
Abstract
Stable isotope analyses and derived population-level metrics were used to quantitatively analyse spatial and seasonal heterogeneity in the fish trophic dynamics in relation to environmental variables in Mwanza Gulf, Lake Victoria (Tanzania). The fish community in Lake Victoria, including the top predator Nile perch, is generally omnivorous with a heavy reliance on invertebrates. This is in contrast to findings based on stomach content analyses of Nile perch, which showed a stronger reliance on fish. We tested two hypotheses: (1) during the rainy seasons multiple carbon sources influence the food-web structure inside the Gulf, leading to increased carbon ranges and trophic diversity. (2) During dry periods, the food-web structure mainly relies on pelagic primary production, reducing carbon ranges and trophic diversity. Carbon sources indeed varied seasonally and spatially, affecting the fish community at the highest trophic levels. With the onset of rains, carbon sources became spatially highly differentiated with enriched δ13C values of fish in shallow water inside the Gulf and depleted δ13C values in open waters. Metrics associated with niche size correlated significantly with seasonally varying environmental variables, while δ13C ranges correlated with spatially varying environmental variables.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
The food web of Lake Victoria underwent major ecological changes, including eutrophication (Hecky, 1993) and changes in the fish community. The endemic haplochromines declined (Witte et al., 1992a), followed by an upsurge of the introduced Nile perch (Lates niloticus, Linnaeus, 1758) in the 1980s. During the explosive population growth of Nile perch, there was also a population increase of the atyid shrimp Caridina nilotica (Roux, 1833), the small cyprinid fish Dagaa [Rastrineobola argentea (Pellegrin, 1904)] and the exotic Nile tilapia [Oreochromis niloticus (Linnaeus, 1758)] (Ogutu-Ohwayo, 1990; Witte et al., 1992b; Goldschmidt et al., 1993; Goudswaard et al., 2006). From the 1980s onwards, the Lake’s fishery thrived, dominated by Nile perch as economically most important fish, and Dagaa and Nile tilapia as second and third most important target species (Wanink, 1999; Cowx et al., 2003; Njiru et al., 2005).
The former complex food web with more than 500 species of endemic haplochromines occupying every niche in the lake (van Alphen et al., 2003) was replaced by a simpler food web, dominated by omnivorous feeders (Ojwang et al., 2010), which opportunistically switched between available food sources. Traditional diet analyses showed that during the Nile perch boom, Nile perch preyed heavily on the endemic haplochromines and, when haplochromines declined, they switched their diet to the increasingly abundant Caridina shrimp, their own juveniles and Dagaa (Hughes, 1986; Mkumbo & Ligtvoet, 1992; Katunzi et al., 2006). Since the 1990s, a recovery of some pelagic haplochromine species was observed in the Mwanza Gulf in the south-east of Lake Victoria (Witte et al., 2000, 2007; Kishe-Machumu et al., 2012) and Nile perch is observed to switch to haplochromines again (Kishe-Machumu et al., 2012; Cornelissen et al., 2015). Other abundant species of Lake Victoria also changed their diets. Former specialist feeders such as the haplochromine species (specialized in detritus or zooplankton), Nile tilapia (algae), Dagaa (zooplankton) and Brycinus sadleri (Boulenger, 1906) (zooplankton) are now including larger invertebrate prey and even fish in their diet (Wanink, 1998; Wanink & Witte, 2000; Katunzi et al., 2003; Njiru et al., 2004; Wanink & Joordens, 2007; Kishe-Machumu et al., 2008).
The increasing eutrophication facilitated the explosive population growth of Nile perch and increased fish production in Lake Victoria (Kolding et al., 2008). However, even though the food web simplified and Nile perch boomed, the lake still displayed great spatial heterogeneity in algal biomass and fish distributions (Getabu et al., 2003; Silsbe, 2004). Water quality and other abiotic factors varied spatially and seasonally within the Mwanza Gulf (Cornelissen et al., 2014). Nile perch distributions were primarily driven by water temperature, depth and oxygen and not by the availability of particular prey species (Goudswaard et al., 2011; Cornelissen et al., 2015). As a result the diet of Nile perch also varied spatially and temporally, since it feeds opportunistically on prey which is available in its abiotic-driven distribution range (Cornelissen et al., 2015).
Most studies in Lake Victoria focused on stock assessments and diet studies of a few commercial species, giving limited and ‘snapshot’ information on the organization of the food web and trophic interactions. Combined carbon (δ13C) and nitrogen (δ15N) stable isotopes provide information on food sources, trophic interactions and food-web structure of the entire communities, covering a longer integrated timespan (Peterson & Fry, 1987; Hesslein et al., 1993; Layman et al., 2012). However, only few isotopic studies on food-web dynamics of Lake Victoria have been conducted, and these were limited to two Gulfs in the northern part of Lake Victoria (Branstrator et al., 2003; Campbell et al., 2003; Ojwang et al., 2010).
In addition to identifying the resource use of individual organisms and species, community-wide metrics can be used to quantify trophic structure on a food-web level of the entire communities or populations in a number of diagnostic metrics (Layman et al., 2007). This approach has been used to compare food-web structures between ecosystems, seasons and habitats (Carlier et al., 2009; Jackson et al., 2012; Doi et al., 2013; Abrantes et al., 2014), and to study the effects of invasive species on trophic interactions of populations (Jackson et al., 2012).
The environment affects productivity rates and resources at the base of the food web which are supporting higher trophic levels. In ecosystems which are strongly affected by its hydrology such as wetlands, rivers, and estuaries, variation in environmental and physical conditions reflects variation in food-web structure (Doi et al., 2013; Hyndes et al., 2013; Schriever, 2015; Thormar et al., 2016). At present little is known about how the environment structures the food web at higher trophic levels in large lake systems such a Lake Victoria.
Mwanza Gulf is spatially and seasonally heterogeneous in water quality and the distribution and diets of fish (Cornelissen et al., 2014, 2015). Therefore, we hypothesize that fish trophic interactions will also vary spatially and seasonally in relation to environmental variables (O’Neill & Thorp, 2014; Schriever, 2015). To test this, we used 8 metrics on population level to detect spatial and seasonal shifts of haplochromines and Nile perch and correlated them to several important environmental variables. We expected that during the rainy periods, when run-off is high, the fish community inside the Gulf will be more influenced by terrestrial and benthic/littoral carbon sources than during the dry season, when primary production will be mainly pelagic. These multiple carbon sources are expected to lead to increased carbon ranges and trophic diversity during the rainy periods, as compared to the dry season.
