
Abstract
In paleolimnological studies, rotifers are usually a very rare component in multiproxy research. These small invertebrates are identified from sediments by their loricas. Analysis of subfossil rotifers was conducted for sediment cores collected from three peatlands in within Poland: the Grel raised bog (S Poland, Carpathians, Orawa-Nowy Targ Basin), the Imszar raised bog (NE Poland, Podlasie Region), and the Podemszczyzna peatland (SE Poland, Sandomierz Basin). Results were compared with multiproxy analyses of peat deposits: pollen of aquatic and mire plants, non-pollen palynomorphs (NPPs), and geochemical analyses. Analysis of the rotifers indicated the occurrence of the bdelloid species Habrotrocha angusticollis (Murray, 1905). In all profiles of the studied peatlands, the occurrence of rotifers was not correlated with pollen of aquatic or mire plants or NPPs; they seem to be more related to geochemical indices. The highest densities of rotifers were found in profile sections without significant fluctuations of geochemical variables, i.e. those with higher Na/K (chemical denudation) ratios and stable redox conditions (Fe/Mn ratio). The study of subfossil rotifers supported by multiproxy analysis of peat deposits could be a useful tool for indication of periods with stable palaeohydrological conditions during the Late Glacial and Holocene.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Peatland deposits preserve unparalleled archives of palaeohydrological, palaeobiological and palaeoenvironmental changes recorded in sedimentary, geochemical and palaeobiological proxies. Paleoecologists commonly use multiproxy analyses to reconstruct precise records of historical and contemporary changes in lakes, watersheds, and peatlands at a local scale, and—ultimately—at global scales. From a paleoecological perspective, most taxa (e.g. pollen, diatoms, amoebas, chironomidae, Cladocera, and Rotifera) have also been readily adopted as valuable indicators of ecological change. Some of these organisms may serve as models in resurrection ecology, but several others offer strong potential because of their prevalence across environmental and temporal scales (eg. Lamentowicz et al., 2010; Chambers et al., 2012; Marcisz et al., 2015). Recent studies show that among the variety of biological proxies, rotifers can provide valuable information on environmental changes (Wojewódka & Hruševar, 2020).
Rotifers are cosmopolitan in distribution, living in a variety of aquatic and semi-aquatic habitats, from the deepest parts of the largest lakes, to the smallest temporary puddles, rock depressions, cemetery urns, and eaves troughs, to damp soil and wet mosses.
So far, in palaeolimnological studies, rotifers are a very rare component in multiproxy research. These small invertebrates are identified from sediments by their loricas and resting eggs, which are particularly conducive to preservation in fossil peat deposits (Frey, 1964; van Geel, 1986; Warner & Chengalath, 1988). Rotifer loricas have been used to reconstruct early postglacial community succession (Swadling et al., 2001) and nutrient dynamics of lakes (Turton & McAndrews, 2006).
The use of Rotifera for palaeoreconstruction is still questionable due to the weak state of preservation of their remains in lake and peatland deposits. They are hardly ever found in lake sediments, except for one core from Lake Crawford (Canada), where numerous fossil rotifer remains were well preserved. Analysis of this core aided the tracing of eutrophication (Turton & McAndrews, 2006; Wojewódka & Hruševar, 2020). On the other hand, Rotifera have a particular role in the functioning of the ecosystem due to their pivotal position in the food chain. They are good indicators of environmental changes because they quickly respond to environmental changes and stressors. Based on the literature, the proportions of the use rotifers as a proxy in studies of palaeoreconstructions and lake restauration is still low (below 2% and 9% respectively) (Wojewódka & Hruševar, 2020).
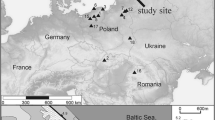
In this work, we combined stratigraphic data from peat compiled on the basis of analyses of pollen and non-pollen palynomorphs (NPPs), thelmatological analyses, Rotifera and geochemical analysis in peatlands, as well as radiocarbon (14C) dates. Analysis of subfossil rotifers was conducted for sediment cores collected from three peatlands in Polish territory: the Grel raised bog (S Poland, Orawa-Nowy Targ Basin), the Imszar raised bog (NE Poland, Podlasie Region), and the Podemszczyzna peatland (SE Poland, Sandomierz Basin) (Fig. 1). The results of the studies of Rotifera were compared with multiproxy analysis of peat deposits including pollen of aquatic and mire plants, non-pollen palynomorphs (NPPs), and geochemical analyses. Multiproxy analysis aimed to document palaeoenvironmental and (hydro) climatic variability in north-eastern Poland since the Allerød Interstadial (Margielewski et al., 2022a, b, c). We examined the impact of climatic changes (mainly climate humidity fluctuation) on the colonisation of the Grel, Imszar and Podemszczyzna peatlands by rotifers.

Location of analysed peatlands, and presentation of study sites on a LiDAR DTM model (A–C)
Materials and methods
Study area
The Grel raised bog is located in the Orawa-Nowy Targ Basin (ONTB) (Inner Carpathians, Southern Poland), in the vicinity of Ludźmierz village (near the town of Nowy Targ). It forms in the highest terrace of the Czarny Dunajec River (ca. 598–601 m a.s.l.) (Fig. 1A). The raised bog (with an original area of 18 ha), exploited for a long time by local communities, has a strongly degraded peat dome (598.4 m a.s.l.). The area of the present-day preserved peat bog reaches ca. 8 ha, while the maximum thickness of the peat deposits is about 3.9 m (Margielewski et al., 2022c).
The Imszar raised bog is located in the Gródek-Michałowo Depression belonging to the mesoregion of the Białystok Heights in the Podlasie region (NE Poland), which is a part of the Central European Plain (Kondracki, 2001). Its area is approx 57 ha (the peat exploitation field covers 36 ha) (Fig. 1B). The thickness of the peat deposits is about 3.3 m (Margielewski et al., 2022a).
The Podemszczyzna peatland is located in the village of Podemszczyzna, in the SE part of the Tarnogród Plateau, part of the Sandomierz Basin, in the vicinity of the Roztocze Upland (Kondracki, 2001). A fen, with an area of 25 ha, was formed in a depression developed within the Świdnica river valley which currently flows in the vicinity (Fig. 1C). The maximum thickness of the peat deposits is 4 m (Margielewski et al., 2022b).
Sampling and laboratory analyses
Coring and sampling
Corings were performed in the central parts of each peatland, using an Instorf sampler, 10 cm in diameter. Complete logs were retrieved, sampled, and subjected to multiproxy analysis (Margielewski et al., 2022a, b, c). In this study, mineral deposits from the bottom parts of the studied logs (sands, silt) were excluded from further analysis due to lack of Rotifera remains.
