
Abstract
Changes in local hydrological and climatic conditions over the last 5500 years have been reconstructed based on geochemical and paleobiological features recorded in the sediments of two mountain lakes. The lakes are located in the Tatra Mountains, in the highest mountain range of the Carpathians (Central Europe), which during the Holocene constituted an important climatic barrier. Because both studied lakes are relatively shallow, even relatively minor fluctuations in water level are clearly recorded both in the geochemical characteristics of the sediments and in the phyto- and zooplankton communities. The multiproxy records indicate several periods of high water stands: 5.4–5.2 ka, 3.5–2.7 ka, 1.4–1.0 ka and 0.5–0 ka, and prominent dry periods 2.7–2.1 ka and 1.7–1.5 ka. Comparison of the reconstructed water levels of the Tatra lakes with records from other European regions suggests that at the boundary of the Middle and the Late Holocene, the hydrological conditions in the Tatras were similar to those in Western and Central Europe. Later, beginning approximately 3500 years ago, records from the Tatras, the northern surroundings of the Pannonian Basin, and the southern part of the Carpathians were unified. In addition to changes in local and regional hydrology, the records from the studied lakes allowed us to reconstruct changes in lake productivity. Relatively high δ13C values, compared to the sedimentary organic matter of other lakes in the region, point to in-lake primary production as a major source of sedimentary organic matter in both lakes. The stable C:N ratio values suggest a constant proportion of organic matter coming from in-lake primary production and transported from the lake catchment. However, the amount of organic carbon and nitrogen and, most of all, differences in the composition of stable C and N isotopes indicate changes in the lake environment. These changes were correlated with some paleotemperature proxies from the region.
Similar content being viewed by others


Introduction
The biota of mountainous regions, including alpine lakes and organisms that inhabit them, are particularly sensitive to changes in environmental conditions. Often living close to their border conditions, in specific ecosystems, such species indicate changes in, for example, climate (Parmesan and Yohe 2003), acidity (Sienkiewicz et al. 2006; Lovett et al. 2009) or geodiversity in the lake catchment (Toivanen et al. 2019) better than those from the lowlands. The Central European mountains are among the most important regions for studying Holocene climate change on this continent. They are located on the border of oceanic and continental climate types, and in the Holocene, they were an effective climate barrier (Hercman et al. 2020).
Despite the high sensitivity of mountain-lake ecosystems as indicators of environmental or climate change, it is sometimes difficult to separate local and regional/global signals recorded by specific proxies. Hence, the selection of the study lake or even the sampling site may have a significant impact on the performed reconstruction. Therefore, it is always better to compare records from at least two sites that differ in certain features (e.g., depth, shore morphology or catchment area and vegetation) to be sure that the interpreted changes have at least regional meaning.
Changes in available moisture significantly affect lakes through the amount of water input, the precipitation/evaporation balance or the transport of mineral matter and nutrients. These processes are often clearly recorded in lake sediments. This is especially true in shallow lakes, where even small changes in recharge can cause relatively large fluctuations in water levels and strongly affect the plants (Heinsalu et al. 2008) and animals (Gąsiorowski and Hercman 2005) that live there. This, in turn, provides an opportunity, through the use of a wide range of geochemical and paleobiological methods, to reconstruct changes in lake hydrology.
In this study, we integrated geochemical (elemental, stable isotope and radioisotopes) and paleobiological (diatoms, Cladocera) data to reconstruct changes, especially related to hydrological conditions, in the ecosystems of two shallow mountain lakes located in the Tatra Mountains (South Poland, Carpathians). These lakes were selected for their stable sediment-deposition conditions, and we hoped to obtain records of environmental change undisturbed by local sedimentary episodes.
Our motivation for the selection of research sites was to obtain complementary records from two lakes located relatively close to each other, which would allow us to separate signals of a local nature from those of at least regional importance. The records span more than the last 5000 years—the entire Late Holocene or Subboreal and Subatlantic periods. This period is considered to be a time of progressive cooling interrupted by several episodes of warming. Although post Holocene Climatic Optimum (9500-5500 cal. yr BP) cooling has been observed globally, short-lasting episodes tended to be regionally significant or asynchronous in different parts of the globe. In this study, we attempt to reconstruct environmental changes in one mountain valley in the Tatra Mountains and compare this reconstruction with other records in the region and worldwide.
Materials and methods
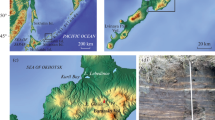
The Kurtkowiec (KUR) and Długi Staw (DLU) lakes are located in the Hala Gąsienicowa Valley in the Tatra Mountains, the highest range of the Carpathian Mountains (Fig. 1). The climate at these sites is characterized by a mean annual temperature of 2.4 °C and a cumulative annual precipitation of 1667 mm (Limanówka 2013). Both are relatively small and shallow oligotrophic lakes located above treeline (Table 1). Their catchment areas are dominated by dwarf pine, mountain meadow and rock (mostly granites with pegmatites and aplites, moraines and screes). The KUR Lake (1686 m a.s.l.) is supplied by the stream draining located above the DLU Lake (1783 m a.s.l.), which also drains water from Zadni Staw Gąsienicowy Lake (1852 m a.s.l.).

Location of the studied sites. The source of the topographic layout: https://pl.mapy.cz/zakladni?x=20.0045519&y=49.2268898&z=15&l=0 (Access date: 12-15-2021)
Field work and coring, lithology and stable isotope analysis
The coring of sediment sequences was performed during the winter season from the ice cover using a Kajak-type gravity corer. The cores were extruded in the field, divided into 0.5- or 1-cm-thick slices and stored in plastic bags in a cold room until subsampling in the laboratory.
Subsamples for elemental and stable isotope analysis were dried at 60 °C over 24 h, ground in an agate mortar and homogenized. The concentrations of total organic carbon (TOC), total nitrogen (TN), total hydrogen (TH) and total sulfur (TS) were measured with a Vario Cube elemental analyzer. Five to ten milligrams of sediment were treated with 10% HCl for carbonate removing, washed twice with distilled water, dried again and transferred to preweighed tin capsules. Then, samples were combusted at a temperature of 1150 °C, and the obtained gases (CO2, N2, H2O and SO2) were separated on a chromatographic column and measured with a thermal conductivity detector. Sulfuric acid was measured with every series of samples as a standard. The measurement uncertainties for TOC, TH, TN and TS were 0.6, 0.21, 0.18 and 0.43%, respectively.
