
Abstract
Generally, nonreproductive sex is thought to act as “social grease,” facilitating peaceful coexistence between subjects that lack close genetic ties. However, specifc nonreproductive sexual behaviors may fulfill different functions. With this study, we aimed to test whether nonreproductive mounts in Barbary macaques are used to 1) assert dominance, 2) reinforce social relationships, and/or 3) solve conflicts. We analyzed nonreproductive mounts (N = 236) and postmount behavior in both aggressive and nonaggressive contexts, in 118 individuals belonging to two semi-free-ranging groups at La Montagne des Singes (France). As predicted by the dominance assertion hypothesis, the probability to be the mounter increased with rank difference, especially in aggressive contexts (increasing from 0.066 to 0.797 in nonaggressive contexts, and from 0.011 to 0.969 in aggressive contexts, when the rank difference was minimal vs. maximal). The strength of the social bond did not significantly predict the proportion of mounts across dyads in nonaggressive contexts, providing no support for the relationship reinforcement hypothesis. Finally, in support of the conflict resolution hypothesis, when individuals engaged in postconflict mounts, 1) the probability of being involved in further aggression decreased from 0.825 to 0.517, while 2) the probability of being involved in grooming interactions with each other increased from 0.119 to 0.606. The strength of the social bond between former opponents had no significant effect on grooming occurrence and agonistic behavior after postconflict mounts. Overall, our findings suggest that nonreproductive mounts in Barbary macaques have different functions that are not affected by the strength of the social bond.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Sociosexual behaviors, or sexual behaviors without a reproductive function, are widespread across animal taxa (e.g., elephants, Elephas maximus: Bailey and Zuk 2009; beluga whales: Lomac-Macnair 2016; spotted hyenas, Crocuta crocuta: East et al. 1993; birds: MacFarlane et al. 2010). Nonreproductive mounts occur when a subject climbs ventrodorsally on a standing partner often with thrusting and typically without penetration. This sociosexual behavior is considered one of the most common ones in the animal kingdom (Thierry 2000; Vasey 1995). In primates, nonreproductive mounts belong to the behavioral repertoire of several species (e.g., bonobos, Pan paniscus: Kano 1980; geladas, Theropithecus gelada: Pallante et al. 2018; baboons, Papio spp.: Owens 1976; Tibetan macaques, Macaca thibetana: Li et al. 2007; pigtailed macaques, Macaca nemestrina: Bernstein 1980; Tonkean macaques, M. tonkeana: De Marco et al. 2014; Barbary macaques, M. sylvanus: Faraut et al. 2015).
Three major and nonmutually exclusive hypotheses have been proposed for the function of nonreproductive mounts: dominance assertion, relationship reinforcement, and conflict resolution (e.g., Preuschoft and van Schaik 2000; Smuts and Watanabe 1990; Wickler 1967). Under the dominance assertion hypothesis, higher-ranking individuals affirm their hierarchical position by mounting lower-ranking individuals (Wickler 1967). This hypothesis has been supported by several studies (e.g., Japanese macaques, Macaca fuscata: Cordischi et al. 1991; mantled guereza, Colobus guereza: Kutsukake et al. 2006; Barbary macaques, Macaca sylvanus: Faraut et al. 2015; golden snub-nosed monkeys, Rhinopithecus roxellana: Huang et al. 2017). Factors other than rank also affect whether the higher-ranking animal in a dyad is the mounter or the mountee, including the mounter’s sex and age, or whether the mount is displayed in an aggressive context (Bernstein 1980; Colmenares 1990, 1991; Hanby et al. 1971; Oi 1990; Owens 1976; Reinhardt et al. 1986; Smuts and Watanabe 1990; Vasey et al. 1998).
The relationship reinforcement hypothesis proposes that nonreproductive mounts and other sociosexual behaviors help to strengthen social relationships and alliance formation in nonaggressive contexts (Smuts and Watanabe 1990). Moreover, they may facilitate peaceful interactions when the risk of aggression from the potential social partner is high, for example, during interactions between animals that rarely interact in a friendly manner or occupy very different dominance positions (Hohmann and Fruth 2000). However, most studies of nonreproductive mounts in nonaggressive contexts (e.g., Dal Pesco and Fischer 2018; De Marco et al. 2014; Fraser and Plowman 2007) include several other behaviors (e.g., penis diddle, embrace, expressive run), so that the relative contribution of mounts to the reinforcement of relationships is unclear.
Finally, the conflict resolution hypothesis proposes that nonreproductive mounts serve as conflict management strategies (Preuschoft and van Schaik 2000). According to this hypothesis, former opponents may exchange a nonreproductive mount in the first minutes after a conflict, as a form of reconciliation. Postconflict nonreproductive mounts may help to reduce the probability of further aggression between former opponents, restore their social relationship, and facilitate other forms of postconflict affiliation, such as grooming (e.g., Aureli et al. 1989; Aureli and van Schaik 1991; Aureli et al. 2002; McFarland and Majolo 2013; Pallante et al. 2018). Moreover, the short-term increase in anxiety that former opponents experience during or after a conflict, as measured by the frequency of self-directed behaviors such as self-scratching, quickly returns to baseline levels if former opponents exchange a nonreproductive mount (e.g., Aureli et al. 19892002; Castles et al. 1996; Cooper et al. 2007; Cords 1992; Hanby 1974; Kano 1980; Majolo et al. 2005; Matsumura 1996; MacFarlane et al. 2010).