Sediment analysis
Organic (peat) deposits were subjected to plant tissue analysis by means of a light microscope. On the basis of telmatological analysis, the peat type was determined in accordance with the classification of Tołpa et al. (1971). The degree of peat decomposition (R) was also determined. Loss on ignition (LOI) analysis of deposits was elaborated for each 2.5 cm sequence of log. The samples were subjected to ignition in a muffle furnace at 550 °C according to the procedure proposed by Heiri et al. (2001) (see Fig. 2).

Sedimentary sequences of the studied peatlands, with loss on ignition curve of sediments of each log. On the right of each profile the modelled age axis is presented, elaborated on the basis of an age-depth model elaborated according to Bronk Ramsey (2008)
Chronology
Based on a large number of radiocarbon 14C dates (LSC, AMS), high resolution age-depth models were established for each core of the analysed peatlands (see Margielewski et al., 2022a, b, c). All three age-depth models were elaborated using the P_sequence function of OxCal 4.3 software (Bronk Ramsey, 2006, 2008) based on the IntCal 20 calibration curve (Reimer et al., 2020).
Pollen and non-pollen palynomorphs (NPPs) analyses
Results of pollen and NPPs analyses of each peat sequence have been recently published (Margielewski et al., 2022a, b, c). Samples (of 1 cm3 in volume) collected from a core taken in the Imszar peatland were analysed by M. Kupryjanowicz and M. Fiłoc from the University of Białystok (Poland) (Fig. 3B) (Margielewski et al., 2022a). Samples from the Grel raised bog and Podemszczyzna peatland were elaborated by K. Korzeń (Fig. 3A and C) (Margielewski et al., 2022b, c). Pollen and non-pollen palynomorphs diagrams of each profile were simplified in order to detect possible relations between Rotifera occurrences with the presence of wetland and aquatic plants and selected NPPs such as fungi, algae, and testate amoebae. The variable amount of arboreal pollen (AP) and terrestrial herbs (NAP) was presented as an AP/NAP ratio, according to the standard procedure proposed by Berglund and Ralska-Jasiewiczowa (1986) and Moore et al. (1991) (Fig. 3). Calculations and presentation of pollen and NPPs data were performed using TILIA Graph software (Grimm, 1991).

Simplified pollen and non-pollen palynomorphs (NPPs) diagrams of deposits of the studied peatlands (excluding mineral sediments underlying the peat), with accumulation rate of sediments and Rotifera (Habrotrocha angusticolis) frequency. Pollen diagrams A and C were elaborated by K. Korzeń, diagram B by M. Kupryjanowicz and M. Fiłoc. Rotifera analysis: A. Pociecha. Graphical elaboration of diagrams by K. Buczek
The aim of the palynological analysis was not to reconstruct local vegetation (which was described in previous articles—see Margielewski et al., 2022a, b, c), but to reconstruct (based on pollen and NPPs) paleohydrological conditions that could affect the occurrence of Habrotrocha angusticollis.
Rotifera remains
Analysis of Rotifera remains was performed using sub-samples of the sediment core. Samples (of 1 cm3 in volume) taken in profile were prepared according to the procedure proposed by Frey (1986, 1987). After the removal of carbonates using HCl, each sample was boiled at a temperature of 60 °C in 10% KOH for 30 min. A magnetic stirrer was used for dispersion. After washing with distilled water, the residue was sieved through a 35-μm mesh sieve. The extracted Rotifera remains were stored in 10 ml of water with glycerine and safranine. Taxa were identified and counted at 200 or ×400 magnification under a Nikon 50i microscope. The results of the analyses are presented in a diagram elaborated using the TILIA Graph computer program (Grimm, 1991) (Fig. 3).
Geochemical and statistical analyses
The bottom parts of the profiles consisted mainly of mineral sediments without the occurrence of Rotifera and were excluded from further analyses. Geochemical analyses were prepared according to the procedure proposed by Pansu & Gautheyrou (2006). The content of macroelements (Ca, Mg, Na, K, Fe) and trace elements (Mn, Cu, Zn, Ni, Pb) were measured using Atomic Absorption Spectrometry (AAS). Geochemical analyses of sedimentary sequences from the Imszar and Podemszczyzna peatlands were conducted by D. Sala and have been recently published (Fig. 4B, C) (Margielewski et al. 2022a, b). Analyses of the Grel peat sequence were developed by R.K. Borówka and J. Tomkowiak from the Geochemical Laboratory at the University of Szczecin (Fig. 4A). Due to the lack of measurable concentrations of Cu, Pb and Ni in the Imszar peatland deposits (Margielewski et al. 2022a), neither these variables nor Cu/Zn ratio were included in this study. The geochemical diagram developed in the TILIA Graph computer program (Grimm, 1991) presents both the variability of the content of the major and trace elements in the profile (Ca, Mg, K, Na, Fe, Mn, Zn,), and the geochemical environment indicators Na/K, Ca/Mg, Na_K_Mg/Ca, Fe/Mn, Fe/Ca (Fig. 4). Concentration of elements, content of organic matter, modelled average accumulation rate [mm/year], and values of basic geochemical indicators were compared in intervals with the presence and absence of Rotifera. The significance of differences between selected groups was tested using the non-parametric Mann–Whitney U test (P < 0.05). The correlation between the density of Habrotrocha and geochemical variables was tested using the non-parametric Spearman rank correlation test at a significance level P < 0.05. Statistical analyses were carried out using Statistica 13 software.

Rotifera distribution in the analysed logs versus geochemical analysis of sediments. Geochemistry of the Grel raised bog (A) by K. Borówka and J. Tomkowiak, of the Imszar and Podemszczyzna peatlands (B, C) by D. Sala
Results
Sedimentary sequence
The bottom section of the log taken from the Grel peatland (3.95–3.05 m) is dominated by mineral, fluvial clayey silts with an admixture of sand and a thin layer of strongly decomposed peat in the middle of the section, which was radiocarbon dated to 13,875 ± 45 BP (17,026–16,661 year. cal BP). Fluvial sediments are overlain with an approx. 1.4 m thick horizon of strongly decomposed minerogenic peat, accumulated since ca. 13,807–13,446 cal BP (Fig. 2A). Peat sedentation was interrupted several times by deposition of thin illuvial horizons caused by flooding of the peatland by the Czarny Dunajec River (Fig. 2A). The minerogenic peat is overlain by a 1.7 m thick horizon of ombrogenic peat, whose accumulation began at ca. 9399–9018 cal BP (Margielewski et al., 2022c). The lower part of the ombrogenic peat sequence (below 0.9 m) is strongly decomposed (R > 65%). The upper part of the horizon is composed mainly of a changeable amount of Sphagnum magellanicum Brid. 1798, S. fallax (Klinggr.) Klinggr. 1881, and Eriophorum vaginatum L. 1753.
