
Abstract
The accuracy of fertiliser recommendations can be improved by knowledge of use efficiency of applied nutrients. The aim of this work was to determine the efficiency with which mature oil palm (Elaeis guineensis Jacq.) recovers and uses fertiliser K and Mg under different environmental conditions in Sumatra, Indonesia. The work was carried out in seven factorial fertiliser trials (NPKMg) at sites with contrasting soils, slope and rainfall (1500–3500 mm annual average). We recorded the palms’ fresh fruit bunch yield (weekly), vegetative growth and nutrient content (annually), and soil properties (once during the study) over three years. Nutrient content of the above-ground palm components was calculated by multiplying concentration by biomass. Palm responses to K and Mg application were analysed at non-limiting values of the other nutrients. Fresh fruit bunch yield was significantly increased by application of K in five of the seven sites (by 4.8–11.9 t ha−1) and by Mg in two of the seven sites (by 7.2–12.3 t ha−1). Application of these fertilisers also significantly increased growth and nutrient content in most situations. Nutrient use efficiency declined with increasing yield and nutrient uptake. In the five sites with most positive yield responses to K, agronomic efficiency of K and Mg differed between sites due to differences in recovery efficiency. Recovery efficiency was correlated positively to soil K, Mg and silt content and rainfall, and negatively to slope and soil moisture deficit. In contrast, physiological efficiency of K and Mg use was similar across sites, despite differences in palm age and planting density. It thus appeared that physiological efficiency was determined by genotype and that variation in agronomic efficiency was mostly a result of differences in recovery efficiency.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Like all crops, the profitable and sustainable production of oil palm relies on rational nutrient management. To maximise profitability of oil palm fruit production, the palms usually need to be supplied with some major nutrients via fertilisers. These often include potassium (K) and magnesium (Mg), which are commonly poorly supplied in tropical soils. Nutrient requirements change as the crop passes through the stages of nursery, immaturity and maturity over a productive life of 25–30 years. Fertiliser requirements for the first stages (nursery and immaturity) are relatively small. However, as fruit bunch production requires a large quantity of nutrients (Ng and Thamboo 1967; Prabowo et al. 2006), fertiliser requirements will be large throughout the mature phase.
The type and amount of fertiliser required varies greatly from site to site. In Southeast Asia, where most commercial oil palm is grown, soils vary from fertile to marginal (Pushparajah 1982; Lubis and Adiwiganda 1996) and on many soils yield responds significantly to fertilisers (BLRS 1992–2009). The variation in palm responses is also affected by climatic factors such as rainfall, which ranges from less than 1500 to more than 4000 mm per annum. As fertiliser application makes up a large proportion (~ 50%) of the fruit production costs (Tohiruddin et al. 2006), appropriate fertiliser recommendations are crucial for maximising commercial gain. Failure to estimate palms’ fertiliser requirement accurately may result in insufficient or excessive nutrient supply, which greatly affects profitability. Unsustainable soil nutrient mining or water pollution could be a direct impact of inappropriate fertiliser recommendations on the surrounding environment.
Foster (2003) compared different methods of assessing oil palm fertiliser requirements and concluded that the diagnosis of nutrient status through frond analysis is essential and as a first approximation can be used on its own for making recommendations. However, knowledge of the efficiency of yield response to fertilisers at the site is also needed to accurately determine the fertilisers required to maximise profitability and to correct the nutritional status determined from frond analysis. Frond analysis is widely used to diagnose nutrient deficiencies and guide fertiliser applications via a range of approaches (Ollagnier et al. 1970; Caliman 1994; Foster 2002; Foster and Prabowo 2002; Goh 2011) and the more developed methods are applicable to most regions (Foster 2003). Initially only leaflet nutrient concentrations were used to diagnose nutrient status of the palm, but more recently concentrations in the rachis have also been shown to be important and are now routinely used together with leaflet concentrations (Teoh and Chew 1988a; Foster and Prabowo 2006).
Recommendations of fertiliser type and amount are challenges since trials show that the amount required for palms to reach an adequate nutrient status usually differs between locations. Therefore, the amount of fruit produced per unit of fertiliser applied, or ‘agronomic efficiency’ (AE) also varies. Agronomic efficiency is the product of ‘uptake or recovery efficiency’ (RE), which is the increase in amount of fertiliser-derived nutrient taken up by palms per unit increase in the amount applied, and ‘utilization or physiological efficiency’ (PE), which is the increase in fruit bunch yield per unit increase in nutrient uptake (Fairhurst 1999). In some situations with annual crops, using PE on its own to compare varieties or species can provide a misleading measure of nutrient utilization efficiency (Siddiqi and Glass 1981). However, in the case of a mature perennial crop such as oil palm, which is in a relatively steady state of annual canopy turnover and fruit production, PE provides a robust measure for comparing sites and varieties. Agronomic efficiency was shown to be related to environmental conditions in Peninsular Malaysia based on the analysis of 48 oil palm fertiliser trials (Foster et al. 1985). However, it was emphasised that the findings are only applicable in the trial regions so cannot be extrapolated to regions where the soils and climate are different.
Knowing how AE and its components (RE and PE) differ between sites is essential for making accurate fertiliser recommendations but measuring them is difficult and time-consuming. The question as to which of the two components plays larger role in determining AE is also crucial to answer. If AE is related to readily observed site characteristics (e.g., soil nutrient content, texture, rainfall, soil moisture deficit or slope), then it could be estimated at any particular site using those characteristics. However, such relationships are not yet known for oil palm in Sumatra. Therefore, the aim of this work was to measure RE and PE for important nutrients viz. K and Mg in NPKMg fertiliser trials across seven contrasting sites in Sumatra and relate it to the characteristics of the sites.