The importance of restoring the relationship between former opponents may vary depending on the value of their relationship, so that postconflict mounts should occur more often after conflicts between individuals with a stronger social bond (Call et al. 2002; Clay and de Waal 2013ab; Cords and Aureli 1993; de Waal and Yoshihara 1983; Kappeler and van Schaik 1992). Most studies of the conflict resolution hypothesis have combined nonreproductive mounts with other affiliative behaviors (e.g., grooming, close proximity, social play; McFarland and Majolo 2013). Thus, it is not clear whether nonreproductive mounts alone can reconcile a conflict, or whether they are more frequently exchanged after a conflict between former opponents that have stronger social bonds.
Barbary macaques live in matrilineal groups, in which males migrate but females stay in the natal group for their entire life (Paul and Kuester 1988). Based on its social behavior, the species is described as being relatively tolerant (Thierry 2000). Typically, both kin and nonkin affiliate with similar frequencies after conflicts (Aureli 1997). Nonreproductive mounts are part of the behavioral repertoire used by Barbary macaques for conflict management (Aureli 1997). Higher-ranking males are usually more likely to be the mounter than the mountee in nonreproductive mounts, while individuals engaging in postconflict mounts are more likely to exchange grooming and have a lower rate of self-scratching than individuals in matched-control sessions (Faraut et al. 2015). However, it remains unclear whether engaging in postconflict mounts also reduces the probability of further aggression in a dyad, because the time window used in previous studies to measure the occurrence of further aggression (Faraut et al. 2015) may have been too narrow (for comparison, see Patzelt et al. 2009).
We aimed to test the relative importance of the three hypothesized functions of nonreproductive mounts (i.e., dominance assertion, relationship reinforcement, and conflict resolution) in two groups of semi-free-ranging Barbary macaques. Here, we expanded on previous work (Faraut et al. 2015) by 1) collecting data in both aggressive and nonaggressive contexts, 2) including direct measures of social bonds between individuals (to better test the relationship reinforcement hypothesis), and 3) directly comparing postconflict periods including mounts to postconflict periods including no mounts (as matched controls) involving the same dyads. If nonreproductive mounts serve to assert dominance, individuals with a higher rank in the dyad should have a higher probability of being the mounter than subordinates, both in aggressive and nonaggressive contexts (prediction 1). If nonreproductive mounts serve to strengthen social bonds, the probability of engaging in mounts during nonaggressive contexts should be higher in dyads with stronger relationships than dyads with weaker social bonds (prediction 2). If nonreproductive mounts act as a form of reconciliation, individuals engaging in a postconflict mount should have a lower probability of being involved in further aggression than when they do not mount (prediction 3a). Moreover, postconflict mounts should be more frequent between former opponents with a stronger rather than a weaker social bond (prediction 3b), because the former gain greater benefits from reconciling a conflict than the latter (Aureli et al. 2002; Majolo et al. 2009). Finally, if nonreproductive mounts serve to facilitate further postconflict affiliation between former opponents, individuals engaging in a postconflict mount should have a higher probability of grooming after the mount than when they do not mount (prediction 3c).
Methods
Subjects
Three observers recorded behavioral data on two groups of semi-free-ranging Barbary macaques at La Montagne des Singes (Kintzheim, France). The two groups consisted of 62 (group A) and 56 (group B) individuals of both sexes and different ages (Table I), which we could recognize individually thanks to their physical characteristics and codes tattooed on their chest. Group B shared their enclosure (11 ha) with another group of Barbary macaques, which we did not study. Although the two groups were free to move in the enclosure, they always occupied different areas. Intergroup interactions were rare, so we excluded them from analysis. Group B spent most of the time in a small wooded area next to an open area with sparse vegetation, crossed by a tourist trail. They were all habituated to humans. During observations we achieved optimal visibility by having one observer in each part of the enclosure frequented by the subjects (i.e., the small wood and the open area). Subjects fed on natural vegetation, but the park staff additionally spread vegetables, fruit, pellets, and wheat all around the area three times a day, to ensure adequate food for all subjects. Fresh water was provided ad libitum. Along the trail, tourists were allowed to feed the monkeys with popcorn distributed at the park entrance. The monkeys appeared to like popcorn, but we saw no conflicts over it during our observations (likely because popcorn was relatively abundant). Subjects in group A (enclosure area: 7 ha) lived in very similar conditions, except that they were not visited by tourists and were thus less habituated to humans.
Procedures
Between September 2016 and June 2017, three researchers conducted behavioral observations on all adults (males ≥5 yr of age; females ≥4 yr), subadults (males 3–4 yr; females 2–3 yr) and juveniles (males 1–3 yr; females 1–2 yr) in the study groups. We conducted behavioral observations based on the ethogram by Thierry and colleagues (2000). The researchers started collecting data independently only after conducting>30 h of observations on the same subjects and reaching interobserver reliability >90%. We estimated interobserver reliability by comparing multiple random samples of behavior (Kaufman and Rosenthal 2009).
We recorded dyadic agonistic interactions using all-occurrences sampling and analyzed interactions with a clear winner–loser outcome to determine the dominance hierarchy and individual rank values using the Elo method (EloRating package, version 0.43; Neumann et al. 2011). We set the k factor at 100 and the starting values at 1000. We took the mean of the Elo values for each individual for the whole study period and standardized them to range 0–1. We refer to these values as Elo ranks, where a value of 0 indicates the lowest rank and a value of 1 the highest one. We ran separate analyses for each group using 1412 interactions for group A and 1253 for group B. We first assessed the degree of hierarchy stability via visual inspection and by using the stab_elo function, which assesses the ratio of rank changes in all individuals across consecutive days (for more details, see Neumann and Kulik 2020; Neumann et al. 2011). The Elo ranks were very stable in both groups (group A, 0.989; group B, 0.925, with 0 suggesting minimum stability with daily rank reversals and 1 suggesting high stability with no rank reversals), so we included no burn in periods.
