
Abstract
The glass eel catch from the international River Minho, western Iberian Peninsula, forms the southernmost Atlantic series in the composite index outside the North Sea assessing European eel recruitment. Here, new experimental yield data from the lower estuary (1981–2022) and fishers´ daily records (1990–2022) were modelled to describe and compare seasonal and interannual trends. Seasonality matched river discharge climatology, possibly a more general feature in the southern range of the species distribution, with the difference between high and low season becoming less marked in recent periods of lower abundance. Glass eel yield showed a sharp decline during the 1980s, in line with the recruitment index outside the North Sea and with total Minho catch, but not with recent local estimates of catch per unit effort. This decline is corroborated by trends in fishers´ daily records, suggesting that the reduction in effort in the Minho was the consequence of a gradual adjustment to recruitment in the catchment mediated by progressively tighter local management measures. The inefficiency of fisheries regulatory measures to guarantee stock recovery alone calls for more comprehensive measures to reverse anthropogenic impacts on continental eel populations, but the task seems Herculean.
Similar content being viewed by others

Introduction
Anguillid eels have complex life cycles that may require cumulative exposure to oceanic, coastal, estuarine, and inland habitats across their life stages (Tesch, 2003). Their wide distribution, predatory behaviour, and diversity of habitats required to complete their life cycle also make eels suitable surrogate species to monitor biodiversity conservation, especially in freshwaters (Itakura et al., 2020). Being of marine origin, the genus retains marine spawning with oceanic dispersal of larvae (leptocephali) until reaching continental waters. On arrival to the continental shelf, the larvae metamorphose into glass eels and generally enter the estuaries of the nearest rivers. After several years of growth (yellow eel phase), the duration of which depends on sex, habitat productivity and latitude, yellow eels undergo a metamorphosis into silver eels and initiate a migration back to their spawning grounds, where they reproduce and die (Jellyman, 2022).
Currently, most species of the genus Anguilla are under some conservation strain. In addition to changes in oceanic conditions, eels have been subject to a host of anthropogenic pressures, such as habitat loss and fragmentation, obstacles to migration, pollution, introduction of parasites and overexploitation, that have acted in their continental habitats for decades (Jacoby et al., 2015) and synergistically led to the current situation of generalised threat (Drouineau et al., 2018). The most prominent conservation risk is for the European eel (Anguilla anguilla (L.)). With a distribution area that spans more than ten thousand km across the temperate north-eastern Atlantic Ocean and the Mediterranean Sea (Tesch, 2003), the European eel remains Critically Endangered in the latest IUCN Red List assessment (Pike et al., 2020) and is listed in Appendix II of CITES, which prohibits international trade. Perilous IUCN classifications have also been attributed to other temperate species (Harrison et al., 2014) and most tropical ones (Jellyman, 2022), although some of the latter are classified as Data Deficient. This leads to a paradoxical situation where one of the most widespread fish genera in the globe, and apparently among the most evolutionary successful due to a range of biological adaptations, is currently facing a practically worldwide conservation threat.
The status of the European eel stock is annually assessed by ICES based on trends in composite indices of glass eel (from 1960 onwards) and yellow eel relative abundance (from 1950 onwards). These indices emerge from the joint statistical modelling of relative abundance indicators available across the species distribution area, retained in the analysis following quality checks and, in the case of the recruitment index, geographically organized as North Sea or elsewhere Europe (ICES, 2022). Currently retained indicators are based on a few fishery-independent monitoring series, frequently associated with obstacles to migration (dams or cooling stations—e.g., Briand et al., 2003; Westerberg and Wickström, 2016) at some distance from the coast, and several fisheries series based on commercial catch or catch per unit effort (CPUE) data (Bornarel et al., 2018). Glass and yellow eel data are often reported jointly in mixed indicators, with 20 mixed series and 37 glass eel series currently used for the assessment of recruitment (ICES, 2022). Data are mainly available in and around the North Sea, with 18 series out of the 22 currently used in the yellow eel index coming from the North Sea (ICES, 2022). The glass eel recruitment index outside the North Sea covers all the remaining range of the species, with a latitudinal (more series in northern than in southern Europe, no series in northern Africa) and a longitudinal (more series in the northeast Atlantic than in the Mediterranean Sea) imbalance. A review of historical trends in eel fisheries from Mediterranean Sea lagoons, also points to important declines in recruitment and local standing stocks (Aalto et al., 2016), although these data are not used by ICES in the assessment indices mentioned above.
The international River Minho/Miño (Minho thereafter) forms the border between northern Portugal and Spain and flows into the Atlantic ocean. Situated at the western edge of the Iberian Peninsula, its river plume (Fernández-Nóvoa et al., 2017) is associated with a narrow continental shelf (Varela et al., 2005). Sediments are exported from the estuary during episodic flood events (Dias et al., 2002) that also contribute to the temporary increase of the turbid plume of the River Minho over a coastal area that can exceed 400 km2 (Fernández-Nóvoa et al., 2017). Complex coastal interactions with the Douro plume and the Rias Baixas circulation are modulated by wind regimes (Mendes et al., 2016) that also influence oceanographic conditions and productivity in the North Iberian Shelf (González-Nuevo and Nogueira, 2014). As a result, the freshwater discharge from the River Minho provides one of the westernmost attractions for continental entry of eastward migrating oceanic eel larvae that initiate their metamorphosis into glass eels (Antunes & Tesch, 1997).
The river Minho has experienced many centuries of diadromous fish exploitation (Araújo et al., 2016). Since the 1970s the most important activity in the estuary has been glass eel fishing using stow nets (Weber, 1986). Since 1981 the fishery is managed jointly by Portugal and Spain (Stratoudakis et al., 2023). An advisory committee, composed of members of the national and regional administration from the two countries, supports the two maritime authorities in the preparation of an annual decree and contributes to communication with national and international scientific bodies and the European Fisheries Administration. A formal meeting of the advisory committee takes place annually in the presence of the representatives of local authorities, where fishery regulations for the coming year are presented and discussed (Stratoudakis et al., 2023). In recent years, the restrictions on glass eel fishing effort have been prominent in the agenda, creating heated debates within the committee and with representatives of local communities and fishers´ associations. The annual catch data reported from this fishery contribute towards the southernmost Atlantic indicator of eel recruitment in the ICES glass eel index outside the North Sea (Bornarel et al., 2018). A local CPUE series has been recently reconstituted, based on total catch adjusted for underreporting and divided by potential fishing effort (Correia et al., 2018). The latter corresponds to fishing opportunities estimated from the number of emitted licences times the number of new moons of possible fishing in the respective glass eel fishing season. The CPUE series showed a moderate decline in the 1980s and some recovery from the 2000s onwards, raising hypotheses of latitudinal differences in population decline within the species distribution area (Correia et al., 2018).
