
Abstract
Legumes are important in sustainable agriculture and particularly so when they are intercropped with other species. In breeding programs, little attention is paid to their agronomic performance in species mixtures. In red clover, improved persistence is an important breeding goal. We identified traits associated with survival of red clover cultivated in pure stands (PS 3) or in mixtures with grasses (MS 3) and managed under a 3-cut system (two locations), as well as in pure stands in a 5-cut system (PS 5, one location). Survivors from replicate plots were collected and a new generation made from each plot. The new generations were characterized in a growth experiment with light or simulated shade, and in a freezing experiment. We show that the traits related to red clover persistence depend on both plant community composition and cutting frequency. MS 3 had more leaves with larger leaf blades and longer petioles during the vegetative stage, followed by earlier stem elongation, higher number of elongating stems, higher biomass (also when accounting for earlier stem elongation) and more leaves in the regrowth after cutting than PS 3. MS 3 also had better freezing tolerance. PS 5 was similar to MS 3 and different from PS 3 in the number of leaves, leaf blade size, petiole length and number of elongating stems. These results show that breeding and cultivar evaluation, which is currently almost exclusively considering performance in pure stands, may miss some variation which provides persistence of red clover in mixtures with grasses.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Legumes (species belonging to the family Fabaceae) are indispensable for the transition to an environmentally and economically sustainable agri-food system (Rubiales et al. 2021). This is due to their symbiosis with nitrogen-fixing bacteria, which strongly reduces the need for application of synthetic N fertilizer, and thus reduces energy-demanding fertilizer production. Although nitrogen losses from legume residues can be a problem (Sturite et al. 2021), losses are generally lower when legumes rather than synthetic fertilizers are the source of nitrogen in cropping systems (Jensen et al. 2012). The nitrogen fixed by legumes is not only utilized by the legumes themselves, but also by non-fixing species like grasses and cereals, which are either cultivated together with or follow the legume crop. Moreover, legumes are rich in protein, a limited resource in many parts of the world (de Visser et al. 2014; Gale et al. 2015). Compared to crops of the grass family (Poaceae), the efforts to produce genetically improved legume varieties through plant breeding has been limited so far (Rubiales et al. 2021). However, the current focus on sustainable agriculture, protein crops and plant-based food has stirred an increased interest from both farmers and plant breeding companies. At the same time there is an increasing awareness of the need to diversify agricultural systems, both in order to utilize resources better and in order to improve resilience in the face of increasingly variable climatic conditions associated with climate change.
Red clover (Trifolium pratense L.) is an important grassland forage crop in temperate regions of the world, mostly used in rotations with annual crops or in regularly re-sown swards, where it provides benefits such as high yields, symbiotic N fixation, good soil structure and high-quality animal feed (Frame et al. 1998; Abberton and Marshall 2005; Lüscher et al. 2014; Phelan et al. 2015). As a forage crop, red clover is mostly cultivated in mixtures with at least one grass species. The persistence of red clover is often lower than that of its companion species (e.g. Ergon et al. 2016; Brophy et al. 2017), and increasing persistence is therefore an important breeding goal (Abberton and Marshall 2005; Riday 2010; Annicchiarico et al. 2015). More persistent red clover would increase sustainability (less use of synthetic fertilizer and fossil fuel, less nutrient losses during frequent ploughing and re-sowing) as well as economic profits for the farmer (less operation costs, higher yields and better forage quality).
Persistence may be defined purely as survival over years, or alternatively, as maintenance of annual yield over years. These two measures can be correlated (e.g., Herrmann et al. 2008), but there may also be negative associations between harvestable biomass produced and survival during a subsequent stressful period, such as a winter (discussed further below). The limited persistence of red clover can be attributed to its life strategy as a short-lived perennial with no vegetative mode of reproduction, limited winter survival ability, susceptibility to fungal root-infecting pathogens and nematodes, as well as to competition from grasses, which is high in sown swards with moderate to high levels of N fertilization (Abberton and Marshall 2005; Taylor 2008; Boller et al. 2010; Ӧhberg 2008; Annicchiarico et al. 2015). In regions with cold winters, red clover populations in sown swards tend to be reduced over winter a few years after sowing, and persistence is therefore strongly associated with winter survival. Winter survival may either be directly due to mechanisms of tolerance to various abiotic and biotic winter stress factors (e.g. freezing, low-temperature pathogenic fungi, ice cover, water-saturated soil, lack of light) or more indirectly, to conditions during the growing season, late fall or early spring, leading to e.g. insufficient cold acclimation, too early de-acclimation, or a general weakening of plants due to pathogens, defoliation or competition (Rapacz et al. 2014). Studies of different genetic materials have revealed a trade-off between shoot biomass potential and winter survival in red clover (Annicchiarico and Pagnotta 2012; Loucks et al. 2018; Zanotto et al. 2021). Strong competition for light and corresponding shade avoidance responses, as well as shoot regrowth after repeated defoliation, involves allocation of resources to the aerial parts of the shoot and may exhaust organic reserves and reduce the winter survival ability (Vik 1955; Frankow-Lindberg et al. 1997; Donaghy and Fulkerson 1998; Bélanger et al. 2006). Disease resistance, competitiveness, and the ability to regrow and survive after repeated defoliation is likely to be related to a range of additional traits, e.g. specific disease resistance mechanisms (Lager and Gerhardson 2002), vegetative growth habit (prostrate vs. erect) (Ford and Barrett 2011) and other architectural traits (Van Minnebruggen et al. 2014). Given this complex nature of persistence, expression of genetic variation and correlation with other traits is highly dependent on the environmental conditions and management that the material is exposed to. For instance, when grown as single plants, traits related to competitivity are not relevant for persistence, while in dense stands they are.