We assessed social networks by recording the closest individual to each animal hourly using group scans (393 group scans in group A, 389 in group B). We used these measures to create an undirected weighted matrix and ran analyses with several R packages (vegan 2.5–3: Oksanen et al. 2019; asnipe 1.1.10: Farine 2018; igraph 1.2.1: Csardi and Nepusz 2006) to assess eigenvector centrality as the sum of the centralities of an individual’s neighbors (Farine 2017; Farine and Whitehead 2015).
We also conducted 20-min focal samples on all the adults, subadults, and juveniles in the groups. We chose the daily focal individuals via a random permutation procedure with ≥60 min between focal follows of the same subject. During focal samples we recorded the duration of grooming involving the subject and the partner identity. We then used grooming duration to assess the bond strength in each dyad as the proportion of time spent grooming out of the sum of focal time of both individuals, and then divided these values to obtain the mean grooming value across all dyads (mean and range of grooming index: 1, 0–50.29). In group A, 74 out of the 861 possible dyads had a social bond higher than the mean and therefore were considered as having a strong bond (dyadic mean index = 6.48). Using the same criterion, in group B 241 out of the 1596 possible dyads were considered to have a strong bond (dyadic mean index = 4.83).
For 3 mo, between March and May 2017, we recorded all occurrences of nonreproductive mounts involving two males (N = 150) or one female and one male (N = 82) ad libitum. Female–female mounts were rare (N = 3), so we excluded them from analyses. We included individuals of all age classes, except for infants. We defined nonreproductive mounts as ventrodorsal sexual contacts with thrusting movements (Thierry et al. 2000; Vasey 1995). We excluded ventrodorsal embraces from the analyses. Since Barbary macaques are seasonal breeders, and since we collected data on mounts outside the mating season (winter), all the mounts we observed were nonreproductive. For each observed mount, we recorded 1) partner identity; 2) the number of other individuals within 10 m; (iii) whether the mount happened within 10s after aggression between the partners; (iv) whether mounter and mountee were involved in further aggression in the hour following the mount (which is an appropriate time window to include all occurrences of further aggression, which typically range between 21 and 1588 seconds in this species; see Patzelt et al. 2009); and (v) whether the dyad exchanged affiliative behaviors in the minute following the mount (former opponents in Barbary macaques mostly exchange affiliative behavior in the first minute postconflict: Aureli et al. 1994). Moreover, for each mount immediately following an aggression (i.e., postconflict mounts), we conducted two matched-control sessions, one lasting 1 min and one lasting 1 h. We conducted matched-control sessions on the next day on which an aggression involved the same subjects, but no postconflict mount occurred. In 1-min matched-control sessions, we assessed the occurrence of affiliative behavior in the dyad in the minute following the aggression. In 1-h matched-control sessions, we assessed the occurrence of a further conflict in the dyad in the hour following the aggression. For the 41 postconflict mounts we recorded, we conducted 41 1-min matched controls, and 32 1-h matched controls (in 9 cases, we could not record the matched controls because the mounter and/or mountee were not visible). We conducted 46% of 1-min matched-control sessions in the week following the postconflict mount, 37% after 1 week but in the following month, and 17% after 1 mo. We conducted 34% of the 1-h matched controls in the week following the matched postconflict mount, 38% after 1 week but in the following month, and 28% after 1 mo.
Statistical Analyses
We performed all analyses in R (R Core Team, version 3.5.0), using the glmmTMB package (version 1.0.1; Brooks et al. 2017) to build generalized linear mixed models (Bolker et al. 2009). To test the dominance assertion hypothesis (prediction 1), we entered one line for each of the two individuals involved in each mount (N = 232). We ran a binomial model to assess whether being the mounter, within each dyad, was predicted by the 2-way interaction of mount context (i.e., whether the mount immediately followed a conflict involving the mounter and/or mountee) with the rank difference between the individual and the partner (also including them as main effects). We included the individuals’ age class, whether the individual had a higher centrality in the dyad, and the number of other group members being present as controls. We excluded individual sex as control predictor because no females ever acted as mounters. We included subject and mount identity as random factors (model 1).
To test the relationship reinforcement hypothesis (prediction 2), we entered one line for each possible male–male and male–female dyad (infants excluded; N = 1578), including only mounts which did not follow a conflict (N = 126). As the dependent variable, we entered the proportion of mounts observed in the dyad out of the overall number of mounts observed in the group, using the cbind function. We then ran a binomial model to assess whether the strength of the social bond predicted the proportion of mounts, controlling for absolute rank difference and sex combination, and including the identities of both individuals as random factors (model 2).