The objectives of this study were to investigate trends in local eel monitoring data and compare with already published series. For that, we recovered and jointly analysed historical and recent glass eel recruitment data that are assumption-free and independent of the current ICES assessment series and the recent reconstitution of the Minho CPUE by Correia et al. (2018). Daily catch records of individual fishers were also used to validate the new series and Generalised Additive Models (GAMs) were used to detect seasonal and inter-annual patterns. Considering what is known about the implementation of the European Eel Management Plans (EMP), emanating from EC Regulation nº 1100/2007 that mainly focused on the restriction of fishing effort, and recent roadmap adjustments proposed to address observed insufficiencies (ICES, 2021), the present study aims to update information on the southern limit of the species distribution in the Atlantic and contribute towards a broader management perspective that simultaneously addresses eel stressors in continental and marine waters.
Materials and methods
Study area
River Minho starts in the highlands of northeast Galicia and ends, after approximately 300 km, in the Atlantic Ocean (41º 51′ 54.39’’ North; 8º 52′ 17.70’’ West), between Caminha (Portugal) and A Guarda (Spain). The river drains a basin of 17.11 × 103 km2, 95% of which is in Spain, and the final part of its course, with a length of 70 km, delimits the northern border between the two countries (international River Minho—Fig. 1). The estuary is mesotidal with a total area of 23 km2, extending about 40 km inland and having a maximum width of 2 km near the mouth. Five dams, constructed between the 1950s and the 1970s, significantly reduced the longitudinal connectivity for diadromous fish in the international main stem and some tributaries (Azeiteiro et al., 2021; Almeida et al., 2023). Damming has also led to increased siltation and reduced water flow in the summer months (Sousa et al., 2008), leading to complaints by fishers and their associations (Stratoudakis et al., 2023). The river has limited industrial and urban pollution and acceptable water quality (Santos et al., 2013), but in recent years there has been an increase in exotic species and a modification of aquatic communities (Ilarri et al., 2022).

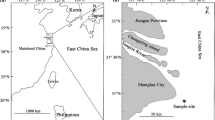
Iberian Peninsula and international River Minho/Miño along the northern border between Portugal and Spain. Rivers along the southern distribution area of the European eel in the northeast Atlantic, indicated in Table 3, are also highlighted
Glass eel fishery
The stow net used in the glass eel fishery in the River Minho was crafted by a local Spanish fisher in the 1960s and rapidly gained prominence due to its efficiency during flood tides (Antunes, personal communication), particularly at nights around new moon (Weber, 1986). Local licensing and systematic recording of total annual catch of glass eel in the River Minho started in the 1973/74 fishing season. The glass eel fishery in Portugal was regulated in 1981, with stow net commercial fishing only allowed at the international section of River Minho. In the remaining Portuguese rivers glass eel fishing was initially allowed with a hand-net, but it became prohibited during 2000, when signs of declining abundance were associated with a generalized use of illegal fishing gear and high levels of bycatch (Antunes and Weber, 1996; Neves et al., 2018). In River Minho, the observed decline in recruitment and the implementation of the local EMP led to a prohibition of fishing for yellow and silver eel during the 2008/09 fishing season and to a progressively tighter regulation of glass eel fishing opportunities (Correia et al., 2018). The number of fishing licences has been steadily decreasing and the total duration of the fishing season has been reduced. During the early years of the activity the fishing season was restricted from November to April to avoid the increasing level of bycatch in the remaining months (Antunes & Weber, 1996). Although there has always been a preference for fishing around the new moon, in more recent years there has been a restriction of fishing to one week before and one week after new moon per month (Correia et al., 2018) and a further reduction in fishing months.
Data availability
Historical data series of experimental stow net fishing, conducted opportunistically in the lower Minho estuary since 1981, were recovered and added to the monitoring data regularly obtained since 2017 following the same sampling protocol. This compilation only considers experimental glass eel fishing events with a stow net at a known fishing location in the lower estuary in which both fishing time duration and glass eel catch records (grams) are available: station 3 in Weber (1986), 5.2 km from the river mouth, and station 1 in Antunes (1990), 2 km from the river mouth. Experimental observations started in the early 1980s (1981/82 fishing season) and have continued opportunistically until 2017, when a regular monitoring program, of monthly sampling from November to May, was initiated under the EU Data Collection Framework. All data correspond to glass eel biomass caught per hour of fishing with a stow net of standard dimensions (20 m long float lines, 15 m long bottom-anchored lead line, 8 m high and 1–2 mm mesh size), with the fishing operation always taking place around the new moon and for 2–3 h after low tide. Some of the earliest data are already published, but the majority are either opportunistic collections within projects or, since 2017, are annually reported to ICES through Eel Data Calls. The new series makes no estimation assumptions, as it is based on standardised glass eel yield per hour of stow net operation during flood tide around the new moon, and closely corresponds to new arrivals, as it takes place near the river mouth and is mainly composed of glass eels at early stages of development (Weber, 1986; Antunes, 1994). Table 1 summarises all standardised experimental glass eel fishing data made available for this study in the international River Minho for the period 1981–2022. These data can provide an independent indicator of catchment recruitment monitoring (according to the definition of Drouineau et al., 2016) for the River Minho and be considered in the future as an additional monitoring series for the composite index outside the North Sea.
The most recent ICES assessment report (ICES, 2022) was used to obtain the series of recruitment indices for the northeast Atlantic and the North Sea and the official Portuguese catch data from the Minho. Table 2 summarizes the information available from daily records of glass eel catch from commercial fishing operations in the River Minho, either through the voluntary disclosure of fishers´ logbooks after retirement or through the anonymous register of logbooks by local traders. Given that commercial fishing operations in the River Minho have always taken place in autumn and winter (Weber, 1986; Antunes and Weber, 1996), calendar months and years are reorganized for this analysis according to the seasonality of glass eel recruitment and the respective fishing season: from October of one year (month 1 in the season of year 1) to September of the next (month 12 of year 1). For example, the last month of the fishing season 2022/2023 in March 2023, corresponds to month 5 in the fishing year of 2022. For this reason, the recruitment indices of ICES are analysed and presented for the year prior to the one reported.