Materials and methods
Study area
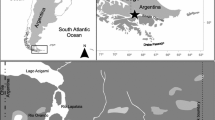
Samples were collected at 6 stations in three areas in Mwanza Gulf (Fig. 1). Mwanza Gulf is located in south-east Lake Victoria in Tanzania and is about 60 km long, 2.5–11 km wide, 3–25 m in depth, and covers a surface area of approximately 500 km2 (Witte & van Densen, 1995). Mwanza City is located in the north near the entrance of the Gulf (Fig. 1); here the shoreline is completely urbanized. Outside the city range, the littoral zone is characterized by a mixed vegetation of papyrus (Cyperus papyrus Linnaeus), reeds, e.g. [Phragmites australis (Cav.) Trin. ex Steud.] and water hyacinth [Eichhornia crassipes, (Martius) Solms-Laubach], and by rock formations. The bottom of Mwanza Gulf consists of soft, fine-grained sediment. The south of the Gulf at area 1 (station 1 and 2) is shallow with a depth of <5 m (Fig. 1). This area can be considered as a littoral habitat with benthic and terrestrial influences. Area 3 (station 5 and 6) is located in the open lake outside the Gulf with a depth of 15–25 m (Fig. 1). This area can be considered as a pelagic habitat. Area 2 (stations 3 and 4) has an intermediate depth range of 5–10 m with both littoral and pelagic habitat characteristics (Fig. 1).

Map of the Mwanza Gulf with six sampling stations. Stations were divided in three areas: (1) shallow = depth range of <5 m, (2) intermediate = depth range of 5–10 m and (3) deep = depth range of 15–25 m
Climate
The climate in Lake Victoria is characterized by three seasons, based on the yearly monsoon cycles. During the cooler dry season, from June to August, strong southerly winds and low precipitation keep the water well-mixed, isothermal and oxygenated (Talling, 1966; Akiyama et al., 1977). During the short rainy season, from September to December, winds decline and rainfall and temperature increase, warming up the surface layer of the water column, which becomes gradually stratified. During the long rainy season, from January to May, precipitation is high and stratification of the water column becomes strong (Talling, 1966). During the sampling period from 2009 to 2011, Mwanza Gulf had similar seasons, although during the long rains of 2010, there was twice as much rainfall as during the long rains of 2011 (Supplementary Fig. S1).
Data collection
Biological samples were collected from September 2009 to March 2011 during the short rains (September–November 2009), long rains (April–May 2010 and January–March 2011) and dry season (July–August 2010) (Table 1). The datasets gathered during the long rains of 2010 and 2011 were pooled together as isotope values did not differ between these years. Areas were sampled on 1–3 days each season (Table 1).
Fish were caught using gill nets and surface trawling at night. Fish were classified to species level, except for haplochromines, which, for this study, were classified as one group. This was justified by the fact that more than 94% of the haplochromines belonged to species classified as pelagic zooplanktivores or benthic detritivores, which, since their recovery in the 1990s, have similar broad diets in the Gulf including larger invertebrates (Kishe-Machumu et al., 2008, I. J. M. Cornelissen unpublished data) and also have similar habitat utilization. Invertebrates were collected during the day. Caridina shrimp and gastropods were collected by hauling a 1 m2 square lift net from bottom to surface. Zooplankton was collected by hauling a 150-μm net from bottom to surface and stored in 50-ml jars.
In fish >5 cm standard length, ca. 0.5 cm3 of skinned dorsal muscle tissue was collected. Fish <5 cm were entirely skinned and filleted. Fish tissue was rinsed in distilled water before preservation. Zooplankton was killed with ethanol and then transferred to 2-ml tubes. Gastropods were kept in regularly refreshed, distilled water for 24 h to allow for emptying their digestive tracts from food particles, after which their shells were removed and body tissue was preserved. Caridina and insect specimens were preserved entirely. All invertebrates and fish samples were preserved in 70% ethanol and stored at −20°C. The preserved samples were shipped to the laboratory of the Netherlands Institute of Ecology (NIOO) in the Netherlands for further sample preparation and analyses. There, samples were drained off, freeze-dried and homogenized with stainless steel balls (4 mm in diameter) in 2-ml Eppendorf tubes. Of each sample, 300–500 μg of tissue powder was weighed before δ13C and δ15N isotope analysis. Stable isotopes of samples collected in 2009 were analysed with an Elemental analyser (Eurovector), interfaced with a Conflo 2 device for continuous flow and an Isotope Ratio Mass Spectrometer (IRMS) (Delta XP Advantage, Thermo, Germany). The samples collected in 2010–2011 were measured with an Elemental analyser Flash2000 (Thermo) interfaced with a Conflo 4 and an IRMS (Delta XP Advantage, Thermo, Germany). The isotope ratios were measured against reference standards of carbon and nitrogen using USGS40 l-glutamic Acid C5H9NO4, with δ15N of −4.5‰ and δ13C of −26.389‰.
C:N ratios are related to lipid content (Post et al., 2007). Our mean C:N ratios varied from 3.13 to 4.44 between taxa (Supplementary Table S1). We therefore applied a posteriori mathematical normalization of δ13C values using the lipid-correction equations given by Post et al. (2007). We corrected for consumer values (invertebrates and fish). The corrections (δ13C) rely on carbon to nitrogen ratios (C:N):
Environmental data of chlorophyll a, mean depth, KPAR (attenuation coefficient) and water temperature were collected during this study in the Mwanza Gulf and calculated as described in Cornelissen et al. (2014). Mean wind speed and rainfall were based on data measured in Mwanza by the National Centers for Environmental Information (NOAA-NCEI data, retrieved on May 10, 2011). The environmental data are listed in Supplementary Table S2.
Nile perch diets were analysed by dissecting the stomachs. Stomach fullness was classified into five categories: 0–1% (empty), 1–25%, 25–50%, 50–75% and >75% (full) (Witte & van Densen, 1995). The relative volume per prey category of Nile perch was estimated into the same five categories. Prey were categorized as Caridina shrimp, haplochromines, Odonata, R. argentea, Nile perch and fish remains.
Data analyses
Baseline correction
δ15N values of primary production sources vary in space and time. Therefore, to reliably compare trophic levels (TL) of consumers, their δ15N values needed to be corrected with baselines. Since samples were collected in two distinct habitats, i.e. shallow, littoral waters inside the Gulf and open pelagic waters near the entrance, we used a two-end-member mixing model with a littoral and a pelagic source for calculating trophic levels (Post, 2002). The δ15N baseline was identified by the two-end-member organisms in δ13C, which represent the littoral (baseline 1) and pelagic baseline (baseline 2). We used primary consumers as end members and identified these for each area and season separately (Supplementary Table S3). δ15N and δ13C of the littoral baseline did not show any seasonal or spatial variation (Supplementary Table S4) and therefore we used the mean littoral δ15N and δ13C to calculate trophic levels. The pelagic baseline of δ15N and δ13C varied across areas (Supplementary Table S4) and therefore we used a mean pelagic δ15N and δ13C of each area to calculate trophic levels. Trophic levels were calculated as (Post, 2002)
where λ is the trophic position of the baseline organism, δ15Nsc is the measured δ15N of the consumer of interest, δ15Nb1 is the measured δ15N of baseline 1, δ15Nb2 is the measured δ15N of baseline 2, and Δn is the enrichment in δ15N per trophic level. We used a λ value of 2 for the trophic position of the baseline organisms. We used 2.4‰ as enrichment per tropic level, which is used in Lake Tanganyika, a rift valley lake (Kilham et al., 2009), and also appropriate for Lake Victoria. The letter α represents the proportion of nitrogen derived from baseline 1 and can be estimated as (Post, 2002)
where δ13Csc is the measured δ13C of the consumer of interest, δ13Cb1 is the measured δ13C of baseline 1, and δ13Cb2 is the measured δ13C of baseline 2.