To test the conflict resolution hypothesis, we ran three models. First, we entered one line for each mount (including only mounts that followed an aggressive interaction; N = 32), and a further line for the matched control (i.e., an aggressive interaction between the same two individuals, not followed by a mount; N = 32). We then ran a binomial model to assess whether the occurrence of affiliative interactions in the dyad in the minute following a conflict was predicted by the occurrence of a mount (i.e., postconflict, N = 41, or matched control, N = 41) and the strength of the dyadic social bond (in a two-way interaction and as main effects), controlling for age and sex combination, including the identities of both individuals as random factors (model 3c, testing prediction 3c). Finally, we ran a binomial model to assess whether the occurrence of further aggression in the dyad in the following hour was predicted by the occurrence of a mount (i.e., whether it was a postconflict mount or matched control) and the strength of the dyadic social bond in a two-way interaction and as main effects, controlling for age and sex combination, including the identities of both individuals as random factors (model 3a, testing prediction 3a). To assess whether postconflict mounts are more likely in dyads with stronger social bonds, we entered one line for each possible male–male and male–female dyad (infants excluded; N = 1578), and used the proportion of postconflict mounts observed in the dyad out of the overall number of mounts observed in the group, using the cbind function as a dependent variable. We then ran a binomial model to assess whether the strength of the social bond predicted the proportion of dyadic postconflict mounts, controlling for absolute rank difference and sex combination, including the identities of both individuals as random factors (model 3b, testing prediction 3b).
We performed full–null model comparisons using a likelihood ratio test (Dobson 2002) to test whether the inclusion of predictors improved model fit; the null model included only the control variables and the random factors. When the difference between full and null model was significant, we conducted a likelihood ratio test and obtained the P-values for each predictor variable using single-term elimination, with the drop1 function in R (Barr et al. 2013). We removed all nonsignificant two-way interactions from the model and included the main terms as predictors in the model. We detected no convergence issues in the models and ruled out collinearity based on Variance Inflaction Factors (VIFs; Field 2005) (maximum VIFs across all models = 1.54). We excluded overdispersion of the models using the R package qcc (Scrucca 2004).
Ethical Note
This research was carried out with permission from La Montagne des Singes. The authors declare they have no conflict of interest.
Data Availability
The data sets supporting these findings are provided as Electronic Supplementary Material.
Results
When testing the dominance assertion hypothesis, the full–null model comparison was significant (model 1: χ2 = 20.50, df = 3, P < 0.001). In particular, the probability of being mounter in the dyad was significantly predicted by the two-way interaction of context and rank difference between the individuals (P = 0.012). When rank difference increased, the probability that the higher-ranking animal was the mounter also increased, especially after aggression. In nonaggressive contexts, in particular, the probability to be the mounter increased from 0.066 (when the difference between the individual’s and the partner’s rank was minimal, i.e., when the individual had a much lower rank than the partner) to 0.797 (when the rank difference was maximal, i.e., when the individual had a much higher rank than the partner), with an increase of 0.964 in the probability to be the mounter for every 1-unit increase in rank difference. In aggressive contexts, the probability to be the mounter increased significantly more quickly, from 0.011 (when the rank difference was minimal) to 0.969 (when the rank difference was maximal), with an increase of 0.998 in the probability to be the mounter for every 1-unit increase in rank difference.
When testing the relationship reinforcement hypothesis, the full model did not significantly differ from the null model (model 2: χ2 = 0.08, df = 1, P = 0.776). The strength of the social bond did not significantly predict the proportion of nonaggressive mounts across dyads (Table II).
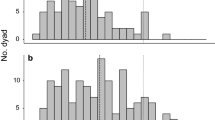
When testing prediction 3a of the conflict resolution hypothesis the full–null model comparison was significant (model 3a: χ2 = 9.78, df = 3, P = 0.021). The occurrence of postconflict mounts significantly predicted a lower probability of further aggression in the dyad (0.517) in comparison to conflict not followed by a mount (0.825), regardless of the strength of their social bond (Table II, Fig. 1).

Number of aggressive events that were followed by a mount (Postconflict) or not (i.e., Matched Control), and that were followed by another aggression (in gray) or not (in black). The data on the bars indicate the number of events and the relative percentage of each category (out of the total aggressive events observed). Data were collected on the Barbary macaques at La Montagne des Singes, France, between March 2017 and May 2017.
When testing prediction 3b, the full model did not significantly differ from the null model (model 3b: χ2 = 0.08, df = 1, P = 0.776), and the strength of the social bond did not significantly predict the proportion of postconflict mounts across dyads (Table II).
Finally, when testing prediction 3c, the full–null model comparison was significant (model 3c: χ2 = 20.59, df = 3, P < 0.001). In particular, the occurrence of postconflict mounts significantly predicted a greater probability of grooming between former opponents (0.606) in comparison to conflicts that were not followed by a mount (0.119) (Fig. 2). The strength of the social bond between former opponents, their age or sex combination had no significant effect on the occurrence of postconflict grooming (Table II).

Number of aggressive events that were followed by a mount (Postconflict) or not (i.e., Matched Control), and that were followed by grooming behavior (in gray) or not (in black). The data on the bars indicate the number of events and the relative percentage of each category (out of the total aggressive events observed). Data were collected on the Barbary macaques at La Montagne des Singes, France, between March 2017 and May 2017.
Discussion
Overall, our study supports the dominance assertion hypothesis and the conflict resolution hypothesis, but not the relationship reinforcement hypothesis. In line with previous findings in primates (e.g., Cordischi et al. 1991; Faraut et al. 2015; Huang et al. 2017; Kutsukake et al. 2006), higher ranking individuals were more likely to mount than to be mounted, after controlling for age, sex, social integration, and presence of other group members (dominance assertion hypothesis). This was true both in nonaggressive and aggressive contexts, but especially in the latter. This pattern probably occurs because dominance assertion is especially important during or immediately after a conflict, when social uncertainty is higher and rank-changing coalitions are more likely (Bissonnette et al. 2015; Young et al. 2014b).