The organic nitrogen and carbon isotopic compositions were analyzed in dry sediment samples using a Thermo Flash 1112HT elemental analyzer connected to a Thermo Delta V Advantage mass spectrometer in continuous flow mode. The samples were washed with ultrapure water to remove pore water, dried at 60 °C for 48 h, treated with weak hydrochloric acid to remove detrital carbonates and dried again. Measurements were calibrated to the USGS 40, USGS 4 and IAEA 600 standards. The results were reported as per mil (‰) deviations versus Vienna Pee Bee Belemnite (δ13C) and atmospheric N2 (δ15N). The analytical errors (1 SD) for the δ13C and δ15N measurements were 0.17 and 0.24‰, respectively.
Sediment dating
Sediment sequences were dated with a combination of 210Pb, 137Cs and radiocarbon methods. Samples for cesium activity analysis were dried, homogenized, transferred to round polyethylene cups (flat cylinder geometry) and measured using low-background gamma spectrometry (Canberra-Packard) with a broad-energy germanium (BE-5030) detector (FWHM = 1.28 keV at 661.7 keV for 137Cs). The activity of 210Pb was determined indirectly by measuring the activity of 210Po (α = 5.31 MeV, T1/2 = 138 days), assuming an isotopic equilibrium between these isotopes (Appleby 2001). A known amount of 208Po (α = 5.11 MeV) and 209Po (α = 4.88 MeV) was added to the weighed sample as an internal yield tracer. Polonium was separated from the sample using strong hydrochloric and nitric acids, and 30 wt% hydrogen peroxide was used to eliminate organic matter. Polonium was deposited onto silver disks (Flynn 1968). The activity of 210Po and 208Po was measured using a DUO alpha spectrometer produced by EG&G ORTEC and corrected for background measurements.
Material for radiocarbon dating was manually picked from sediment samples. For sediment samples with no visible macrofossils, pollen concentrates were prepared. The pollen preparation included the removal of calcium carbonate with 38 wt% hydrochloric acid and humic acids with 10 wt% potassium hydroxide from 2 to 4 cm3 sediment samples. The samples were carbonized at 600 °C in an argon atmosphere, combusted to CO2 at 1000 °C, purified with cupric oxide and silver wire and lastly graphitized. The analysis was performed in Beta Analytic and in the Poznań Radiocarbon Laboratory with a 1.5 SDH-Pelletron Compact Carbon AMS instrument (National Electrostatics Corporation). Conventional radiocarbon dates were calibrated using OxCal ver. 4.4 software (Bronk Ramsey 2009) and the IntCal20 calibration curve (Reimer et al. 2020). Age–depth models were calculated in OxCal software using the P_Sequence function (Bronk Ramsey and Lee 2013).
Diatom analysis
Sediment samples for diatom analysis were prepared according to the standard method (Battarbee 1986). Samples in a volume of 1 cm3 were treated with 10% HCl to remove carbonates and were heated with 30% H2O2 until all organic matter was oxidized. The prepared samples were diluted (to 25–100 ml) with distilled water to the appropriate concentration and thoroughly mixed (homogenized) to achieve an equal representation of the diatom community on the final slides. Permanent slides were mounted in Naphrax® (R.I. = 1.75). In each sample, more than 300 diatom valves were identified using an Olympus BX51 light microscope. Diatom identification was based on Krammer and Lange-Bertalot (1986, 1988, 1991a, b) and Lange-Bertalot and Metzeltin (1996). The nomenclature of diatoms has been upgraded according to AlgaeBase data (Guiry and Guiry 2020). The diatom stratigraphy was divided into diatom zones using Constrained Incremental Sum of Squares Cluster Analysis (CONISS; Grimm 1987) for clustering the diatom assemblages powered by RiojaPlot 1.03.
The reconstruction of pH in the sediments of DLU and KUR lakes was based on the WA method with classical deshriking using the diatom-inferred pH transfer function in the POL_SLOV dataset (Sienkiewicz et al. 2021). It has the best relationships between the observed and estimated pH values (R2 = 0.94) and modern and fossil diatom communities. The snow water equivalent (SWE) was calculated based on the proportion of diatom taxa of the genus Aulacoseira related to turbid/clear water (Pedraza Garzon 2021) and expressed as the logarithm of the relative abundance ratio.
Cladocera analysis
Preparation of samples for Cladocera analysis was based on the standard procedure of Frey (1986). One cubic cm of wet sediment was sampled every 1 cm. The sediment was heated in a 10 wt% water solution of potassium hydroxide, and calcium carbonate was eliminated with 10 wt% chloric acid. The sample was sieved with 33-µm mesh, and the residuum was transferred to 10 ml of distilled water. Microscope slides were prepared with 0.1 ml of the homogenized solution per slide. Cladocera remains were identified and counted with a Zeiss Axioskop 2 optical microscope at 100, 200 and 400 × magnifications. Two–four slides were analyzed from every sample. Identification was based on keys by Flössner (2000) and Szeroczyńska and Sarmaja-Korjonen (2007), and the number of individuals was calculated based on the most numerous portion of the skeleton. The results are presented as a diagram of relative abundance. The trophic and habitat preferences of specific species were adopted from Bjerring et al. (2009).
Statistical analysis
Zonation of diatom and Cladocera diagrams was based on constrained single link clustering (ConsLink analysis) and principal component analysis (PCA). The number of statistically significant zones was determined on the basis of the broken stick model (Jackson 1993). Analysis of similarities (ANOSIM) was used to test the significance of differences between identified zones (Clarke 1993). Computations were performed with Canoco 5.03 (ter Braak and Šmilauer 2012) and PAST 3.10 software (Hammer et al. 2001). Reconstruction of changes in lake-water pH based on diatom-species composition was performed with C2 software (Juggins 2001).
Results
Lithology and stable isotope analysis
The core of sediments from Kurtkowiec Lake was macroscopically homogenous and consisted of dark brown sapropelic mud. The TOC and TN contents were higher in the lower part of the core (up to 18 cm) and equaled ~ 20% and ~ 1.5%, respectively (Fig. 2). Upper in the core, the content of both elements decreased and attained their minima (11% for C and 1% for N) in the surface samples. The C:N ratio was stable along the entire core and varied between 13 and 16. Stable carbon isotopes varied slightly between − 22.3 and − 21.0‰ along the core with two prominent peaks to higher values at depths of ~ 20 cm (up to − 18.9‰) and 8–9 cm (up to − 20.0‰). The curve of δ15N in the Kurtkowiec sediment core started with values above 3.5‰, but shortly after that, it decreased to ~ 3‰ at depths of 41–7 cm. In the uppermost portion of the core, δ15N values dropped to as low as 0.3‰ at a depth of 2 cm.