In the Imszar peatland, organic deposits were accumulated directly on medium-grained sand drilled at the interval 3.4–3.3 m. The beginning of peat accumulation (at the depth of 3.3 m.) was dated by 14C to 12,492 − 11,813 cal BP. In the interval 3.3–2.2 m, minerogenic peat occurs: firstly (in the bottom part) strongly decomposed, whereas in the upper part of the sedimentary sequence, detrital peat was developed: sedge-reed peat, with an admixture of woody peat (Fig. 2B). The upper part of the peat complex (above the depth 2.2 m) was formed by ombrogenic peat: eriophorum peat (with an admixture of pine wood), and (in the last 90 cm of the profile) sphagnum peat (Fig. 2B). On the basis of pollen analysis, at the depth of 205–210 cm, a hiatus (sedimentary gap) was found.
The Podemszczyzna peatland was developed on river sediments formed in the Older Dryas Stadial: in the bottom part of the log, in the interval 4.5–4.0 m, sandy silt (locally with an admixture of organic matter) occurs. The beginning of peat accumulation at a depth of 4.0 m was dated by 14C to ca. 13,583–13,110 cal years BP. In the bottom part (4.0–2.5 m), highly decomposed peat (R > 60%) occurs. In the upper part of the profile (2.5–1.6 m), an insertion of detrital peat occurs: sedge peat with an admixture of common reed [Phragmites australis (Cav.) Trin. ex Steud, 1841] macroremnants. In the interval 1.6–0.9 m, strongly decomposed (R > 65%) sedge peat was accumulated again (Fig. 2C). In the upper part of the peat complex (0.9–0.2 m.), sedge-reed peat (Magnocaricioni) was accumulated. The peat complex is covered by a thin (20 cm) horizon of strongly decomposed peat (R > 65%), with an admixture of a large amount of mineral sediments (Fig. 2C).
Pollen and non-pollen palynomorphs (NPPs) analyses
Pollen analysis indicates that all of the analysed peatlands began to form in the Late Glacial. The beginning of the vegetation history in the Grel raised bog in the light of pollen records was dated at the Oldest Dryas Stadial. In the lower part of the sedimentary sequence (minerogenic stage of mire formation: 300–240 cm) the palynological diagram shows a high proportion of cyperaceae (up to 30%) and bryales (up tu 40%) (Fig. 3A). In the lower parts of the profile, there is also a distinct greater share of herbaceous plants (NAP) (Fig. 3A). From a depth of 110 cm (dated at ca 6.2 ka mod. inch BP), the share of plants typical of raised bogs increases: ericaceae appears, Vaccinium, Sphagnum). There are also numerous NPPs typical for the raised bogs (Entophlyctis lobata Willoughby & Townley 1961, Arcella, Assulina seminulum (Ehrenberg, 1838) Leidy, 1879) (Fig. 3A). In the top parts of the sedimentary sequence (from the depth of 40 cm), the share of ericaceae increases significantly, with the simultaneous disappearance of NPPs (except for E. lobata).
The beginning of the accumulation of organic sediments in the Imszar peatland took place at the beginning of the Alleørd Interstadial. The profile of sediments are characterized by a lower share of herbaceous pollen (NAP) than in the Grel peatland. (Fig. 3A, B). In the lower parts of the sedimentary sequences (300–200 cm) there is the largest share of cyperaceae (up to 10%) (locally accompanied by bryales and Equisetum), typical for a mineogenic fen (Fig. 3B). In the lower parts of the sediments (290–280 cm), there is a local, significant increase in the share of Sphagnum, which disappears almost completely in the interval 270–200 cm (Fig. 3B). The decrease in the share of sedge pollen upwards in the profile is accompanied by a sudden and significant increase in the share of Sphagnum spores (up to 30% in the depth of ca 200 cm), accompanied by taxa characteristic of the raised bog: Vaccinum, Calluna vulgaris (L.) Hull 1808, Ledum (Fig. 3B). Along with the change of accumulation from minerogenic to ombrogenic peat, numerous NPPs also appear in the organic deposits: Tilletia sphagni Navashin (1890), E. lobata, and locally, Arcella and Assulina (Fig. 3B). Characteristic for the Imszar peatland development, is the negligible amount of pollen of aquatic plants, represented only by Lemna and Typha, occurring very locally in the lower part of the sedimentary sequence.
The Podemszczyzna peatland was formed during the Older Dryas Stadial climate cooling, as indicated by palynological analysis. In the pollen profile, the negligible share of herbaceous plants (NAP) is noteworthy: only in the bottom parts of the peat sedimentary sequence, it increases very locally and is accompanied by the spot appearance of ericaceae, Calluna and Vaccinum pollen (Fig. 3C). The share of cyperaceae pollen is small (up to 5%), but quite even in profile. This is characteristic of a minerogenic fen: the lack of Sphagnum spores is also symptomatic here (Fig. 3C). The share of NPPs is very small, locally even trace. The share of aquatic plants is also small, as for a fluviogenic fen (Sparganium, Typha) (Fig. 3C).
Rotifers
Analyses of rotifers indicated one bdelloid species, H. angusticollis, in the Grel, Imszar, and Podemszczyzna peatlands. H. angusticollis is a species typically associated with peat vegetation; it lives among Sphagnum and Polytryhum, and also less commonly in reservoirs and rivers. In the Grel profile, the highest densities of H. angusticollis lorica were found in two layers from 55 (117 ind./cm3) to 65 (52 ind./cm3) cm, and from 95 (46 ind./cm3) to 115 (18 ind./cm3) cm. Single rotifer specimens were found in layers from 275 to 305 cm, which may indicate the reappearance of water in the peatbog zone (Fig. 3A). In the Imszar sedimentary sequence, lorica of H. angusticollis were found in the sediments of the Subatlantic and Subboreal Phases in significant densities. The highest densities of the species were found in the layers at the depths 65 cm–77 ind./cm3; 85 cm–55 ind./cm3, and 130 cm–44 ind./cm3 (Fig. 3B). In the Podemszczyzna profile, lorica of H. angusticollis were found in significant densities in the sediment samples corresponding to the end part of the Preboreal Phase (PB). The highest densities of these species were identified at the depths of 160 cm–41 ind./cm3; 170 cm–35 ind./cm3; 210 cm–15 ind./cm3; and 220 cm–22 ind./cm3 (Fig. 3C).
The development of the rotifer H. angusticollis coincided with the intensive development of two species of microorganisms: a parasitic fungus growing on Sphagnum (E. lobata) and a testate amoeba (A. seminulum) in the Grel profile, as well as two species of parasitic fungi growing on Sphagnum (E. lobata, T. sphagni) in the Imszar profile (Fig. 3A and B). These organisms are strictly related to a wet or submerged Sphagnum vegetation environment.