Materials and methods
Overall procedure and field experiment design
To determine nutrient (K and Mg) use efficiency, fresh fruit bunch (FFB) yield and nutrient uptake were measured in several contrasting environments (Table 1), each with fertilisers applied at a range of rates. General measurements (yield, growth and nutrient content) were taken over a three-year period and averaged to obtain values on an annual basis. However, specific measurements for the efficiency study, such as above-ground (leaf, petiole, frond, trunk) dry matter, and nutrient content analysis of trunks and bunches were done only once. Agronomic efficiency was calculated as the amount of FFB yield per unit of nutrient applied. Recovery efficiency was calculated as the increase in amount of nutrient taken up by the above-ground parts of the palm per unit increase in the amount of nutrient applied as fertiliser (Fairhurst 2003). It does not take into account the recycling of nutrients in pruned leaves and male inflorescences. Annual nutrient uptake in above-ground parts of the palms was calculated as the sum of annual uptake by fruit bunches, male inflorescences, fronds (leaflets, rachises and petioles) and trunk, with each component being the product of annual biomass production and its nutrient concentration. The petiole bases remaining attached to the trunk after frond pruning were not included in the calculation. Total annual uptake does not equal the annual increase in above-ground stock of nutrients because of loss or removal of palm parts. The stock of nutrients in fronds, fruit bunches and male inflorescences remains fairly constant over time in mature palms, with the rate of growth being balanced by the rate of loss. Pruned leaves and male inflorescences are recycled in-situ whilst fruit bunches are harvested when ripe and exported. Physiological efficiency was calculated as the increase in FFB yield per unit increase in nutrient uptake.
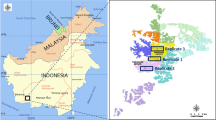
This work was carried out in seven factorial (N, P, K and Mg) fertiliser trials in mature oil palm plantations in North Sumatra (NS, Trials 231, 232, 275 and 277) and South Sumatra (SS, Trials 1403, 1411 and 1412) provinces, Indonesia (Table 1). Soil and climate differed considerably between sites. The trial sites had acidic soils with low cation exchange capacity (CEC) formed on rhyolite (Trials 231, 275 and 277), sandstone (Trial 232) and dacite and claystone (Trials 1403, 1411 and 1412). The annual rainfall ranged from 1571 to 3484 mm. Trials in NS were planted in 1985, replacing rubber (except Trial 232, which replaced forest) whereas the SS trials were planted in 1996–1997, replacing forest. The trial areas ranged from 13 to 25 ha each. The trials had 36 plots (Trials 275, 277, 1411, 1412 and 1413) or 81 plots (Trials 231 and 232). Plot size in NS trials was 49 palms (7 × 7), of which the outer 40 palms were treated as double guard rows and the inner 9 palms (3 × 3) were recorded for yield, growth and nutrient content. Being on contour terrace (40% slope), plots in Trial 232 included three terraces, with 12 palms on each (the outer two palms on each side were guard palms and not recorded). Palms on the upper terrace functioned as a guard row and were not recorded. The SS trial plots were 64 palms (8 × 8) in size, with 48 double guard row palms leaving inner 16 recorded palms (4 × 4). In addition to the double guard row palms, trenches (0.5 m wide, 1.0 m deep) were installed around each individual plot to prevent possible nutrient poaching from neighbouring plots (except for Trial 232 on contour terraces). In all cases the palms were mature Tenera (Deli Dura x AVROS Pisifera). Fertiliser treatments (urea and muriate of potash at all sites, triple super phosphate and kieserite in NS, and rock phosphate and dolomite in SS) were in place for 4–12 years prior to the measurement period (Table 2). The treatments were applied in all factorial combinations of the application rates. Units of measurement for AE, RE and PE were kg (mass or weight), ha (area) and year (time).
Palm measurements and calculations of nutrient use efficiency
Since yield response to nutrient application and nutrient use efficiency decline as yield increases, and both depend on the supply of other nutrients, these factors were taken into account so that the sites could be compared fairly. To compare AE and RE at the same yield level, quadratic equations were fitted to the relationship between yield or nutrient uptake and nutrient application rate, and from them the application rate at given levels of yield attained in all trials was derived. As AE and RE may also depend on the supply of other nutrients, the yield and uptake responses to K and Mg were calculated at optimum (non-limiting) levels of the other applied nutrients (Gourley et al. 1994). For example, if the highest yield in Trial 231 was obtained at the NPKMg fertiliser rates of 2222 (i.e. the highest rates of all those elements, being 1.8 kg N + 0.8 kg P + 2.49 kg K + 0.51 kg Mg palm−1, see Table 2), then to determine the effect of K at optimal levels of the other nutrients, we compared yields in the plots with NPKMg rates of 2202, 2212 and 2222.
Annual FFB yield was recorded on an individual palm basis and analysed on a plot basis. Nutrient uptake was calculated by multiplying annual FFB production by nutrient concentration. FFB nutrient content was measured once during the 2002–2004 period, on all plots. To obtain a sample for nutrient analysis, fresh bunches were separated into their components (stalk, spikelet, staminode, mesocarp, shell, kernel, trash) before drying. The dried samples were then mixed together according to proportions determined in a preliminary investigation of 206 fruit bunches collected in different trials. The average dry matter content of a bunch was found to be 59.8%. The dry matter content of bunch components ranged from 21 to 82%, with the shell and kernel having the highest values. The average dry weight of bunches comprised of 67.3% mesocarp, 10.7% spikelet, 9.6% kernel, 6.2% shell, 3.3% staminode and 2.8% stalk. Number of male inflorescences (MI) was counted in all trial plots. One MI sample was randomly taken from each trial plot for fresh and dry weights. Fresh weight of a MI ranged from 2.6 to 4.4 kg with a moisture content of 74%. Spikelets made up 91% of the dry weight and stalks 9%. Using this dry weight proportion, the spikelets and stalk was mixed for nutrient analysis.