Mixtures of forage species tend to outperform pure stands of individual species in terms of biomass yield (e.g. Kirwan et al. 2007). In spite of this, almost all breeding and cultivar evaluation is conducted using pure stands (PS). However, genotypes with optimal performance in PS are not necessarily those with the best performance in mixed stands (MS), and researchers have therefore asked whether selection in MS would be better (reviewed by Litrico and Violle 2015; Annicchiarico et al. 2019). In sown grassland swards, only a fraction of the sown plants survives the first years due to competition and stress. For example, in pure red clover stands sown at a rate of 18 kg ha−1, 40, 27 and 18% of the number of plants expected to germinate according to a germination test, had survived by the end of the first (establishment year), second and third growing season, respectively (Marley et al. 2003). This “natural selection” is used by breeders aiming for improved persistence, but it can also be utilised to identify traits that provide persistence under different environmental conditions and managements, e.g. PS and MS. Studies of such survivor populations of white clover, poly-crossed separately and then compared to the original populations in a common environment, have demonstrated that genetic shifts occur in presumably adaptive traits after just a few years in the field (Collins et al. 2001; Frankow-Lindberg 2001; Helgadóttir et al. 2001). In a previous study at Ås, Norway, we have shown that red clover winter survival differed between plots subjected to different managements. During the winter after the second production year, winter mortality was more severe in PS harvested 3 times per year (PS 3) than in MS harvested 3 times per year (MS 3) (see Fig. 7 in Ergon et al. 2016). Better winter survival in mixtures is commonly observed, but it is not known if it is due to an effect of plant community composition on winter stress factors or on the winter survival ability of red clover. In the field experiment referred to above, we also showed (with molecular markers) that already prior to this third winter, there were detectable shifts in the genetic composition of the subpopulations in the plots as compared to the originally sown population, and that subpopulations in MS plots differed from subpopulations in PS plots, indicating divergent selection (Ergon et al. 2019). In the present study we characterize this divergent selection in terms of phenotypic traits, recorded in offspring of survivor populations collected from this experiment after the third production year (Fig. 1), as well as survivor populations after the third production year in another experiment at a different location (Stjørdal, Norway). The aim was to test if there is differential selection in red clover growing in PS vs. MS that can be detected phenotypically in the offspring, and to identify traits that are favourable for survival in either type of stand. We also tested for differential selection in PS 5 vs. PS 3 (at Ås only). Our hypotheses were as follows:
-
1.
Plants with early stem elongation compete better for light with grasses (which elongate earlier in the season than red clover). MS survivor populations therefore start stem elongation earlier than PS survivor populations.
-
2.
Plants with long stems, large shoots and strong shade avoidance responses compete well for light in stands with a high amount of standing biomass. Survivor populations from PS 3 and MS 3 therefore develop longer stems, larger shoots and respond more to shade than survivor populations from PS 5.
-
3.
At Ås PS 3 went through a stronger selection during the last strong winter than MS 3. This resulted in survivor populations with higher freezing tolerance in PS 3 than in MS 3.
-
4.
Plants with many nodes and branches close to the ground and rapid regrowth after cutting tolerate frequent cutting and compete well in the regrowth. Survivor populations from PS 5 therefore have more nodes and branches at the plant base and more rapid regrowth than survivor populations from PS 3.