Geochemistry
Analysis of average values of selected elements and geochemical indicators has shown some statistically significant differences (P < 0.05) between levels with H. angusticollis (“Y”) and those without it (“N”) (Tables 1, 2 and 3). In the Grel raised bog peat, levels with the occurrence of H. angusticollis are characterised by lower concentrations of Mg (Me:0.23 mg/g), a two-fold lower concentration of K (Table 1), and two-fold higher values of the Na/K ratio (Fig. 4A). “Y” horizons in the Imszar peat bog are characterised by significantly lower concentrations of Ca (Me: 3.9 mg/g; P = 0.028), Fe (4.9 mg/g; P = 0.036) and a higher content of Mg (Me: 1.47 mg/g; P = 0.036) and Zn (two-fold higher). The appearance of rotifer communities is correlated with a significantly higher average value of the Na + K + Mg/Ca ratio (associated with a two-fold lower Ca content) and a more oxidizing environment (lower Fe/Mn ratio). The values of Ca, Fe, Ca/Mg, Fe/Mn, Fe/Ca, and accumulation rate [AR] are noticeably less diverse (more stable) in the “Y” group (Table 2) (Fig. 4B). In the Podemszczyzna peatland, the “Y” group is correlated with a higher AR (Me: 0.59 mm/year), greater content of organic matter (Me: 91.1%), lesser concentrations of K (fourfold less compared to the “N” group), Fe, Mn, and Zn. Values of geochemical variables in peat sequences with H. angusticollis are predominantly less dispersed (expressed as a quartile coefficient of dispersion) compared to other peat sections (Tables 1, 2 and 3). In general, the sediment sequence of the Grel peat bog is characterised by higher variability of geochemical variables (CQD varies from 1.4 to 86.7% (Me: 43%) compared to the Imszar site (2.3–45.5%; Me: 25.9%) and the Podemszczyzna peatland (0.9–70.5%; Me: 18.2%). A considerably lower variability of the geochemical proxies was found in the H. angusticollis horizons “Y” of the Grel site for the lithogenic elements Ca, Mg, K, and Na as well as for Ca/Mg, Fe/Mn, and Fe/Ca ratios. Levels with rotifer occurrence in the Imszar raised bog correspond with significantly more stable Ca (CQD: 21.6%) and Fe (CQD: 30%) concentrations, and also Ca/Mg, Na/K, Fe/Mn, and Fe/Ca values. Similarly to the Grel sediment sequence, the “Y” horizons of the Podemszczyzna profile are characterised by less varied concentrations of K and Na (CQD: 14.3 and 14.4% respectively) and more stable values of Ca/Mg, Fe/Mn, and Fe/Ca ratios. CQD values are also significantly lower in the case of Na/K (threefold) and Na + K + Mg/Ca (threefold) ratios (Tables 1, 2 and 3) (Fig. 4C).
The density of H. angusticollis in the Grel peat sequence is highly negatively correlated with the concentration of the lithophilic elements Mg (− 0.64), K (− 0.67), and Fe (− 0.62) and positively with content of organic matter (0.59) and Na/K ratio (0.82) (Table 4). Similarly to the Grel site, the density of the analysed rotifer in the Imszar raised bog was strongly positively correlated with Na/K ratio (0.61) and conversely negatively correlated with Fe/Ca ratio (− 0.56) (Table 4). In the Podemszczyzna peatland, some weak and moderate relationships between H. angusticollis and the analysed variables can be observed: Na (0.49), OM (0.38), Fe/Ca (− 0.37), Fe (− 0.35), and Mn (− 0.34), although at such a small sample size (N = 15) they are not-significant at P < 0.05 (Table 4).
Disscusion
The appearance of H. angusticollis versus the palaeobotanical records
However, only in the Grel peatland were a few individuals of H. angusticollis found in the horizons of sediments deposited in the Late Glacial, corresponding to the end of the Allerød Interstadial and the Younger Dryas Stadial (Fig. 3A) (see Margielewski et al., 2022c). In the other two peatlands, Habrotrocha occur exclusively in the Holocene sequences of sediments. In the palynological profiles of all the analysed peatlands, the significant presence of cyperaceae pollen in the entire profile is noteworthy, especially in the minerotrophic stage of peatland formation (the bottom parts of the Grel and Imszar peatlands and the entire Podemszczyzna peatland profile), as well as in the ombrogenic peats of the Grel and Imszar peatlands) (Fig. 3A–C). The significant share of ericaceae pollen in the ombrogenic peat sequences of the Grel and Imszar peatlands is also characteristic, with a very small share of this pollen taxa in the minerogenic (and at the same time fluviogenic) Podemszczyzna peatland (Fig. 3).
Pollen of water plants indicating the high level of the water table in peat bogs (Nuphar, Stratiotes aloides L. 1753) appears only in the Grel peatland, mainly in its bottom parts (Fig. 3A). At the other sites, water plants are represented mainly by shallow water or reed communities (Typha, Sparganium, Caltha). Among the NPPs, spores of the fungi E. lobata appear an masse within the ombrogenic peats in the Grel and Imszar peatlands (Fig. 3A, B), and their occurrence in the profile of the Podemszczyzna mineogenic peatland is in trace amounts (Fig. 3C). This species is generally associated with humid environments (Kuhry, 1985), less often with shallow water (Shumilovskikh et al., 2015). In the Grel and Podemszczyzna peatlands, a greater share of E. lobata spores occurs in sedimentary sequences where a local decrease in the share of H. angusticollis was recorded (Fig. 3A and C). Similar trends in relation to the decrease in the share of rotifers in the sediments are shown by the local increase in the share of T. sphagni, a species considered to be an indicator of the transition from a dry to wet environment (Shumilovskikh et al., 2015). In addition, in the Grel peatland, the increase in the share of Habrotrocha is well correlated with the local disappearance of the amoeba Archerella flavum (Archer,1877) Loeblich and Tappan, 1961 in the deposits. (Fig. 3A), considered to be an indicator of a high water level in peat bogs (Payne et al., 2012). In turn, in the Imszar, these relationships are less clear: an increase in the share of E. lobata spores accompanies both a decrease in the share of Rotifera and (to a lesser extent) a local decrease in the share of spores of Habrotrocha. The same trend is visible in the share of T. sphagni spores (Fig. 3B).