Population-level metrics
We used 8 diagnostic population-level metrics (Table 2) based on the δ13C and δ15N isotope data to reveal spatial and seasonal differences in niche use and trophic diversity of populations of haplochromines and Nile perch in Mwanza Gulf. The metrics nitrogen range (dNr), carbon range (dCr), corrected standard ellipse area (SEAc) and mean distance to centroid (CD) indicate the total isotopic space used in the δ13C–δ15N bi-space by a trophic group. The metrics nitrogen range (dNr) and carbon range (dCr) indicate the total carbon or nitrogen range used by a trophic group and are calculated as the difference between the lowest and highest values of δ15N or δ13C within a trophic group (Layman et al., 2007). We used SEAc as a measure of the core niche area. It is calculated from the variance and covariance between δ13C and δ15N, containing about 40% of the data points (Batschelet, 1981) and is corrected for small samples sizes according to Jackson et al. (2011). Mean δ13C and mean δ15N represent the centre of the core niche area. CD is calculated as the mean Euclidean distance of each individual to the δ13C–δ15N centroid, and is a metric of mean degree of trophic diversity (Layman et al., 2007).
The metrics packing density (MNND) and trophic evenness (SDNND) give information on the dispersion of isotopic values in the bi-space and are related to trophic redundancy. MNND is calculated as the mean of nearest neighbour distance between individual data points and measures the density of data point packing (Layman et al., 2007). A group of individuals with different trophic ecology will be more widely spread in isotopic bi-space and exhibit higher MNND values than a group of individuals with similar trophic ecology. Higher MNND values indicate decreased redundancy. SDNND indicates how evenly (low value) or clustered (high value) the isotope values are spread in isotopic bi-space (Layman et al., 2007). Clustering means that carbon sources and diets of organisms within a cluster are similar, whereas carbon sources and diets differ between separate clusters.
Jackson et al. (2011) developed a Bayesian statistical method to compare metrics across communities with the assumption of a normal distribution. However, in our study, normality of residuals of the isotope data could not be achieved directly or after transformations. In addition, sample sizes varied between areas and seasons. Therefore, bootstrapped means and 95% confidence intervals of the Layman metrics were calculated for each area and season. Spatial and seasonal differences in metrics were significant if 95% confidence intervals did not overlap. We used 10,000 replicates and a minimum of 8 observations to perform the bootstrap analysis (Table 2). As a comparison to the more standard analysis of Bayesian metrics, where normality is assumed, we also analysed Bayesian standard ellipses (SEAb) to compare to the bootstrapped version of SEAc. By comparing both methods, a robust result was acquired.
Moreover, we used Bayesian mixing models to estimate the relative contributions of resources to the diet of Nile perch as compared to results from stomach contents. We ran separate models with prior information on stomach content proportions of prey of each area and season. In this way, the spatial and seasonal variability in resources were included and corrected for the diet estimates. Based on main prey found in stomach contents, Caridina shrimp, Nile perch <10 cm TL, haplochromines, Odonata larvae and R. argentea were used as resources.
For some area–season combinations, resource isotope values were lacking (e.g. Caridina shrimp in areas 1 and 2 in the short rains). We then used the mean of δ15N and δ13C of the resource of samples collected during the study period in the remaining areas. Unfortunately, a similar analysis could not be performed for haplochromine diets, due to the lack of isotope values of detritus and phytoplankton, two major diet items of haplochromines (I. J. M. Cornelissen, unpublished data). We assumed a fractionation factor per trophic level of 2.4‰ for δ15N (Lake Tangyanika) (Kilham et al., 2009) and 0.4‰ for δ13C (Post, 2002).
Kendall tau correlations were used to relate significant differences in metric values of Table 2 with the environmental variables of Supplementary Table S2. R version 3.1.0 (R Core Team, 2014) was used for the Kendall tau correlations. Diet contributions were calculated using the SIAR package. Metrics were calculated using the SIBER analyses (Jackson et al., 2011) within the SIAR package (Parnell et al., 2008).
Results
The number of fish sampled varied between 20 and 119 for the different areas and seasons (Table 1). In general, haplochromines and Nile perch were the most dominant fish taxa caught in Mwanza Gulf (7–55% and 27–82% of total numbers, respectively). 86% of Nile perch caught were juveniles (standard length of <40 cm) with a mean standard length of 21.8 cm and a size range of 2–67 cm, which did not vary significantly over space and time. Mean δ13C isotope ratios of each taxon in Mwanza Gulf ranged from −21.4 [Caecomastacembelus frenatus (Boulenger, 1901)] to −14.2‰ [Bagrus docmak (Forsskäl, 1775)] (Fig. 2, Supplementary Table S5). The variation in the δ13C isotope ratios caused taxa to overlap (see error bars in Fig. 2). Synodontis afrofischeri Hilgendorf, 1888 was most variable in δ13C values, with standard errors ranging from −19.3 to −14.9‰.

Mean δ13C and δ15N values with standard errors of taxa found in the Mwanza Gulf. Symbols represent trophic groups of taxa (haplochromine cichlids = reversed triangle, Nile perch = squares, other fish = diamonds and invertebrates = circles). Core niche areas (SEAc-ellipses) of trophic groups are indicated for haplochromine cichlids (blue), Nile perch (red), other fish (yellow) and invertebrates (green). Details of trophic groups are listed in Table 1
Mean δ15N values ranged from 4.9 (gastropods) to 10.1‰ (B. docmak)) in Mwanza Gulf (Fig. 2, Supplementary Table S5). Zygoptera nymphs showed large variation in mean δ15N, with standard errors ranging from 3.3 to 7.3‰. Amongst the fish, Schilbe intermedius (Rüppell, 1832), Nile perch, haplochromines, Brycinus sadleri and Mormyrus kannume Forsskäl, 1775 had similar δ15N values of ca. 8.5‰. Rastrineobola argentea, Synodontis victoriae Boulenger, 1906, B. docmak and C. frenatus showed the highest δ15N values (ca. 10‰) and Oreochromis niloticus the lowest (5.2‰).
The core niche area (SEAc-ellipses in Fig. 2) of all invertebrates combined was larger than that of haplochromines, Nile perch and other fish. These three fish groups showed large overlap amongst each other, but not with invertebrates.