Our study did not support the relationship reinforcement hypothesis (Smuts and Watanabe 1990), in that dyads with stronger social bonds were not more likely to engage in mounts in nonaggressive contexts, as compared to dyads with weaker social bonds. This contrasts with studies showing that mounts and other sociosexual behaviors strengthen social bonds and alliance formation in nonaggressive contexts (Dal Pesco and Fischer 2018; De Marco et al. 2014; Fraser and Plowman 2007; Smuts and Watanabe 1990). There are at least two reasons for these differences. First, our study is based on a relatively small sample size, because nonreproductive mounts and conflicts occur at low rates even between animals that share a strong social bond (Ventura et al. 2006). It is therefore possible that longer-term studies using a composite index to assess social bonds (sensu Silk et al. 2013) would show a link between strength of social bonds and occurrence of nonreproductive mounts, as 1) longer studies and 2) more detailed social bond indexes (including, e.g., proximity or body contact) may be needed to reliably differentiate social relationships in macaques (see Davis et al. 2018). Second, in contrast to previous studies (e.g., Dal Pesco and Fischer 2018; De Marco et al. 2014), we specifically examined nonreproductive mounts and excluded other sociosexual behaviors (e.g., genital touch, ventroventral embrace, antiparallel embrace). Therefore, it is possible that our study differs from older ones because mounts have a different function from other sociosexual behaviors.
Our study provided some support for the conflict resolution hypothesis (Aureli et al. 2002; de Waal and van Roosmalen 1979). In particular, individuals engaging in postconflict mounts were less likely to exchange further aggression than those that engaged in a conflict but not in a mount. These results contrast with previous studies of reconciliation in Barbary macaques, which found that reconciliation does not decrease the risk of further aggression, particularly for the victim (Faraut et al. 2015; McFarland and Majolo 2011a; Patzelt et al. 2009). Instead, our results suggest that postconflict mounts reduce aggression between former opponents and thus exert an important function in conflict management. These different results may be due to methodological differences in these studies. First, when comparing postconflict mount to matched-control sessions following an aggression, we selected matched-control sessions that included conflicts between exactly the same subjects as in the postconflict mount session. In this way, we were able to test the effect of nonreproductive mounts in conflict management more precisely than studies using different dyads in postconflict mount and matched-control sessions. Second, by using longer postconflict and matched-control sessions (i.e., 1 h instead of 10 or 20 min in Faraut et al. 2015; McFarland and Majolo 2011a; Patzelt et al. 2009), we could better detect whether nonreproductive mounts decrease the probability of further aggression: during an aggression, subordinate subjects typically run far away from the aggressor and therefore victims may not be in the aggressor’s visual range in the 10–20 min after an aggression.
In line with the conflict resolution hypothesis, engaging in postconflict mounts increased the probability of grooming between former opponents. If postconflict mounts simultaneously indicate that the conflict is over and that the subordinate “acknowledges” the higher-ranking position of the dominant former opponent, then no further aggression is needed. This is particularly true if, as we found, postconflict mounts increase the likelihood of grooming exchange between former opponents, because such affiliative interaction eliminates the need for the dominant to coerce grooming through aggression (McFarland and Majolo 2011b). These results are in line with previous work on the occurrence of postconflict affiliative behavior, with postconflict mounts significantly increasing the probability of affiliative behavior after a conflict (Aureli et al. 1994; Faraut et al. 2015; McFarland and Majolo 2011a; Patzelt et al. 2009).
For nonaggressive contexts, our study revealed no link between strength of social bonds and the probability of mounts after an aggression. This contrasts with studies of reconciliation (Call et al. 1999; McFarland and Majolo 2011a; Patzelt et al. 2009). However, our results are comparable to those of previous studies of postconflict sexual behaviors that also found no relation between rate of postconflict sexual behaviors and strength of social bonds (Call et al. 2002; Clay and de Waal 2013ab; Moscovice et al. 2017; Tokuyama and Furuichi 2016). Since the frequency of nonreproductive mounts is affected by the rank difference between the mounter and the mountee, we argue that the dominance assertion function of the mounts may mask or reduce the use of this behavior for establishing or maintaining social bonds.
Finally, sex and sex combination (which we included as control in the models) strongly affected our response variables. In particular, females were never mounters in the dyad (model 1), and male–male dyads had a higher probability of engaging in mounts than male–female ones, both in aggressive (model 3b) and nonaggressive contexts (model 2). These results suggest that mounts may be especially relevant for male–male dyads to reduce the negative effects of conflicts. Moreover, being the mounter in nonreproductive mounts may not be part of the behavioral repertoire of Barbary macaque females and thus they may use other behaviors to resolve conflicts.
Overall, our study showed that mounts appear to be multifunctional, serving to assert dominance, reduce conflict escalation, and facilitate postconflict affiliative interactions, regardless of the social bond between former opponents. Animals may employ a range of different behaviors to achieve the same goal and studies sometimes integrate different behaviors into broad categories. However, studies like ours that target specific sexual behaviors in different contexts are important to investigate the function of each behavior within the behavioral repertoire of a species.
References
Aureli, F. (1997). Post-conflict anxiety in nonhuman primates: The mediating role of emotion in conflict resolution. Aggressive Behavior, 23, 315–328. https://doi.org/10.1002/(SICI)1098-2337(1997)23:57<315::AID-AB2>3.0.CO;2-H.
Aureli, F., Cords, M., & Van Schaik, C. P. (2002). Conflict resolution following aggression in gregarious animals: A predictive framework. Animal Behaviour, 64(3), 325–343. https://doi.org/10.1006/anbe.2002.3071.