Total organic carbon (TOC) and total nitrogen (TN) content and C and N stable isotope composition in the sediments of Kurtkowiec (KUR) and Długi Staw (DLU) lakes
The sedimentary core of Długi Staw Lake could be macroscopically divided into two parts. The first portion, up to 25 cm, was characterized by a dark brown color and a relatively high (~ 20%) TOC content (Fig. 2). The color changed to pale brown above 25 cm to the top of the sediment column, and the carbon content decreased to 11–16%. The nitrogen concentration did not change much along the core and was between 1.2 and 2.0%. The molar C:N ratio followed changes in the carbon content and was ~ 13 in the lower part of the core and ~ 9 in the upper part, and only at a depth of 2–4 cm did it increase above 10 again. The stable isotope composition of carbon changed in a pattern different from that of the carbon content. In the lower part of the core, δ13C increased from − 26.15‰ to a maximal value of − 24.75‰ at a depth of 29 cm. From a depth of 29 cm, δ13C started to decrease, and from a depth of 13 cm, it stabilized at ~ − 26.2‰. The exception was the layer at a depth of 2–4 cm, where δ13C rose again to ~ − 25.8‰. The curve of δ15N in the Długi Staw sediment core showed two sections: the first, up to 39 cm, was characterized by relatively high values (mostly above 2‰), and the second was characterized by lower values (0.2–1.7‰) and more dynamic changes (Fig. 2).
Sediment chronology and deposition rate
The age–depth relationship in the KUR sediment sequence is almost linear over the entire analyzed section (Fig. 3). 137Cs was recorded only in the 0.5-cm-thick layer in the uppermost part of the sediment column, while allogenic 210Pb was measured only in the two uppermost samples (0–1 cm depth). The lowest sample taken (depth 47 cm) was dated to 5645 ± 107 cal. yr BP (Table 2) and, together with cesium and lead data, suggests a low deposition rate throughout the core (i.e., 8–9 mm per 100 years).

Chronology of the studied sedimentary sequences. Age-depth models based on calibrated radiocarbon dates were calculated using OxCal ver. 4.4 software. The young ages for the topmost samples were confirmed by measurements of 210Pb and 137Cs
Model calculation for DLU indicated that the topmost 60 cm of the sediment column was deposited during the last 2700 ± 118 years (Fig. 3). 137Cs was present only in the topmost sample, while allogenic 210Pb was present in the two topmost samples. The sedimentation rate changed in the core and was 36 mm per 100 years in the lower portion and slowed down to 20 mm per 100 years during the last ca. 1600 years. These values are comparable with the sedimentation rate calculated for the core from DLU collected in 1993 (Appleby and Piliposian 2006).
Diatom analysis and pH changes
Kurtkowiec Lake
A total of 130 diatom taxa belonging to 26 genera were identified from the sediments of Kurtkowiec Lake. The diatom stratigraphy was divided into five zones (Fig. 4). Dominant diatoms were benthic/periphytic taxa.

Diatom relative abundance, diatom records’ zonation, the first Principal Component axis and diatom-inferred pH reconstructions from Kurtkowiec Lake (KUR) and Długi Staw Lake (DLU) in the Tatra Mountains
Diatom-assemblage zone (DAZ) 1 (47-31 cm; 5645-3758 cal. yr BP)
This zone was dominated by Aulacoseira italica (Ehr.) Simonsen, Pseudostaurosira brevistriata (Grun.) D. M. Williams & Round, and Sellaphora seminulum (Grun.) Mann with the highest abundance in the core. In the upper part of the zone, there was a slight increase in Aulacoseira lirata (Ehr.) Ross and Pseudostaurosira elliptica (Schum.) Edlund, Morales & Spauld. The highest abundance of planktonic diatoms occurs in this zone.
DAZ 2 (31–17.5 cm; 3758–2251 cal. yr BP)
This part of the core was dominated by Staurosirella pinnata (Ehrenberg) D.M.Williams & Round, while A. italica almost completely disappeared in the upper part of the zone. The decrease of S. seminulum in relation to the previous zone was also noted. A high abundance of benthic diatoms (i.e., > 90%) occurred at the bottom and at the top of this zone.
DAZ 3 (17.5–7.5 cm; 2251–995 cal. yr BP)
This zone was characterized by a significant decrease in S. pinnata and an increase in planktonic A. lirata and periphytic Psammothidium sacculum (Carter) Bukhtiyarova. At the top of the zone, the highest abundance (above 20%) of Pinnularia interrupta W. Smith was observed. Small forms of other Psammothidium spp., such as P. levanderi (Hust.) Bukhtiyarova & Round and P. curtissimum (Carter) Aboal, also increased in abundance.
DAZ 4 (7.5-2.5 cm; 995-290 cal. yr BP)
This zone was dominated by small forms of Fragilaria sensu lato, such as Pseudostaurosira elliptica (Schumann) Williams & Round and Staurosirella pinnata. At that time, almost 100% of the diatom community consisted of benthic taxa. Some planktonic diatoms occurred as single valves (e.g., A. italica) or at a very low percentage abundance.
DAZ 5 (2.5-0 cm; 290 cal. yr BP—present)
Substantial changes in the diatom assemblage were observed in the youngest sediments. In comparison with the previous zone, a large decrease in Fragilaria s.l. and a slight increase in small forms of Achnanthes s.l. (e.g., Psammothidium curtissimum, P. marginulatum (Grun.) Bukhtiyarova & Round and P. subatomoides (Hust.) Bukhtiyarova & Round) were noted.
In the sediments of KUR Lake, the diatom PCA 1 axis shows higher values between ~ 5500 and ~ 2700 cal. yr BP (DAZ 1—lower part of DAZ 2) and lower values from ~ 2700 cal. yr BP to the present (higher part of DAZ 2–DAZ 5). Higher values reflect the increase in A. italica, P. brevistriata, S. pinnata and S. seminulum, while lower values reflect opposite trends in species abundance, especially of P. elliptica in DAZ 4.
Długi Staw Gąsienicowy Lake
A total of 105 diatom taxa belonging to 22 genera were identified from the sediments of DLU Lake. Three DAZs were determined (Fig. 4):
DAZ 1 (63–51 cm; 2730–2327 cal. yr BP)
This zone was dominated by Aulacoseira spp., such as A. distans (Ehr.) Simonsen, A. alpigena (Grun.) Krammer and A. lirata and periphytic Psammothidium curtissimum, P. subatomoides.
DAZ 2 (51–20 cm; 2327–1487 cal. yr BP)
In this part of the core, the highest abundance (above 40%) of A. distans was noted. The abundance of P. curtissimum, P. subatomoides and P. levanderi remained almost constant. A slight decrease in the abundance of A. alpigena and A. lirata occurred in this zone.