To explore the relationship between seasonality of glass eel arrivals in estuaries at the southernmost range of the species distribution (from southern France to northern Morocco in the northeast Atlantic—see Fig. 1) and river discharge, the results of the present analysis in the River Minho and metadata from other publications were used. Mean monthly discharge data from the River Minho were obtained from Santos et al. (2021) and associated with the seasonality patterns resulting from the modelling of the experimental yield data. For neighbouring rivers within the southern distribution area of eel, mean monthly discharge values and months of glass eel arrivals were obtained from the literature (see discharge metadata citation in parentheses within Table 3 below).
Statistical analysis
The date of fishing operations was used to estimate the lunar luminosity, using the library lunar in R. This corresponds to the nominal nocturnal luminosity at a given latitude and date and does not account for local meteorological conditions. Temporal patterns in experimental glass eel yield from the lower Minho estuary (Table 1) and daily fisher records (Table 2) were modelled with a Generalised Additive Model (GAM—Wood & Augustin, 2002). All experimental yield data from Table 1 were used to describe the seasonality of glass eel arrivals in the Minho estuary and explore for changes over time. To this end, a smooth effect of the month was considered, as well as the effect of the period (high vs low abundance period, two level factor) and the smooth effect of the year. Only the experimental yield data from the main fishing months were then used together with the fisher catch data from Table 2 to model the interannual yield trend, to compare between the two sets (experimental vs fisher data) and with the ICES indices. For that, only the smooth effect of the year was considered for experimental yield during the fishing season, while the smooth effect of luminosity was also considered for the fisher catch daily data.
Adequacy of the model complexity was evaluated using the Akaike´s Information Criterion (AIC). In the case of the seasonality model a forward stepwise selection procedure was followed and the model with the lowest AIC was selected. A negative binomial error distribution was used with a logarithmic link function, iteratively modifying the theta parameter until optimising the distribution of residuals through visual inspection (Venables and Ripley, 2002). The logarithm of the number of experimental fishing hours per night was used as an offset in the experimental yield data and the number of fishers with records in the daily catch data. Predictions were standardised as average daily yield, assuming three hours of commercial fishing operation on the night with the lowest luminosity in the fisher data model. To evaluate management effectiveness over time, luminosity during fishing nights was compared between temporal phases using the non-parametric Wilcoxon rank sum test (W-score for non-parametric equivalent of the two-sample t test when the normality assumption is relaxed). Further, cross-correlations between landings and fishing licenses series were tested in the periods prior and after the EMP, using Kendall´s rank correlation (tau-score for non-parametric measure of association between random variables, able to capture monotonic dependencies when the normality assumption is relaxed and under no or limited auto-correlation). Statistical and graphical analysis was performed in R 4.2.0 (R Core Team, 2022), also using the packages mgcv and MASS.
Results
Although glass eels entered the Minho estuary all year round, experimental observations were generally skewed towards the months of higher recruitment (Table 1). A smooth seasonal effect, distinct for the early and the later period, explained 73% of the deviance in the data series (n = 105 daily observations, 1981–2022). Until the early 1990s there was a clear separation between the high season, from November to April, and the low season, from May to October. Peak yields declined by almost an order of magnitude after the early 1990s, but there was no significant change in mean yield during the low season (Fig. 2). As a result, although seasonality remained evident in the more recent period, the difference between high and low season became less marked and showed a close link with the discharge climatology. Similar relationships were observed in the seasonality of discharge and glass eel entry in the metadata from neighbouring rivers in the southernmost range of the species distribution in the Atlantic (Table 3, for rivers depicted in Fig. 1). Reported seasonality from other rivers in the region confirmed that estuarine entry at lower latitudes takes place throughout the year, but also showed an earliest peak in estuaries of the western Iberian shelf. This difference was evident both in the northernmost Spanish and French estuaries (Bay of Biscay) and in the southernmost Spanish and Moroccan estuaries (Gulf of Cadiz). Further, the climatology of river discharge also seemed to be in line with the above geographical patterns. In most rivers for which mean monthly data were available, the four months of highest discharge coincided with the period of highest glass eel recruitment (Table 3). The highest mean monthly freshwater discharge was reported earlier in rivers whose estuaries are located in the western edge of the Iberian Peninsula, compared to rivers further south or further north.

Left panels: Boxplot comparisons of observed glass eel yield during experimental fishing in the early (up to mid-1990s, green) and later periods (in blue), during both the high season (a) and the low season (c). Note the difference in scale of the y-axis between the two seasons. Right panels: Comparisons of modelled monthly seasonality between the early (lines and circles, in green) and the later periods (solid line, in blue), in absolute (b) and relative (d) yield values. In the latter, the dashed lines with arrows indicate the relative mean monthly freshwater discharge in Minho River
Beyond the drastic reduction in glass eel yield in the Minho estuary since the mid-1990s and the corresponding impacts on seasonality, the addition of a smooth effect of time (year, as determined by the start of the fishing season) to the model of the experimental yield data contributed a small but significant increase in the explained deviance (79%). The predicted value for the 2022/23 fishing season was 6.6% of the predicted value during the onset of sampling four decades earlier. This term inclusion confirmed that the most significant part of the reduction in yield took place up to the early 1990s, when mean yield had already reduced to 20% of its initial value, and that a smoother decline possibly continued to the present days. On the contrary, moon luminosity did not significantly improve the model fit, due to the predominance of observations around the new moon. A very similar smooth effect of time (fishing year) was obtained when the model only considered experimental yield data from the months important to fishing (November to April, 1981–2022, n = 74), explaining 62% of the total deviance. Changes in fishing regulations, over the period analysed, led to a progressive reduction in fishing opportunities and some reorganization of fishing activity. This can be seen in the distribution of moon luminosity during reported fishing nights across three temporal phases from 1990 onwards (Fig. 3). Although there were no significant differences in the overall distribution (recent vs. intermediate W = 61,848, p = 0.070; recent vs. early W = 63,780, p = 0.444), fishing activity in recent years (Fig. 3c) was never registered during the full moon half-cycle, in contrast to earlier periods.