Aureli, F., Das, M., Verleur, D., & van Hooff, J. A. R. M. (1994). Postconflict social interactions among barbary macaques (Macaca sylvanus). International Journal of Primatology, 15(3), 471–485.
Aureli, F., & Van Schaik, C. P. (1991). Post-conflict behaviour in long-tailed macaques (Macaca fascicularis). Ethology, 89, 89–100. https://doi.org/10.1111/j.1439-0310.1991.tb00297.x.
Aureli, F., Van Schaik, C. P., & Van Hooff, J. A. R. A. M. (1989). Functional aspects of reconciliation among captive long-tailed macaques (Macaca fascicularis). American Journal of Primatology, 19, 39–51. https://doi.org/10.1002/ajp.1350190105.
Bailey, W., & Zuk, M. (2009). Same-sex sexual behavior and evolution. Trends in Ecology & Evolution, 24(8), 439–460. https://doi.org/10.1016/j.tree.2009.03.014.
Barr, D. J., Levy, R., Scheepers, C., & Tily, H. J. (2013). Random effects structure for confirmatory hypothesis testing: Keep it maximal. Journal of Memory and Language, 68(3), 255–278.
Bernstein, I. S. (1980). Activity patterns in a stumptail macaque group (Macaca arctoides). Folia Primatologica, 33, 20–45. https://doi.org/10.1159/000155926.
Bissonnette, A., Perry, S., Barrett, L., Mitani, J. C., Flinn, M., et al (2015). Coalitions in theory and reality: A review of pertinent variables and processes. Behaviour, 152(1), 1–56.
Bolker, B. M., Brooks, M. E., Clark, C. J., Geange, S. W., Poulsen, J. R., et al (2009). Generalized linear mixed models: A practical guide for ecology and evolution. Trends in Ecology & Evolution, 24(3), 127–135. https://doi.org/10.1016/j.tree.2008.10.008.
Brooks, M., Kristensen, K., van Benthem, K. J., Magnusson, A., Berg, C. W., et al (2017). glmmTMB balances speed and flexibility among packages for zeroinflated generalized linear mixed modeling. The R Journal, 9(2), 378–400.
Call, J., Aureli, F., & de Waal, F. B. M. (1999). Reconciliation patterns among stumptail macaques: A multivariate approach. Animal Behaviour, 58, 165–172.
Call, J., Aureli, F., & de Waal, F. B. M. (2002). Postconflict third-party affiliation in stumptailed macaques. Animal Behaviour, 63(2), 209–216. https://doi.org/10.1006/anbe.2001.1908.
Castles, D. L., Aureli, F., & de Waal, F. B. M. (1996). Variation in conciliatory tendency and relationship quality across groups of pigtail macaques. Animal Behaviour, 52, 389–403.
Clay, Z., & de Waal, F. B. M. (2013a). Bonobos respond to distress in others: Consolation across the age spectrum. PLoS ONE, 8, e55206.
Clay, Z., & de Waal, F. B. M. (2013b). Development of socio-emotional competence in bonobos. Proceeding of the National Academy of Sciences of the USA, 110, 18121–18126.
Colmenares, F. (1990). Greeting behaviour in male baboon, I: Communication, reciprocity and symmetry. Behaviour, 113, 81–116.
Colmenares, F. (1991). Greeting behaviour between male baboons: Oestrous females. rivalry and negotiation. Animal Behaviour, 41, 49–60.
Cooper, M. A., Aureli, F., & Singh, M. (2007). Sex differences in reconciliation and post-conflict anxiety in bonnet macaques. Ethology, 113, 26–38.
Cordischi, C., Cozzolino, R., Aureli, F., & Scucchi, S. (1991). Influence of context on mounting and presenting among mature male Japanese macaques. Folia Primatologica, 56, 211–213.
Cords, M. (1992). Post-conflict reunions and reconciliation in long-tailed macaques. Animal Behaviour, 44, 57–61.
Cords, M., & Aureli, F. (1993). Patterns of reconciliation among juvenile long-tailed macaques. In M. E. Pereira & L. A. Fairbanks (Eds.), Juvenile primates: Life history, development and behavior (pp. 271–284). New York: Oxford University Press.
Csardi, G., & Nepusz, T. (2006). The Igraph software package for complex network research (p. 1695). International Journal: Complex Systems http://igraph.org.
Dal Pesco, F., & Fischer, J. (2018). Greetings in male Guinea baboons and the function of rituals in complex social groups. Journal of Human Evolution, 125, 87–98. https://doi.org/10.1016/j.jhevol.2018.10.007.
Davis, G. H., Crofoot, M. C., & Farine, D. R. (2018). Estimating the robustness and uncertainty of animal social networks using different observational methods. Animal Behaviour, 141, 29–44.
De Marco, A., Sanna, A., Cozzolino, R., & Thierry, B. (2014). The function of greetings in male Tonkean macaques. American Journal of Primatology, 76, 989–998. https://doi.org/10.1002/ajp.22288.
de Waal, F. B. M., & van Roosmalen, A. (1979). Reconciliation and consolation among chimpanzees. Behavioral Ecology and Sociobiology, 5, 55–66. https://doi.org/10.1007/BF00302695.
de Waal, F. B. M., & Yoshihara, D. (1983). Reconciliation and redirected affection in rhesus monkeys. Behaviour, 85, 241.
Dobson, A. J. (2002). An introduction to generalized linear models. Boca Raton: Chapman & Hall.