DAZ 3 (20–0 cm; 1487 cal. yr BP—present)
The youngest sediments were also dominated by A. distans and P. curtissimum. An increase in the abundance of small forms of Sellaphora seminulum was observed. In the upper part of the zone, the abundance of P. levanderi decreased and Pseudostaurosira elliptica disappeared.
PCA Axis 1 for DLU exhibits the lowest values from ~ 2730 to ~ 792 cal. yr BP (DAZs 1 and 2) and higher values from ~ 792 cal. yr BP to present (DAZ 3). The lower values in zones DAZ 1 and DAZ 2 are driven primarily by an increase in Aulacoseira spp. (A. alpigena, A. distans, A. lirata). Higher PCA 1 values reflect a decrease in the abundance of Aulacoseira spp. and Pseudostaurosira elliptica and an increase in Sellaphora seminulum.
Reconstruction of pH (DI-pH) in the sediments of KUR and DLU lakes
The reconstructed pH values in sediments of KUR and DLU varied from 6.60 to 6.94 and from 5.82 to 6.39, respectively. The curves in both lakes do not point to an unequivocal trend. During the last few 1000 years, the water pH in KUR Lake was almost neutral, while in DLU Lake, it was moderately acidic. The pH values changed by 0.34 and 0.57 units, respectively.
Cladocera analysis
Kurtkowiec Lake
The most numerous remains belonged to chydorids, mostly Acroperus harpae, Alona affinis and Eurycercus lamellatus (Fig. 5). Despite the generally higher diversity of Cladocera in KUR than in DLU, vertical changes in species composition were relatively small, and the entire core was divided into only two zones. The lower Cladocera assemblage zone (CAZ 1; 47–14 cm; 5645–1830 cal. yr BP) was characterized by higher diversity and the presence of Alona quadrangularis, which was absent in the topmost portion of the sediment core. In the lower part of the zone, few remains of Daphnia were recorded. CAZ 2 (14–0 cm; 1830 cal. yr BP—present) was dominated by remains of Acroperus harpae and in the lower portion by Eurycercus lamellatus, whose numbers, however, decreased in the topmost sediment. The PC1 values generally follow the ratio of Alona affinis to other chydorids. This axis has a negative value until 18 cm, a positive value above 13 cm, and with the transition zone (18–13 cm), where relatively abrupt changes in the proportion of chydorid species were observed.

Cladocera records from Kurtkowiec Lake (KUR) and Długi Staw Lake (DLU) in the Tatra Mountains. The PC axis1 for DLU was nonsignificant and thus not presented
Długi Staw Gąsienicowy Lake
The Cladocera assemblage from DLU differed strongly from that of KUR (Fig. 5). Remains of five Cladocera species were found in DLU sediments. Chydorus sphaericus s.l., which was only an accessory species in KUR, dominated the entire core in DLU. The second taxon present in all samples was Daphnia pulex-group. Despite the low species diversity along the core, it was divided into three cladoceran zones. Only C. sphaericus s.l. and Daphnia pulex were found in the upper part of CAZ 1 (63–42 cm; 2730–2076 cal. yr BP). The middle part of the core was separated as the second zone (CAZ 2; 42–10 cm; 2076–714 cal. yr BP) based on the presence of benthic taxa, namely, Acroperus harpae, Alona affinis and Alona guttata. These benthic chydorids were absent in the topmost portion of the core (CAZ 3; 10–0 cm; 714 cal. yr BP—present), with the exception of Alona guttata, which was numerous (over 30% relative abundance) in the surface sediment sample. The low taxonomic diversity and only minor changes in relative abundance caused all PC axes to be statistically nonsignificant and thus were not analyzed.
Discussion
Environmental interpretation of the proxy records
The studied lakes have many similarities. Both are located at a similar altitude under the same climatic conditions. Both are oligotrophic lakes and have a similar surface area and catchment area. However, some of their characteristics differ. The most important are depth, the dominant vegetation in the catchment (meadow/rock vs. dwarf pine zone) and the shore morphology determined by the lake origin. These features were sufficient to make the geochemical and paleobiological records in the two lakes very different.
DLU is a postglacial lake occupying a postglacial cirque, and on its east side it directly borders the steep rocky slope of Mt. Kościelec (Fig. 1). This results in a higher input of detrital material and nutrients to the lake, causing a sedimentation rate twice as high as that in the KUR. The sedimentation rate in KUR was very low during the last century (only one centimeter of sediment column was deposited), much lower even compared to Greenland lakes (Appleby 2008), and has been stable for at least the last five millennia.
The relatively high δ13C values compared to the sedimentary organic matter (OM) of other lakes in the region (Gąsiorowski and Sienkiewicz 2013), noted especially in KUR, are mainly a result of the low water depth in the studied lakes (Piscia et al. 2018) but also point to in-lake primary production as a major source of the sedimentary OM in both lakes (Torres et al. 2012). Since there are significant differences in δ13C values between specific phytoplankton taxa, such values could also be interpreted as an indicator of the major group of producers in the lake, with high values characteristic of cyanobacteria (Taipale et al. 2016). Stable C:N ratios (~ 9–12 in DLU and ~ 15 in KUR) suggest a stable proportion of organic matter coming from primary production in lakes and transported from the lakes’ catchments. However, the amount of organic carbon and nitrogen, and most of all, changes in the stable isotope composition of C and N indicate changes in the lake environment. In KUR, changes in δ13C followed changes in C content, which additionally confirms the stable source of OM deposited into sediments. In DLU, this relationship was not as obvious. δ13C values increased while the C content was stable in the lower portion of the core (depth > 25 cm). In younger sediments (depth < 20 cm), δ13C started to follow the C content. The switch coincided with the change in the diatom assemblage (DAZ2/DAZ3) related to a decrease in the abundance of Aulacoseira spp. and Pseudostaurosira elliptica and an increase in Sellaphora seminulum.
Diatom records from Arctic and mountain lakes are often related to climate change (Korhola et al. 2000; Bigler and Hall 2003). Changes in air temperature directly affect the water temperature in the lake but also determine the thickness and duration of the ice cover. Diatom PC1 follows the general cooling trend in the Late Holocene (Fig. 6) but also stresses some climatic events, with the most visible change at the Subboreal/Subatlantic chronozones transition (2.8 kyr event) and minimum during the Little Ice Age (LIA). Diatoms react not only to changes in temperature but also to water transparency and circulation—some taxa (e.g., Aulacoseira lirata) prefer clear water, while others (e.g., Aulacoseira alpigena and A. italica) need mixing to stay in the water column (Pedraza Garzon 2021).