Dry weight of palm fronds and their components could be predicted reliably (R2 = 0.91) from the area of the petiole cross section (pcs) (Fig. 1). These data were derived from 335 fronds from Tenera palms of 14 different ages (4–22 years after field planting) in Sumatra. Dry weight of leaflets, rachis and petiole were 38%, 39% and 23%, respectively, of the total frond weight. In this work, our dry weight prediction equations (Fig. 1) were used because they provided a better fit for the advanced planting material than previously published models. As a comparison, a previous estimate of frond dry weight based on pcs (R2 = 0.94) (Corley et al. 1971) conducted on Dura palms was slightly overestimating A recent allometric relationship (Aholoukpe et al. 2013) estimated frond biomass using rachis dry weight (R2 = 0.62), which is less simple than using pcs.

Allometric relationships developed to estimate dry biomass of palm components: a frond, b leaflet, c rachis, d rachis + petiole from petiole cross section (pcs), and e dry trunk density from palm age
Annual frond biomass production was calculated as frond production rate multiplied by individual frond weight, and nutrient uptake was calculated by multiplying annual frond biomass production by nutrient concentration. Frond no.17 from all plots was sampled and analysed annually for nutrient concentration (leaflets and rachises analysed separately). Petiole and rachis were assumed to have the same K and Mg concentration, as analyses from 5 fertiliser trials in Malaysia showed no consistent difference in K concentrations between them (Teoh and Chew 1988b) and our unpublished results showed a similar lack of difference for Mg concentrations. The dry weight of fronds and their components (leaflets, rachis, petiole) was estimated by making non-destructive measurements and relating them to the weight using the allometric relationships developed earlier (Prabowo et al. 2002, 2006).
The annual increment in trunk weight was determined using height increase, diameter and density and nutrient uptake was calculated by multiplying annual trunk weight increment by nutrient concentration. Trunk nutrient content was measured once during the 2002–2004 period, on all plots. Trunk samples were taken from the axil of frond no.41, using a corer that penetrated 20 cm deep. Trunk density was estimated (R2 = 0.37) using the relationship between palm age and density (Fig. 1) based on destructive sampling of 88 palms of eight different ages (6–30 years after field planting) (Prabowo et al. 2002, 2006). A trunk portion (after removing the frond butts) within the upper 1 m below frond no.41 was cut (10 cm thick) after felling and its volume and oven-dried weight were measured.
Soil analysis and correlation of efficiency values with site characteristics
Soil samples were taken at 0.0–0.4 m depth from between the palm circle and frond heaps of 4–6 central palms in the unfertilised control plot of each trial. The samples were mixed thoroughly for a composite sample of each trial (Table 1). Soils were analysed for texture by the pipette method and exchangeable cations using 1 M ammonium acetate. Concentration of K was also measured using concentrated HCl boiled at 100 °C (Singh and Ratnasingam 1971; Rengel and Damon 2008). This method gives much higher concentrations than 1 M ammonium acetate extracts and reflects the long-term reserve. Slope was measured to represent the whole trial area. Five years data of rainfall and moisture deficit was obtained from manual rain gauge stations within 3 km of the trial sites.
Bullet 2.4 Statistical analysis
Effects of the treatments on FFB and nutrient uptake by various components of the palms (Table S1) were analysed as detailed in Note S5. Following the analysis, the significance of the difference between the Control (with no added K and Mg fertiliser) and each other treatment level was determined by least significance difference (LSD) tests (Table S4). Each site was analysed separately and only one time was analysed (the experimental period). The soil and climatic properties were correlated with yield, uptake and nutrient use efficiency data using linear regression and Pearson’s correlation coefficient. IBM SPSS 20 software was used for statistical data analysis. P values for the analyses are presented and we use the term ‘significantly’ where p < 0.05.
Results
Effects of fertilisers on FFB yield and uptake of K and Mg
FFB yield and nutrient uptake in different palm components (trunks, leaflets, rachises, male inflorescences and bunches) were affected by K and Mg application in most of the trials (Tables S1 and S4). At the first fertiliser increment (0–1), the fertiliser effects were highly significant (at p = 0.010 and 0.001 levels) (Table S2). The effect of K fertiliser was greater than that of Mg.
Application of K significantly increased FFB yield in five of the seven trials and increased K uptake in the main palm components in all trials. However, K fertiliser decreased Mg uptake in the rachis and leaf in five trials (Table S1). Mg fertiliser significantly increased FFB yields in only two trials (and decreased it in one trial) and increased Mg uptake in all palm components in most trials. The greater the uptake of K and Mg in the unfertilised palms, the lower the yield response was to application of those nutrients (Fig. 2).

Maximum FFB yield responses, i.e., difference between maximum value of yield reached in the trial and yield in the Control plots with no added K and Mg fertilisers, in relation to a K and b Mg uptake, and in relation to c soil K and d soil Mg content (in unfertilised control fertiliser plots)
The cation uptake was affected by N application but not by P application in most trials (Table S1). Nitrogen fertiliser increased yield in five trials and increased K uptake in the rachises, leaflets and bunches in most trials whilst P fertiliser significantly increased yield in four trials. The effects of Mg and P fertilisers on K uptake and of N and P fertilisers on Mg uptake were mostly not significant. Interactions between treatments were generally not significant or not consistent (Table S4) but taken together they had an appreciable effect on yield and nutrient uptake. Generally, interactions were due to the deficiency of one nutrient limiting the response to another nutrient, which gives a false indication of the potential response to the latter nutrient.