Three of the red clover sub-populations used in the study, one year after sowing (top) and one month before collecting survivors. Survivors were crossed within sub-population to make a new generation, which was phenotyped under controlled conditions. MS 3, mixed stand and 3 cuts per year; PS 3, pure stand and 3 cuts per year; PS 5, pure stand and five cuts per year. Note that PS 5 was photographed almost a month earlier than MS 3 and PS 3 in May 2011
Material and methods
Plant material
Red clover (Trifolium pratense L., diploid cv. Lea, Graminor, Norway) survivors were collected from experimental plots that had been established in 2010 at two locations: Stjørdal (63°30'N, 10°51'E) and Ås (59°40’N, 10°47’E). The plots were either pure stands (PS) or mixed stands (MS), sown at a total rate of 20 (Ås), 15 (PS at Stjørdal) or 24 (MS at Stjørdal) kg ha−1. At Ås the MS plots consisted of perennial ryegrass (cv. Fagerlin, Graminor), tall fescue (cv. Kora, DLF-Trifolium, Denmark), white clover (cv. Milkanova, DLF-Trifolium) and red clover (each species constituted 25% of the total seed weight sown). At Stjørdal MS plots consisted of timothy (cv. Grindstad, Tollef Chr. Grindstad, Norway, 54%), meadow fescue (cv. Fure, Graminor, 25%) and red clover (21%). The plots were harvested from 2011 to 2013. PS and MS plots were harvested 3 times per year at both locations (PS 3 and MS 3) and at Ås there was an additional treatment with PS plots harvested five times per year (PS 5). Three cuts per year is the normal harvesting frequency of grassland crops for cattle at Ås, while in Stjørdal both two and three cuts are common. Annual total yields and red clover yields are provided in Online Resource 1. Plots were fertilized with a compound fertilizer at a total rate of 100 (Ås) and 130 (mixtures, Stjørdal) kg N ha−1 year−1. PS at Stjørdal were applied a PK-fertilizer without N. In October 2013 sixty survivors were collected from each of two replicate plots in Ås while forty survivors were collected from each of three replicate plots at Stjørdal. The survivors were planted in large pots and kept in a greenhouse at Ås over the winter at a cool temperature (minimum 2 °C). Prior to flowering the following year survivors from each plot, considered a population here, were covered with a large net and one bumble bee nest with 50 bees (Pollinering AS, Bryne, Norway) was placed inside each net. After maturation seeds were collected from each population. Together with two replicate samples of seeds from the seed lot originally sown at Ås, this resulted in a total of 14 subpopulations belonging to six different population types (Stjørdal: PS 3, MS 3, Ås: original population, PS 3, MS 3 and PS 5).
Phenotypic characterization
For characterization of growth and development, seeds were sown in a peat soil and germinated at 16 °C and 20 h photoperiod (metal halide lamps, approximately 110 µmol m−2 s−1 photosynthetically active radiation (PAR)). After 2 weeks, individual young seedlings were transplanted into peat soil in 10 cm × 10 cm pots, grown under the same conditions and watered and fertilized regularly with a fertilizer containing 12% N, 4% P, 18% K and micronutrients. There were 32 seedlings of each population, organized into 32 blocks, with one seedling per population in each block (but two seedlings of the original Ås population). During the first 4 weeks after pricking, pots were placed at a density of approximately 90 pots per m2 under 240 μmol m−2 s−1 PAR and a red light to far red light ratio (R:FR) of 2.25 (referred to as the normal light treatment). Six weeks after sowing, when the first plants showed signs of stem elongation, half of the blocks were moved to a shade treatment with 100 μmol m−2 s−1 PAR and a R:FR of 1.15 (referred to as the shade treatment), while the rest of the plants remained. From then on the pot density were approximately 45 pots per m2 in both light treatments. Traits related to growth and development were recorded weekly (see Online Resource 2 for details). The number of days from sowing until stem elongation was recorded and plants that did not elongate during the experiment were given the maximum value + 1 = 76. Twelve weeks after sowing, the plants were cut at 5 cm height and the dry weight recorded. All plants were then placed in the normal light treatment at a pot density of 90 pots per m2 for regrowth. After 9 days, the number of leaves and the number of stems with no bud outgrowth were recorded for each plant, and after 2 weeks of regrowth the plants were again cut at 5 cm height, and the dry weight recorded.
For characterization of susceptibility to freezing, seeds were sown in a peat soil in early January and germinated in a greenhouse (59°40’ N, 10°47’ E) at 16 °C with natural light supplied with additional metal halide lamps (approximately 110 µmol m−2 s−1 PAR) to a photoperiod of 12 h. After germination, individual young seedlings were transplanted into individual pots with peat soil, organized as 96 replicate blocks with one seedling per population in each block (but two seedlings of the original Ås population), and grown under the same conditions. When the plants were 6 weeks old they were moved to cold acclimating conditions at 3–4 °C, 12 h photoperiod and 110 µmol m−2 s−1 PAR, for 2 weeks. Thirty-two blocks were placed in each of three replicate programmed freezing chambers initially set at 2 °C. The temperature was first lowered from 2 °C to − 3 °C at 1 °C h−1 and kept at this level for 12 h to ensure even freezing, after which the temperature was lowered again by 1 °C h−1. When the temperature reached one of the test temperatures, − 7 °C, − 9 °C, − 11 °C and − 13 °C, eight blocks were removed from each freezing chamber and allowed to thaw at 2 °C. After thawing, plants were placed back in the greenhouse and kept there for 3 weeks, before survival of each plant was recorded.