Correlation of different proxies from sedimentary records of KUR and DLU lakes in the extra-regional context. A humid climate phases reconstructed for Central (Haliuc et al. 2017, Demeny et al. 2019) and Western (Magny 2004) Europe; B compilation of hydrological, sedimentary and biological indicators from the KUR and DLU lake records, from top to bottom: blue squares indicate higher proportion of Daphnia remains, blue curves are for snow water equivalent (SWE), orange curves summarize changes in organic matter elemental and isotopic composition, green and pale blue lines are PC1 axis score for diatom and Cladocera assemblages, respectively; C stable oxygen isotope record from JMr-Hol stalagmite collected in the Demänová Cave System (Hercman et al. 2020); D local pollen zones based on the records from the Kurtkowiec and Zielony Staw lakes (Obidowicz 1996); E pine (blue) and hazelnut (red) pollen fluxes for Gościąż Lake record (Ralska-Jasiewiczowa et al. 1998); F relative changes in total solar irradiance (Steinhilber et al. 2009); G climate periods and events during Late Holocene: IA—Iron Age, RWP—Roman Warm Period, MP—Migration Period, LALIA—Late Antique Little Ice Age, MWP—Medieval Warm Period, LIA—Little Ice Age; H—radiocarbon age in years before present (AD 1950)
The general trend of changes in Cladocera assemblages in KUR is consistent with that of diatoms and may be linked with the Late Holocene cooling (Fig. 6). Moreover, Daphnia are a good indicator of water-level changes due to the need for a minimal depth for vertical migration, which in shallow lakes is possible only during high water stands. The increase in Daphnia in DLU coincided with minor changes in the Cladocera assemblage in KUR (i.e., decrease in PC1), suggesting that in both lakes, Cladocera are good indicators of changes in regional hydrology.
Changes in lakes related to climate
Before the Late Holocene cooling
The Middle/Late Holocene transition is recorded only in KUR lake sediments. The lowest portion of the core has been dated to 5600–5000 cal. yr BP, representing the youngest part of the Atlantic period. At that time, a coniferous forest dominated by pine and spruce grew in the vicinity of the lake, and the tree line was positioned higher than today (Obidowicz 1996). The diatom flora of the lake was dominated by Pseudostaurosira brevistriata, Staurosirella pinnata, Sellaphora seminulum and Aulacoseira italica. The latter species require turbulent water to remain in suspension in the photic zone (Rühland et al. 2015), and in alpine lakes, this is related to the higher snowmelt discharge (Pedraza Garzon 2021). The high SWE and Daphnia remains indicate a relatively high input of meltwater during the spring seasons and a high water level (Fig. 6). Additionally, the C content and the C:N ratio indicate relatively high lake productivity. These data suggest a humid climate at approximately 5.2 kyr, which is coherent with data from Western Europe (Roland et al. 2015) and the southern part of the Carpathians (Constantin et al. 2007; Drăguşin et al. 2014), where the climate was relatively humid and warm at that time.
Subboreal and Subboreal/Subatlantic transition (2.8 kyr climate event)
With the onset of the Subboreal period, the Late Holocene cooling started, at least in the mid- and high latitudes (Luoto and Nevalainen 2015; Bader et al. 2020). The lower SWE after ca. 5000 cal. yr BP and the lack of Daphnia remains in KUR Lake sediments suggest a decreasing water level equivalent to the dry phase recorded in lakes and peatland in Central and North Poland (Ralska-Jasiewiczowa and Starkel 1988; Gąsiorowski and Kupryjanowicz 2009), but in the southern part of the Carpathians, the wet phase lasted as long as 4200 yr BP (Haliuc et al. 2017). Spruce became the dominant tree in the forest around the lake (Obidowicz 1996). The 4200 cal. yr BP climate event, similar to the Polish lowlands (Pleskot et al. 2020), did not significantly affect the environment of KUR Lake or its catchment area, but the SWE curve indicated low water input to the lake. Another reason for the lacking record of this event in the KUR sediments may be the low temporal resolution of this core. The 2800 cal. yr BP event was characterized in Western Europe by rather cool and wet conditions (van Geel et al. 1999, 2014; Martin-Puertas et al. 2012). Additionally, in our records, higher humidity is confirmed at a relatively high water level, expressed by a high SWE value in KUR and Daphnia increase in DLU sediments. Although the 2800 cal. yr BP episode is not clearly recorded in the KUR sediments, probably like the 4200 cal. yr BP episode due to low temporal resolution, it is synchronous with major changes in the lake ecosystem. Shortly after that (at 2700 cal. yr BP), the water level clearly decreased, suggesting a drier climate. The water level, at least in the KUR Lake, has no longer returned to the level before the 2800 cal. yr BP episode.
Biological turnover at the beginning of the Roman Warm Period and during the Medieval Warm Period
The general cooling trend of the global climate during the Late Holocene was interrupted by several relatively warm periods of global or regional importance. One of the better documented such periods in Europe was the Roman Warm Period (RWP) lasting from ~ 2200 to 1550 cal. yr BP (Margaritelli et al. 2020). This period in our records was manifested by the turnover in cladoceran PC1 in KUR Lake caused mainly by the replacement of Alona quadrangularis by A. affinis, change in diatom assemblage and drop in DI-pH in DLU and significant decrease in the TOC content in both lakes. Additionally, δ13C values changed during this time and increased in KUR while they decreased in DLU. All these records suggest periphyton development (Tremel et al. 2000) and low water levels, probably caused by higher evapotranspiration and hence lower plankton productivity in KUR. In DLU, the lake level changed, with higher stands during the second part of the RWP. In the close vicinity of this lake, there were probably also peatbogs, which caused a decrease in pH. The RWP in the Mediterranean region ended with cooling related to the so-called Late Antique Little Ice Age (Büntgen et al. 2016). In our lakes, this event has been recorded only slightly with higher TOC content and higher δ13C values in KUR, probably indicating a higher water level.
The Medieval Warm Period was again a time of relatively low water level, which is reflected in the lower SWE values in both lakes and the lower proportion of Daphnia in DLU (Fig. 6). Additionally, the diatom flora, dominated by the small benthic Pseudostaurosira elliptica (Fig. 4.), the species common in high alpine oligotrophic lakes and streams (Niyatbekov and Barinova 2018), indicated that the water level was low and, despite milder temperature, the lake was frozen to the bottom.