East, M. L., Hofer, H., & Wickler, W. (1993). The erect ‘penis’ is a flag of submission in a female-dominated society: Greetings in Serengeti spotted hyenas. Behavioral Ecology and Sociobiology, 33, 355–370. https://doi.org/10.1007/BF00170251.
Faraut, L., Northwood, A., & Majolo, B. (2015). The functions of non-reproductive mounts among male Barbary macaques (Macaca sylvanus). American Journal of Primatology, 77(11), 1149–1157. https://doi.org/10.1002/ajp.22451.
Farine, D. (2018). asnipe. https://www.rdocumentation.org/packages/asnipe.
Farine, D., & Whitehead, H. (2015). Constructing, conducting and interpreting animal social network analysis. The Journal of Animal Ecology, 84, 1144–1163.
Farine, D. R. (2017). A guide to null models for animal social network analysis. Methods in Ecology and Evolution, 8, 1309–1320. https://doi.org/10.1111/2041-210X.12772.
Field, A. P. (2005). Discovering statistics using SPSS. 2nd Edition, Sage Publications, London.
Fraser, O., & Plowman, A. B. (2007). Function of notification in Papio hamadryas. International Journal of Primatology, 28, 1439–1448.
Hanby, J. P. (1974). Male-male mounting in Japanese monkeys (Macaca fuscata). Animal Behaviour, 22, 836–849.
Hanby, J. P., Robertson, L. T., & Phoenix, C. H. (1971). Sexual behaviour in a confined troop of Japanese macaques. Folia Primatologica, 16, 123–143.
Hohmann, G., & Fruth, B. (2000). Use and function of genital contacts among female bonobos. Animal Behaviour, 60, 107–120.
Huang, P., He, X., Zhang, E., & Chen, M. (2017). Do same-sex mounts function as dominance assertion in male golden snub-nosed monkeys (Rhinopithecus roxellana)? American Journal of Primatology, 79, e22636. https://doi.org/10.1002/ajp.22636.
Kano, T. (1980). Social behavior of wild pygmy chimpanzees (Pan paniscus) of Wamba: A preliminary report. Journal of Human Evolution, 9, 255–260.
Kappeler, P. M., & Van Schaik, C. P. (1992). Methodological and evolutionary aspects of reconciliation among primates. Ethology, 92, 51–69.
Kaufman, A. B., & Rosenthal, R. (2009). Can you believe my eyes? The importance of interobserver reliability statistics in observations of animal behaviour. Animal Behaviour, 78(6), 1487–1491.
Kutsukake, N., Suetsugu, N., & Hasegawa, T. (2006). Pattern, distribution, and function of greeting behavior among black-and-white colobus. International Journal of Primatology, 27, 1271–1291.
Li, J., Yin, H., & Zhou, L. (2007). Non-reproductive copulation behavior among Tibetan macaques (Macaca thibetana) at Huangshan. China. Primates, 48(1), 64–72. https://doi.org/10.1007/s10329-006-0002-5.
Lomac-Macnair, K. S., Smultea, M. A., Cotter, M. P., Thissen, C., & Parker, L. (2016). Socio-sexual and probable mating behavior of cook inlet beluga whales, Delphinapterus leucas, observed from an aircraft. Marine Fisheries Review. https://doi.org/10.7755/MFR.77.2.2.
MacFarlane, G. R., Blomberg, S. P., & Vasey, P. L. (2010). Homosexual behaviour in birds: Frequency of expression is related to parental care disparity between the sexes. Animal Behaviour, 80(3), 375–390. https://doi.org/10.1016/j.anbehav.2010.05.009.
Majolo, B., Ventura, R., & Koyama, N. F. (2005). Postconflict behavior among male Japanese macaques. International Journal of Primatology, 26, 321–336.
Majolo, B., Ventura, R., & Koyama, N. F. (2009). Anxiety level predicts post-conflict behaviour in wild Japanese macaques (Macaca fuscata yakui). Ethology, 115, 986–995. https://doi.org/10.1111/j.1439-0310.2009.01685.x.
Matsumura, S. (1996). Post-conflict affiliative contacts between former opponents among wild moor macaques (Macaca maurus). American Journal of Primatology, 38, 211–219.
McFarland, R., & Majolo, B. (2011a). Reconciliation and the costs of aggression in wild Barbary macaques (Macaca sylvanus): A test of the integrated hypothesis. Ethology, 117, 928–937. https://doi.org/10.1111/j.1439-0310.2011.01948.x.
McFarland, R., & Majolo, B. (2011b). Grooming coercion and the post-conflict trading of social services in wild Barbary macaques. PLoS ONE, 6(10), e26893.
McFarland, R., & Majolo, B. (2013). The importance of considering the behavioral form of reconciliation in studies of conflict resolution. International Journal of Primatology, 34, 15–29. https://doi.org/10.1007/s10764-012-9643-y.
Moscovice, L. R., Douglas, P. H., Martinez-Iñigo, L., Surbeck, M., Vigilant, L., & Hohmann, G. (2017). Stable and fluctuating social preferences and implications for cooperation among female bonobos at LuiKotale, Salonga National Park, DRC. American Journal of Physical Anthropology, 163, 158–172. https://doi.org/10.1002/ajpa.23197.
Neumann, C., Duboscq, J., Dubuc, C., Ginting, A., Irwan, A. M., et al (2011). Assessing dominance hierarchies: Validation and advantages of progressive evaluation with Elo-rating. Animal Behaviour, 82(4), 911–921. https://doi.org/10.1016/j.anbehav.2011.07.016.