Histogram of lunar luminosity during fishing nights in the early (a, 1990–1998), intermediate (b, 1998–2014) and recent (c, 2017–2023) period
Daily fisher records (n = 1101 individual fishing events, 1990–2022) also showed that the daily catches were significantly higher in the first half of the 1990s than in subsequent periods (Fig. 4). Further, when fishers with known activity in the lower estuary were compared, there was a significant increase in the most recent years (Fig. 4b), although with catches still much lower than in the early reporting period in the 1990s (Fig. 4c). In recent years daily catch was significantly higher in the lower than in the upper estuary (W = 26,676, p = 0.016). Comparing the predicted yield, from the model fitted to the experimental glass eel catch per hour fished, with the corresponding yield predicted from the reported fisher daily catch (assuming a daily effort of three hours), shows that the two independent sources of evidence are nearly overlapping and point to very similar temporal trends (Fig. 4d). Extending the comparison of the fitted yield with ICES recruitment indices and reported Minho catch and potential fishing effort (Fig. 5) also shows that the temporal trend in the experimental yield from the lower Minho estuary is congruent with the ICES elsewhere Europe recruitment index (Fig. 5b) and the total Portuguese landings in the River Minho (Fig. 5c). On the other hand, it shows a delay of a few years in the decline, when compared to the North Sea recruitment index (Fig. 5a), and precedes, by a few years, the decline in the number of fishing licences in the Portuguese River Minho (Fig. 5d). Although the highest cross-correlation of the predicted yield in the monitoring series with all remaining series was without any lag, there was an interesting relationship between the official landings and licences (Fig. 6). In the latter case, the cross-correlation was maximal with a lag-phase of two years for the period 1973–1995 (corresponding to the phase from the onset of licensing to the end of high abundance), whereas no lag was found in the period 2008–2022 (phase since the implementation of the EMP). Although the above relationship may be inflated by the high autocorrelation in the licence series of the early period, the change in pattern between the two periods remains valid.

Top: Boxplots of distribution of daily catch (glass eel biomass per fisher per night) in different time periods in the River Minho—a unknown fishing area and b in the lower estuary. Bottom: Fitted temporal trend (thick blue line) to fishers´ catch records, assuming catch during the new moon night (c) and comparison of fitted experimental yield (dotted line) with estimated yield from fitted catch records (blue line), assuming a daily fishing effort of three hours (d). Data from daily fisher records, as reported in Table 2, and experimental yield records, as reported in Table 1, are superposed on the respective graphs as open circles (highest values are truncated in the y-axis to facilitate visual comparison of the fitted models)

Comparison of temporal trends in experimental yield of glass eel in the River Minho (1981–2022) with: a ICES recruitment index for the North Sea (1960–2022); b ICES recruitment index for Elsewhere Europe (1960–2022); c Glass eel catch in the Minho commercial fishery (Portuguese landings, 1973–2022); and d Glass eel fishing effort in the Minho (Portuguese fishing licences, 1973–2022). Experimental yield values correspond to predicted GAM values (within one standard error envelope). All series under comparison are scaled to the secondary y-axis

Cross-correlation coefficients for total glass eel landings and the respective number of fishing licences in the Portuguese side of the international River Minho during two periods: a during the first two decades of registered fishing (early period, 1973–1995); b from the implementation of the EMP onwards (recent period, 2008–2022). The dashed line (in blue) indicates the limit for statistical significance of the comparisons
Discussion
The recovery and compilation of experimental glass eel fishing data in the lower estuary of River Minho presented here provide a new source of fishery-independent information to monitor river catchment recruitment (following the definition of Drouineau et al., 2016) in the southernmost part of the European eel range, in the northeast Atlantic. Existing recruitment series in that region cover relatively narrow time periods (Arribas et al., 2012; Aranburu et al., 2016; Stratoudakis et al., 2018), combine observations along the estuary and upstream waters (Lobón-Cerviá and Iglesias, 2008; Aranburu et al., 2016) or involve fishery-dependent estimates (Bru et al., 2009; Aranburu et al., 2016; Stratoudakis et al., 2018). The recovery of data from the first experimental monitoring in the Minho estuary (Weber, 1986; Antunes, 1994; Antunes and Weber, 1996) and the replication of these sampling methods in an ongoing monitoring program, in place since 2017, allowed the estimation of temporal trends over the last 40 years. Although the recruitment monitoring in the River Minho has been sporadic and opportunistic (Iglesias et al., 2010), the experimental protocol, including fishing location, practice and gear used (Weber, 1986), has not changed over time, providing compatible observations that are amenable to joint modelling and interpolation. A further strength of the new series is that observations are independent of commercial fishing and possible contaminations from underreporting or variations in fishing effort (Correia et al., 2018). Finally, as this study focuses exclusively on the lower estuary, close to the river mouth, it maximizes the probability of sampling new arrivals (Weber, 1986; Antunes, 1994), prior to experiencing fishing mortality, while avoiding later pigmentation stages and elvers that may be found further upstream (Gascuel et al., 1995; O’leary et al., 2022; Monteiro et al., 2023).
The GAM-based temporal trend in the experimental yield series suggested a large decline of glass eel in the lower Minho estuary since the onset of monitoring in 1981. Most of the observed decline took place during the 1980s. This pattern, but also the absolute values of yield, were corroborated by the estimates derived from the daily fishers´ records assuming three hours of fishing operation per night. The fact that the two independent sources provided congruent yield levels and temporal trends (Fig. 4d), under minimal (in the case of commercial data) or no assumptions (in the case of experimental yield), strengthens the reliability of the new monitoring series. In addition, this trend was in line with the ICES index for elsewhere Europe (Fig. 5b), which includes landings in the Minho, despite the vast geographic area that the latter represents (from the northern Bay of Biscay to the western Mediterranean Sea—Bornarel et al., 2018). Similarly, it was in line with the reduction in leptocephali distribution area in the NE Atlantic (Westerberg et al., 2018) from the 1960s onwards. The new series trend also matched the trend in total landings in River Minho (Fig. 5c). This match is indicative of systems where local effort control does not influence population abundance (Kaifu et al., 2021). The results from this study indicate that although the potential fishing effort is currently below 5% of its maximum values and the glass eel fishery has been banned in all other Portuguese rivers for more than 20 years, there are no signs of recovery in the experimental yield of new glass eel arrivals in the Minho estuary, apart from the possible stabilization of a low catchment recruitment regime.