Little Ice Age
LIA records from KUR and DLU lakes, similar to other sites in the Tatra Mountains (Bitušik et al. 2009; Gąsiorowski and Sienkiewicz 2010), suggest the duality of this period. The first part, dated to ~ 1300–1500 AD, was characterized by low SWE values in the KUR Lake and low C concentrations and δ13C values in the DLU Lake and had relatively stable conditions with a cool and dry climate. The second part, which started ~ 1500 AD, was wet and cold by contrast (Gąsiorowski and Sienkiewicz 2010; Hercman et al. 2010), with intense snowfalls and short cool summers, which was documented by higher SWE values (but never to the level calculated for the Subboreal period) and a higher proportion of Daphnia in the cladoceran assemblages in DLU. The higher C content in DLU was probably not related to higher productivity, but together with a clear increase in the C:N ratio (Fig. 2), it documented a higher input of the allogenic OM to the lake, caused by higher slope activity around the lake. Similar events of slope activity and intensification of mineral deposition in lakes were also reported from other valleys in the region (Kotarba 2006).
Paleohydrological changes in the Tatra Mountains in the extraregional context
Several phases of higher humidity expressed by a higher water level in the studied lakes can be distinguished over the last 5500 years. Higher water stands were dated to the following periods: 5400–5200 cal yr BP, 3500–2700 cal yr BP, 2000–1700 cal yr BP, 1400–1000 cal yr BP and 500–0 cal yr BP. These series of prominent and some short-lasting changes in water level reconstructed in the KUR and DLU lakes had their equivalents in other paleoenvironmental archives of the region and Europe. The oldest recorded high water-stand period was similar to the phase of water-level increase in lakes located in Western Europe (Magny 2004) but was not noticed in some records from central Europe and the Carpatho-Balkan region. This suggests a stronger connection between the western and northern European climates at this time. In fact, the northern Carpathian range became influenced by the oceanic air mass from the North Atlantic as early as ~ 9000 cal. yr BP (Hercman et al. 2020), while in the southern Carpathians, the dominance of the ‘Atlantic’ circulation started ca. 4700 cal. yr BP (Perșoiu et al. 2017). Thus, both speleothem records from the Lower Tatra Mountains and lake-sediment records from this study stress the important role of the Carpathians in shaping the regional climate at that time. After climate unification in the entire Carpathians region ca. 4700 cal. yr BP, the hydrological phases in Western Europe, the Tatra Mountains and the Pannonian Basin, and to some extent, in the southern Carpathians, became similar. Especially clear record of wet phase at 3500–2700 cal. yr BP was related to humid and relatively warm period. In contrast, the wet phase dated to the Migration Period (1400–1000 cal. yr BP) was related to cooling in the region, as recorded, for example, in speleothems in the Demianova Cave System (Hercman et al. 2020) and caused by a prominent minimum in solar activity (Fig. 6). Some wet phases, however, have not been recorded throughout Central Europe. Wet phases at ~ 2500 cal yr BP were noted in Western Europe and the southern part of the Carpathians (Haliuc et al. 2017) but not in the Pannonian Basin or the Tatra Mountains. The reason may be that this phase was related to the negative phase of the North Atlantic Oscillation (NAO−) and weaker penetration of the humid Atlantic air masses in the northern part of the continent, including the northern Carpathians (Olsen et al. 2012).
Conclusions
The multiproxy study of two shallow mountain lakes suggests that hydrological events, namely, snowmelt input and lake-water-level fluctuations, had a stronger influence on the biota of the study lakes than changes in the temperature during the Late Holocene. Organisms living in such an ecosystem might experience not only episodes of water-level lowering but also a lack of water during the winter season due to freezing to the bottom of shallow water, despite higher mean annual temperatures. Some of the climatic and ecological episodes were recorded only in the sediments of one of the lakes studied (e.g., the rise in water level in the DLU ~ 2000 yr BP interpreted on the basis of the increased proportion of Daphnia remains), and in that case should be treated as a record of local environmental conditions and their significance on a regional scale should be interpreted with caution. Short-lived climatic episodes were not recorded in the sediments of the studied lakes due to insufficient temporal resolution although the effects of some of them, such as the 2800 cal. yr BP episode, caused long-term changes in these ecosystems. This research also confirmed strong role of Carpathians as climatic barrier during earlier phases of Late Holocene and unification of climate (or at least precipitation conditions) in almost the entire region after ~ 4700 yr BP.
References
Appleby PG (2001) Chronostratigraphic techniques in recent sediments. In: Last WM, Smol JP (eds) Tracking environmental changes using lake sediments, vol 1. Basin Analysis, Coring, and Chronological Techniques. Kluwer Academic Publishers, Dordrecht, pp 171–203
Appleby PG (2008) Three decades of dating recent sediments by fallout radionuclides: a review. Holocene 18:83–93
Appleby PG, Piliposian GT (2006) Radiometric dating of sediment records from mountain lakes in the Tatra Mountains. Biolo Bratisl 61(Suppl. 18):S51–S64
Bader J, Jungclaus J, Krivova N, Lorenz S , Maycock A, Raddatz T, Schmidt H, Toohey M, Wu C-J, Claussen M (2020) Global temperature modes shed light on the Holocene temperature conundrum. Nat Commun 11:4726
Battarbee RW (1986) Diatom analysis. In: Berglund BE (ed) Handbook of Holocene palaeoecology and palaeohydrology. John Willey and Sons, Hoboken, pp 527–570
Bigler C, Hall RI (2003) Diatoms as quantitative indicators of July temperature: a validation attempt at century—scale with meteorological data from northern Sweden. Palaeogeogr Palaeoclimatol Palaeoecol 198:147–160
