
Abstract
Biological processes rely on interactions between many binding partners. Binding results in the modulation of the conformational landscape of the interacting molecules, a phenomenon rooted in folding and binding cooperativity underlying the allosteric functional regulation of biomacromolecules. The conformational equilibrium of a protein and the binding equilibria of different interacting and cooperative ligands are coupled giving rise to a complex scenario in which protein function can be finely tuned and modulated. Binding cooperativity and allostery add additional levels of complexity in protein function regulation. Here we will review some important concepts associated with binding, cooperativity and allostery in protein interactions, illustrated with several representative protein-dependent biological systems related to drug discovery and physiological mechanisms characterization and studied by isothermal titration calorimetry.





Similar content being viewed by others


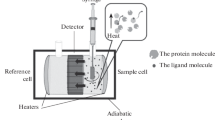
References
Wyman J, Gill SJ. Binding and linkage: functional chemistry of biological macromolecules. Mill Valley: University Science Books; 1990.
Schellman JA. Macromolecular binding. Biopolymers. 1975;14:999–1018.
Wyman J. The binding potential, a neglected linkage concept. J Mol Biol. 1965;11:631–44.
Wyman J. Linked functions and reciprocal effects in hemoglobin—A second look. Adv Protein Chem. 1964;19:223–86.
Velazquez-Campoy A. Geometric features of the Wiseman isotherm in isothermal titration calorimetry. J Thermal Anal Calorim. 2015;122:1477–83.
Velazquez-Campoy A. Allostery and cooperative interactions in proteins assessed by isothermal titration calorimetry. In: Bastos M, editor. Biocalorimetry—foundations and contemporary approaches. Boca Raton: CRC Press; 2016. p. 223–46.
Vega S, Abian A, Velazquez-Campoy A. A unified framework based on the binding polynomial for characterizing biological systems by isothermal titration calorimetry. Methods. 2015;76:99–115.
Freire E, Schön A, Velazquez-Campoy A. Isothermal titration calorimetry: general formalism using binding polynomials. Methods Enzymol. 2009;455:127–55.
Bohr C, Hasselbalch K, Krogh A. Ueber einen in biologischer beziehung wichtigen einfluss, den die kohlensäurespannung des blutes auf dessen sauerstoffbindung übt. Skand Archiv Physiol (Acta Physiol). 1904;16:402–12.
Monod J, Wyman J, Changeux JP. On the nature of allosteric transitions a plausible model. J Mol Biol. 1965;12:88–118.
Monod J, Changeux JP, Jacob F. Allosteric proteins and cellular control systems. J Mol Biol. 1963;6:306–29.
Gunasekaran K, Ma B, Nussinov R. ls allostery an intrinsic property of all dynamic proteins? Proteins. 2004;57:433–43.
Eftink M, Biltonen RL. Thermodynamics of interacting biological systems. Beezer AE, Biological calorimetry”. London: Academic Press; 1980. p. 343–412.
Courter JR, Madani N, Sodroski J, Schön A, Freire E, Kwong PD, Hendrickson WA, Chaiken IM, LaLonde JM, Smith AB 3rd. Structure-based design, synthesis and validation of CD4-mimetic small molecule inhibitors of HIV-1 entry: conversion of a viral entry agonist to an antagonist. Acc Chem Res. 2014;47:1228–37.
Williams R, Holyoak T, McDonald G, Gui C, Fenton AW. Differentiating a ligand’s chemical requirements for allosteric interactions from those for protein binding. Phenylalanine inhibition of pyruvate kinase. Biochemistry. 2006;45:5421–9.
Jobichen C, Fernandis AZ, Velazquez-Campoy A, Leung KY, Mok YK, Wenk MR, Sivaraman J. Identification and characterization of the lipid-binding property of GrlR, a locus of enterocyte effacement regulator. Biochem J. 2009;13(420):191–9.
Rodriguez-Cardenas A, Rojas AL, Conde-Gimenez M, Velazquez-Campoy A, Hurtado-Guerrero R, Sancho J. Streptococcus pneumoniae TIGR4 flavodoxin: structural and biophysical characterization of a novel drug target. PLoS ONE. 2016;11:e0161020.
Cremades N, Velazquez-Campoy A, Freire E, Sancho J. The flavodoxin from Helicobacter pylori: structural determinants of thermostability and FMN cofactor binding. Biochemistry. 2008;47:627–39.
Irun MP, Garcia-Mira MM, Sanchez-Ruiz JM, Sancho J. Native hydrogen bonds in a molten globule: the apoflavodoxin thermal intermediate. J Mol Biol. 2001;306:877–88.