Seasonal and spatial dynamics in fish interactions
We observed clear seasonal and spatial shifts along the δ13C-axis in the position of core niche areas of Nile perch and haplochromines in Mwanza Gulf (SEAc-ellipses in Fig. 3, for seasonal and spatial mean δ13C and δ15N values of all species see Supplementary Table S6). During the short rains, strong spatial differences between niches occurred along the δ13C axis for both haplochromine cichlids and Nile perch (Fig. 3a, d). Seasonal mean δ13C values were especially different in area 1 (highest values) compared to areas 2 and 3 (lowest values) (Table 2). This means that niches of both haplochromines and Nile perch were spatially differentiated (Fig. 3a, d). This differentiation was enhanced by the relatively small core niche area and carbon range in area 1 and/or areas 2 and 3 during the short rains (SEAc and dCr, Table 2). A similar, spatial effect was visible during the long rains, but less strong (Fig. 3b, e), because mean δ13C values were more similar and in Nile perch also because of the relatively large core niche areas (SEAc, Table 2). During the dry season, mean δ13C values and core niche areas of the three areas overlapped (Fig. 3c, f, mean δ13C, Table 2), with a large core niche area and dCr in area 1 of haplochromines (SEAc, dCr, Table 2).

Spatial and seasonal variation of mean δ13C and δ15N values and core niche areas (SEAc-ellipses) of (a–c) haplochromines, d–f Lates niloticus (Nile perch). Error bars represent 95% confidence intervals. Means were bootstrapped. For location of sampling stations and the three areas see Fig. 1
Trophic diversity (CD) varied less than core niche area (SEAc). Trophic diversity of haplochromines did not vary spatially and seasonally (Table 2). Trophic diversity of Nile perch was smallest in area 1 in short rains and largest in area 3 in long rains (Table 2).
There was no overall spatial and seasonal pattern in packing density (MNND), trophic evenness (SDNND) and mean δ15N of haplochromines and Nile perch (Table 2). Mean δ15N did show some variation with lowest values in area 1 during the long rains for both haplochromines and Nile perch. MNND and SDNND did not vary spatially and seasonally (Table 2).
Depth was negatively correlated with δ13C values, whereas chlorophyll was positively correlated with δ13C values for Nile perch and haplochromines (Table 3). Core niche areas (SEAc) and carbon range (dCr) correlated with seasonal environmental variables. Core niche area of haplochromines was negatively related to water temperature and carbon range was negatively related to rainfall and water temperature (Table 3). Core niche area, trophic diversity (CD) and carbon range of Nile perch were negatively related to wind speed (Table 3).
Haplochromines and Caridina both dominated the diet of Nile perch in stomach contents (Table 4). Haplochromines, Caridina, Nile perch and Odonata dominated the diet of Nile perch based on isotopes (Table 4). Relative contribution of Caridina to the diet of Nile perch based on both isotopes and stomach contents was highest in areas 2 and 3 in short and long rains (Table 4). Relative contribution of haplochromines decreased from area 1 to area 3 during the short and long rains and was similar for areas during the dry season based on isotopes and stomach content (Table 4). The relative contributions of R. argentea were low for both isotopes and stomach contents (Table 4). The contributions of Odonata and Nile perch were higher based on isotopes than based on stomach contents, which resulted in lower contribution of haplochromines based on isotopes than on stomach contents (Table 4).
Discussion
We used δ13C and δ15N stable isotope ratios, quantitative population-level metrics and diets to investigate the trophic dynamics of fish species in Mwanza Gulf, Lake Victoria. However, there are some caveats, such as the fact that our samples were preserved in ethanol before isotope analyses. Although samples were preserved consistently, ethanol could have affected isotope values. Ethanol reduces lipid content of tissue and, therefore, ethanol mostly affect δ13C (Kaehler & Pakhomov, 2001; Cummings et al., 2010; Syvärante et al., 2011; Ford et al., 2016) and effects are species-specific (Kelly et al., 2006; Syvärante et al., 2008). The C:N ratios of our taxa ranged between 3.1 and 4.4, indicating relatively low lipid contents between taxa (Supplementary Table S1) and thus a minimal lipid effect on δ13C values. Furthermore, we used a lipid normalization (Post et al., 2007), which corrects for variable lipid contents affecting δ13C values. We are convinced that in this way ethanol effects were minimal.
We used bootstrapped metrics instead of the commonly used Bayesian metrics. We compared the bootstrapped standard ellipse area (SEAc) with the Bayesian standard ellipse area (SEAb) for validation. In general, SEAc corresponded well with SEAb, especially when sample sizes were large as in the case of Nile perch. This validates our use of bootstrapped metrics.
Most fish in the Mwanza Gulf were positioned mid-range compared to the carbon ranges of invertebrates in the food web. This indicates omnivory at higher trophic levels and fish relying heavily on invertebrates. This is in contrast with the large diversity of species occupying many different feeding niches, based on stomach and gut analyses, observed in Lake Victoria before the Nile perch upsurge (van Alphen et al., 2003). Many abundant fish species, for instance O. niloticus, B. sadleri, R. argentea and haplochromines, that were originally known as phytoplankton, zooplankton or detritus feeders extended their diets with larger invertebrates, such as insects, Caridina shrimps and even small fishes (Wanink, 1998; Wanink & Witte, 2000; Katunzi et al., 2003; Njiru et al., 2004; Wanink & Joordens, 2007; Kishe-Machumu et al., 2008).
Lake Victoria’s top predator, Nile perch, showed relatively low δ15N values in the food web of Mwanza Gulf compared to other consumers. Similar δ15N values between Nile perch and haplochromines and an enrichment of 2–3‰ between Nile perch and invertebrates indicate that Nile perch relies more on invertebrates than fish. This agrees with a diet of Caridina shrimps found in stomachs of juvenile Nile perch after the decline of haplochromines (Katunzi et al., 2006). However, more recent diet analyses of stomach contents in the Mwanza Gulf revealed that haplochromines and Caridina both dominated the diet of juvenile Nile perch of <40 cm (Kishe-Machumu et al., 2012). This is confirmed by stomach analyses in this study. We therefore observed large differences in the diet estimations between stomach content and isotope analyses. The contribution of haplochromines to the diet of Nile perch was smaller based on isotopes than based on stomach contents. Odonata and small Nile perch showed a considerable contribution in isotope diets, whereas in stomach contents they were only contributing little to the diet of Nile perch.