Statistical analyses
Populations from the two field locations were analysed separately, using data from each individual and not population averages. Analysis of variance of data from the growth experiment was performed using PROC MIXED in SAS Enterprise 7.1 (SAS Institute Inc., Cary, NC, USA). Light treatment was included as a fixed factor from the time point when it was introduced into the experiment and replicate block was then nested in light treatment as a random effect factor. The number of days to elongation was also included as a fixed effect co-factor in the analyses of traits that were recorded from then on, except for leaf traits and number of branches from the base. In all analyses, two-way interactions among fixed factors where initially included and removed in a step-wise manner if not significant (P > 0.05). The significance of pairwise differences among population types at Ås were tested with a Tukey test option in PROC MIXED. Correlations among traits were tested using PROC CORR in SAS. Binary mortality data from the freezing experiment were analysed with logistic regression using PROC LOGISTIC in SAS, using freezing temperature, replicate freezing chamber and population type as factors.
Results
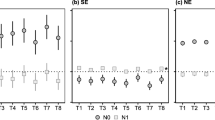
Differences were seen between population types from Ås during the first weeks. PS 3 plants were significantly smaller with fewer branches and fewer leaves with shorter middle leaflets and shorter petioles than plants from MS 3 and PS 5 (Table 1a, b, Fig. 2, P < 0.05 according to Tukey pairwise comparison tests). Plants from the original population were intermediate. The populations from Stjørdal both had large plants, and although the differences were similar, the distinction between PS 3 and MS 3 plants was less clear and significant for length of petiole and middle leaflet at some time points only (Table 1a, Fig. 2).

Characteristics of the studied red clover survivor populations during early growth. Plants were grown under “normal” light conditions in controlled conditions, and half the plants were transferred to a simulated shade treatment after 6 weeks (w). After 6 weeks the averages of the two light treatments are shown. Original, the original population sown at Ås; PS 3, pure stand harvested 3 times a year; PS 5, pure stand harvested 5 times a year (Ås only); MS 3, mixed stand harvested 3 times a year. See Table 1a and b for statistical analysis
The first signs of stem elongation were seen in some individuals around 6 weeks after sowing, but population averages for days to elongation ranged from 59 to 70 days. MS 3 populations from Stjørdal had earlier onset of stem elongation and a higher number of elongating stems, internodes per stem and branches on elongating stems than PS 3 populations (Table 1b, Fig. 3, 4). There was a similar but smaller and not significant difference in days to elongation and number of elongating stems between PS 3 and MS 3 at Ås.

Characteristics of the studied red clover populations during stem elongation in the studied survivor populations. Averages across two light treatments are shown. Original, the original population sown at Ås; PS 3, pure stand harvested 3 times a year; PS 5, pure stand harvested 5 times a year (Ås only); MS 3, mixed stand harvested 3 times a year. See Table 1b for statistical analysis

Timing of stem elongation and characteristics measured at harvest and after regrowth in the studied red clover survivor populations from Ås and Stjørdal. Averages of 2 (Ås) or 3 (Stjørdal) replicate survivor populations per population type are shown together with standard errors. Original, the original population sown at Ås; PS 3, pure stand harvested 3 times a year; PS 5, pure stand harvested 5 times a year (Ås only); MS 3, mixed stand harvested 3 times a year. See Table 1b for statistical analysis
Late stem elongation (high number of days to elongation) was negatively correlated with biomass in the normal light treatment at 12 weeks after sowing (R = − 0.68, P < 0.0001 and R = − 0.59, P < 0.0001, in the light and shade treatment, respectively), and positively correlated with biomass of the regrowth (R = 0.69, P < 0.0001 and R = 0.71, P < 0.0001 in the light and shade treatments, respectively). MS 3 populations had significantly higher biomass above 5 cm at 12 weeks than PS 3 populations at both locations (Table 1b, Fig. 4, P < 0.05 according to Tukey pairwise comparison test for Ås), also when accounting for the positive effect of early stem elongation on biomass in the statistical model. MS 3 from Stjørdal also had a higher number of nodes below 5 cm and a larger number of leaves in the regrowth after cutting than the PS 3 populations. There was a similar but smaller and not significant difference between PS 3 and MS 3 in the number of leaves in the regrowth at Ås.
Light treatment had a very strong effect on many of the traits (Table 1). Plants in the shade treatment had longer petioles and internodes, fewer branches from the base, fewer elongating branches and branches on stems, shorter stems, lower biomass above 5 cm height and fewer nodes below 5 cm height than plants in the normal light treatment (Online Resource 3). After cutting and regrowth under normal light conditions, plants from the shade treatment had significantly lower biomass, fewer leaves and more stems with no bud regrowth. The timing of stem elongation was not significantly affected by shading, and neither was the number of internodes. No significant interaction between light treatment and population type on any of the traits could be detected.
Almost all plants survived freezing at -7 and -9 °C and data from these two temperatures were therefore not included in the analysis. Among Stjørdal populations, PS 3 populations survived freezing better than MS 3 populations (Table 1b, Fig. 5). This tendency was seen at Ås also.