Bitušík P, Kubovčík V, Štefková E, Appleby PB, Svitok M (2009) Subfossil diatoms and chironomids along an altitudinal gradient in the High Tatra Mountain lakes: a multi-proxy record of past environmental trends. Hydrobiologia 631:65–85
Bjerring R, Becares E, Declerck S, Gross EM, Hansson LA, Kairesalo T, Nykanen M, Halkiewicz A, Kornijow R, Conde-Porcuna JM, Seferlis M, Nöges T, Moss B, Amsinck SL, Odgaard BV, Jeppesen E (2009) Subfossil Cladocera in relation to contemporary environmental variables in 54 Pan—European lakes. Freshw Biol 54:2401–2417
Bronk Ramsey C (2009) Bayesian analysis of radiocarbon dates. Radiocarbon 51:337–360
Bronk Ramsey C, Lee S (2013) Recent and planned developments of the program OxCal. Radiocarbon 55(2–3):720–730
Büntgen U, Myglan VS, Ljungqvist FC, McCormick M, Di Cosmo N, Sigl M, Jungclaus J, Wagner S, Krusic PJ, Esper J, Kaplan JO, de Vaan MAC, Luterbacher J, Wacker L, Tegel W, Kirdyanov AV (2016) Cooling and societal change during the late antique little ice age from 536 to around 660 AD. Nat Geosci 9:231–236
Clarke KR (1993) Non-parametric multivariate analysis of changes in community structure. Aust J Ecol 18:117–143
Constantin S, Bojar A-V, Lauritzen S-E, Lundberg J (2007) Holocene and late Pleistocene climate in the sub-mediterranean continental environment: a speleothem record from Poleva Cave Southern Carpathians, Romania. Palaeogeogr Palaeoclimatol Palaeoecol 243:322–338
Demény A, Kern Z, Németh A, Frisia S, Hatvani IG, Czuppon G, Leél-Őssy S, Molnár M, Óvári M, Surányi G, Gilli A, Wu C-C, Shen C-C (2019) North Atlantic influences on climate conditions in East-Central Europe in the late-Holocene reflected by flowstone compositions. Quat Int 512:99–112
Drăguşin V, Staubwasser M, Hoffmann DL, Ersek V, Onac BP, Veres D (2014) Constraining Holocene hydrological changes in the Carpathian—Balkan region using speleothem δ18O and pollen-based temperature reconstructions. Clim Past 10:1363–1380
Flössner D (2000) Die Haplopoda und Cladocera (ohne Bosminidae) Mitteleuropas. Backhuys Publishers, Leiden, pp 1–428
Flynn WW (1968) The determination of low-levels of polonium-210 in environmental materials. Anal Chim Acta 43:221–227
Frey DG (1986) Cladocera analysis. In: Berglund BE (ed) Handbook of Holocene palaeoecology and palaeohydrology. John Wiley & Sons, Chichester, pp 667–692
Gąsiorowski M, Hercman H (2005) Recent sedimentation and eutrophication of Kruklin Lake after artificial drop in water-level in the middle of 19th century. Studia Quaternaria 22:17–25
Gąsiorowski M, Kupryjanowicz M (2009) Lake–peat bog transformation recorded in the sediments of the Stare Biele mire (Northeastern Poland). Hydrobiologia 631:143–154
Gąsiorowski M, Sienkiewicz E (2010) The little ice age recorded in sediments of a small dystrophic mountain lake in southern Poland. J Paleolimnol 43:475–487
Gąsiorowski M, Sienkiewicz E (2013) The sources of Carbon and Nitrogen in mountain lakes and the role of human activity in their modification determined by tracking stable isotope composition. Water Air Soil Pollut 224:1498
Grimm E (1987) CONISS: a Fortran 77 program for stratigraphically constrained cluster analysis by the method of incremental sum of squares. Comp Geosci 13:13–25
Guiry MD, Guiry GM (2020) AlgaeBase. World-wide electronic publication, National University of Ireland, Galway. https://www.algaebase.org; Accessed 26 Mar 2020
Haliuc A, Veres D, Brauer A, Hubay K, Hutchinson SM, Begy R, Braun M (2017) Palaeohydrological changes during the mid and late Holocene in the Carpathian area, central-eastern Europe. Glob Planet Change 152:99–114
Hammer Ø, Harper DAT, Ryan PD (2001) PAST: paleontological statistics software package for education and data analysis. Palaeontol Electron 4:9
Heinsalu A, Luup H, Alliksaar T, Nõges P, Nõges T (2008) Water level changes in a large shallow lake as reflected by the plankton:periphyton-ratio of sedimentary diatoms. Hydrobiologia 599:23–30
Hercman H, Gąsiorowski M, Gradziański M, Kicińska D (2010) The first dating of cave ice from the Tatra Mountains, Poland and its implication to palaeoclimate reconstructions. Geochronometria 36:31–38
Hercman H, Gąsiorowski M, Pawlak J, Błaszczyk M, Gradzińsk M, Matoušková Š, Zawidzki P, Bella P (2020) Atmospheric circulation and the differentiation of precipitation sources during the Holocene inferred from five stalagmite records from Demänová Cave System (Central Europe). Holocene 30:834–846
Jackson DA (1993) Stopping rules in principal components analysis: a comparison of heuristical and statistical approaches. Ecology 74:2204–2214
Juggins S (2001) C2—software for ecological and palaeoecological data analysis and visualization user guide version 1.5., p 73
Korhola A, Weckström J, Holmström L, Erästö P (2000) A quantitative Holocene climate record from diatoms in northern Fennoscandia. Quat Res 54:284–294
Kotarba A (2006) The little ice age in the high Tatra mountains. Studia Quaternaria 23:47–53
Krammer K, Lange-Bertalot H (1986) Süßwasserflora von Mitteleuropa. Bacillariophyceae. I. Teil: Naviculaceae. Gustav Fischer Verlag, Stuttgart
Krammer K, Lange-Bertalot H (1988) Süßwasserflora von Mitteleuropa. Bacillariophyceae. 2. Teil: Bacillariaceae, Epithemiaceae, Surirellaceae. Gustav Fischer Verlag, Stuttgart
Krammer K, Lange-Bertalot H (1991) Süßwasserflora von Mitteleuropa. Bacillariophyceae. 3. Teil: Centrales, Fragilariaceae, Eunotiaceae. Gustav Fischer Verlag, Stuttgart
Krammer K, Lange-Bertalot H (1991) Süßwasserflora von Mitteleuropa. Bacillariophyceae. 4. Teil: Achnanthaceae, Kritische Ergänzungen zu Navicula (Lineolatae) und Gomphonema Gesamtliteraturverzeichnis Teil. Gustav Fischer Verlag, Stuttgart, pp 1–4
Lange-Bertalot H, Metzeltin D (1996) Ecology–diversity–taxonomy. Indicators of oligotrophy–800 taxa representative of three ecologically distinct lake types. In: Lange-Bertalot H (ed) Iconographia Diatomologica 2. Koeltz Scientific Books, Koenigstein
Limanówka D (red.) (2013) 100 lat pomiarów meteorologicznych na Hali Gąsienicowej, IMGW-PIB, Kraków, p103
Lovett GM, Tear TH, Evers DC, Findlay SEG, Cosby BJ, Dunscomb JK, Driscoll CT, Weathers KC (2009) Effects of air pollution on ecosystems and biological diversity in the Eastern United States. Ann N Y Acad Sci 1162:99–135