Wyman J. Heme proteins. Adv Protein Chem. 1948;4:407–531.
Wyman J. Allosteric linkage. J Am Chem Soc. 1967;89:2202–18.
Koshland DE Jr, Nemethy G, Filmer D. Comparison of experimental binding data and theoretical models in proteins containing subunits. Biochemistry. 1966;5:365–85.
Di Cera E, Gill SJ, Wyman J. Binding capacity: cooperativity and buffering in biopolymers. Proc Natl Acad Sci USA. 1988;85:449–52.
Ji Y, Postis VL, Wang Y, Bartlam M, Goldman A. Transport mechanism of a glutamate transporter homologue GltPh. Biochem Soc Trans. 2016;44:898–904.
Groeneveld M, Slotboom DJ. Na(+): aspartate coupling stoichiometry in the glutamate transporter homologue Glt(Ph). Biochemistry. 2010;49:3511–3.
Boudker O, SeCheol O. Isothermal titration calorimetry of ion-coupled membrane transporters. Methods. 2015;76:171–82.
Kohl NE, Emini EA, Schleif WA, Davis LJ, Heimbach JC, Dixon RA, Scolnick EM, Sigal IS. Active human immunodeficiency virus protease is required for viral infectivity. Proc Natl Acad Sci USA. 1988;85:4686–90.
Brik A, Wong C-H. HIV-1 protease: mechanism and drug discovery. Org Biomol Chem. 2003;1:5–14.
Velazquez-Campoy A, Luque I, Todd MJ, Milutinovich M, Kiso Y, Freire E. Thermodynamic dissection of the binding energetics of KNI-272, a potent HIV-1 protease inhibitor. Protein Sci. 2000;9:1801–9.
Baker BM, Murphy KP. Evaluation of linked protonation effects in protein binding reactions using isothermal titration calorimetry. Biophys J. 1996;71:2049–55.
Krishnamurthy R. Role of pKa of nucleobases in the origins of chemical evolution. Acc Chem Res. 2012;45:2035–44.
Ross D, Siegel D. Functions of NQO1 in cellular protection and CoQ10 metabolism and its potential role as a redox sensitive molecular switch. Front Physiol. 2017;8:595.
Pey AL, Megarity CF, Timson DJ. FAD binding overcomes defects in activity and stability displayed by cancer-associated variants of human NQO1. Biochim Biophys Acta—Mol Basis Dis. 2014;1842:2163–73.
Claveria-Gimeno R, Velazquez-Campoy A, Pey AL. Thermodynamics of cooperative binding of FAD to human NQO1: implications to understanding cofactor-dependent function and stability of the flavoproteome. Arch Biochem Biophys. 2017;636:17–27.
Lee AL, Sapienza PJ. Thermodynamic and NMR assessment of ligand cooperativity and intersubunit communication in symmetric dimers: application to thymidylate synthase. Front Mol Biosci. 2018;5:47.
Tomei L, Failla C, Santolini E, De Francesco R, La Monica N. NS3 is a serine protease required for processing of hepatitis C virus polyprotein. J Virol. 1993;67:4017–26.
Kwong AD, Kim JL, Rao G, Lipovsek D, Raybuck SA. Hepatitis C virus NS3/4A protease. Antivir Res. 1998;40:1–18.
Martinez-Julvez M, Abian O, Vega S, Medina M, Velazquez-Campoy A. Studying the allosteric energy cycle by isothermal titration calorimetry. Methods Mol Biol. 2012;796:53–70.
Martinez-Julvez M, Medina M, Velazquez-Campoy A. Binding thermodynamics of ferredoxin:NADP + reductase: two different protein substrates and one energetics. Biophys J. 2009;96:4966–75.
Velazquez-Campoy A, Goñi G, Peregrina JR, Medina M. Exact analysis of heterotropic interactions in proteins: characterization of cooperative ligand binding by isothermal titration calorimetry. Biophys J. 2006;91:1887–904.
Du W, Liu W-S, Payne DJ, Doyle ML. Synergistic inhibitor binding to Streptococcus pneumoniae 5-enolpyruvylshikimate-3-phosphate synthase with both monovalent cations and substrate. Biochemistry. 2000;39:10140–6.
Jonckheere AI, Smeitink JAM, Rodenburg RJT. Mitochondrial ATP synthase: architecture, function and pathology. J Inherit Metab Dis. 2012;35:211–25.
Pulido NO, Salcedo G, Perez-Hernandez G, Jose-Nuñez C, Velazquez-Campoy A, Garcia-Hernandez E. Energetic effects of magnesium in the recognition of adenosine nucleotides by the F(1)-ATPase beta subunit. Biochemistry. 2010;49:5258–68.