Mortality of the studied red clover survivor populations after freezing, expressed as the proportion of tested plants that survived. Averages of 2 (Ås) or 3 (Stjørdal) replicate survivor populations per population type are shown together with standard errors. Original, the original population sown at Ås; PS 3, pure stand harvested 3 times a year; PS 5, pure stand harvested 5 times a year (Ås only); MS 3, mixed stand harvested 3 times a year. See Table 1c for statistical analysis
Discussion
Rapid growth and development appear to be more advantageous for long-term survival in mixtures with grasses than in pure stands
Plants of MS 3 survivor populations started stem elongation earlier than those of PS 3 survivor populations. Thus hypothesis 1 was supported. Additionally, MS 3 populations had generally faster development than PS 3 populations both before and during the stem elongation phase, resulting in larger shoot size. A similar difference was seen after cutting, with a higher number of leaves in the regrowth of MS 3 populations than of PS 3 populations. We did not observe any significant interaction between population type and light treatment on any of the recorded traits, suggesting that the more rapid growth and development of MS 3 was largely constitutive, and not depending on shading by neighbouring plants in order to be expressed.
Rapid growth could possibly provide the ability to compete for light during establishment and regrowth when growing next to grasses. Similarly, genotypes with early stem elongation could be expected to compete better with grasses, which enter the stem elongation stage earlier than red clover. Hoekstra et al. (2018), comparing several Swiss Mattenklee and Ackerklee cultivars grown in sward plots together with perennial ryegrass in the Netherlands, also found positive associations between persistence (measured both as plant density and as persistence of yield in the third production year), stem length and shoot biomass. As the cultivars were not tested in pure stands in parallel, it is not known from their results to which extent this association is dependent on stand type or not. Here we have shown that a more rapid increase of shoot size (corrected for earliness of stem elongation), independently of shading, appears to provide a stronger selective advantage and better persistence in MS 3 than in PS 3. But – does red clover really suffer from competition with grasses? Red clover is frequently not inferior to grasses during the first 2 years, particularly if N fertilization is limited. This was for example the case in our studied MS 3 stands, where the harvested red clover biomass increased by 35 and 83% from year 1 to year 2 at Ås and Stjørdal, respectively, as compared to 4 and 18% increase of the total harvested biomass (Online Resource 1, see also Ergon et al. (2016)). The increase in red clover biomass yield of the PS 3 stands also increased, but to a lesser extent (26 and 36% from year 1 to year 2 at Ås and Stjørdal, respectively, as compared to 29 and 51% increase of the total harvested biomass). This suggests that red clover was a stronger competitor than the companion species during the two first years, which could be expected to give shoot size a stronger competitive advantage in PS 3 than in MS 3 (according to our hypothesis 2). We obtained the opposite result when we collected survivors after year 3, thus hypothesis 2 was rejected. This may be related to the strong selection during the last winter, which is discussed further in the section below.
High tolerance to freezing appears to be more advantageous for long-term survival in pure stands than in mixtures with grasses
PS 3 populations tended to be more freezing tolerant (lower mortality) than MS 3 populations at both locations. Thus, hypothesis 3 was supported. The last winter was strong at both locations, leading to a strong decline in red clover proportion in year 3 (Online Resource 1), particularly at Ås, where a very high winter mortality was also documented, particularly in PS 3 (see Introduction and Ergon et al. 2016). A much lower mortality in the MS 3 and PS 5 populations suggests that the high mortality in PS 3 could be related to both stand type and harvesting regime. As red clover usually produces more biomass in late summer than grasses, we can speculate that the higher mortality in PS 3 was a result of more allocation of resources to the shoot in order to compete in a dense red clover stand in late summer (due to shade avoidance responses and/or genotypic selection), leading to physiologically weakened plants with insufficient organic reserves for winter survival. In a study of Nordic gene bank material, rapid petiole growth and potential shoot size was associated with lower winter survival and lower freezing tolerance (Zanotto et al. 2021). Thus, the last winter may have selected individuals with better freezing tolerance, and indirectly also slower growth and smaller shoot size. This illustrates that care must be taken in the interpretation of the results. A trait that differs between population types may do so simply because it is correlated with a trait that gives a selective advantage. This may occur due to physiological associations, genetic linkage or population structure. Moreover, the time point for selection of survivors may be crucial for the results; sampling of survivors prior the last winter may have yielded very different results. Verwimp et al. (2018) have earlier shown that the genetic composition of perennial ryegrass, another perennial forage species, can vary strongly from year to year.
Rapid growth and development appear to be advantageous under high defoliation frequency
We could not detect a significantly higher number of nodes and branches close to the ground or more rapid regrowth in PS 5 than in the other Ås populations. Hypotheses 4 was therefore not supported. Like MS 3, PS 5 had more rapid growth during the first weeks, with a higher number of leaves, larger leaves, longer petioles and more branches than PS 3. This suggests that more rapid growth provides a selective advantage in frequently cut plots, however, it may also be an indirect effect of lower winter mortality in PS 5 plots than in PS 3 plots, combined with a negative association between growth potential and winter survival ability as described in the section above.