Habrotrocha angusticollis appearances in connection to climatic and paleohydrological changes since the Late Glacial
In the Podemszczyzna peatland, the first abrupt rise of rotifer density (1–22 ind/cm3) was proceeded by a short-term period of higher water level causing around 9830–9790 cal BP a dying off phase of the local bog pine forest (Margielewski et al., 2022b). High abundances of Habrotrocha between 9700 and 9530 cal BP co-occurred with decreasing content of Mg and Fe, a dropping catchment erosion ratio (Na + K + Mg/Ca) as well as a distinct increase in the Na/K ratio, which points to limited delivery of allochthonous material from the bog’s catchment and/or a less prominent groundwater supply to the bog (Borówka, 1992, 2007). In the Podemszczyzna peat sequence, the first Habrotrocha “crisis” occurred between ca. 9190 and 9030 cal. BP, coinciding with a 9.2 ka worldwide climate anomaly event (Rasmussen et al., 2007; Fleitmann et al., 2008; Lang et al., 2010). A climate cooling dated to ca. 9.3 ka has been recorded in Greenland ice cores (Rasmussen et al., 2007) and in Atlantic sea cores (Bond et al., 1997, 2001, 2008). Overlapped cooler and wetter events were recorded in several sites in Europe. In the Alps, a period of tree line lowering and higher water level in lakes occurred between 9.6 and 9.1 cal. b2k (Haas et al., 1998). Wet and cold shifts were recognised in northern and western England (Barber et al., 2003; Lang et al., 2010). In Asia, a significant weakening of the summer monsoon and general dry conditions have been recorded (Zhang et al., 2018). A short-term period of higher water level (the appearance of Nuphar pollen in the pollen diagram) was also noticed at around 9500 cal BP in the nearby (ca. 50 km) Hamernia raised bog (Margielewski et al., 2015). The density of Habrotrocha had been decreasing gradually since ca. 9530 cal BP, completely disappearing around 9030 (± 96) cal. BP. The rapid reappearance of Habrotrocha after a relatively short period (ca. 170 years) correlates with the rather short-term duration (200–150 year) of the 9.2 ka anomaly around the world (Fleitmann et al., 2008). A “Habrotrocha optimum” following the 9.2 ka event was characterised by the highest rotifer density (35–41 ind/cm3) in the Podemszczyzna peat sequence. This short-lived phase estimated at between 8860 and 8690 cal. BP was not clearly marked in the geochemical record, although slightly lower contents of Ca, Mg, Fe and Mn were noticed during this period. Another sharp decrease of rotifer density (from 41 to 2 ind/cm3) was dated to ca. 8520 cal. BP. From that time onwards, specimens of H. angusticollis become less abundant (1–5 ind./cm3) and finally disappear after ca. 7840 cal BP. According to geochemical data, the decline of Habrotrocha coincides with a decreasing content of Mg, gradually rising Ca/Mg ratio, dropping Fe/Mn ratio and significant fluctuation of the chemical denudation ratio (Na/K). Based on geochemical data, it can be assumed that deterioration of Habrotrocha habitat conditions was associated with a substantial, long-term lowering of the water level probably as a result of a series of droughts. This period can be linked with a well-studied global, cold climate anomaly occurring between 8.6 and 8.0 cal BP (Wanner et al., 2011) (Fig. 5).

Rotifera occurrence in the studied logs in relation to palaeoclimatic changes in the Northern Hemisphere (following various authors)
In the Grel raised bog, the highest densities of H. angusticollis occurs in moderately decomposed ombrogenic peat (Ombro-Sphagnoni, Eriophoro-Sphagneti) at the depth of 55–65 cm dated to 4390–4780 cal BP. In a very similar period (ca. 4190–4620 cal. BP), one of the highest densities of rotifer specimens was recorded in the Imszar peat sequence. In both peat bogs, this period is characterised by a notably higher concentration of Na and an associated rise of the Na/K ratio (Fig. 4). This phase in the Grel site is marked by a twofold lower content of calcuim (Me: 0.24 mg/g) compared to the average value for the whole peat sequence (Me: 0.47 mg/g). The highest density of rotifers in the Grel site coincides with the predominance of A. seminulum, which become the most abundant species of testate amoebae during this period (5.2–14.2%). In the same period, the sphagnum parasite fungi E. lobata (T 13) become less abundant and another fungi, Microthyrium sp., declines completely, accompanied by a decreasing percentage of cyperaceae (from 2.0 to 0.8%) and the disappearance of ericaceae and Vaccinium. During a similar period in the Imszar peat bog, the percentage of Sphagnum spores and its parasitic fungi T. sphagni gradually decrease. In contrast, Gaeumannomyces and Gelasinospora sp. become more frequent, which points to drier conditions (van Geel, 1978; Shumilovskikh et al., 2015) and/or possible fire episodes (Kuhry, 1985, 1997; Muller et al., 2008).
The significant decreasing density of H. angusticollis specimens (from 17 to 1 ind./cm3) registered in the Imszar site after ca. 4180 cal BP coincides with a distinct rise in the Fe/Mn ratio (from 212 to 365), which points to more reducing conditions probably due to a higher water level in the peat bog. At the same time, the concentration of Mg, Ca and Fe increased significantly whereas zinc content decreased six times (from 34.7 to 6 µg/g). A notably higher water level at the Imszar site after around 4190 cal BP was also confirmed by an NPPs analysis which shows a sharp, short-term decrease of sphagnum spores accompanied by rising cyperaceae. Concurrently in the Grel peat bog, the sharp decline in H. angusticollis (117–7 ind/cm3) was associated with a more oxidized environment (Fe/Mn decrease from 227 to 146) and a distinct yet short-lived increase in concentration of almost all elements, which suggests a possible lower water level and humification of the peat (Damman, 1978). Hydrological shifts registered in both peatlands coincided with one of the well-studied Holocene climate anomalies known as the 4.2 ka event (Bond et al., 1997; Wanner et al., 2011). The diverse hydrological response recorded in the studied peatlands is well matched to the spatial heterogeneity of the ‘4.2 ka’ record at the global and continental scale (Weiss et al., 1993; Wanner et al., 2015; Pleskot et al., 2020). So far in Central Europe, especially in Poland, it is considered to be weakly expressed (Starkel et al., 2013; Pleskot et al., 2020), in some cases correlated with an increase in climate humidity and cooling (Magny et al., 2003; Gałka et al., 2013; Karpińska-Kołaczek et al., 2018) either with relatively stable climatic conditions (Gałka et al., 2015; Pleskot et al., 2020) or even droughts (Słowiński et al., 2016).