Luoto TP, Nevalainen LE (2015) Late Holocene precipitation and temperature changes in Northern Europe linked with North Atlantic forcing. Climate Res 66:37–48
Magny M (2004) Holocene climate variability as reflectedby mid-European lake-level fluctuations and its probable impact on prehistoric human settlements. Quat Int 113:65–79
Margaritelli G, Cacho I, Català A, Barra M, Bellucci LG, Lubritto C, Rettori R, Lirer R (2020) Persistent warm Mediterranean surface waters during the Roman period. Sci Rep 10:10431
Martin-Puertas C, Matthes K, Brauer A, Muscheler R, Hansen F, Petrick C, Aldahan A, Possnert G, van Geel B (2012) Regional atmospheric circulation shiftFs induced by a grand solar minimum. Nat Geosci 5:397–401
Niyatbekov T, Barinova SS (2018) Bioindication of aquatic habitats with diatom algae in the Pamir Mountains. Tajikistan MOJ Eco Environ Sci 3:117–120
Obidowicz A (1996) A Late Glacial — Holocene history of the formation of vegetation belts in the Tatra Mts. Acta Palaeobot 36:159–206
Olsen J, Anderson NJ, Knudsen MF (2012) Variability of the North Atlantic Oscillation over the past 5200 years. Nat Geosci 5:1–14
Parmesan C, Yohe G (2003) A globally coherent fingerprint of climate change impacts across natural systems. Nature 421:37–42
Pedraza Garzon EL (2021) Deciphering the Ecology of Aulacoseira Taxa in Alpine Lakes: Implications for Paleoclimate Reconstructions. Electronic Theses and Dissertations. p 3394
Perșoiu A, Onac BP, Wynn JG, Blaauw M, Ionita M, Hansson M (2017) Holocene winter climate variability in Central and Eastern Europe. Sci Rep 7:1196
Piscia R, Boggio E, Bettinetti R, Mazzoni M, Manca M (2018) Carbon and nitrogen isotopic signatures of zooplankton taxa in five small subalpine lakes along a trophic gradient. Water 10:94
Pleskot K, Apolinarska K, Kołaczek P, Suchora M, Fojutowski M, Joniak T, Kotrys B, Kramkowski M, Słowiński M, Woźniak M, Lamentowicz M (2020) Searching for the 4.2 ka climate event at Lake Spore, Poland. Catena 191:104565
Ralska-Jasiewiczowa M, Starkel L (1988) Record of the hydrological changes during the Holocene in the lake, mire and fluvial deposits of Poland. Folia Quaternaria 57:91–127
Ralska-Jasiewiczowa M, Goslar T, Madeyska T, Starkel L (1998) Lake Gościąż, central Poland. A Monographic Study. Part 1. W. Szafer Institute of Botany, Polish Academy of Sciences, Kraków
Reimer P, Austin W, Bard E, Bayliss A, Blackwell P, Bronk Ramsey C, Butzin M, Cheng H, Edwards R, Friedrich M, Grootes P, Guilderson T, Hajdas I, Heaton T, Hogg A, Hughen K, Kromer B, Manning S, Muscheler R, Palmer J, Pearson C, van der Plicht J, Reimer R, Richards D, Scott E, Southon J, Turney C, Wacker L, Adolphi F, Büntgen U, Capano M, Fahrni S, Fogtmann-Schulz A, Friedrich R, Köhler P, Kudsk S, Miyake F, Olsen J, Reinig F, Sakamoto M, Sookdeo A, Talamo S (2020) The IntCal20 Northern Hemisphere radiocarbon age calibration curve (0–55 cal kBP). Radiocarbon 62:725–757
Roland TP, Daley TJ, Caseldine CJ, Charman DJ, Turney CSM, Amesbury MJ, Thompson GJ, Woodley EJ (2015) The 5.2 ka climate event: evidence from stable isotope and multi-proxy palaeoecological peatland records in Ireland. Quat Sci Rev 124:209–223
Rühland KM, Paterson AM, Smol JP (2015) Lake diatom responses to warming: reviewing the evidence. J Paleolimnol 54:1–35
Sienkiewicz E, Gąsiorowski M, Hercman H (2006) Is acid rain impacting Sudetic lakes? Sci Total Environ 369:139–149
Sienkiewicz E, Gąsiorowski M, Hamerlík L, Bitušík P, Stańczak J (2021) A new diatom training set for the reconstruction of past water pH in the Tatra Mountain lakes. J Paleolimnol 65:445–459
Steinhilber F, Beer J, Fröhlich C (2009) Total solar irradiance during the Holocene. Geophys Res Lett 36:L19704
Szeroczyńska K, Sarmaja-Korjonen K (2007) Atlas of subfossil Cladocera from Central and Northern Europe. Friends of Lower Vistula Society, Świecie, pp 1–84
Taipale SJ, Vuorio K, Brett MT, Peltomaa E, Hiltunen M, Kankaala P (2016) Lake zooplankton δ13C values are strongly correlated with the δ13C values of distinct phytoplankton taxa. Ecosphere 7(8):e01392
ter Braak C, Šmilauer P (2012) Canoco reference manual and user’s guide: software of ordination (version 5.0). Microcomputer Power, Ithaca, NY
Toivanen M, Hjort J, Heino J, Tukiainen H, Aroviita J, Alahuhta J (2019) Is catchment geodiversity a useful surrogate of aquatic plant species richness? J Biogeogr 46:1711–1722
Torres IC, Inglett PW, Brenner M, Kenney WF, Reddy KR (2012) Stable isotope (δ13C and δ15N) values of sediment organic matter in subtropical lakes of different trophic status. J Paleolimnol 47:693–706
Tremel B, Frey SEL, Yan ND, Somers KM, Pawson TW (2000) Habitat specificity of littoral chydoridae (Crustacea, Branchiopoda, Anomopoda) in Plastic Lake, Ontario, Canada. Hydrobiologia 432:195–205
van Geel B, Raspopov OM, Renssen H, van der Plicht J, Dergachev VA, Meijer HAJ (1999) The role of solar forcing upon climate change. Quat Sci Rev 18:331–338
van Geel B, Heijnis H, Charman DJ, Thompson G, Engels S (2014) Bog burst in the eastern Netherlands triggered by the 2.8 kyr BP climate event. Holocene 24:1465–1477
Acknowledgements
We would like to thank Agata Jaskółka and Wojtek Sienkiewicz for their assistance during the field work. To the employees of the Tatra National Park, we are grateful for their assistance during the planning of the survey and the collection of lake sediments. This work was funded by National Science Centre of Poland (Grant No. 2018/31/B/ST10/00664).
Author information
Authors and Affiliations
Contributions
MG, conceptualization, methodology, data curation, formal analysis, writing—original draft, supervision; ES, conceptualization, methodology, data curation, formal analysis; UK, writing—original draft and editing; KK, writing—original draft and editing, art work.
Corresponding author
Ethics declarations
Conflict of interest
The authors declare no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Gąsiorowski, M., Sienkiewicz, E., Kowalewska, U. et al. Paleohydrological changes during the Late Holocene recorded in the sediments of two mountain lakes in the Tatra Mountains (Central Europe). J Paleolimnol 70, 293–310 (2023). https://doi.org/10.1007/s10933-023-00294-9
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10933-023-00294-9