, enhancing landscape naturalization (García-Ruiz et al. 2020). According to some authors, the presence of human activity is related to increasing the complexity and biodiversity of these landscapes (Bauer et al. 2009; Lasanta et al. 2015). This human activity is the origin of most of the grassland ecosystem which provides multiple ecosystem services such as water supply and flow regulation, carbon storage, erosion control, climate mitigation, pollination, and cultural values (Li et al. 2018; Bengtsson et al. 2019; Jarque-Bascuñana et al. 2022). However, according to other authors, the natural recovery of scrublands and forests as a means of recovering the refuge of numerous plant and animal species is the best alternative for nature conservation (Navarro and Pereira 2015; Malhi et al. 2016; Pettorelli et al. 2018). Because different wild ungulates present browsing behaviour (i.e. red deer, Iberian ibex), land abandonment and the consequential replacement of grasslands with shrublands can potentially favour these wild ungulate species (Velamazán et al. 2023). Hence, although this natural succession implies a decrease in open grassland, the progressive establishment of shrublands can enhance the biodiversity of landscapes previously dominated by livestock farming (Speed et al. 2019).
The loss of some of these cultural landscapes seems inevitable as the abandonment of traditional uses is increasing, and thus passive rewilding. However, this process could be providing a unique opportunity to create more diverse landscapes. Warming has less effect on the persistence of trees or shrubs (Brown and Archer 1999; Dulamsuren et al. 2013) as they are less dependent on the water on the upper layers of the soil profile. Thus, by providing suitable landscape management, this process of shrub encroachment at intermediate levels could improve landscape heterogeneity and help buffer the effects of droughts and climatic oscillations.
Conclusion
The abandonment of traditional uses is increasing, and consequently, passive rewilding is expected to occur in different regions. Although domestic and wild ungulates may partially present similarities in their roles in trophic interactions through ingestion of excess plant tissue or by affecting nutrient cycling in grazed areas (Milchunas et al. 1989; Veblen et al. 2016; Barbosa et al. 2020), they also present important differences on their effects on vegetation. We, therefore, suggest focusing management efforts on maintaining both systems and evaluating transhumant landscapes as a form of sustainable ecological management that can also enhance food security. In addition, we should have special attention to areas where passive rewilding is taking place as areas for refuge and habitat for other types of wild species. With the increasing frequency of extreme weather events and rising temperatures, more studies should question how these different landscapes will be conserved in the long-term. In this context, our study helps to advance our understanding of vegetation dynamics in the context of different herbivory scenarios and under a framework of long-term climate patterns that ultimately may provide information for decision making in the management of Mediterranean landscapes.
References
Aguilera-Alcalá N, Arrondo E, Pascual-Rico R et al (2022) The value of transhumance for biodiversity conservation: vulture foraging in relation to livestock movements. Ambio 51:1330–1342
Augustine DJ, McNaughton SJ (1998) Ungulate effects on the functional species composition of plant communities: herbivore selectivity and plant tolerance. J Wildl Manage 62:1165
Austrheim G, Speed JDM, Martinsen V et al (2014) Experimental effects of herbivore density on aboveground plant biomass in an alpine grassland ecosystem. Arctic, Antarct Alp Res 46:535–541
Babcock C, Finley AO, Looker N (2021) A Bayesian model to estimate land surface phenology parameters with harmonized Landsat 8 and Sentinel-2 images. Remote Sens Environ. https://doi.org/10.1016/j.rse.2021.112471
Bailey DW, Gross JE, Laca EA et al (1996) Mechanisms that result in large herbivore grazing distribution patterns. J Range Manag. https://doi.org/10.2307/4002919
Barbosa JM, Pascual-Rico R, Eguia Martínez S, Sánchez-Zapata JA (2020) Ungulates attenuate the response of mediterranean mountain vegetation to climate oscillations. Ecosystems 23:957–972.
Bauer N, Wallner A, Hunziker M (2009) The change of European landscapes: human-nature relationships, public attitudes towards rewilding, and the implications for landscape management in Switzerland. J Environ Manage 90:2910–2920.