The rapid decline of Habrotrocha density recorded in the Imszar peat sequence between ca. 1810 and 1330 cal BP was associated with a distinct rise in Fe concentration and simultaneous drop in Ca, Mg and Mn, which points to a higher water level fluctuation or even desiccation of the peat dome. The abruptly decreasing percentage of E. lobata and T. sphagni accompanied by raised Hyalosphenia subflava Cash, 1909, higher decomposition of the peat (R > 65%), and rapid decline of accumulation rate (Margielewski et al., 2022a) supports the hypothesis of the strong desiccation of the Imszar peatland basin after ca. 1810 cal BP. Dry conditions during a similar period (1850–1550 cal BP) were recognised in a peatland located in central Poland (Marcisz et al., 2015). A short-term, drought-related decline and disappearance of H. angusticollis was also noted between 1740 and 1520 cal BP in the Puścizna Wielka raised bog in the Polish Inner Carpathians (Krąpiec et al., 2016). As in our research, the fluctuation of Habrotrocha density in the Puścizna Wielka raised bog was not correlated with Sphagnum percentage or A. flavum, which is commonly considered to be an indicator of a high and stable water level in peat bogs (Payne et al., 2012). It is believed that the climate anomalies registered in northern and central Europe (a series of droughts) around 1600–1400 cal BP were caused by a periodic weakening of North Atlantic Oscillation circulation and were a main push factor for the Migration Period (Drake, 2017).
Conclusions
In the Podemszczyzna peatland, the Habrotrocha crises were correlated both with a moistening and cooling of the climate as well as long-term drainage related to recorded worldwide climatic events: 9.2 ka and 8.2 ka. The climate anomaly known as the 4.2 ka event was responsible for the decline of rotifers in the Grel and Imszar raised bogs, although in the case of the Grel peatland it was probably related to a lower water level in contrast to the Imszar site where a period of higher water level was established by both geochemical and NPPs analysis. Habrotrocha angusticollis decline events were established at ca. 6.0 ka BP, 5.7 ka and 4.2 ka BP (Fig. 5). The estimated duration of phases with H. angusticollis is varied: from ca. 110 to ca. 590 years (average 300 years) in the case of the Grel peatland, from ca. 180 to ca. 1500 years (410 years on average) in the Imszar peatland, and from ca. 1030 to ca. 1200 years in the Podemszczyzna peatland (Figs. 3 and 5).
In the studied profiles, Habrotrocha variability did not overlap with visible lithological changes in the sediment sequences. In the Imszar and Grel peatlands, the occurrence of rotifers was mainly limited to ombrogenic peat; however, they appear only in 45–50% of the samples. In contrast, in the Podemszczyzna peatland Habrotrocha occur in moderately and strongly decomposed sedge peat.
The occurrence of Rotifera in the investigated peatland sediments was correlated with stable redox condition (low Fe/Mn ratio values) and moderate and stable Na/K ratios, which points to periods of higher evapotranspiration without a higher water level and delivery of allochtonous material from the surrounding area as well as a moderate and fairly stable content of all main and trace elements. Palynological analysis of both profiles showed no correlation of rotifers and local vegetation changes (especially with Sphagnum and water plants).
In all profiles, Rotifera occurred between extreme climatic events (wet, dry, cold, warm) recorded in the Northern Hemisphere. In relation to paleoclimatic changes in the Northern Hemisphere, rotifers occurred between wet and dry events and also between cold and warm climatic events. It seems that rotifers preferred stable climatic conditions in the past (Fig. 5).
Multi-proxy analysis of peat deposits and peat stratigraphy are effective in the documentation and reconstruction of periods of dryness and, generally, fluctuation of climate humidity.
The study of subfossil rotifers supported by multi-proxy analysis of peat deposits could be a potential proxy for the indication of periods with stable paleohydrological and palaeoecological conditions (Fig. 5).
The fossil biota including rotifers remains might provide information about changes caused by climate change and human activity.
Data availability
Enquiries about data availability should be directed to the authors.
References
Barber, K. E., F. M. Chambers & D. Maddy, 2003. Holocene palaeoclimates from peat stratigraphy: macrofossil proxy climate record from three oceanic raised bogs in England and Ireland. Quaternary Science Reviews 22: 521–539. https://doi.org/10.1016/S0277-3791(02)00185-3.
Berglund, B. E. & M. Ralska-Jasiewiczowa, 1986. Pollen analysis and pollen diagrams. In Berglund, B. E. (ed), Handbook of Holocene Palaeoecology and Palaeohydrology Wiley, Chichester: 455–484.
Bond, G., W. Showeres, M. Cheseby, R. Lotti, P. Almasi, P. de Menocal, P. Priore, H. Cullen, I. Hajdas & G. Bonani, 1997. A pervasive millennial-scale cycle in North Atlantic Holocene and Glacial climates. Science 278: 1257–1266.
Bond, G., B. Kromer, J. Beer, R. Muscheler, M. N. Evans, W. Showers, S. Hoffmann, R. Lotti-Bond, I. Hajdas & G. Bonani, 2001. Persistent solar influence on north Atlantic climate during the Holocene. Science 294: 2130–2136.
Bond, G., M. N. Evans & R. Muscheler, 2008. North Atlantic Holocene Drift Ice Proxy Data. IGBP PAGES/World Data Center for Paleoclimatology Data Contribution Series # 2008-018. NOAA/NCDC Paleoclimatology Program, Boulder CO, USA. Found at ftp://ftp.ncdc.noaa.gov/pub/data/paleo/contributions_by_author/bond2001/bond2001.txt.
Borówka, R. K., 1992. The pattern and magnitude of denudation in intraplateau sedimentary basins during the Vistulian and Holocene (in Polish with English summary), Vol. 54. Adam Mickiewicz University Press, Seria Geografia: 1–177.
Borówka, R. K., 2007. Geochemiczne badania osadów jeziornych strefy umiarkowanej. Studia Limnologica Et Telmatologica 1: 33–42.
Bronk Ramsey C., 2006. OxCal program v 4.0 [software and documentation]. http://c14.arch.ox.ac.uk/oxcal/hlp_contents.html.
Bronk Ramsey, C., 2008. Deposition models for chronological records. Quaternary Science Reviews 27(1–2): 42–60. https://doi.org/10.1016/j.quascirev.2007.01.019.
Chambers, F. M., R. K. Booth, F. De Vleeschouwer, M. Lamentowicz, G. Le Roux, D. Mauquoy, J. E. Nichols & B. van Geel, 2012. Development and refinement of proxy-climate indicators from peats. Quaternary International 268: 21–33. https://doi.org/10.1016/j.quaint.2011.04.039.
Damman, A. W. H., 1978. Distribution and movement of elements in ombrotrophic peat bogs. Oikos 30: 480–495. https://doi.org/10.2307/3543344.
Drake, B. L., 2017. Changes in North Atlantic oscilation drove population migrations and the collapse of the Western Roman Empire. Scientific Reports 7: 1227.
Fleitmann, D., M. Mudelsee, S. J. Burns, R. S. Bradley, J. Kramers & A. Matter, 2008. Evidence for a widespread climatic anomaly at around 9.2 ka before present. Paleoceanography 23: PA1102. https://doi.org/10.1029/2007PA001519.