Agronomic efficiency (AE) of K and Mg
Without fertiliser, FFB yield was greater in the South than in the North Sumatra trials except the non-responsive Trials 275 and 277 (Table 3). Agronomic efficiency decreased with increasing fertiliser rate and yield, as seen by the decreasing slope of the lines in Fig. 3 with increasing fertiliser rate. Agronomic efficiency was similar for both nutrients for most trials at a given rate of fertiliser (see slope of lines in Fig. 3 at a given fertiliser rate) but differed substantially between trials when compared at a given rate of yield (values given in Table 4).

FFB yield response to a K and b Mg fertiliser in all trials, calculated at non-limiting levels of other nutrients
Agronomic efficiency of K was 12–50 kg FFB kg−1 K (at a yield of 27,000 kg ha−1 a−1), being highest in Trials 1403 and 1411, lowest in Trials 232, 275 and 277 and intermediate in Trials 231 and 1412. Agronomic efficiency of Mg ranged from 29 kg FFB kg−1 Mg in Trial 232 to 202 kg FFB kg−1 Mg in Trial 231 and was negative in all the other trials. In most cases, the fertiliser requirement to reach a given level of yield was larger in sites with poorer AE-K or AE-Mg except for the non-responsive trials (Table 4). The relationship between yield response and site nutrient status was closer for K than Mg.
Recovery efficiency (RE) of K and Mg
The amount of K and Mg taken up, and the recovery efficiency of applied K and Mg, differed substantially between sites (Fig. 4). In contrast to the FFB yield, in the absence of fertiliser total nutrient uptake was lower in South than North Sumatra trials except the non-responsive Trials 275 and 277 (Table 3). RE-K ranged from 46 to 148% (at the same FFB yield level of 27,000 kg ha−1 a−1). As with the yield response, K and Mg recovery tended to decline with nutrient uptake level, so to compare RE fairly, the values were compared at the same level of yield (Table 5). Compared in this way, RE-K was highest in Trial 1403, lowest in Trial 232 and intermediate in Trials 1411 and 1412. Of the trials with a positive yield response to Mg, at the same level of yield (27,000 kg FFB ha−1 a−1), RE-Mg was highest in Trial 231 (111%) and lowest in Trial 232 (14%) (Table 5). Recovery efficiency was even lower in the trials in which yield did not respond positively to Mg fertiliser.

Uptake by palms of a K in relation to K applied and b Mg in relation to Mg applied in all trials, calculated at non-limiting levels of other nutrients
Physiological efficiency (PE) of K and Mg
Physiological efficiency of both K and Mg was similar in most trials (Fig. 5), except for K in Trial 1411, which had a much higher efficiency. Comparison of the regression lines showed significant differences (p = 0.0063) if Trial 1411 was included. However, the remaining four trials (Trials 231, 232, 1403 and 1412) displayed statistically parallel linear curves (p = 0.4496).

FFB yield in relation to a K and b Mg uptake in all trials, calculated at non-limiting levels of other nutrients. As for Table 6, PE is calculated from the equations (for example, PE-K for Trial 231 is 33.618 kg FFB kg−1 K uptake)
Unlike recovery efficiency, there was no change in PE with yield or fertiliser application rate. In contrast to RE, the PE was much lower for K (17–64 kg FFB kg−1 K in all the seven trial sites) than for Mg (172–187 kg FFB kg−1 Mg in two sites). PE-K was very similar and ranked “medium” for Trials 231 (34 kg FFB kg−1 K), 232 (39 kg FFB kg−1 K), 1403 (30 kg FFB kg−1 K) and 1412 (26 kg FFB kg−1 K), but higher for Trial 1411 (64 kg FFB kg−1 K). PE-Mg for Mg-responsive Trials 231 and 232 was also similar viz. 172 and 187 kg FFB kg−1 Mg, respectively.
Effects of environmental factors on FFB yield and uptake of K and Mg
The response of yield, nutrient uptake and RE to fertiliser application was significantly correlated with several environmental variables. The maximum yield response, i.e., yield difference between level 0 with either level 1 or 2, to applied K and Mg was negatively correlated with uptake of the corresponding nutrients (Fig. 2). The uptake was large in trials not responding to K (Trials 275 and 277) or to Mg (Trials 275, 277, 1403, 1411 and 1412). Yield response also declined with increasing soil nutrient concentrations in the unfertilised control plots. Soil silt content and water availability significantly affected nutrient use efficiency (Fig. 6).

Effects of environmental factors on the efficiency of response to K and uptake of the cationic nutrient (calculated at FFB yield = 27,000 kg ha−1 a−1)
Of the soil properties measured (Table 1), yield response to K was most closely related to hot-acid extractable K with a large response occurring at values of less than 1.5 cmol kg−1. The soil exchangeable K in all these sites was not correlated to yield response. The two trials that did not significantly respond to K fertiliser (275 and 277) had extractable K levels of more than 4 cmol kg−1 soil (Fig. 2). The only two trials showing a significant response to Mg fertiliser (231 and 232) were distinguished by having soil exchangeable Mg values of 0.12 cmol kg−1 or lower. The other 5 trials had soil exchangeable Mg values of 0.12 cmol kg−1 or above and most showed a significant decline in yield response to application of Mg fertiliser (Fig. 2).
Agronomic efficiency was significantly correlated with several environmental variables for K but not for Mg (Fig. 6). AE-K was 5 times higher at sites with high rainfall (p = 0.0305) and low soil moisture deficit (p = 0.0124) than at drier sites with moisture deficit of more than 200 mm a−1. Linear regression of AE-K showed that R2 increased from 0.73 (rainfall only) to 0.77 with the slope effect added (p = 0.1134). The trial on sloping topography (232) showed even smaller AE-K. Of the soil parameters, silt content was highly correlated with K recovery (R2 = 0.89) (Fig. 6).