Implications for plant breeding and evaluation of candidate cultivars
Although effects of species interactions (facilitation and competition) between forage legumes and forage grasses are well known, practical breeding and cultivar evaluation have paid little attention to genetic variation in the ability of species to coexist and yield over time without outcompeting each other. Although red clover is mostly cultivated in species mixtures, almost all breeding and cultivar evaluation is conducted using pure stands. There are good reasons for this; performance in mixtures depends not only on genetic variation in red clover and effects of environment and management, but also on variation in the other components of the mixture (species, cultivars, proportions, and interactions with environment, management and the red clover component), adding complexity to the breeding process and cultivar evaluation. It is therefore important to quantify the added value of evaluating genetic material in species mixtures and evaluate if it is large enough to warrant extra costs that it will incur. A range of options for incorporating mixture performance specifically in breeding goals are given by Litrico and Violle (2015), Annicchiarico et al. (2019) and Sampoux et al. (2020).
An ideotype approach can complement breeding programs by helping to prioritize among traits, as well as generate testable hypotheses about how changes in one trait could affect the overall performance of the plant or crop community (Tork et al. 2019). As more information is gained, an ideotype model describing the relationship between traits and performance in mixtures could be improved. We consider here improved persistence of red clover cultivated in mixtures with grasses in regions with long winters as a breeding goal. Based on the limited information gained in this study, a hypothetical ideotype would have rapid growth, early stem elongation and large shoot size. First of all, the hypothesis that these traits provide better persistence in mixtures relative to pure stands would have to be tested for relevant breeding material and across relevant environments. Persistence is already selected for in most breeding programs, but in pure stands, which, as shown here is not quite the same as persistence in mixtures. Therefore, selecting on the basis of persistence in mixtures, or alternatively, applying additional selection for traits that are particularly important for persistence in mixed stands, could possibly help achieving breeding goals faster. Early petiole and stem elongation could be easily scored in a large number of individuals under controlled conditions and selected individuals could be used for crossing.
In a recent study of a collection of Nordic red clover accessions (Zanotto et al. 2021), petiole length, shoot size and late stem elongation under controlled conditions were to some extent negatively correlated with winter survival and freezing tolerance. Also, time to stem elongation under controlled conditions (non-vernalized plants, 23 h photoperiod) did not correlate significantly with flowering date in the field, suggesting that either there is a genotype x environment effect due to these different testing environments, or initiation of stem elongation and initiation of flowering are differently regulated. Although we used more realistic temperatures and photoperiods in the present study, the association between early stem elongation and persistence in mixed stands should be tested under field conditions. Moreover, the material studied by Zanotto et al. (2021) consisted of mainly wild populations and landraces, which may have different association among traits than cultivated material. In any case it would be necessary to carefully monitor possible trade-off effects of selection of more rapid petiole elongation, earlier stem elongation and shoot size on winter survival. Grasses produce more biomass in early summer while red clover produce more later in the season, so another possible trade-off to consider is a reduced yield in mixtures during the first years after sowing due to a reduced temporal niche differentiation between red clover and grasses. If earlier stem elongation in the red clover component leads to less productivity later in the season, it could potentially reduce the differentiation in resource utilization that contributes to high yields in mixtures, an effect that should be weighed against the advantages of better persistence. Earlier stem elongation in red clover is, as in grasses, associated with a reduction in protein content and digestibility when harvesting time is kept constant (e.g. Nadeem et al. 2019), but on the other hand, a more synchronized growth and development between red clover and grasses would allow for better optimization of harvesting time points and ensure a more stable species composition throughout the growing season and better utilization of the complementarity between red clover and grasses on forage quality.
Conclusion
We have here shown that specific traits providing persistence to red clover depend on both plant community composition and cutting frequency. Rapid growth and development were associated with persistence in mixtures with grasses and in frequently harvested pure stands, while high freezing tolerance is associated with persistence in pure stands harvested less frequently. Our results indicate that breeding and cultivar evaluation should pay more attention to performance in mixtures with grasses in order to exploit the full potential of red clover in sustainable agriculture.