The trend in the new series diverged from the temporal pattern in the North Sea glass eel recruitment index of ICES, the latter showing an earlier and more pronounced decline, currently at 2–3% of starting values (Fig. 5a). This divergence possibly corresponds to the earlier detection of declining recruitment in the peripheral areas of the oceanic migration. The new series from Minho also diverged from the CPUE series estimated by Correia et al. (2018), who used total catch and estimated fishing opportunities for the Portuguese fishing fleet since 1974. In the CPUE series the decline of glass eel in the Minho estuary did not exhibit a very marked decline, never dropping below 20% of the 1974–1983 baseline and even showing signs of recovery in the most recent decade. This discrepancy may result from disproportional overestimation of fishing effort and underreporting by Correia et al. (2018) over the study period. Potential fishing opportunities assume all licences are active and fully operational along the fishing period. The lag of a few years in the decline of licences in relation to yield (Fig. 5d) and the uneven seasonal distribution of activity in the fishers´ records indicate that observed fishing effort has been lower than the potential and that the discrepancy may be greater in the period of higher abundance. This is further supported by the comparison of cross-correlations between catch and number of licences for the periods up to 1995 and from 2008 onwards. In the initial period of the fishery, the highest correlation found between catch and licences series was with a lag of two years, whereas after the implementation of the EMP, no lag was observed (Fig. 6). Similarly, it is very likely that underreporting of catch has decreased considerably in recent decades, rendering a fixed correction factor for the whole series unrealistic. Finally, the new series also revealed a decline that is more pronounced than the one reported by Stratoudakis et al. (2018) from the River Lis, central Portugal. In the latter, there was a significant reduction from 1989 to 1996, not as sharp as the one reported here, but, when only considering fisheries-independent data, there were no significant differences between the late 1990s and the 2010s. A likely explanation for the latter, in the light of the present series, is that the decline in recruitment to the River Lis (approximately 250 km southwards of Minho) also occurred mainly in the 1980s.
Fisheries-independent sampling in the lower estuary also provided an opportunity to describe patterns in seasonality of entry more reliably than in monitoring systems that aggregate inland observations or depend on the fishing season. It is known that the recruitment season for glass eel in the southern range of the Atlantic distribution is wider (Jorge and Sobral, 1989; Domingos, 1992; Arribas et al., 2012; Stratoudakis et al., 2018; ICES, 2020) and takes place earlier (Tesch, 2003; ICES, 2020) than in the northern part of the distribution range. However, this study also showed that the large reduction in recruitment in this catchment was disproportionately distributed over the year, with glass eels continuing to enter to the Minho estuary all year round and currently showing a close relationship with the monthly seasonality of river discharge climatology (Fig. 2). The latter is possibly a more general feature, at least in the southern part of the species´ Atlantic distribution region, that deserves closer attention. If river discharge mainly drives the seasonality of recruitment in catchments under low population abundance conditions, then the impact of flow regulation and further changes in the hydrological cycle due to climate change (Guerreiro et al., 2017) will interfere with the geographical repartition of glass eel recruitment.
Finally, the trend of declining recruitment in the River Minho reported in this study occurs in an Iberian background of progressive loss of continental habitat for diadromous species along centuries (Clavero & Hermoso, 2015), especially after large-scale dam construction in the twentieth century. The impact of the latter is most evident in anadromous species (Almeida et al., 2023), where orders of magnitude reduction in local abundance were observed during the decades of intense dam construction, particularly from the 1950s to the 1970s. However, regulation of water flow and retention of large water volumes in dams (Syvitski et al., 2005) affects the seasonality and oceanic extension of turbid plumes (Fernández-Nóvoa et al., 2017). This possibly modifies the dynamics of glass eel attraction to estuaries (Stratoudakis et al., 2018) that is highly influenced by continental odours such as geosmine (Tosi & Sola, 1993). In addition to the reduction in longitudinal continuity, the same period has been faced with increased degradation of coastal and continental habitats relevant for eel growth (Costa et al., 2008). Currently, there is no eel restocking programme in Portugal and in general no mitigation of stressors such as habitat loss and degradation. In recent years, translocations of 12–15 cm eels, which accumulate in the first dam (Frieira) of the River Minho, have been carried to tributaries of the international basin. More effective local actions, such as exemptions to maintain a regulated glass eel fishery that is heavily controlled due to the reality of illegal fishing in the country, and the banning of yellow and silver eel fishing to promote spawner escapement, show the difficulty of a concerted national management strategy. This becomes more difficult when other stakeholders with conflicting interests are involved, such as conservationists, water managers and the hydropower industry (Verhelst et al., 2021; Waldman and Quinn, 2022). The historical decline of the stock shows that uncoordinated actions by local managers alone will not sustain the stock (Feunteun, 2002; Dekker, 2016; Kaifu et al., 2021). The reality of the status of the eel stock is more likely to lead to total prohibition measures than to measures aimed at tackling illegal fishing, such as the opening of controlled glass eel fisheries. However, management measures that focus only on restricting fishing may intensify illegal fishing and have an effect contrary to the original objective, while not addressing the remaining drivers of the observed decline (Verhelst et al., 2021; Waldman & Quinn, 2022).
Conclusion
The new monitoring series in the lower estuary of River Minho supports the consistent decline of continental eel catchment recruitment across the species distribution range and shows that two decades of more drastic fishery measures have at best stabilized the species to a low recruitment regime. The inefficiency of fisheries regulatory measures to guarantee alone stock recovery call for more comprehensive measures to reverse anthropogenic impacts on continental eel populations. Such measures will need to be globally relevant and locally comprehensible in order to be effective. In particular, it will be necessary to develop a broader perspective that also takes into account the discharge dynamics in continental basins and the progressive degradation of eel growth habitats in the remaining distribution areas up to the first insurmountable barriers upstream. Observed and predicted declines in river discharge intensity and seasonality resulting from renewable energy requirements and changes in the hydrological cycles is a stress factor towards glass eel catchment recruitment that has not been adequately addressed. Further, habitat loss that results from the above energy needs is aggravated by flood prevention and control of saline intrusion works in the lowlands, contributing to ever decreasing habitat quality as well as quantity. Unless such stressors also become addressed and trajectories are reverted, drastic reduction or even elimination of fishing mortality seems unlikely to revert the European eel to its abundance levels 40 years ago.
Data availability
The datasets generated and analysed during the current study are available from the last author (CA) on reasonable request. The new series will also be made available to the ICES Working Group on Eel WGEEL.
References
Aalto, E., F. Capoccioni, J. T. Mas, M. Schiavina, C. Leone, G. D. Leo & E. Ciccotti, 2016. Quantifying 60 years of declining European eel (Anguilla anguilla L., 1758) fishery yields in Mediterranean coastal lagoons. ICES Journal of Marine Science 73: 101–110. https://doi.org/10.1093/icesjms/fsv084.
Almeida, P. R., C. S. Mateus, C. M. Alexandre, S. Pedro, J. Boavida-Portugal, A. F. Belo, et al., 2023. The decline of the ecosystem services generated by anadromous fish in the Iberian Peninsula. Hydrobiologia 850: 2927–2961. https://doi.org/10.1007/s10750-023-05179-6.