There are several reasons that might explain the discrepancy between isotopes and stomach contents. Firstly, the Bayesian models used for estimating isotope diets might have resulted in some bias in estimates. Most datasets were not normally distributed. The model also showed difficulties distinguishing between sources, especially between diets of small Nile perch and haplochromines, Odonata and Caridina. This was mainly caused by a high variability within sources in each area and season, which results in a under- or over-estimation in sources, i.e higher contributions of Odonata and Nile perch and lower contributions of haplochromines to the isotope diets. This highlights the importance of incorporating resource variation in mixing models (Hyndes et al., 2013). Furthermore, we used a constant enrichment in δ15N between Nile perch and the different prey types in the model. However, enrichment varies between different trophic transfers because of differences in δ15N assimilation between types of prey (McCutchan et al., 2003). Still, this cannot fully explain the similar δ15N levels we observed between Nile perch and its haplochromine prey, because the enrichment is larger for fish predators feeding on high-protein preys such as fish, than for fish feeding on invertebrate prey (McCutchan et al., 2003).
Secondly, the discrepancy between isotope diet estimations and stomach contents may be caused by Nile perch’s feeding behaviour. The relative high proportions of fish found in stomachs were of Nile perch caught at night and only represent its night-time feeding behaviour. As Nile perch exhibits a diel feeding pattern of consuming more shrimps during the day and more fish at night (Mkumbo & Ligtvoet, 1992; Goudswaard et al., 2004), the contribution of fish in its diet as based on stomach content analyses is probably overestimated.
Thirdly, isotopes cover an integrated timespan of weeks compared to the timespan of hours in diets of stomach analyses. A Nile perch sampled in the Gulf may have been feeding on haplochromines there, resulting in a higher proportion of fish in stomach analyses, but may have been feeding previously on Caridina shrimps outside the Gulf where Caridina densities are higher and where it is still a major prey in Nile perch diets (Ngupula & Mlaponi, 2010; Cornelissen et al., 2015). Nevertheless, Nile perch isotope values correspond more to an invertebrate-based diet and haplochromines are a less important food source for Nile perch than is suggested by stomach content analyses alone. This highlights the importance of both isotope and stomach content methods to study resource use in consumers.
After the Nile perch upsurge in the late 1980s, the Lake Victoria food web simplified with Caridina shrimp, R. argentea, O. niloticus, zooplanktivorous and detritivorous haplochromine cichlids and Nile perch dominating the food web. When comparing our estimates of isotopic trophic positions, most taxa in our study correspond with the estimates reported for 1977, 2000 and 2005 in Lake Victoria (Table 5) (Matsuishi et al., 2006; Downing, 2012). Therefore, despite the loss of biodiversity in Lake Victoria since the Nile perch upsurge in the 1980s (Witte et al., 1992a), the overall trophic structure of the food web seems unchanged since 1977 when haplochromines still dominated the food web. Yet, Nile perch in our study, especially of >40 cm, showed lower trophic levels than in 1977, 2000 and in 2005 (Table 5). However, the higher trophic levels observed in those years for Nile perch were all solely based on stomach content analyses and are therefore probably overestimates.
We found large seasonal differences in carbon sources of fish in Mwanza Gulf. In contrast to our hypothesis, differentiation in the niche position characterized fish during the short rainy season, because of a large spatial differentiation in carbon sources and smaller carbon ranges and core niche area especially in area 1 during the short rains (dCr, SEAc, Table 2; Fig. 3). This spatial differentiation of carbon sources was probably caused by the increased run-off from the land into the Gulf, the effect of which was greatest in area 1, deep inside the Gulf and decreased towards area 3 in the open water outside the Gulf. The negative δ13C values found in the open water outside the Gulf indicate that isotopic composition of fish here was mainly influenced by pelagic phytoplankton (France, 1995; Hecky & Hesslein, 1995; Keough et al., 1996; Schindler et al., 1997).
The extreme positive δ13C values inside the gulf in shallow water indicate an increased effect of littoral/benthic influence and/or increased primary production on fish (France, 1995; Casey & Post, 2011). Littoral primary producers such as macrophytes, benthic algae and increased phytoplankton growth often seen in shallow inshore areas all result in enriched δ13C values. However, macrophytes probably contribute little to the food chain of haplochromines and Nile perch in Mwanza Gulf because they do not feed on macrophytes (Katunzi et al., 2003; Kishe-Machumu et al., 2008; Cornelissen et al., 2015), nor do their prey that feed on detritus, phytoplankton, zooplankton and invertebrates. This is consistent with studies in a satellite lake of Lake Victoria, where macrophytes contribute little to consumers in the food web (Hecky & Hesslein, 1995; Mbabazi et al., 2010).
In eutrophic water, like Mwanza Gulf, phytoplankton production often causes light limitation for the benthic primary production pathway in the food web (Vadeboncoeur et al., 2003). Therefore, benthic algae are probably of minimal influence on fish. High primary productivity and high rates of N-fixation with associated phytoplankton blooms may also result in an enrichment of δ13C (Mugidde, 2001; Casey & Post, 2011). The higher phytoplankton biomass in shallow waters compared to deep waters (Supplementary Table S2) indicates higher primary production rates in shallow waters, which might have contributed to an enriched δ13C of fish in shallow water. N-fixation rates were low in Mwanza Gulf (Cornelissen et al., 2014) and, therefore, N-fixing bacteria are unlikely to be a major carbon source for fish. Terrestrial carbon also has typically enriched δ13C values. The rainfall in September to November 2009 probably increased the run-off of terrestrial carbon and therefore might also explain the spatial differentiation in carbon sources and the enriched δ13C values of fish in shallow waters during the short rains. The long rains showed a similar, but less extreme spatial shift in carbon sources as the short rains, indicating a more mixed use of carbon sources by fish. The long rains seemed to represent a transition period where carbon sources start to diffuse over the Gulf, before becoming mixed during the dry season.
The dry season showed similar δ13C ranges of fish between areas which resulted in overlapping niches between areas (Fig. 3). This indicates that during this time fish were omnivorous, using a mixture of carbon sources regardless of their location in the Gulf. This can be explained by the combination of increasing winds and little rainfall during the dry season (Fig. S1), which resulted in a well-mixed column and no run-off. This caused a mixed carbon environment throughout the gulf at the base of the food web.
Diet analyses of isotopes and stomach contents showed decreasing proportions of Caridina shrimp and increasing proportions of haplochromines in the diet of Nile perch from shallow to deep water during the rainy periods. This was probably caused by spatial variation in Nile perch size at which ontogenetic diet shifts occur (Cornelissen et al., 2015). The seasonal high abundance of Caridina in deep water and the corresponding high occurrence of Caridina in their diets indicate here a late dietary shift of Nile perch from invertebrates to fish (Cornelissen et al., 2015). In shallow water, Nile perch shifted from Caridina to fish at a much smaller size (Cornelissen et al., 2015), leading to a higher contribution of haplochromines in shallow water.