Frey, D. G., 1964. Remains of animals in Quaternary lake and bog sediments and their interpretation. Archiv Fur Hydrobiologie Beihefte. Ergebnisse Der Limnologie 2: 1–114.
Frey, D. G., 1986. Cladocera analysis. In Berglund, B. E. (ed), Handbook of Holocene Palaeoecology and Palaeohydrology Wiley, New York: 667–692.
Frey, D. G., 1987. The taxonomy and biogeography of the Cladocera. Hydrobiologia 145: 5–17.
Grimm, E. C., 1991. TILIA and TILIA Graph, Illinois State Museum, Springfield:
Gałka, M., G. Miotk-Szpiganowicz, T. Goslar, M. Jęśko, W. Van der Knaap & M. Lamentowicz, 2013. Palaeohydrology, fires and vegetation succession in the southern Baltic during the last 7500years reconstructed from a raised bog based on multiproxy data. Palaeogeography, Palaeoclimatology, Palaeoecology 370: 209–221. https://doi.org/10.1016/j.palaeo.2012.12.011.
Gałka, M., G. Miotk-Szpiganowicz, M. Marczewska, J. Barabach, W. Van der Knaap & M. Lamentowicz, 2015. Palaeoenvironmental changes in Central Europe (NE Poland) during the last 6200 years reconstructed from a high-resolution multi-proxy peat archive. Holocene 25: 421–434. https://doi.org/10.1177/0959683614561887.
Haas, J. N., I. Richoz, W. Tinner & L. Wick, 1998. Synchronous Holocene climatic oscillations recorded on the Swiss Plateau and at the timberline in the Alps. The Holocene 8: 301–309.
Heiri, O., A. F. Lotter & G. Lemcke, 2001. Loss on ignition as a method for estimating organic and carbonate content in sediments: reproducibility and comparability of results. Journal of Paleolimnology 25: 101–110.
Karpińska-Kołaczek, M., M. Woszczyk, R. Stachowicz-Rybka, A. Obidowicz & P. Kołaczek, 2018. The impact of climate changes during the last 6000 years on a small peatland in North-Eastern Poland: a multi-proxy study. Review of Palaeobotany and Palynology 259: 81–92. https://doi.org/10.1016/j.revpalbo.2018.09.013.
Kondracki, J., 2001. Geografia Regionalna Polski, PWN Warszawa, Warszawa:
Krąpiec, M., W. Margielewski, K. Korzeń, E. Szychowska-Krąpiec, D. Nalepka & A. Łajczak, 2016. Late Holocene palaeoclimate variability: the significance of bog pine dendrochronology related to peat stratigraphy. The Pu´scizna Wielka raised bog case study (Orawa—Nowy Targ Basin, Polish Inner Carpathians). Quaternary Science Reviews 148: 192–208. https://doi.org/10.1016/j.quascirev.2016.07.022.
Kuhry, P., 1985. Transgressions of a raised bog across a coverstand ridge originally covered with an oak-lime forest. Review of Palaeobotany and Palynology 44: 313–353.
Kuhry, P., 1997. The palaeoecology of a treed bog in western boreal Canada: a study based on microfossils, macrofossils and physico-chemical properties. Review of Palaeobotany and Palynology 96: 182–224.
Lamentowicz, M., W. O. Van der Knaap, J. F. N. Van Leeuwen, S. Hangartner, E. A. D. Mitchell, T. Goslar, W. Tinner & C. Kamenik, 2010. A multi proxy high resolution approach to reconstructing past environmental changes from an Alpine peat archive. PAGES News 18: 13–20.
Lang, B., A. Bedford, S. J. Brooks, R. T. Jones, N. Richardson, H. J. B. Birks & J. D. Marshall, 2010. Early-Holocene temperature variability inferred from chironomid assemblages at Hawes Water, northwest England. The Holocene 20: 943–954. https://doi.org/10.1177/0959683610366157.
Magny, M., C. Begeot, J. Guiot & O. Payron, 2003. Contrasting patterns of hydrological changes in Europe in response to Holocene climate cooling phases. Quaternary Science Reviews 22: 1589–1596. https://doi.org/10.1016/S0277-3791(03)00131-8.
Marcisz, K., W. Tinner, D. Colombaroli, P. Kołaczek, M. Słowiński, B. Fiałkiewicz-Kozieł, E. Łokas & M. Lamentowicz, 2015. Long-term hydrological dynamics and fire history over the last 2000 years in CE Europe reconstructed from a high-resolution peat archive. Quaternary Science Reviews 112: 138–152. https://doi.org/10.1016/j.quascirev.2015.01.019.
Margielewski, W., M. Krąpiec, L. Jankowski, J. Urban & V. Zernitskaya, 2015. Impact of aeolian processes on peat accumulation: late Glacial-Holocene history of the Hamernia peat bog (Roztocze region, south-eastern Poland). Quaternary International 2015(386): 212–225. https://doi.org/10.1016/j.quaint.2015.07.016.
Margielewski, W., M. Krąpiec, M. Kupryjanowicz, M. Fiłoc, K. Buczek, R. Stachowicz-Rybka, A. Obidowicz, A. Pociecha, E. Szychowska-Krąpiec, D. Sala & A. Klimek, 2022a. Bog pine dendrochronology related to peat stratigraphy: palaeoenvironmental changes reflected in peatland deposits since the Late Glacial (case study of the Imszar raised bog, Northeastern Poland). Quaternary International 613: 61–80. https://doi.org/10.1016/j.quaint.2021.11.007.
Margielewski, W., M. Krąpiec, K. Buczek, K. Korzeń, E. Szychowska-Krąpiec, A. Pociecha, J. Pilch, A. Obidowicz, D. Sala & A. Klimek, 2022b. Bog pine and deciduous trees chronologies related to peat sequences stratigraphy of the Podemszczyzna peatland (Sandomierz Basin, Southeastern Poland). Radiocarbon 64: 1557–1575. https://doi.org/10.1017/RDC.2022.38.
Margielewski, W., D. J. Michczyńska, K. Buczek, A. Michczyński, K. Korzeń & A. Obidowicz, 2022c. Towards the understanding of the present-day human impact on peatland deposits formed since the late glacial: a “retrospective” age–depth model of the Grel raised bog (Polish Inner Carpathians). Radiocarbon 64: 1525–1543. https://doi.org/10.1017/RDC.2022.62.
Moore, P. D., J. A. Webb & M. E. Collinson, 1991. Pollen Analysis, Blackwell Scientific Publications, Oxford:
Muller, S. D., P. J. H. Richard & B. Talon, 2008. Impact of disturbance on the Holocene development of a temperate peatland (Southern Quebec). Vegetation History and Archaeobotany 17: 713–721.