Antunes, C., 1994. The seasonal occurrence of glass eels Anguilla anguilla (L.) in the Rio Minho between 1991 and 1993 (North of Portugal). Internationale Revue Der Gesamten Hydrobiologie 79: 287–294.
Antunes, C. & M. Weber, 1990. Abundance and distribution of eels (Anguilla anguilla) in the Rio Minho. Internationale Revue Der Gesamten Hydrobiologie 75: 795.
Antunes, C. & M. Weber, 1996. The glass ell fishery and the bycatch in the rio Minho after one decade (1981–1982 and 1991–1992). Archives of Polish Fisheries 4: 131–139.
Antunes, C. & F.-W. Tesch, 1997. Eel larvae (Anguilla anguilla L.) caught by R.V. “Heincke” at the European continental slope in autumn 1991. Ecology of Freshwater Fish 6: 50–52.
Aranburu, A., E. Díaz & C. Briand, 2016. Glass eel recruitment and exploitation in a south European estuary (Oria, Bay of Biscay). ICES Journal of Marine Science 73: 111–121. https://doi.org/10.1093/icesjms/fsv116.
Araújo, M.J., S. Silva, Y. Stratoudakis, M. Gonçalves, R. Lopez, M. Carneiro, et al., 2016. Sea lamprey fisheries in the Iberian Peninsula. In “Jawless fishes of the World” Volume 2, Chapter: 20, Publisher: Cambridge Scholars Publishing, Editors: A. M. Orlov, Richard Beamish, pp.115–148.
Arribas, C., C. Fernández-Delgado, F. J. Oliva-Paterna & P. Drake, 2012. Oceanic and local environmental conditions as forcing mechanisms of the glass eel recruitment to the southernmost European estuary. Estuarine Coastal and Shelf Science 107: 46–57. https://doi.org/10.1016/j.ecss.2012.04.024.
Azeiteiro, U. M., M. U. Pereira, A. M. V. M. Soares, H. O. Braga, F. Morgado, M.C. Sousa MC, et al., 2021. Dynamics of two anadromous species in a dam intersected river: analysis of two 100-years datasets. Fishes 6: 21. https://doi.org/10.3390/fishes6020021.
Bornarel, V., P. Lambert, C. Briand, C. Antunes, C. Belpaire, E. Ciccotti, et al., 2018. Modelling the recruitment of European eel (Anguilla anguilla) throughout its European range. ICES Journal of Marine Science 75:541–552. https://doi.org/10.1093/icesjms/fsx180
Briand, C., D. Fatin, G. Fontenelle & E. Feunteun, 2003. Estuarine and fluvial recruitment of the European glass eel, Anguilla anguilla, in an exploited Atlantic estuary. Fisheries Management and Ecology 10: 377–384.
Bru, N., P. Prouzet & M. Lejeune, 2009. Daily and seasonal estimates of the recruitment and biomass of glass eels runs (Anguilla anguilla) and exploitation rates in the Adour open estuary (Southwestern France). Aquatic Living Resources 22: 509–523. https://doi.org/10.1051/alr/2009050.
Clavero, M. & V. Hermoso, 2015. Historical data to plan the recovery of the European eel. Journal of Applied Ecology 52: 960–968. https://doi.org/10.1111/1365-2664.12446.
Correia, M. J., J. L. Costa, C. Antunes, G. De Leo & I. Domingos, 2018. The decline in recruitment of the European eel: new insights from a 40-year-long time-series in the Minho estuary (Portugal). ICES Journal of Marine Science 75: 1975–1983. https://doi.org/10.1093/icesjms/fsy073.
Costa, J. L., I. Domingos, C. A. Assis, P. R. Almeida, F. Moreira, E. Feunteun & M. J. Costa, 2008. Comparative ecology of the European eel, Anguilla anguilla (L., 1758), along a large Iberian River. Environmental Biology of Fishes 81: 421–434. https://doi.org/10.1007/s10641-007-9229-2.
Dias, J. M. A., J. M. Jouanneau, R. Gonzalez, M. F. Araújo, T. Drago, C. Garcia, et al., 2002. Present day sedimentary processes on the northern Iberian shelf. Progress in Oceanography 52: 249–259.
Dekker, W., 2016. Management of the eel is slipping through our hands! Distribute control and orchestrate national protection. ICES Journal of Marine Science 73: 2442–2452. https://doi.org/10.1093/icesjms/fsw094.
Domingos, I., 1992. Fluctuations of glass eel migration on the Mondego estuary (Portugal) in 1988 and 1989. Irish Fisheries Investigations Series A (freshwater) 36: 1–4.
Drouineau, H., C. Briand, P. Lambert & L. Beaulaton, 2016. GEREM (glass eel recruitment estimation model): a model to estimate glass eel recruitment at different spatial scales. Fisheries Research 174: 68–80. https://doi.org/10.1016/j.fishres.2015.09.003.
Drouineau, H., C. Durif, M. Castonguay, M. Mateo, E. Rochard, G. Verreault, et al., 2018. Freshwater eels: A symbol of the effects of global change. Fish and Fisheries 19: 903–930. https://doi.org/10.1111/faf.12300.
Feunteun, E., 2002. Management and restoration of European eel population (Anguilla anguilla): an impossible bargain. Ecological Engineering 18: 575–591. https://doi.org/10.1016/S0925-8574(02)00021-6.
Fernández-Nóvoa, D., M. DeCastro, M. Des, X. Costoya, R. Mendes & M. Gómez-Gesteira, 2017. Characterization of Iberian turbid plumes by means of synoptic patterns obtained through MODIS imagery. Journal of Sea Research 126: 12–25. https://doi.org/10.1016/j.seares.2017.06.013.
Fernández-Nóvoa, D., X. Costoya, M. deCastro & M. Gómes-Gesteira, 2019. Dynamic characterization of the main Cantabrian river plumes by means of MODIS. Continental Shelf Research 183: 14–27. https://doi.org/10.1016/j.csr.2019.06.005.
Fonseca, A., C. Botelho, R. A. R. Boaventura & V. J. P. Vilar, 2014. Integrated hydrological and water quality model for river management: A case study on Lena River. Science of the Total Environment 485–486: 474–489. https://doi.org/10.1016/j.scitotenv.2014.03.111.