Neumann, C., & Kulik, L. (2020). EloRating: A brief tutorial. https://rdrr.io/cran/EloRating/f/inst/doc/EloRating_tutorial.pdf.
Oi, T. (1990). Patterns of dominance and affiliation in wild pigtailed macaques (Macaca nemestrina nemestrina) in West Sumatra. International Journal of Primatology, 11, 339–356.
Oksanen, J., Blanchet, G., Friendly, M., Kindt, R., Legendre, P., et al.. (2019). An R package for community ecologists. https://cran.r-project.org/web/packages/vegan/index.html.
Owens, N. W. (1976). The development of socio-sexual behaviour in free-living baboons. Papio anubis. Behaviour, 57, 241–259.
Pallante, V., Stanyon, R., & Palagi, E. (2018). Calming an aggressor through spontaneous post-conflict triadic contacts: Appeasement in Macaca tonkeana. Aggressive Behavior, 44, 406–415.
Patzelt, A., Pirow, R., & Fischer, J. (2009). Post-conflict affiliation in Barbary macaques is influenced by conflict characteristics and relationship quality, but does not diminish short-term renewed aggression. Ethology, 115, 658–670. https://doi.org/10.1111/j.1439-0310.2009.01649.x.
Paul, A., & Kuester, J. (1988). Life-history patterns of Barbary macaques (Macaca sylvanus) at Affenberg Salem. In J. E. Fa & C. H. Southwick (Eds.), Ecology and behavior of food-enhanced primate groups (pp. 199–228). New York: Alan R. Liss.
Preuschoft, S., & van Schaik, C. P. (2000). Dominance and communication. In F. Aureli & F. B. M. de Waal (Eds.), Natural conflict resolution (pp. 77–105). Berkeley: University of California Press.
Reinhardt, V., Reinhardt, A., Bercovitch, F. B., & Goy, R. W. (1986). Does intermale mounting function as a dominance demonstration in rhesus monkeys? Folia Primatologica, 47, 55–60.
Scrucca, L. (2004). qcc: An R package for quality control charting and statistical process control. R News, 4(1), 11–17.
Silk, J., Cheney, D., & Seyfarth, R. (2013). A practical guide to the study of social relationships. Evolutionary Anthropology, 22(5), 213–225. https://doi.org/10.1002/evan.21367.
Smuts, B. B., & Watanabe, J. M. (1990). Social relationships and ritualized greetings in adult male baboons (Papio cynocephalus anubis). International Journal of Primatology, 11, 147–172.
Thierry, B. (2000). Covariation of conflict management patterns across macaque species. In F. Aureli & F. B. M. de Waal (Eds.), Natural conflict resolution (pp. 106–128). University of California Press.
Thierry, B., Bynum, E. L., Baker, S., Kinnaird, M. F., Matsumura, S, Muroyama Y, et al. (2000). The social repertoire of Sulawesi macaques. Primate Research, 16, 203–226. https://doi.org/10.2354/psj.16.203.
Tokuyama, N., & Furuichi, T. (2016). Do friends help each other? Patterns of female coalition formation in wild bonobos at Wamba. Animal Behaviour, 119, 27–35. https://doi.org/10.1016/j.anbehav.2016.06.021.
Vasey, P. L. (1995). Homosexual behavior in primates: A review of evidence and theory. International Journal of Primatology, 16, 173–204.
Vasey, P. L., Bernard, C., & Gauthier, C. (1998). Mounting interaction between female Japanese macaques: Testing the influence of dominance & aggression. Ethology, 104, 387–398.
Ventura, R., Majolo, B., Koyama, N. F., Hardie, S., & Schino, G. (2006). Reciprocation and interchange in wild Japanese macaques: Grooming, cofeeding and agonistic support. American Journal of Primatology, 68(12), 1138–1149.
Wickler, W. (1967). Sociosexual signals and their intra-specific imitation among primates. In D. Morris (Ed.), Primate ethology (pp. 69–147). Aldine.
Young, C., Majolo, B., Heistermann, M., Schülke, O., & Ostner, J. (2014a). Responses to social and environmental stress are attenuated by strong male bonds in wild macaques. Proceedings of the National Academy of Sciences of the USA, 111, 18195–18200.
Young, C., Majolo, B., Schülke, O., & Ostner, J. (2014b). Male social bonds and rank predict supporter selection in cooperative aggression in wild Barbary macaques. Animal Behaviour, 95, 23–32. https://doi.org/10.1016/j.anbehav.2014.06.007.
Acknowledgments
We are extremely grateful to Ellen Merz for providing extensive and generous support throughout the study, as well as thoughtful comments on the manuscript. We further thank La Montagne des Singes for allowing us access to the monkeys, and to the whole staff, who kindly supported our investigation. We thank María Teresa Martínez Navarrete, who contributed to the data collection; Lauriane Faraut for the kind comments; Roger Mundry for the precious R functions; and the editor and two anonymous reviewers for their suggestions and comments.
Funding
Open Access funding enabled and organized by Projekt DEAL.
Author information
Authors and Affiliations
Corresponding author
Additional information
Handling Editor: Joanna Setchell.
Supporting Information
ESM 1
(XLSX 156 kb)
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Anzá, S., Majolo, B. & Amici, F. The Function of Mounts in Free-Ranging Barbary Macaques (Macaca sylvanus). Int J Primatol 42, 426–441 (2021). https://doi.org/10.1007/s10764-021-00210-w
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10764-021-00210-w