Pansu, M. & J. Gautheyrou, 2006. Handbook of Soil Analysis. Mineralogical, Organic and Inorganic Methods, Springer, Berlin:
Payne, R. J., M. Lamentowicz, W. O. van der Knaap, J. F. N. van Leeuwen, E. A. D. Mitchell & Y. Mazei, 2012. Testate amoebae in pollen slides. Review of Palaeobotany and Palynology 173: 68–79. https://doi.org/10.1016/j.revpalbo.2011.09.006.
Pleskot, K., K. E. Apolinarska, P. Kołaczek, M. Suchora, M. Fojutowski, T. Joniak, B. Kotrys, M. Kramkowski, M. Słowiński, M. Woźniak & M. G. Lamentowicz, 2020. Searching for the 4.2 ka climate event at Lake Spore, Poland. Catena 191: 1–13. https://doi.org/10.1016/j.catena.2020.104565.
Rasmussen, S. O., B. M. Vinther, H. B. Clausen & K. K. Andersen, 2007. Early Holocene climate oscillations recorded in three Greenland ice cores. Quaternary Science Reviews 26: 1907–1914. https://doi.org/10.1016/j.quascirev.2007.06.015.
Reimer, P., W. Austin, E. Bard, A. Bayliss, P. Blackwell, C. Bronk Ramsey, M. Butzin, H. Cheng, R. Edwards, M. Friedrich, P. Grootes, T. Guilderson, I. Hajdas, T. Heaton, A. Hogg, K. Hughen, B. Kromer, S. Manning, R. Muscheler, J. Palmer, C. Pearson, J. van der Plicht, R. Reimer, D. Richards, E. Scott, J. Southon, C. Turney, L. Wacker, F. Adolphi, U. Büntgen, M. Capano, S. Fahrni, A. Fogtmann-Schulz, R. Friedrich, P. Köhler, S. Kudsk, F. Miyake, J. Olsen, F. Reinig, M. Sakamoto, A. Sookdeo & S. Talamo, 2020. The IntCal20 Northern Hemisphere radiocarbon age calibration curve (0–55 cal. ka BP). Radiocarbon 62: 725–757.
Shumilovskikh, L. S., F. Schlütz, I. Achterberg, A. Bauerochse & H. H. Leuschner, 2015. Non- pollen palynomorphs from mid-holocene peat of the raised bog Borsteler Moor (Lower Saxony, Germany). Studia Quaternaria 32: 5–18. https://doi.org/10.1515/squa-2015-0001.
Słowiński, M., K. Marcisz, M. Płóciennik, M. Obremska, D. Pawłowski, D. Okupny, S. Słowińska, R. Borówka, P. Kittel, J. Forysiak, D. J. Michczyńska & M. Lamentowicz, 2016. Drought as a stress driver of ecological changes in peatland—a palaeoecological study of peatland development between 3500 BCE and 200 BCE in central Poland. Palaeogeography, Palaeoclimatology, Palaeoecology 461: 272–291. https://doi.org/10.1016/j.palaeo.2016.08.038.
Starkel, L., D. J. Michczyńska, M. Krąpiec, W. Margielewski, D. Nalepka & A. Pazdur, 2013. Holocene chrono-climatostratigraphy of Polish territory. Geochronometria 40: 1–21.
Swadling, K. M., H. J. Dartnall, J. A. Gibson, E. Saulnier-Talbot & W. F. Vincent, 2001. Fossil rotifers and the early colonization of an Antarctic lake. Quaternary Research 55: 380–384. https://doi.org/10.1006/qres.2001.2222.
Tołpa, S., M. Jasnowski & A. Pałczynski, 1971. New classification of peat based on phytosociological methods. Bulletin International Peat Society 2: 9–14.
Turton, C. L. & J. H. McAndrews, 2006. Rotifer loricas in second millennium sediment of Crawford Lake, Ontario, Canada. Review of Palaeobotany and Palynology 141: 1–6. https://doi.org/10.1016/j.revpalbo.2006.03.017.
van Geel, B., 1978. A palaeoecological study of Holocene peat bog sections in Germany and The Netherlands, based on the analysis of pollen, spores and macro- and microremains of fungi, algae, cormophytes and animals. Review of Palaeobotany and Palynology 25: 1–120.
van Geel, B., 1986. Application of fungal and algal remains and other microfossils in palynological analyses. In Berglund, B. E. (ed), Handbook of Holocene Palaeoecology and Palaeohydrology Wiley, Chichester: 497–505.
Wanner, H., O. Solomina, M. Grosjean, S. P. Ritz & M. Jetel, 2011. Structure and origin of Holocene cold events. Quaternary Science Reviews 30: 3109–3123. https://doi.org/10.1016/j.quascirev.2011.07.010.
Wanner, H., Mercolli, L., Grosjean, M., Ritz, S.P., 2015. Holocene climate variability and change: a data based review. Journal of the Geological Society 172: 254–263.
Wojewódka, M. & D. Hruševar, 2020. The role of paleolimnology in climate and environment reconstruction and lake restoration in light of research on selected bioindicators. The Holistic Approach to Environment 10: 16–28. https://doi.org/10.33765/thate.10.1.3.
Warner, B. G. & R. Chengalath, 1988. Holocene fossil Habrotrocha angusticollis (Bdelloidea: Rotifera) in North America. Journal of Paleolimnology 1: 141–147.
Weiss, H., M.-A. Courty, W. Wetterstrom, T. Guichard, L. Senior, R. Meadow & A. Curnow, 1993. The genesis and collapse of third millennium north Mesopotamian civilization. Science 261: 995–1004. https://doi.org/10.1126/science.261.5124.995.
Zhang, W., H. Yan, J. Dodson, P. Cheng, C. Liu, J. Li, F. Lu, W. Zhou & Z. An, 2018. The 9.2 ka event in Asian summer monsoon area: the strongest millennial scale collapse of the monsoon during the Holocene. Climate Dynamics 50: 2767–2782.
Funding
This study was supported with funds from the National Science Centre, Poland, Grant No. 2017/25/B/ST10/02439 (2018–2022).
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare no conflict of interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Guest editors: Maria Špoljar, Diego Fontaneto, Elizabeth J. Walsh & Natalia Kuczyńska-Kippen / Diverse Rotifers in Diverse Ecosystems
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Pociecha, A., Buczek, K., Margielewski, W. et al. Appearance of the rotifer community as a potential indicator of stable paleohydrological conditions in peatlands since the Late Glacial: a case study of three wetlands in Poland. Hydrobiologia 851, 2965–2981 (2024). https://doi.org/10.1007/s10750-023-05339-8
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10750-023-05339-8