Gascuel, D., E. Feunteun & G. Fontenelle, 1995. Seasonal dynamics of estuarine migration in glass eels (Anguilla anguilla). Aquatic Living Resources 8: 122–133.
González-Nuevo, G. & E. Nogueira, 2014. Temporal and spatial variability of river plumes in the NW and N Iberian shelf (1987–2007). Continental Shelf Research 91: 95–108. https://doi.org/10.1016/j.csr.2014.09.005.
Guerreiro, S. B., S. Birkinshaw, C. Kilsby, H. J. Fowler & E. Lewis, 2017. Dry getting drier – The future of transnational rivers basins in Iberia. Journal of Hydrology: Regional Studies 12: 238–252. https://doi.org/10.1016/j.ejrh.2017.05.009.
Harrison, A. J., A. M. Walker, A. C. Pinder, C. Briand & M. Aprahamian, 2014. A review of glass eel migratory behaviour, sampling techniques and abundance estimates in estuaries: implications for assessing recruitment, local production and exploitation. Reviews in Fish Biology and Fisheries 24: 967–983. https://doi.org/10.1007/s11160-014-9356-8.
ICES, 2020. Workshop on the temporal migration patterns of European eel (WKEELMIGRATION). ICES Scientific Reports. 2:25. 108 pp. https://doi.org/10.17895/ices.pub.5993
ICES, 2021. Workshop on the future of eel advice. ICES Scientific Reports 3:13. 67 pp. https://doi.org/10.17895/ices.pub.5988
ICES, 2022. European eel (Anguilla anguilla) throughout its natural range. ICES advice on fishing opportunities, catch and fishing effort. Ele2733.nea. Published 03 November 2022
Iglesias, T., J. Lobon-Cerviá, S. Costa-Dias & C. Antunes, 2010. Variation of life traits of glass eels of Anguilla anguilla (L.) during the colonization of Rios Nalón and Minho estuaries. Hydrobiologia 651: 213–223. https://doi.org/10.1007/s10750-010-0300-1.
Ilarri, M., A. Souza, E. Dias & C. Antunes, 2022. Influence of climate change and extreme weather events on an estuarine fish community. Science of the Total Environment 827: 154190. https://doi.org/10.1016/j.scitotenv.2022.154190.
Itakura, H., R. Wakiya, M. Gollock & K. Kaifu, 2020. Anguillid eels as a surrogate species for conservation of freshwater biodiversity in Japan. Scientific Reports 10: 8790. https://doi.org/10.1038/s41598-020-65883-4.
Jacoby, D. M. P., J. M. Casselman, V. Crook, M. B. DeLucia, H. Ahn, K. Kaifu, et al., 2015. Synergistic patterns of threat and the challenges facing global anguillid eel conservation. Global Ecology and Conservation 4: 321–333. https://doi.org/10.1016/j.ecco.2015.09.007.
Jellyman, D. J., 2022. An enigma: how can freshwater eels (Anguilla spp.) be such a successful genus yet be universally threatened? Reviews in Fish Biology and Fisheries 32: 701–718. https://doi.org/10.1007/s11160-021-09658-8.
Jorge, I., & M. Sobral, 1989. Contribuição para o conhecimento da pescaria do meixão (Anguilla anguilla L.). Dados preliminares sobre a eficiência das principais artes de pesca e importância das capturas acessórias no estuário do Mondego. Relatórios Científicos e Técnicos INIP No 16.
Kaifu, K., K. Yokouchi, M. J. Miller & I. Washitani, 2021. Management of glass eel fisheries is not a sufficient measure to recover a local Japanese eel population. Marine Policy 134: 104806. https://doi.org/10.1016/j.marpol.2021.104806.
Lobón-Cerviá, J. & T. Iglesias, 2008. Long-term numerical change and regulation in a river stock of European eel Anguilla anguilla. Freshwater Biology 53: 1832–1844. https://doi.org/10.1111/j.1365-2427.2008.02008.x.
Mendes, R., M. C. Sousa & M., deCastro, M. Gómez-Gesteira, & J.M. Dias, 2016. New insights into the Western Iberian Buoyant Plume: Interactions between the Douro and Minho river plumes under winter conditions. Progress in Oceanography 141: 30–43. https://doi.org/10.1016/j.pocean.2015.11.006.
Monteiro, R. M., I. Domingos, P. R. Almeida, J. L. Costa, E. Pereira, A. F. Belo, et al., 2023. Upstream movement of juvenile eels (Anguilla anguilla L.) in a southwestern European river. Environmental Biology of Fishes 106: 1313–1325. https://doi.org/10.1007/s10641-023-01417-x.
Neves, J. B., F. Martinho, M. A. Pardal, 2018. Effect of illegal glass eel (Anguilla anguilla) fishery on estuarine fish stocks: a case study in the Mondego Estuary, Portugal. Marine and Freshwater Research 69: 1692–1703. https://doi.org/10.1071/MF17364
O’leary, C., S. Healy, R. Cruikshanks, K. Kelly & P. Gargan, 2022. Assessment of environmental drivers of European glass eel (Anguilla anguilla) recruitment in transitional waters. Environmental Biology of Fishes 105: 1203–1217. https://doi.org/10.1007/s10641-022-01340-7.
Petus, C., V. Marieu, S. Novoa, G. Chust, N. Bruneau & J. M. Froidefond, 2014. Monitoring spatio-temporal variability of the Adour River turbid plume (Bay of Biscay, France) with MODIS 250-m imagery. Continental Shelf Research 74: 35–49. https://doi.org/10.1016/j.csr.2013.11.011.
Pike, C., V. Crook, & M. Gollock, 2020. Anguilla anguilla. The IUCN Red List of Threatened Species 2020: e.T60344A152845178. https://doi.org/10.2305/IUCN.UK.2020-2.RLTS.T60344A152845178.en.
R Core Team, 2022. R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. URL https://www.R-project.org/
Rato, A. S., C. M. Alexandre, P. R. Almeida, J. L. Costa & B. R. Quintella, 2021. Effects of hydropeaking on the behaviour, fine-scale movements and habitat selection of an Iberian cyprinid fish. River Research Applications 37: 1365–1375. https://doi.org/10.1002/rra.3848.
Sabatié R., & G. Fontenelle, 2007. The eel in Morocco: a resource on the edge? Vie et Milieu – Life and Environment 57:213–221.
Santos, S., V. J. P. Vilar, P. Alves, R. A. R. Boaventura & C. Botelho, 2013. Water quality in Minho/Miño River (Portugal/Spain). Environmental Monitoring and Assessment 185: 3269–3281. https://doi.org/10.1007/s10661-012-2789-4.