Spatial and seasonal changes in carbon sources at the base of the food web affected carbon use and in some extent niche size and trophic diversity of fish at higher trophic levels. In our study, depth and chlorophyll a correlated with the spatial distribution of available carbon sources. This is also observed in estuaries where abiotic factors such as nutrients and light reflect the spatial variability of carbon sources (Hyndes et al., 2013). In our study, seasonal environmental factors such as wind, rainfall and temperature correlated with trophic diversity and niche size. In estuaries, seasons also influence trophic diversity of food-web structures where higher variabilities of carbon sources due to primary production and terrestrial inputs result in increased trophic diversities after rainy periods (Abrantes et al., 2014).
Conclusion
We conclude that fish in the food web of Lake Victoria’s Mwanza Gulf are omnivorous with a heavy reliance on invertebrates and that environmental factors influence the higher trophic levels of the food web on spatial and seasonal scales. The significant effect of environmental factors on food-web dynamics has previously been shown in highly fluctuating systems such as ponds along a hydroperiod gradient, rivers, wetlands and coastal areas (Zeug & Winemiller, 2008; Hyndes et al., 2013; O’Neill & Thorp, 2014; Schriever, 2015; Thormar et al., 2016). The patterns of heterogeneity in food webs of lakes should be investigated further in order to explain responses to environmental changes such as eutrophication. Eutrophication often results in a loss of diversity and a simplified food web (Coll et al., 2011; Thormar et al., 2016; Xu et al., 2016). In Lake Victoria, the increasing eutrophication and introduction of Nile perch led to a high degree of omnivory and functional redundancy in the food web and the dominance of resilient species (Ojwang et al., 2010). However, a simplified food web with few trophic links is more vulnerable for collapse and as eutrophication continues in Lake Victoria, it is unknown how this will affect food-web dynamics in the future.
References
Abrantes, K. G., A. Barnett & S. Bouillon, 2014. Stable isotope-based community metrics as a tool to identify patterns in food web structure in east African estuaries. Functional Ecology 28: 270–282.
Akiyama, T., A. A. Kajumulo & S. Olsen, 1977. Seasonal variation of plankton and physicochemical condition in Mwanza Gulf, Lake Victoria. Bulletin of Freshwater Fisheries Research Laboratory 27: 48–61.
Batschelet, E., 1981. Circular Statistics in Biology. Academic Press, London.
Branstrator, D. K., L. Mwebaza-Ndawula & J. P. Montoya, 2003. Resource–consumer relationships in Lake Victoria, East Africa. Hydrobiologia 493: 27–34.
Campbell, L. M., R. E. Hecky & S. B. Wandera, 2003. Stable isotope analyses of food web structure and fish diet in Napoleon and Winam Gulfs, Lake Victoria, East Africa. Journal of Great Lakes Research 29: 243–257.
Carlier, A., P. Riera, J. M. Amouroux, J. Y. Bodio, M. Desmalades & A. Gremare, 2009. Spatial heterogeneity in the food web of a heavily modified Mediterranean coastal lagoon: stable isotope evidence. Aquatic Biology 5: 167–179.
Casey, M. M. & D. M. Post, 2011. The problem of isotopic baseline: reconstructing the diet and trophic position of fossil animals. Earth-Science Reviews 106: 131–148.
Coll, M., A. Schmidt, T. Romanuk & H. K. Lotze, 2011. Food-Web structure of seagrass communities across different spatial scales and human impacts. PLoS ONE 6: e22591.
Cornelissen, I. J. M., G. Silsbe, J. A. J. Verreth, E. van Donk & L. A. J. Nagelkerke, 2014. Dynamics and limitations of phytoplankton biomass along a gradient in Mwanza Gulf, southern Lake Victoria (Tanzania). Freshwater Biology 59: 127–141.
Cornelissen, I. J. M., P. A. M. van Zwieten, H. K. Peter & L. A. J. Nagelkerke, 2015. Nile perch distribution in south-east Lake Victoria is more strongly driven by abiotic factors, than by prey densities. Hydrobiologia 755: 239–255.
Cowx, I. G., M. van der Knaap, L. I. Muhoozi & A. Othina, 2003. Improving fishery catch statistics for Lake Victoria. Aquatic Ecosystem Health & Management 6: 299–310.
Cummings, D. O., D. J. Booth, R. W. Lee, S. J. Simpson & A. J. Pile, 2010. Ontogenetic diet shifts in the reef fish Pseudanthias rubrizonatus from isolated populations on the North-West Shelf of Australia. Marine Ecology Progress Series 419: 211–222.
Doi, H., E. I. Zuykova, E. Kikuchi, H. Ota, N. I. Yurlova & E. Yadrenkina, 2013. Isotopic evidence for the spatial heterogeneity of the planktonic food webs in the transition zone between river and lake ecosystems. PeerJ 1: e222.
Downing, A.S., 2012. Seeing the water for the fish: building on perspectives of Lake Victoria PhD. thesis, Wageningen University, Wageningen, The Netherlands.
Ford, A. G. P., L. Rüber, J. Newton, K. K. Dasmahapatra, J. D. Balarin, K. Bruun & J. J. Day, 2016. Niche divergence facilitated by fine-scale ecological partitioning in a recent cichlid fish adaptive radiation. Evolution 70–12: 2718–2735.
France, R. L., 1995. Differentiation between littoral and pelagic food webs in lakes using stable carbon isotopes. Limnology & Oceanography 40: 1310–1313.
Getabu, A., R. Tumwebaze & D. N. Maclennan, 2003. Spatial distribution and temporal changes in the fish populations of Lake Victoria. Aquatic Living Resources 16: 159–165.
Goldschmidt, T., F. Witte & J. H. Wanink, 1993. Cascading effects of the introduced Nile perch on the detritivorous/phytoplanktivorous species in the sublittoral areas of Lake Victoria. Conservation biology 7: 686–700.
Goudswaard, K. P. C., J. H. Wanink, F. Witte & E. F. B. Katunzi, 2004. Diel vertical migration of major fish-species in Lake Victoria, East Africa. Hydrobiologia 513: 141–152.
Goudswaard, K. P. C., F. Witte & J. H. Wanink, 2006. The shrimp Caridina nilotica in Lake Victoria (East Africa), before and after the Nile perch increase. Hydrobiologia 563: 3–44.
Goudswaard, K. P. C., E. F. B. Katunzi & F. Witte, 2011. Distribution of Nile perch Lates niloticus in southern Lake Victoria is determined by depth and dissolved oxygen concentrations. African Journal of Aquatic Science 36: 147–153.
Hecky, R. E., 1993. The eutrophication of Lake Victoria. Verhandlungen der internationalen Vereinigung für Limnologie 25: 39–48.
Hecky, R. E. & R. H. Hesslein, 1995. Contributions of benthic algae to lake food webs as revealed by stable isotope analysis. Journal of the North American Benthological Society 14: 631–653.
Hesslein, R. H., K. A. Hallard & P. Ramlal, 1993. Replacement of sulfur, carbon, and nitrogen in tissue of growing broad whitefish (Coregonus nasus) in response to a change in diet traced by δ34S, δ13C, and δ15N. Canadian Journal of Fisheries and Aquatic Science 50: 20170–22076.