Salcedo G, Cano-Sanchez P, Tuena de Gomez-Puyou M, Velazquez-Campoy A, Garcia-Hernandez E. Isolated noncatalytic and catalytic subunits of F1-ATPase exhibit similar, albeit not identical, energetic strategies for recognizing adenosine nucleotides. Biochim Biophys Acta—Bioenerg. 2014;1837:44–50.
Peselis A, Gao A, Serganov A. Cooperativity, allostery and synergism in ligand binding to riboswitches. Biochimie. 2015;117:100–9.
Seo M-H, Park J, Kim E, Hohng S, Kim H-S. Protein conformational dynamics dictate the binding affinity for a ligand. Nat Commun. 2014;5:3724.
Guo J, Zhou H-X. Protein Allostery and conformational dynamics. Chem Rev. 2016;116:6503–15.
Riera TV, Zheng L, Josephine HR, Min D, Yang W, Hedstrom L. Allosteric activation via kinetic control: potassium accelerates a conformational change in IMP dehydrogenase. Biochemistry. 2011;50:8508–18.
Stock G, Hamm P. A non equilibrium approach to allosteric communication. Philos Trans R Soc Lond B Biol Sci. 2018;373:20170187.
Nussinov R, Tsai C-J. Allostery in disease and drug discovery. Cell. 2013;153:293–305.
Abdel-Magid AF. Allosteric modulators: an emerging concept in drug discovery. ACS Med Chem Lett. 2015;6:104–7.
Lu S, Ji M, Ni D, Zhang J. Discovery of hidden allosteric sites as novel targets for allosteric drug design. Drug Discov Today. 2018;23:359–65.
Ni D, Lu S, Zhang J. Emerging roles of allosteric modulators in the regulation of protein-protein interactions (PPIs): a new paradigm for PPI drug discovery. Med Res Rev. 2019. https://doi.org/10.1002/med.21585.
Hilser VJ, Wrabl JO, Motlagh HN. Structural and energetic basis of allostery. Annu Rev Biophys. 2012;41:585–609.
Li J, Hilser VJ. Assessing allostery in intrinsically disordered proteins with ensemble allosteric model. Methods Enzymol. 2018;611:531–57.
Zhang L, Li M, Liu Z. A comprehensive ensemble model for comparing the allosteric effect of ordered and disordered proteins. PLoS Comput Biol. 2018;14:e1006393.
Hilser VJ, Thompson EB. Intrinsic disorder as a mechanism to optimize allosteric coupling in proteins. Proc Natl Acad Sci USA. 2007;104:8311–5.
Berlow RB, Dyson HJ, Wright PE. Expanding the paradigm: intrinsically disordered proteins and allosteric regulation. J Mol Biol. 2018;430:2309–20.
Li J, White JT, Saavedra H, Wrabl JO, Motlagh HN, Liu K, Sowers J, Schroer TA, Thompson EB, Hilser VJ. Genetically tunable frustration controls allostery in an intrinsically disordered transcription factor. eLife. 2017;6:e30688.
Felix J, Weinhäupl K, Chipot C, Dehez F, Hessel A, Gauto DF, Morlot C, Abian O, Gutsche I, Velazquez-Campoy A, Schanda P, Fraga H. Mechanism of the allosteric activation of the ClpP protease machinery by substrates and active-site inhibitors. Sci Adv. 2019. https://doi.org/10.1101/578260.
Taneva SG, Bañuelos S, Falces J, Arregi I, Muga A, Konarev PV, Svergun DI, Velazquez-Campoy A, Urbaneja MA. A mechanism for histone chaperoning activity of nucleoplasmin: thermodynamic and structural models. J Mol Biol. 2009;393:448–63.
Acknowledgements
This work was supported by Miguel Servet Program from Instituto de Salud Carlos III (CPII13/00017 to OA); Fondo de Investigaciones Sanitarias from Instituto de Salud Carlos III, and European Union (ERDF/ESF, ‘Investing in your future’) (PI15/00663 and PI18/00349 to OA); Spanish Ministry of Economy and Competitiveness (BFU2016-78232-P to AVC); Diputación General de Aragón (Protein Targets and Bioactive Compounds Group E45_17R to AVC, and Digestive Pathology Group B25_17R to OA); and Centro de Investigación Biomédica en Red en Enfermedades Hepáticas y Digestivas (CIBERehd).