Santos, A. I., A. Oliveira, D. Carinhas, J. P. Pinto & M. C. Freitas, 2021. Hydrodynamic and sediment transport patterns in the Minho and Douro estuaries (NW Portugal) based on ADCP monitoring data: Part 2—Statistical interpretation of bottom moored datasets. Coasts 1: 56–72. https://doi.org/10.3390/coasts1010004.
Snoussi, M., S. Haida & S. Imassi, 2002. Effects of the construction of dams on the water and sediment fluxes of the Moulouya and the Sebou Rivers, Morocco. Regional Environmental Change 3: 5–12. https://doi.org/10.1007/s10113-001-0035-7.
Sobrino, I., F. Baldó, D. García-González, J. A. Cuesta, A. Silva-García, C. Fernández-Delgado, et al., 2005. The effect of estuarine fisheries on juvenile fish observed within the Guadalquivir estuary (SW Spain). Fisheries Research 76: 229–242. https://doi.org/10.1016/j.fishres.2005.06.016.
Sousa, R., S. C. Dias, L. Guilhermino & C. Antunes, 2008. Minho river tidal freshwater wetlands: threats to faunal biodiversity. Aquatic Biology 3: 237–250. https://doi.org/10.3354/ab00077.
Stratoudakis, Y., P. B. Oliveira, A. Teles-Machado, J. M. Oliveira, M. J. Correia & C. Antunes, 2018. Glass eel recruitment to the river Lis: Ingress dynamics in relation to oceanographic processes in the western Iberian margin and shelf. Fisheries Oceanography 27: 536–547. https://doi.org/10.1111/fog.12274.
Stratoudakis, Y., C. Antunes, C. Correia, A. F. Belo & P. R. Almeida, 2023. Riverine communities and management systems for anadromous fisheries in the Iberian Peninsula: global strategy, local realities. Reviews in Fish Biology and Fisheries 33: 875–892. https://doi.org/10.1007/s11160-022-09742-7.
Syvitski, J. M. P., C. J. Vorosmarty, A. J. Kettner & P. Green, 2005. Impact of humans on the flux of terrestrial sediment to the global coastal ocean. Science 308: 376–380. https://doi.org/10.1126/science.1109454.
Tesch, F.-W., 2003. The Eel. Biology and management of anguillid eels. 5th edn. Blackwell Science, Oxford. 408 pp.
Tosi, L. & C. Sola, 1993. Role of geosmin, a typical inland water odour, in guiding glass eels Anguilla anguilla (L.) migration. Ethology 95: 177–185.
Varela, R. A., G. Rosón, J. L. Herrera, S. Torres-López & A. Fernández-Romero, 2005. A general view of the hydrographic and dynamical patterns of the Rías Baixas adjacent sea area. Journal of Marine Systems 54: 97–113. https://doi.org/10.1016/j.jmarsys.2004.07.006.
Verhelst, P., J. Reubens, D. Byusse, P. Goethals, J. van Wicehelen, & T. Moens, 2021. Toward a roadmap for diadromous fish conservation: the big five considerations. Frontiers in Ecology and the Environment 19:396–403. https://doi.org/10.1002/fee.2361
Venables, W.N., & B.D. Ripley, 2002. Modern Applied Statistics with S, Fourth edition. Springer, New York. ISBN 0–387–95457–0, https://www.stats.ox.ac.uk/pub/MASS4/
Waldman, J.R., & T.P. Quinn, 2022. North American diadromous fishes: Drivers for decline and potential for recovery in the Anthropocene. Science Advances 8:eabl5486. https://doi.org/10.1126/sciadv.abl5486
Weber, M., 1986. Fishing method and seasonal occurrence of glasseels (Anguilla anguilla, L.) in the Rio Minho, west coast of the Iberian Peninsula. Vie Et Milieu 36: 243–250.
Westerberg, H. & H. Wickström, 2016. Stock assessment of eels in the Baltic: reconciling survey estimates to achieve quantitative analysis. ICES Journal of Marine Science 73: 75–83. https://doi.org/10.1093/icesjms/fsv049.
Westerberg, H., M. J. Miller, K. Wysujack, L. Marohne, M. Freese, J. D. Pohlmann, et al., 2018. Larval abundance across the European eel spawning area: An analysis of recent and historic data. Fish and Fisheries 19: 890–902. https://doi.org/10.1111/faf.12298.
Wood, S. N. & N. Augustin, 2002. GAMs with integrated model selection using penalized regression splines and applications to environmental modelling. Ecological Modelling 157: 157–177. https://doi.org/10.1016/S0304-3800(02)00193-X.
Yahyaoui, A., P. Aguesse & P. Beaubrun, 1983. Etude du recrutement en civelles (Anguilla anguilla L., 1758) dans l’estuaire du Sebou au cours de la saison de pêche 1980–1981. Bulletin de l´Institut Scientifique. Rabat 7: 149–164.
Acknowledgements
We thank Manuela Azevedo (IPMA) for insight and support to establish a national eel monitoring plan in 2017 under the Data Collection Framework in Portugal (EU funding) and Rita Vasconcelos and Carla Gameiro (IPMA) for smooth handling of CIIMAR subcontracting. The contribution of YS is under the project PPCENTRO. Eduardo Martins and José Benito Guerreiro contributed to the fieldwork and Rodrigo López edited Figure 1. Thanks are also due to the fishers and traders that have contributed their personal records. YS and CA acknowledge other CPIRM members and coordinators, from both countries, for the tough but insightful discussions on the management of the local eel fishery.
Funding
Open access funding provided by FCT|FCCN (b-on). This research was supported by FCT—Foundation for Science and Technology through the project UIDB/04292/2020 awarded to MARE and project LA/P/0069/2020 granted to the Associate Laboratory ARNET.
Author information
Authors and Affiliations
Contributions
YS—Conceptualization, Methodology, Investigation, Writing—Original Draft Preparation; ID—Methodology, Writing—Review and Editing; CA—Data curation, Methodology, Writing—Review and Editing, Funding Acquisition.
Corresponding author
Ethics declarations
Conflict of interest
No interest to declare.
Additional information
Handling editor: Zhi Mao
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Stratoudakis, Y., Domingos, I. & Antunes, C. Glass eels at the continental edge of Europe: revisiting catchment recruitment at the international River Minho/Miño. Hydrobiologia 851, 2591–2607 (2024). https://doi.org/10.1007/s10750-023-05459-1
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10750-023-05459-1