Discussion
Recovery efficiency differed between sites, causing variation in agronomic efficiency
Comparison of the yield response and nutrient uptake efficiency at the same yield level in trials responsive to K or Mg (Table 6) showed that RE of both nutrients varied more than PE. Physiological efficiency was similar for all trials except Trial 1411. We have not identified any site property that might account for the much higher PE in this trial, so the reason remains unknown. In fact, this trial site had similar properties to Trial 1412, which had the lowest PE. The general lack of variation in PE was likely due to the genetic similarity of the palms in the trials. Fairhurst (1999) suggested several factors that are likely to affect PE viz. planting material, density, frond pruning, harvesting, pests and diseases. Planting density and palm age appeared to have little effect on PE over the ranges covered in this study (density 110 to 143 ha−1 and age 7–21 years). FFB yield for a given K uptake was higher for the younger palms planted on ex-forest in South Sumatra than the older palms planted mostly on ex-rubber in North Sumatra. This difference in FFB yield was therefore attributed to a difference in palm age and previous land use rather than a difference in the other environmental characteristics.
Previous studies have found strong interactions between oil palm genotype and leaflet nutrient concentrations, nutrient uptake and yield response to fertiliser (Tan and Rajaratnam 1978; Kushairi et al. 2001; Donough et al. 1996; Cochard and Tristan 2005; Corley and Tinker 2016; Ollivier et al. 2017; Dassou et al. 2022), with genetic variation in K and Mg uptake and utilization contributed by Dura and Tenera parents (Ollivier et al. 2017). Genotypic differences in AE of K and Mg in oil palm appear to be due to differences in RE and PE (Ollivier et al. 2017). Genotypic differences in nutrient use efficiency of oil palm are well worth exploring further as they might lead to better fertiliser use recommendations (Dassou et al. 2022) and may be exploited in breeding programs. For example, the improvement in wheat yields in Mexico due to breeding over 1950–1980 (Epstein and Bloom 2005) or in maize (Ciampitti and Vyn 2012) was associated with an increase in PE of nutrient use (Good et al. 2004; Yang et al. 2010; Canizella et al. 2014). Improved PE-K can be achieved through various mechanisms such as better K translocation into different plant components observed at the molecular level (Very and Sentenac 2003) or greater capacity to substitute Na+ for K+ (Clark 1983; Rengel and Damon 2008). Variations in PE-K between genotypes have also been attributed to differences in biomass and K partitioning patterns (Wang and Chen 2012) and the size of the root system or its ability to solubilise soil K (Steingrobe and Claassen 2000).
As PE was similar between sites, variation in AE was therefore mostly caused by RE, which was affected by inherent site properties. Differences in RE due to soil and climatic variables are likely to be responsible for the well-known site-specific nature of fertiliser responses (Foster and Prabowo 2003; Corley and Tinker 2016). FFB yield responses to fertilisers may vary up to 270% between sites (Goh et al. 1994; Kee et al. 1994).
Across the seven trials, the factor most closely related to RE-K was the soil silt content (Table 1, R2 = 0.89). The two trials with a high K uptake efficiency can be clearly distinguished by their high soil silt content of 28% (Trial 231) and 35% (Trial 1403) compared with 7–18% in the other five trials (Fig. 6). The high RE-K in these two trials was presumably due to a large root system, high specific absorption capacity of roots, greater availability of K for uptake within the root zone (e.g., due to a longer period of moisture) or a combination of these factors. Soil silt content may improve the recovery of K fertiliser because it is associated with the soil K buffer capacity (Foster and Chang 1989). Thus, the higher the soil silt content the higher the capacity of the soil to retain applied K in the root zone against leaching losses. The association between RE-K and slope was presumably due to loss of applied nutrients in runoff, and shallower soil and rooting depth resulting in greater susceptibility to moisture stress (Kee and Soh 2002).
The two trials with a significant yield response to Mg fertiliser (Trials 231 and 232) had very different RE-Mg values (Table 5). Trial 231 had RE-Mg more than 100% up to a yield level of 27,000 kg FFB ha−1 a−1, whereas at the same yield level Trial 232 had a RE-Mg of only 14%. The poor Mg recovery in Trial 232 might be attributed to its low soil silt content and steep slopes, the two factors found to limit K recovery, and high moisture deficit.
Climatic variables such as moisture deficit and rainfall were associated with AE-K (Fig. 6). The SS sites and Trial 231 sites had medium–high AE benefiting from the zero to very low moisture deficit and high rainfall (Table 1). It is worth noting that whilst increasing soil moisture increases K uptake by oil palm, excess drainage can have a negative effect on yield response (Tohiruddin et al. 2010) associated with loss of K by leaching (Ollagnier et al. 1987). In contrast, the other NS trials had low AE, presumably due to the high moisture deficit. This result was in line with an investigation of yield response of oil palm to K fertiliser in 20 trials on sedentary soils in Malaysia (Foster and Goh 1977) where annual rainfall was found to have a significant positive effect and soil slope a significant negative effect. Both variables add explanation to the low AE status of Trial 232 (Fig. 6) although the effect of slope was weakly significant (p = 0.1134; R2 = 0.77) perhaps because the trial was the only one with steep slope. There was no noticeable effect of previous land use on RE of K and Mg, although land use was confounded with soil parent material.