Data availability
Datasets used for statistical analyses, figures and tables presented in the current study are available in the NMBU Open Research Data repository, https://doi.org/10.18710/MRGWV
References
Abberton MT, Marshall AH (2005) Progress in breeding perennial clovers for temperate agriculture. J Agr Sci 143:117–135. https://doi.org/10.1017/s0021859605005101
Annicchiarico P, Pagnotta MA (2012) Agronomic value and adaptation across climatically contrasting environments of Italian red clover landraces and natural populations. Grass Forage Sci 67:597–605. https://doi.org/10.1111/j.1365-2494.2012.00887.x
Annicchiarico P, Barrett B, Brummer EC, Julier B, Marshall AH (2015) Achievements and challenges in improving temperate perennial forage legumes. Crit Rev Plant Sci 34:327–380. https://doi.org/10.1080/07352689.2014.898462
Annicchiarico P, Collins RP, De Ron AM, Firmat C, Litrico I, Hauggaard-Nielsen H (2019) Do we need specific breeding for legume-based mixtures? Adv Agron 157:141–215. https://doi.org/10.1016/bs.agron.2019.04.001
Bélanger G, Castonguay Y, Bertrand A, Dhont C, Rochette P, Couture L, Drapeau R, Mongrain D, Chalifour F-P, Michaud R (2006) Winter damage to perennial forage crops in eastern Canada: causes, mitigation, and prediction. Can J Plant Sci 86:33–47. https://doi.org/10.4141/P04-171
Boller B, Schubiger FX, Kölliker R (2010) Red clover. In: Boller B et al (eds) Fodder crops and amenity grasses - Handbook of plant breeding. Fodder crops and amenity grasses. Springer, Berlin, pp 439–455
Brophy C, Finn JA, Lüscher A, Suter M, Kirwan L, Sebastià MT, Helgadóttir Á, Baadshaug OH, Bélanger G, Black A, Collins RP, Čop J, Dalmannsdottir S, Delgado I, Elgersma A, Fothergill M, Frankow-Lindberg BE, Ghesquiere A, Golinska B, Golinski P, Grieu P, Gustavsson AM, Hӧglind M, Huguenin-Elie O, Jørgensen M, Kadziuliene Z, Kurki P, Llurba R, Lunnan T, Porqueddu C, Thumm U, Connolly J (2017) Major shifts in species’ relative abundance in grassland mixtures alongside positive effects of species diversity in yield: a continental-scale experiment. J Ecol 105:1210–1222. https://doi.org/10.1111/1365-2745.12754
Collins RP, Helgadóttir Á, Fothergill M, Rhodes I (2001) Variation amongst survivor populations of two white clover cultivars collected from sites across Europe: morphological and reproductive traits. Ann Bot 88:761–770. https://doi.org/10.1006/anbo.2001.1462
Donaghy DJ, Fulkerson WJ (1998) The importance of water-soluble carbohydrate reserves on re-growth and root growth of Lolium perenne. Grass Forage Sci 53:211–218
Ergon Å, Kirwan L, Bleken M, Skjelvåg AO, Collins RP, Rognli OA (2016) Species interactions in a grassland mixture under low nitrogen fertilization and two cutting frequencies: I. dry matter yield and dynamics of species composition. Grass Forage Sci 71:667–682. https://doi.org/10.1111/gfs.12250
Ergon Å, Skøt L, Sæther VE, Rognli OA (2019) Allele frequency changes provide evidence for selection and identification of candidate loci for survival in red clover (Trifolium pratense L.). Front Plant Sci 10:718. https://doi.org/10.3389/fpls.2019.00718
Ford JL, Barrett BA (2011) Improving red clover persistence under grazing. Proc New Zealand Grassl Assoc 73:119–124
Frame J, Charlton JFL, Laidlaw AS (1998) Temperate forage legumes. CABI International, Wallingford, UK
Frankow-Lindberg BE, Svanӓng GK, Hӧglind M (1997) Effects of an autumn defoliation on overwintering, spring growth and yield of a white clover/ grass sward. Grass Forage Sci 52:360–369
Frankow-Lindberg BE (2001) Adaptation to winter stress in nine white clover populations: changes in non-structural carbohydrates during exposure to simulated winter conditions and spring regrowth potential. Ann Bot 88:745–751. https://doi.org/10.1006/anbo.2001.1432
Gale F, Hansen J, Jewison M (2014) China’s Growing Demand for Agricultural Imports, EIB-136, U.S. Department of Agriculture, Economic Research Service
Herrmann D, Boller B, Studer B, Widmer F, Kölliker R (2008) Improving persistence in red clover: insights form QTL analysis and comparative phenotypic evaluation. Crop Sci 48:269–277. https://doi.org/10.1007/s00122-005-0158-1
Helgadóttir Á, Dalmannsdóttir S, Collins RP (2001) Adaptational changes in white clover populations selected under marginal conditions. Ann Bot 88:771–780. https://doi.org/10.1006/anbo.2001.1438
Hoekstra NJ, De Deyn GB, Xu Y, Prinsen R, Van Eekeren N (2018) Red clover varieties of Mattenklee type have higher production, protein yield and persistence than Ackerklee types in grass–clover mixtures. Grass Forage Sci 73:297–308. https://doi.org/10.1111/gfs.12307