Baumann M, Kamp J, Pötzschner F et al (2020) Declining human pressure and opportunities for rewilding in the steppes of Eurasia. Divers Distrib 26:1058–1070.
Benavente A (2008) Flora y vegetacion: parque natural de las sierras de cazorla, segura y Las Villas. Anu Del Adelantamiento Cazorla 50:149–153
Bengtsson J, Bullock JM, Egoh B et al (2019) Grasslands—more important for ecosystem services than you might think. Ecosphere. https://doi.org/10.1002/ecs2.2582
Bonet A, Pausas JG (2012) Species Richness and cover along a 60-year chronosequence in old-fields of Southeastern Spain Author ( s ): Andreu Bonet and Juli G. Pausas reviewed work ( s ): Published by : Springer along. Plant Ecol 174:257–270
Bowen ME, McAlpine CA, House APN, Smith GC (2007) Regrowth forests on abandoned agricultural land: a review of their habitat values for recovering forest fauna. Biol Conserv 140:273–296
Brown JR, Archer S (1999) Shrub invasion of grassland: recruitment is continuous and not regulated by herbaceous biomass or density. Ecology 80:2385–2396
Buchhorn M, Lesiv M, Tsendbazar N-E et al (2020) Copernicus global land cover layers—collection 2. Remote Sens 12:1044
Bürkner PC (2021) Bayesian item response modeling in R with brms and Stan. J Stat Softw. https://doi.org/10.18637/JSS.V100.I05
Castillo-Garcia M, Alados CL, Ramos J et al (2022) Understanding herbivore-plant-soil feedbacks to improve grazing management on Mediterranean mountain grasslands. Agric Ecosyst Environ 327:107833.
Doughty CE (2017) Herbivores increase the global availability of nutrients over millions of years. Nat Ecol Evol 1:1820–1827
Dulamsuren C, Wommelsdorf T, Zhao F et al (2013) Increased summer temperatures reduce the growth and regeneration of Larix sibirica in southern boreal forests of Eastern Kazakhstan. Ecosystems 16:1536–1549.
Dupke C, Bonenfant C, Reineking B et al (2017) Habitat selection by a large herbivore at multiple spatial and temporal scales is primarily governed by food resources. Ecography (cop) 40:1014–1027.
Ellison L (1960) Influence of grazing on plant succession of Rangelands. Bot Rev 26:1–78.
Frank D, Inouye R, Huntly N et al (1994) The biogeochemistry of a north-temperate grassland with native ungulates: nitrogen dynamics in Yellowstone National Park. Biogeochemistry. https://doi.org/10.1007/BF00002905
Frank DA, Wallen RL, White PJ (2016) Ungulate control of grassland production: grazing intensity and ungulate species composition in Yellowstone Park. Ecosphere. https://doi.org/10.1002/ecs2.1603
Gamon JA, Field CB, Goulden ML et al (1995) Relationships between NDVI, canopy structure, and photosynthesis in three californian vegetation types. Ecol Appl 5:28–41.
García-Ruiz JM, Lasanta T, Nadal-Romero E et al (2020) Rewilding and restoring cultural landscapes in Mediterranean mountains: Opportunities and challenges. Land Use Policy. https://doi.org/10.1016/j.landusepol.2020.104850
Gelman A, Hill J (2006) Data Analysis Using Regression and Multilevel/Hierarchical Models. Cambridge University Press, Cambridge
Giguet-Covex C, Pansu J, Arnaud F et al (2014) Long livestock farming history and human landscape shaping revealed by lake sediment DNA. Nat Commun 5:3211
Gómez Sal A (2017) Patterns of Vegetation Cover Shaping the Cultural Landscapes in the Iberian Peninsula. The Vegetation of the Iberian Peninsula. Springer International Publishing, Cham, pp 459–497
Gómez-Mercado F (2011) Vegetación y flora de la Sierra de Cazorla. Guineana 17:1–481
Gorelick N, Hancher M, Dixon M et al (2017) Google Earth Engine: Planetary-scale geospatial analysis for everyone. Remote Sens Environ 202:18–27.