Nutrient efficiency knowledge can help finetune fertiliser recommendations and land resource assessment
In the early days of making fertiliser recommendations for oil palm, calculations were based on a nutrient balance sheet including the soil supply of nutrients indicated by soil analysis (Hew and Ng 1968). As a rough guide to fertiliser recommendation for a new site where oil palm is not yet grown at large scale, soil analysis data are worthwhile. Subsequently, FFB yield response to K fertiliser at different sites was found to be predicted more accurately by hot-acid extractable soil K than by the more commonly used exchangeable soil K analysis (Fig. 2). It was K from non-exchangeable clay fraction that became available for palm uptake (Foster and Prabowo 1996; Dubos and Ollivier 2011). Hot acid extractable K reflects the total reserve K including the large non-exchangeable portion fixed tightly by the soil. The value changes little within 12 months, which is the normal period for determining fertiliser recommendation for oil palm. Soil K buffering capacity is probably another factor stabilising plant-available K supply during short time periods (Wang et al. 2004). In addition, spatial variability in soil nutrient concentrations within a field is of concern for agronomists as it makes it difficult to obtain a representative value. Foster and Azhar (1978) concluded that soil analysis results were spatially too variable for accurate calculations of the nutrient balance sheet. However, that conclusion was based on exchangeable K and it is possible that hot acid-extractable K is less spatially variable.
The results of this research indicate that yield responses to K and Mg fertilisers are better predicted by palm uptake of K and Mg in the absence of these fertilisers than by soil analyses (Fig. 2). Therefore, diagnosis of K and Mg requirements of oil palm is more reliably achieved using foliar diagnosis than soil analysis. Later, when a plantation is established and regular annual frond sampling is feasible, palm uptake can be determined. However, if the task is only to determine current nutrient status, measurement of palm uptake is not necessary. Nutrient uptake data should be utilised to fine-tune the current fertiliser recommendation system to determine how much fertilisers to apply to fields with particular site characteristics.
The amount of fertiliser needed to increase yield can be calculated using AE information. If we take Table 4 and Fig. 2 as an example, to target a yield of 24,000 kg FFB ha−1 a−1 Site 231 requires only 76 kg K ha−1 whilst Site 232 needs more than double the amount (173 kg K ha−1). This is because Site 232 has low RE due to its steep slope, low soil silt content and high moisture deficit. At this yield level AE-K is 29 kg FFB kg−1 K applied. If the yield target is 27,000 kg FFB ha−1 a−1 then 319 kg K ha−1 is needed because AE-K is less at this higher yield target viz. 12 kg FFB kg−1 K. It is also noteworthy that this site will never achieve 30,000 kg FFB ha−1 a−1 because the yield response efficiency reaches 0 at 27,653 kg FFB ha−1 a−1. However, in Trial 231 it is still realistic to target up to 34,172 kg FFB ha−1 a−1. As a comparison the other trial sites have maximum FFB yields of 40,133 (Site 1403), 33,215 (Site 1411) and 31,706 kg FFB ha−1 a−1 (Site 1412).
In the case of Trial 277, the site is inherently high-yielding as shown by the high intercept (29,700 kg FFB ha−1 a−1 at 0 kg K ha−1) but the AE-K suggests that fertiliser application will increase yield by only 13 kg FFB kg−1 K fertiliser applied. Therefore, K fertiliser application would be less economically desirable at this site than the others.
The results of this work might improve land suitability evaluation of new areas or detailed characterization of planted fields, by enabling additional interpretations of data collected during soil surveys. Land suitability surveys (Paramananthan 1995; Lubis and Adiwiganda 1996) classify land into S-1 (highly suitable), S-2 (moderately suitable), S-3 (marginally suitable) or NS (not suitable). During survey, field assessments are made (soil type, slope, soil texture, drainage, flood risk, rainfall and number of dry months) and laboratory data collected (soil nutrient concentrations and cation exchange capacity). Our results show that soil texture (silt content), to rainfall and water deficit could be used to help estimate RE and AE of sites being assessed. These estimates of nutrient use efficiency could help in the grouping of fields for management purposes. The integration of information derived from trials (Prabowo et al. 2010), including nutrient use efficiency, with the usual soil survey parameters might also facilitate more accurate estimation of site yield potential of new areas or yield gap analysis of planted fields.
Conclusions
Variation in agronomic efficiency of K and Mg between sites was caused by differences in recovery efficiency of K and Mg, which were strongly correlated with soil silt content. Other site properties directly related to agronomic efficiency included rainfall, moisture deficit and slope. On the other hand, physiological efficiency of both K and Mg were relatively unaffected by site characteristics and were most likely determined by plant genotype. These conclusions are more robust for K than for Mg, due to the higher number of sites with a response to K fertiliser than Mg fertiliser. Knowledge about the uptake of K and Mg, and yield response efficiency to K and Mg fertilisers could be used to increase accuracy of current fertiliser recommendation systems. In addition, the relationship between nutrient efficiency and commonly measured site characteristics such as soil silt content, slope, rainfall and water deficit could be utilised to improve accuracy of land assessment for plantation establishment and management, and inform yield gap analysis in existing plantations.