Hughes, N. F., 1986. Changes in the feeding biology of the Nile perch, Lates niloticus, (L). (Pisces: Centropomidae), in Lake Victoria, East Africa since its introduction in 1960, and its impact on the native fish community of the Nyanza Gulf. Journal of Fish Biology 29: 541–548.
Hyndes, G. A., C. E. Hanson & M. A. Vanderklift, 2013. The magnitude of spatial and temporal variation in d15N and d13C differs between taxonomic groups: implications for food web studies. Estuarine, Coastal and Shelf Science 119: 176–187.
Jackson, A. L., R. Inger, A. C. Parnell & S. Bearhop, 2011. Comparing isotopic niche widths among and within communities: SIBER – Stable Isotope Bayesian Ellipses in R. Journal of Animal Ecology 80: 595–602.
Jackson, M. C., I. Donohue, A. L. Jackson, J. R. Britton, D. M. Harper & J. Grey, 2012. Population-level metrics of trophic structure based on stable isotopes and their application to invasion ecology. PLoS ONE 7: 1–12.
Kaehler, S. & E. A. Pakhomov, 2001. Effects of storage and preservation on the δ13C and δ15N signatures of selected marine organisms. Marine Ecology Progress Series 219: 299–304.
Katunzi, E. F. B., W. L. T. van Densen, J. H. Wanink & F. Witte, 2006. Spatial and seasonal patterns in the feeding habits of juvenile (Lates niloticus, L.), in the Mwanza Gulf of Lake Victoria. Hydrobiologia 568: 121–133.
Katunzi, E. F. B., J. Zoutendijk, T. Goldschmidt, J. H. Wanink & F. Witte, 2003. Lost zooplanktivorous cichlid from Lake Victoria reappears with a new trade. Ecology of Freshwater Fish 12: 237–240.
Kelly, B., J. B. Dempson & M. Power, 2006. The effects of preservation on fish tissue stable isotope signatures. Journal of Fish Biology 69: 1595–1611.
Keough, J. R., M. E. Sierszen & C. A. Hagley, 1996. Analysis of a Lake Superior coastal food web with stable isotope techniques. Limnology & Oceanography 41: 136–146.
Kilham, S. S., M. Hunte-Brown, P. Verburg, C. Pringle, M. R. Whiles, K. R. Lips & E. Zandona, 2009. Challenges for interpreting stable isotope fractionation of carbon and nitrogen in tropical aquatic ecosystems. Verhandlungen der internationale Vereinigung für Limnologie 30: 749–753.
Kishe-Machumu, M. A., F. Witte & J. H. Wanink, 2008. Dietary shift in benthivorous cichlids after the ecological changes in Lake Victoria. Animal Biology 58: 401–417.
Kishe-Machumu, M. A., F. Witte, J. H. Wanink & E. F. B. Katunzi, 2012. The diet of Nile perch, Lates niloticus (L.) after resurgence of haplochromine cichlids in the Mwanza Gulf of Lake Victoria. Hydrobiologia 682: 111–119.
Kolding, J., P. A. M. van Zwieten, O. C. Mkumbo, G. Silsbe & R. E. Hecky, 2008. Are the Lake Victoria fisheries threatened by exploitation or eutrophication? Towards an ecosystem-based approach to management. In Bianchi, G. & H. R. Skjoldal (eds), The Ecosystem Approach to Fisheries. ICP, New York: 309–354.
Layman, C. A., D. A. Arrington, C. G. Montana & D. M. Post, 2007. Can stable isotope ratios provide for community-wide measures of trophic structure? Ecology 88: 42–48.
Layman, C. A., M. S. Araujo, R. Boucek, C. M. Hammerschlag-Peyer, E. Harrison, Z. R. Jud, P. Matich, A. E. Rosenblatt, J. J. Vaudo, L. A. Yeager, D. M. Post & S. Bearhop, 2012. Applying stable isotopes to examine food-web structure: an overview of analytical tools. Biological Reviews 87: 545–562.
Matsuishi, T., L. Muhoozi, O. C. Mkumbo, Y. Budewa, M. Njiru, A. Asila, A. Othina & I. G. Cowx, 2006. Are the exploitation pressures on the Nile perch fisheries resources of Lake Victoria a cause for concern? Fisheries Management and Ecology 13: 53–71.
Mbabazi, D., B. Makanga, F. Orach-Meza, R. E. Hecky, J. S. Balirwa, R. Ogutu-Ohwayo, P. Verburg, L. Chapman & E. Muhumuza, 2010. Intra-lake stable isotope ratio variation in selected fish species and their possible carbon sources in Lake Kyoga (Uganda): implications for aquatic food web studies. African Journal of Ecology 48: 667–675.
McCutchan, J. H., W. M. Lewis, C. Kendall & C. C. Mcgrath, 2003. Variation in trophic shift for stable isotope ratios of carbon, nitrogen, and sulfur. OIKOS 102: 378–390.
Mkumbo, O. C. & W. Ligtvoet, 1992. Changes in the diet of Nile perch, Lates niloticus (L.), in the Mwanza Gulf. Lake Victoria. Hydrobiologia 232: 79–83.
Mugidde, R., 2001. Nutrient status and planktonic nitrogen fixation in Lake Victoria. PhD Thesis, University of Waterloo, Waterloo, Canada.
National Centers for Environmental Information (NOAA-NCEI). Retrieved from http://www1.ncdc.noaa.gov/pub/orders/CDO9535004896519.txt, viewed on May 10, 2011.
Ngupula, G. W. & E. Mlaponi, 2010. Changes in abundance of Nile shrimp, Caridina nilotica (Roux) following the decline of Nile perch and recovery of native haplochromine fishes, Lake Victoria, Tanzanian waters. Aquatic Ecosystem Health & Management 13: 196–202.
Njiru, M., J. B. Okeyo-Owuor, M. Muchiri & I. G. Cowx, 2004. Shifts in the food of Nile tilapia, (Oreochromis niloticus, L.) in Lake Victoria, Kenya. African Journal of Ecology 42: 163–170.
Njiru, M., E. Waithaka, M. Muchiri, M. van Der Knaap & I. G. Cowx, 2005. Exotic introductions to the fishery of Lake Victoria: What are the management options? Lakes & Reservoirs: Research and Management 10: 147–155.
Ogutu-Ohwayo, R., 1990. The decline of the native fishes of lakes Victoria and Kyoga (East Africa) and the impact of introduced species, especially the Nile perch, Lates niloticus, and the Nile tilapia, Oreochromis niloticus. Environmental Biology of Fishes 27: 81–96.
Ojwang, W. O., J. E. Ojuok, D. Mbabazi & L. Kaufman, 2010. Ubiquitous omnivory, functional redundancy and the resiliency of Lake Victoria fish community. Aquatic Ecosystem Health & Management 13: 269–276.