Hamel S, Garel M, Festa-Bianchet M et al (2009) Spring normalized difference vegetation index (NDVI) predicts annual variation in timing of peak faecal crude protein in mountain ungulates. J Appl Ecol. https://doi.org/10.1111/j.1365-2664.2009.01643.x
Hernández F, Ríos C, Perotto-Baldivieso HL (2019) Evolutionary history of herbivory in the Patagonian steppe: the role of climate, ancient megafauna, and guanaco. Quat Sci Rev 220:279–290
Hinrichsen R, Holmes E (2009) Using multivariate state-space models to study spatial structure and dynamics. In: Cantrel S, Cosner C, Shinghui R (eds) Spatial Ecology. CRC Press, pp 145–165
Hobbs NT (1996) Modification of ecosystems by ungulates. J Wildl Manage 60:695.
Holmes EE, Ward Eric J, Wills K (2012) MARSS: multivariate autoregressive state-space models for analyzing time-series data. R J. https://doi.org/10.32614/RJ-2012-002
Holmes EE (2014) Computation of Standardized Residuals for MARSS Models
Hu Z, Li S, Guo Q et al (2016) A synthesis of the effect of grazing exclusion on carbon dynamics in grasslands in China. Glob Chang Biol 22:1385–1393
Huete A, Didan K, Miura T et al (2002) Overview of the radiometric and biophysical performance of the MODIS vegetation indices. Remote Sens Environ 83:195–213
Hurvich CM, Tsai C-L (1989) Regression and time series model selection in small samples. Biometrika 76:297–307
IUSS Working Group WRB (2015) Base referencial mundial del recurso suelo 2014, Actualización 2015. Sistema internacional de clasificación de suelos para la nomenclatura de suelos y la creación de leyendas de mapas de suelos. Informes sobre recursos mundiales de suelos 106. FAO, Roma
Jarque-Bascuñana L, Calleja JA, Ibañez M et al (2022) Grazing influences biomass production and protein content of alpine meadows. Sci Total Environ 818:151771.
Kaarlejärvi E, Hoset KS, Olofsson J (2015) Mammalian herbivores confer resilience of Arctic shrub-dominated ecosystems to changing climate. Glob Chang Biol 21:3379–3388.
Knapp AK, Smith MD (2001) Variation among biomes in temporal dynamics of aboveground primary production. Science (80-) 291:481–484
Lasanta T, Nadal-Romero E, Arnáez J (2015) Managing abandoned farmland to control the impact of re-vegetation on the environment. The state of the art in Europe. Environ Sci Policy 52:99–109
Li W, Li X, Zhao Y et al (2018) Ecosystem structure, functioning and stability under climate change and grazing in grasslands: current status and future prospects. Curr Opin Environ Sustain 33:124–135
MacDonald D, Crabtree J, Wiesinger G et al (2000) Agricultural abandonment in mountain areas of Europe: environmental consequences and policy response. J Environ Manage 59:47–69
Maestre FT, Le Bagousse-Pinguet Y, Delgado-Baquerizo M et al (2022) Grazing and ecosystem service delivery in global drylands. Science (80-) 378:915–920
Malhi Y, Doughty CE, Galetti M et al (2016) Megafauna and ecosystem function from the Pleistocene to the Anthropocene. Proc Natl Acad Sci 113:838–846
Martínez Martínez T (2002) Comparison and overlap of sympatric wild ungulate diet in Cazorla, segura and las Villas Natural Park. Pirineos 157:103–116
McNaughton SJ (1979) Grazing as an optimization process: Grass-ungulate relationships in the Serengeti. Am Nat 113:691–703
McNaughton SJ (1985) Ecology of a grazing ecosystem: The Serengeti. Ecol Monogr 55:259–294
Milchunas DG, Lauenroth WK (1993) Quantitative effects of grazing on vegetation and soils over a global range of environments. Ecol Monogr 63:327–366.