Author information
Authors and Affiliations
Corresponding authors
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Appendix
Appendix
The overall association constants β i can be expressed as a function of β 1
It can be demonstrated that the overall stoichiometric association constant βsi for the formation of complex PsAi can be expressed as a function of the total number of binding sites, n, the number of ligand A molecules bound per macromolecule, i, the intrinsic site-specific association constant for ligand A, βs1 (that is, the overall stoichiometric association constant for the formation of complex PsA), and the cooperativity constant accounting for homotropic cooperativity effects for ligand A binding, αsi:
If a protein conformational state P has n identical and independent binding sites for ligand A, the binding polynomial factorizes into identical terms, each one representing one of the ligand binding sites as a subsystem:
where K is the intrinsic site-specific association constant for each of the ligand binding sites. From that expression, it is obvious that:
with:
and from that:
Then, potential homotropic cooperative effects can be accounted for by introducing an additional factor αi, with α1 = 1, because there is no homotropic effect for the first bound ligand:
This expression is completely general and it can be applied to any situation (i.e., identical or nonidentical binding sites, cooperative or non-cooperative sites) by decomposing β1 and αi into additional factors [8]. Such decomposition will help in reducing the number of binding parameters required in the model [61].
From Eq. (46), the binding polynomial can be factorized into identical terms if, for all i’s, it is fulfilled that [1]:
And it can be factorized into nonidentical terms if, for all i’s, it is fulfilled that [1]:
Therefore, the ligand binding sites are identical and independent if:
and the ligand binding sites are nonidentical and independent if:
Thus, the independent and identical nature of the ligand binding sites can be judged by comparing the overall association constants (β’s) or the homotropic cooperativity parameters (α’s).
The extent of the heterotropic effect between ligand A and ligand B is fully reciprocal
The heterotropic effect between two ligands is fully reciprocal or symmetric. Because of the energy conservation principle, if ligand B modifies the intrinsic affinity for ligand A by a factor αij, then ligand A causes the same effect on ligand B. The constant βij can be split into two factors: βij = βi0βj/i, where βi0 is the overall association constant for P + Ai ↔ PAi and βj/i is the overall association constant for PAi + Bj ↔ PAiBj. Alternatively, βij can be split into two factors: βij = β0jβi/j, where βoj is the overall association constant for P + Bj ↔ PBj and βi/j is the overall association constant for PBj + Ai ↔ PAiBj. Furthermore, the constants βj/i and βi/j can be factorized as: βj/i = β0jαij and βi/j = βi0αij.
The maximal allosteric effect occurs at concentrations around the dissociation constant of the ligand
From Eq. 20, if, for the sake of simplicity, only a single binding site for ligand B is considered:
ΔnLB,s,B will be zero for zero and infinite ligand B concentration, and for βs = β0. Deriving with respect to ligand B concentration:
and a maximal value for ΔnLB,s,B will be achieved when:
whose solutions are: (1) βs = β0 and any value of [B] (trivial case); or (2) βs ≠ β0 and [B] = (βsβ0)−1/2, the inverse of the geometric mean of the overall association constants, which is the concentration of ligand B for a maximal allosteric effect on the apparent conformational constant \( \gamma_{\text{s}}^{\text{app}} \) for Ps.
The maximal heterotropic effect occurs at concentrations around the dissociation constant of the secondary ligand
From Eq. 25, if there are several (i = 1…n) binding sites for ligand A and, for the sake of simplicity, a single binding site for ligand B:
ΔnLB,i,B will be zero for zero and infinite ligand B concentration, and for αi1 = 1. Deriving with respect to ligand B concentration:
and a maximal value for ΔnLB,B will be achieved when:
whose solutions are: (1) αi1 = 1 and any value of β01 and [B] (trivial case); or (2) αi1 ≠ 1 and [B] = β−101α−1/2i1, which is the concentration of ligand B for a maximal heterotropic effect on the apparent overall association constant \( \beta_{\text{i}}^{\text{app}} \) for PAi.
Enthalpy changes associated with equilibrium constants are calculated by applying the van’t Hoff equation
Every equilibrium constants has an associated enthalpy change (besides changes in other thermodynamic potentials). The corresponding enthalpy change can be calculated by applying the van’t Hoff equation:
From that, if a given equilibrium constant is a function of other equilibrium constants, then, the corresponding enthalpy can be calculated by applying Eq. 53:
Rights and permissions
About this article

Cite this article
Vega, S., Abian, O. & Velazquez-Campoy, A. Handling complexity in biological interactions. J Therm Anal Calorim 138, 3229–3248 (2019). https://doi.org/10.1007/s10973-019-08610-0
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10973-019-08610-0