References
Aholoukpe H, Dubos B, Flori A, Deleporte P, Amadji G, Chotte JL, Blavet D (2013) Estimating aboveground biomass of oil palm: allometric equations for estimating frond biomass. For Ecol Manage 292:122–129. https://doi.org/10.1016/j.foreco.2012.11.027
BLRS (1992–2009) Annual reports, Vol. I, PT. PP. London Sumatra Indonesia
Caliman JP, Daniel C, Tailliez B (1994) Oil palm mineral nutrition. Plant Rech Dev 1:36–54
Canizella BT, Moreira A, Moraes LAC, Fageria NK (2015) Efficiency of magnesium use by common bean varieties regarding yield, physiological components, and nutritional status of plants. Commun Soil Sci Plant Anal 46(11):1376–1390
Ciampitti IA, Vyn TJ (2012) Physiological perspectives of changes over time in maize yield dependency on nitrogen uptake and associated nitrogen efficiencies: a review. Field Crop Res 133(2012):48–67. https://doi.org/10.1016/j.fcr.2012.03.008
Clark RB (1983) Plant genotype differences in the uptake, translocation, accumulation, and use of mineral elements required for plant growth. In: Sarić MR, Loughman BC (eds) Genetic Aspects of Plant Nutrition. Springer Netherlands, Dordrecht, pp 49–70
Cochard B, Tristan DG (2005) Oil palm genetic improvement and sustainable development. OCL Oilseeds Fats Crops Lipids 12(2):141–147. https://doi.org/10.1051/ocl.2005.0141
Corley RHV, Hardon JJ, Tan GY (1971) Analysis of growth of the oil palm (Elaeis guineensis Jacq.). I. Estimation of growth parameters and application in breeding. Euphytica 20:307–315
Corley RHV,Tinker PB (2016) The Oil Palm. Fifth edition. World Agricultural Series. Wiley Blackwell
Dassou OS, Bonneau X, Aholoukpe H, Vanhove W, Ollivier J, Peprah S, Flori A, Duran-Gaselin T, Mensah AG, Sinsin BA, Van Damme P (2022) Oil palm (Elaeis guineensis Jacq.) genetic differences in mineral nutrition: potassium and magnesium effects on morphological characteristics of four oil palm progenies in Nigeria (West Africa). OCL. https://doi.org/10.1051/ocl/2022024
Donough CR, Corley RHV, Law IH, Ng M (1996) First results from an oil palm clone x fertiliser trial. Planter 72:69–87
Dubos B, Ollivier J (2011) Potassium uptake and storage in oil palm organs: the role of chlorine and the influence of soil characteristics in the Magdalena valley. Colomb Nutr Cycl Agroecosyst 89(2):219–227. https://doi.org/10.1007/s10705-010-9389-x
Epstein E, Bloom J (2005) In: Mineral nutrition of plants: principles and perspective, Second Edition. Sinauer Associates, Inc., Sunderland, Massachusetts. Crop Yields and Nutrient Efficiency, pp 260–265
Fairhurst, T (1999) Nutrient use efficiency in oil palm: Measurement and management, ISP Central Johore Branch
Fairhurst TH (2003) Efficient management of natural resources in oil palm plantations. In: 2003 international planters conference. ISP, Malaysia, 16–18 June 2003
Foster HL, Prabowo NE (1996) Variation in the potassium fertiliser requirements of oil palm in North Sumatra. In: Proceedings of 1996 PORIM International Palm Oil Congress–Agriculture, pp 143–152
Foster HL, Prabowo NE (2002) Overcoming the limitations of foliar diagnosis in oil palm. In: 2002 international oil palm conference and exhibition. IOPRI, Bali, Indonesia
Foster HL, Prabowo NE (2003) Efficient use of fertilisers in oil palm for increased productivity in North Sumatra. In: Proceedings of 2003 PIPOC international oil palm conference, Malaysian Palm Oil Board, Putrajaya
Foster HL, Prabowo NE (2006) Partition and transfer of nutrients in the reserve tissues and leaves of oil palm. Workshop on nutrient needs in oil palm–a dialogue among experts. Singapore, 17–18 October 2006
Foster HL, Azhar A (1978) Yield response of oil palm to fertilisers in West Malaysia. IV Soil nutrient levels. MARDI Res Bull 6(2):150–164
Foster HL, Chang KC (1989) Factors limiting maximum oil palm yields in Peninsular Malaysia. Oleagineux 44:1–7
Foster HL, Goh HS (1977) Yield response of oil palm to fertilisers in West Malaysia II. Influence of soil and climatic factors. MARDI Res Bull 5(1):6–22
Foster HL, Chang KC, Dolmat MT, Mohammed AT, Zakaria ZZ (1985) Oil palm yield responses to N and K fertilisers in different environments in Peninsular Malaysia. PORIM Occasional Paper No. 16
Foster HL (2003) Assessment of oil palm fertiliser requirements. In: Management for large and sustainable yields. In: Fairhurst TH and Hardter R (eds.) Potash and Phosphate Institute, pp 231–257
Goh KJ, Chew PS, Teo CB (1994) Maximising and maintaining oil palm yields on commercial scale in Malaysia. 1995 Incorporated Societies of Planters Conference, Malaysia
Goh KJ (2011) Fertiliser recommendation systems for oil palm: estimating the fertiliser rates. In: agronomic principles & practices of oil palm cultivation (eds. Goh KJ, Chiu SB & Paramananthan S)
Good AG, Shrawat AK, Muench DG (2004) Can less yield more? Is reducing nutrient input into the environment compatible with maintaining crop production? Trends Plant Sci 9(12):597–605. https://doi.org/10.1016/j.tplants.2004.10.008
Gourley CJP, Allan DL, Russelle MP (1994) Plant nutrient efficiency: a comparison of definitions and suggested improvement. Plant Soil 158(1):29–37
Hew CK, Ng SK (1968) A general schedule for manuring oil palm in West Malaysia. Planters 44:417–429
Kee KK, Soh AC (2002) Management of oil palms on slope lands in Malaysia. In: 2002 international oil palm conference, Nusa Dua, Bali, Indonesia, July 8–12, 2002
Kee KK, Goh KJ, Chew PS, Tey SH (1994) An integrated site-specific fertiliser recommendation system (INFERS) for high productivity in mature oil palms.