O’Neill, B. J. & J. H. Thorp, 2014. Untangling food-web structure in an ephemeral ecosystem. Freshwater Biology 59: 1462–1473.
Parnell, A., R. Inger, S. Bearhop & A.L. Jackson, 2008. SIAR: stable isotope analysis in R. Retrieved from: http://Cran.R-Project.Org/Web/Package/Siar.
Peterson, B. J. & B. Fry, 1987. Stable isotopes in ecosystem studies. Annual Review of Ecology and Systematics 18: 293–320.
Post, D. M., 2002. Using stable isotopes to estimate trophic position: models, methods, and assumptions. Ecology 83: 703–718.
Post, D. M., C. A. Layman, D. A. Arrington, G. Takimoto, J. Quattrochi & C. G. Montaña, 2007. Getting to the fat of the matter: models, methods and assumptions for dealing with lipids in stable isotope analyses. Oecologia 152: 179–189.
Schindler, D. E., S. R. Carpenter, J. J. Cole, J. F. Kitchell & M. L. Pace, 1997. Influence of food web structure on carbon exchange between lakes and the atmosphere. Science 277: 248–251.
Schriever, T. A., 2015. Food webs in relation to variation in the environment and species assemblage: a multivariate approach. PLoS ONE 10: e0122719.
Silsbe, G.M., 2004. Phytoplankton production in Lake Victoria, East Africa. MSc Thesis, University of Waterloo, Waterloo.
Syvärante, J., S. Vesala, M. Rask, J. Ruuhijrviärvi & R. I. Jones, 2008. Evaluating the utility of stable isotope analyses of archived freshwater sample materials. Hydrobiologia 600: 121–130.
Syvärante, J., A. Martino, D. Kopp, R. Céréghino & F. Cantoul, 2011. Freezing and chemical preservatives alter the stable isotope values of carbon and nitrogen of the Asiatic clam (Corbicula fluminea). Hydrobiologia 658: 383–388.
Talling, J. F., 1966. The annual cycle of stratification and phytoplankton growth in Lake Victoria (East Africa). Internationale Revue der gesamten Hydrobiologie 51: 545–621.
Thormar, J., H. Hasler-Sheetal, S. Baden, C. Boström, K. K. Clausen, D. Krause-Jensen, B. Oleson, J. R. Rasmussen, C. J. Svensson & M. Holmer, 2016. Eelgrass (Zostera marina) food web structure in different environmental settings. PLoS ONE 11: e0146479.
Vadeboncoeur, Y., E. Jeppesen, J. M. Vander Zanden, H. H. Schierup, K. Christoffersen & D. M. Lodge, 2003. From Greenland to green lakes: cultural eutrophication and the loss of benthic pathways in lakes. Limnology & Oceanography 48: 1408–1418.
van Alphen, J. J. M., O. Seehausen & F. Galis, 2003. Speciation and radiation in african haplochromine cichlids. In Dieckmann, U., J. A. J. Metz, M. Doebeli & D. Tautz (eds), Adaptive Speciation. Cambridge University Press, Cambridge: 171–189.
Wanink, J.H., 1998. The pelagic cyprinid Rastrineobola argentea as a crucial link in the disrupted ecosystem of Lake Victoria. Dwarfs and giants - African Adventures. PhD Thesis, Leiden University, Wageningen.
Wanink, J. H., 1999. Prospects for the fishery on the small pelagic Rastrineobola argentea in Lake Victoria. Hydrobiologia 407: 183–189.
Wanink, J. H. & F. Witte, 2000. The use of perturbation as a natural experiment: effects of predator introduction on the community structure of zooplanktivorous fish in Lake Victoria. Advances in Ecological Research 31: 553–570.
Wanink, J. H. & J. C. A. Joordens, 2007. Dietary shifts in Brycinus sadleri (Pisces: Characidae) from southern Lake Victoria. Aquatic Ecosystem Health & Management 10: 392–397.
Witte, F. & W. L. T. van Densen, 1995. Fish stocks and fisheries of Lake Victoria; a handbook for field observations. Samara Publishing Limited, Cardigan.
Witte, F., T. Goldschmidt, J. H. Wanink, M. van Oijen, K. Goudswaard, E. Witte-Maas & N. Bouton, 1992a. The destruction of an endemic species flock quantitative data on the decline of the Haplochromine cichlids of Lake Victoria. Environmental Biology of Fishes 34: 1–28.
Witte, F., T. Goldschmidt, P. C. Goudswaard, W. Ligtvoet, M. J. P. van Oijen & J. H. Wanink, 1992b. Species extinction and concomitant ecological changes in Lake Victoria, Netherlands. Journal of Zoology 42: 214–232.
Witte, F., B. S. Msuku, J. H. Wanink, O. Seehausen, E. F. B. Katunzi, P. C. Goudswaard & T. Goldschmidt, 2000. Recovery of cichlid species in Lake Victoria: an examination of factors leading to differential extinction. Reviews in Fish Biology and Fisheries 10: 233–241.
Witte, F., J. H. Wanink, M. Kishe-Machumu, O. C. Mkumbo, P. C. Goudswaard & O. Seehausen, 2007. Differential decline and recovery of haplochromine trophic groups in the Mwanza Gulf of Lake Victoria. Aquatic Ecosystem Health and Management 10: 416–433.
Xu, D., Y. Cai, H. Jiang, X. Wu, X. Leng & S. An, 2016. Variations of food web Structure and energy availability of shallow lake with long-term eutrophication: a case study from Lake Taihu, China. CLEAN-Soil, Air, Water 44: 1306–1314.
Zeug, S. C. & K. O. Winemiller, 2008. Evidence supporting the importance of terrestrial carbon in a large-river food web. Ecology 89: 1733–1743.
Acknowledgements
This work is part of the integrated project “Exploitation or eutrophication as threats for fisheries? Disentangling social and ecological drivers of ecosystem changes in Lake Victoria (SEDEC)”, supported by the Netherlands Organisation for Scientific Research (NWO/WOTRO) grant number W01.65.304.00. Thanks to Hans Hoogveld (NIOO) for help with plankton classification. Special thanks to Klaas Gerber, Alex Mangassa, Gerardi Rajabu, Paulo and Emma for their valuable field assistance and support, and to the Tanzanian Fisheries Research Institute (TAFIRI) in Mwanza for providing laboratory space and use of field equipment during this research.
Author information
Authors and Affiliations
Corresponding author
Additional information
Handling editor: Odd Terje Sandlund
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.
About this article

Cite this article
Cornelissen, I.J.M., Vijverberg, J., van den Beld, A.M. et al. Heterogeneity in food-web interactions of fish in the Mwanza Gulf, Lake Victoria: a quantitative stable isotope study. Hydrobiologia 805, 113–130 (2018). https://doi.org/10.1007/s10750-017-3297-x
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10750-017-3297-x