Milchunas DG, Lauenroth WK, Chapman PL, Kazempour MK (1989) Effects of grazing, topography, and precipitation on the structure of a semiarid grassland. Vegetatio. https://doi.org/10.1007/BF00049137
Miranda JDD, Armas C, Padilla FMM, Pugnaire FII (2011) Climatic change and rainfall patterns: effects on semi-arid plant communities of the Iberian Southeast. J Arid Environ 75:1302–1309
Molinillo M, Lasanta T, García-Ruiz JM (1997) Managing mountainous degraded landscapes after farmland abandonment in the Central Spanish Pyrenees. Environ Manage 21:587–598
Mooney HA, Parsons DJ, Kummerow J (1974) Plant Development in Mediterranean Climates. In: Lieth H (ed) Phenology and Seasonality Modelling. Ecological Studies, vol 8. Springer, Berlin, Heidelberg. https://doi.org/10.1007/978-3-642-51863-8_22
Mowll W, Blumenthal DM, Cherwin K et al (2015) Climatic controls of aboveground net primary production in semi-arid grasslands along a latitudinal gradient portend low sensitivity to warming. Oecologia 177:959–969.
Navalgund RR, Jayaraman V, Roy PS (2007) Remote sensing applications : an overview. Curr Sci Assoc 93:1747–1766
Navarro LM, Pereira HM (2015) Rewilding Abandoned Landscapes in Europe. Rewilding European Landscapes. Springer International Publishing, Cham, pp 3–23
Niu K, He J-S, Lechowicz MJ (2016) Grazing-induced shifts in community functional composition and soil nutrient availability in Tibetan alpine meadows. J Appl Ecol 53:1554–1564
Nogués-Bravo D, Simberloff D, Rahbek C, Sanders NJ (2016) Rewilding is the new Pandora’s box in conservation. Curr Biol 26:R87–R91
O’Mara FP (2012) The role of grasslands in food security and climate change. Ann Bot 110:1263–1270
Olofsson J, Post E (2018) Effects of large herbivores on tundra vegetation in a changing climate, and implications for rewilding. Philos Trans R Soc B Biol Sci 373:20170437
Oteros-Rozas E, González JA, Martín-López B et al (2012) Evaluating Ecosystem Services in Transhumance Cultural LandscapesAn Interdisciplinary and Participatory Framework. GAIA: Ecol Perspect Sci Soc 21:185–193
Padilla FM, Pugnaire FI (2007) Rooting depth and soil moisture control Mediterranean woody seedling survival during drought. Funct Ecol. https://doi.org/10.1111/j.1365-2435.2007.01267.x
Patton BD, Dong X, Nyren PE, Nyren A (2007) Effects of grazing intensity, precipitation, and temperature on forage production. Rangel Ecol Manag 60:656–665
Peña-Angulo D, Khorchani M, Errea P et al (2019) Factors explaining the diversity of land cover in abandoned fields in a Mediterranean mountain area. Catena 181:104064
Perevolotsky A, Seligman NG (1998) Role of grazing in mediterranean rangeland ecosystems. Bioscience 48:1007–1017
Pettorelli N, Ryan S, Mueller T et al (2011) The normalized difference vegetation index (NDVI): unforeseen successes in animal ecology. Clim Res 46:15–27
Pettorelli N, Barlow J, Stephens PA et al (2018) Making rewilding fit for policy. J Appl Ecol 55:1114–1125
Pettorelli N, Durant SM, du Toit JT (2019) Rewilding: a captivating, controversial, twenty-first-century concept to address ecological degradation in a changing world. In: Rewilding. Cambridge University Press, pp 1–11
Pueyo Y, Beguería S (2007) Modelling the rate of secondary succession after farmland abandonment in a Mediterranean mountain area. Landsc Urban Plan 83:245–254
Rebollo S, Gómez-Sal A (2004) Aprovechamiento sostenible de los pastizales. Ecosistemas 12:10
Rey Benayas JM, Martínez-Baroja L, Pérez-Camacho L et al (2015) Predation and aridity slow down the spread of 21-year-old planted woodland islets in restored Mediterranean farmland. New for 46:841–853
Sala OE, Gherardi LA, Reichmann L et al (2012) Legacies of precipitation fluctuations on primary production: theory and data synthesis. Philos Trans R Soc B Biol Sci 367:3135–3144
San Miguel-Ayanz A, Perea García-Calvo A, Fernández-Olalla R (2010) Wild Ungulates vs. extensive livestock. Looking back to face the future. Options Meditérr 92:27–34
Speed JDM, Austrheim G, Kolstad AL, Solberg EJ (2019) Long-term changes in northern large-herbivore communities reveal differential rewilding rates in space and time. PloS one 14:e0217166.