Kushairi A, Rajanaidu N, Jalani BS (2001) Response of oil palm progenies to different fertiliser rates. J Oil Palm Res 13:84–96
Lubis AU, Adiwiganda R (1996) Agronomic management practices of oil palm plantation in Indonesia based on land conditions. In: Proceedings of ISOPA/IOPRI Seminar, Pekanbaru, 1996
Ng SK, Thamboo S (1967) Nutrient contents of oil palms in Malaya. I. Nutrients required for reproduction: fruit bunches and male inflorescences. Malaysian Agric J 46(1):3–45
Ollagnier M, Ochs R, Martin G (1970) The manuring of the oil palm in the world. Fertilite 36:3–63
Ollagnier M, Daniel C, Fallavier P, Ochs R (1987) The influence of climate and soil on potassium critical level. In: Proceedings of 1987 PORIM oil palm/palm oil conference–agriculture, pp 235–248
Ollivier J, Flori A, Cochard B, Amblard P, Turnbull N, Syahputra I, Suryana E, Lubis Z, Surya E, Sihombing E, Durand-Gaselin T (2017) Genetic variation in nutrient uptake and nutrient use efficiency of oil palm. J Plant Nut. https://doi.org/10.1080/01904167.2016.1262415
Paramananthan S (1995) Soil survey and management on tropical soils. Param Agricultural Surveys, Kuala Lumpur
Prabowo, NE, Tohiruddin L, Fairhurst T, Foster HL, Nasution EN (2002) Efficiency of fertiliser recovery by oil palm in Sumatra. In: international oil palm conference. Bali
Prabowo NE, Foster HL, Webb MJ (2006) Nutrient uptake and fertiliser recovery efficiency. Workshop on nutrient needs in oil palm–a dialogue among experts. Singapore, 17–18 October 2006
Prabowo NE, Tohiruddin L, Tandiono J, Foster HL (2010) Identification and evaluation of problem fields: an implementation of a yield prediction tool based on palm data. In: Proceedings of 2010 international oil palm conference, Jogjakarta, Indonesia
Pushparajah E (1982) Nitrogenous fertilisers for plantation crops with special reference to rubber and oil palm. In: Proceedings of the seminar on nitrogen in Malaysian agriculture. Malaysian societies of soil science, Kuala Lumpur. Pp 123–146
Rengel Z, Damon PM (2008) Crops and genotypes differ in efficiency of potassium uptake and use. Physiol Plant. https://doi.org/10.1111/j.1399-3054.2008.01079.x
Singh MM, Ratnasingham K (1971) Manual of laboratory methods of chemical soil analysis. Rubber Research Institute of Malaysia, Kuala Lumpur
Steingrobe B, Claassen N (2000) Potassium dynamics in the rhizosphere and K efficiency of crops. J Plant Nutr Soil Sci. https://doi.org/10.1002/(SICI)1522-2624(200002)163:1%3c101::AID-JPLN101%3e3.0.CO;2-J
Tan GY, Rajaratnam JA (1978) Genetic variability of leaf nutrient concentration in oil palm. Crop Sci 18:548–550
Teoh KC and Chew PS (1988a) Use of rachis Analysis as an indicator of K nutrient status in oil palm. In: Proceedings of 1987 international oil palm/ palm oil conferences, progress & prospects. Kuala Lumpur, Malaysia
Teoh K.C. & Chew P.S. (1988b) Potassium in the oil palm eco‐system and some implications to manuring practice. In: Proceedings of 1987 international oil palm/ palm oil conferences, progress & prospects. Kuala Lumpur, Malaysia
Tohiruddin L, Prabowo NE, Foster HL (2006) Comparison of the response of oil palm fertilisers at different locations in North and South Sumatra. In: International oil palm conference, Bali, Indonesia
Tohiruddin L, Prabowo NE, Tandiono J, Foster HL (2010) A comprehensive approach to the determination of optimal N and K fertiliser recommendations for oil palm in Sumatra. In: Proceedings of 2010 international oil palm conference, indonesian oil palm research institute, Jogjakarta, Indonesia
Very AA, Sentenac H (2003) Molecular mechanisms and regulation of K+ transport in higher plants. The EMBO J. https://doi.org/10.1146/annurev.arplant.54.031902.134831
Wang L, Chen F (2012) Genotypic variation of potassium uptake and use efficiency in cotton (Gossypium hirsutum). J Plant Nutr Soil Sci. https://doi.org/10.1002/jpln.201100199
Wang JJ, Harrell DL, Bell PF (2004) Potassium buffering characteristics of three soils low in exchangeable potassium. Soil Sci Soc Am J 68(2):654–661. https://doi.org/10.2136/sssaj2004.6540
Yaeesh Siddiqi M, Glass ADM (1981) Utilization index: a modified approach to the estimation and comparison of nutrient utilization efficiency in plants. J Plant Nut 4(3):289–302. https://doi.org/10.1080/01904168109362919
Yang F, Wang G, Zhiyong Zhang A, Eneji E, Duan L, Li Z, Tian X (2010) Genotypic variations in potassium uptake and utilization in cotton. J Plant Nutr 34(1):83–97. https://doi.org/10.1080/01904167.2011.531361
Acknowledgements
We would like to thank Agronomy staff of BLRS, PT. London Sumatra Indonesia Tbk.—Tohirudin Lubis, Joko Tandiono and Mahadani Lubis—who helped with field trial recordings and data collection in North and South Sumatra. Special thanks to the statistician, Evi Nafisah who assisted with statistical data analysis of the trials. The work was funded by PT. London Sumatra Indonesia and we express our appreciation to the Director of Research, Dr. Stephen Nelson, the President Director, Mr. Benny Tjoeng and all members of the Board of Directors for the support and permission to publish the research work.
Funding
Open Access funding enabled and organized by CAUL and its Member Institutions.
Author information
Authors and Affiliations
Contributions
All authors reviewed the manuscript
Corresponding author
Ethics declarations
Conflict of interest
The authors have no financial or competing interests to declare.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Prabowo, N.E., Foster, H.L. & Nelson, P.N. Potassium and magnesium uptake and fertiliser use efficiency by oil palm at contrasting sites in Sumatra, Indonesia. Nutr Cycl Agroecosyst 126, 263–278 (2023). https://doi.org/10.1007/s10705-023-10289-7
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10705-023-10289-7