Sullivan S, Rohde R (2002) On Non-equilibrium in arid and semi-arid grazing systems. J Biogeogr 29:1595–1618
Tebaldi C, Knutti R (2010) Climate Models and Their Projections of Future Changes. In: Advances in Global Change Research. Springer International Publishing, pp 31–56
Valle Tendero F, Navarro Reyes FB, Jiménez Morales MN et al (2004) Datos botánicos aplicados a la Gestión del Medio Natural Andaluz I: Bioclimatología y Biogeografía. Junta de Andalucia. Consejería de Medio Ambiente, Sevilla
Veblen KE, Porensky LM, Riginos C, Young TP (2016) Are cattle surrogate wildlife? Savanna plant community composition explained by total herbivory more than herbivore type. Ecol Appl. https://doi.org/10.1890/15-1367.1
Vehtari A, Gelman A, Simpson D, et al (2019) Rank-normalization, folding, and localization: An improved Rb for assessing convergence of MCMC. arXiv
Velamazán M, Sánchez-Zapata JA, Moral-Herrero R et al (2023) Contrasting effects of wild and domestic ungulates on fine-scale responses of vegetation to climate and herbivory. Landsc Ecol. https://doi.org/10.1007/s10980-023-01676-0
Vetter S (2005) Rangelands at equilibrium and non-equilibrium: recent developments in the debate. J Arid Environ 62:321–341
Ward EJ, Chirakkal H, González-Suárez M et al (2010) Inferring spatial structure from time-series data: using multivariate state-space models to detect metapopulation structure of California sea lions in the Gulf of California, Mexico. J Appl Ecol 47:47–56.
Xu C, Li Y, Hu J et al (2012) Evaluating the difference between the normalized difference vegetation index and net primary productivity as the indicators of vegetation vigor assessment at landscape scale. Environ Monit Assess 184:1275–1286.
Acknowledgements
We would like to thank Paloma Prieto, David Cuerda and Francisco Martinez, and the Sierras de Cazorla, Segura, and Las Villas Natural Park for their support in field campaigns and historical information. We would like to thank Evan Alexander Netherton Marks and Jorge Mataix Solera for their assessment on lithology, geology, and soil characterisation. We also like to thank the Agrupación de Defensa Sanitaria Ganadera Los Campos (ADSG) for the provided information on livestock production.
Funding
Open Access funding provided thanks to the CRUE-CSIC agreement with Springer Nature. This study forms part of the AGROALNEXT (2022/038) programme and was supported by MCIN/AEI/https://doi.org/10.13039/501100011033 with funding from European Union NextGenerationEU (PRTR-C17.I1) and Generalitat Valenciana. This study also forms part of the DIGITALPAST (TED2021-130005B-C21) project and was supported by MCIN/AEI/https://doi.org/10.13039/501100011033 with funding from Plan NextGenerationEU. This study also forms part of the TRASCAR (RTI2018-099609-B-C21) project and was supported by MCIN/AEI/https://doi.org/10.13039/501100011033 with funding from European Regional Development Fund (ERDF). JMB and MRM were supported by Generalitat Valenciana with the program PlanGenT (CIDEGENT/2020/030). X.B. was supported by Agencia Estatal de Investigación for Grant PID2019-106341 GB-I00 (jointly financed by the European Regional Development Fund, FEDER) supported by the MCIN/AEI/https://doi.org/10.13039/501100011033.
Author information
Authors and Affiliations
Contributions
MRM, JASZ and JMB defined the study conception and design. MRM, XB and JMB contributed to the data analysis. MRM, JASZ and JMB wrote the manuscript. All authors read and approved the final version of the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare they have no financial interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Rincon-Madroñero, M., Sánchez-Zapata, J.A., Barber, X. et al. Long-term vegetation responses to climate depend on the distinctive roles of rewilding and traditional grazing systems. Landsc Ecol 39, 1 (2024). https://doi.org/10.1007/s10980-024-01806-2
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s10980-024-01806-2