Jensen ES, Peoples MB, Boddey RM, Gresshoff PM, Hauggaard-Nielsen H, Alves BJR, Morrison MJ (2012) Legumes for mitigation of climate change and the provision of feedstock for biofuels and biorefineries. A review. Agron Sustain Dev 32:329–364. https://doi.org/10.1007/s13593-011-0056-7
Kirwan L, Lüsher A, Sebastiá MT, Finn JA, Collins RP, Porqueddu C, Helgadóttir Á, Baadshaug OH, Brophy C, Coran C, Dalmannsdóttir S, Delgado I, Elgersma A, Fothergill M, Frankow-Lindberg B, Golinkski P, Grieu P, Gustavsson A-M, Höglind M, Huguenin-Elie O, Iliadis C, Jørgensen M, Kadziuliene Z, Karyotis T, Lunnan T, Malengier M, Maltoni S, Meyer V, Nyfeler D, Nykanen-Kurki P, Parent J, Smit HJ, Thumm U, Connally J (2007) Evenness drives consistent diversity effects in intensive grassland systems across 28 European sites. J Ecol 95:530–539
Lager J, Gerhardson B (2002) Pathogenicity of clover root pathogens to pea, bean and lucerne. J Plant Dis Protect 109:142–151
Litrico I, Violle C (2015) Diversity in plant breeding: a new conceptual framework. Trends Plant Sci 20:604–613. https://doi.org/10.1016/j.tplants.2015.07.007
Loucks CES, Deen W, Gaudin ACM, Earl HJ, Bowley SR, Martin RC (2018) Genotypic differences in red clover (Trifolium pratense L.) response under severe water deficit. Plant Soil 425:401–441. https://doi.org/10.1007/s11104-018-3594-9
Lüscher A, Mueller-Harvey I, Soussana JF, Rees RM, Peyraud JL (2014) Potential of legume-based grassland–livestock systems in Europe: a review. Grass Forage Sci 69:206–228. https://doi.org/10.1111/gfs.12124
Marley CL, Fychan R, Fraser MD, Winters A, Jones R (2003) Effect of sowing ratio and stage of maturity at harvest on yield, persistency and chemical composition of fresh and ensiled red clover/lucerne bi-crops. Grass Forage Sci 58:397–406. https://doi.org/10.1111/j.1365-2494.2003.00392.x
Nadeem S, Steinshamn H, Sikkeland EH, Gustavsson A-M, Bakken AK (2019) Variation in rate of phenological development and morphology between red clover varieties: Implications for clover proportion and feed quality in mixed swards. Grass Forage Sci 74:403–414. https://doi.org/10.1111/gfs.12427
Ӧhberg H (2008) Studies of the persistence of red clover cultivars in Sweden with particular reference to Sclerotinia trifoliorum. Dissertation, Umeå: Swedish University of Agricultural Studies
Phelan P, Moloney AP, McGeough EJ, Humphreys J, Bertilsson J, O’Riordan EG, O’Kiely P (2015) Forage legumes for grazing and conserving in ruminant production systems. Crit Rev Plant Sci 34:281–326. https://doi.org/10.1080/07352689.2014.898455
Rapacz M, Ergon Å, Höglind M, Jørgensen M, Jurczyk B, Østrem L, Rognli OA, Tronsmo AM (2014) Overwintering of herbaceous plants in a changing climate. Still more questions than answers. Plant Sci 225:34–44. https://doi.org/10.1016/j.plantsci.2014.05.009
Riday H (2010) Progress made in improving red clover (Trifolium pratense L.) through breeding. Int J Plant Breed 1:22–29
Rubiales D, Annicchiarico P, Vaz Patto MC, Julier B (2021) Legume breeding for the agroecological transition of global agri-food systems: A European perspective. Front Plant Sci 12:782574. https://doi.org/10.3389/fpls.2021.782574
Sampoux J-P, Giraud H, Litrico I (2020) Which recurrent selection scheme to improve mixtures of crop species? Theor Expect G3(10):89–107. https://doi.org/10.1534/g3.119.400809
Sturite I, Rivedal S, Dörsch P (2021) Clover increases N2O emissions in boreal leys during winter. Soil Biol Biochem 163:108459. https://doi.org/10.1016/j.soilbio.2021.108459
Taylor NL (2008) A century of clover breeding developments in the United States. Crop Sci 48:1–13. https://doi.org/10.2135/cropsci2007.08.0446
Tork DG, Anderson NO, Wyse DL, Betts KG (2019) Domestication of perennial flax using an ideotype approach for oilseed, cut flower, and garden performance. Agron 9:707. https://doi.org/10.3390/agronomy9110707
Van Minnebruggen A, Roldan-Ruiz I, Van Bockstaele E, Haesaert G, Cnops G (2014) The relationship between architectural characteristics and regrowth in Trifolium pratense (red clover). Grass Forage Sci 70:507–518. https://doi.org/10.1111/gfs.12138
Verwimp C, Ruttink T, Muylle H, Van Glabeke S, Cnops G, Quataert P, Honnay O, Roldán-Ruiz I (2018) Temporal changes in genetic diversity and forage yield of perennial ryegrass in monoculture and in combination with red clover in swards. PLoS ONE 13:e0206571. https://doi.org/10.1371/journal.pone.0206571
Vik K (1955) Forsøk med engvekster og engdyrking II. Forskning Og Forsøk i Landbruket 6:173–318 (In Norwegian)
de Visser CLM, Schreuder R, Stoddard F (2014) The EU’s dependency on soya bean import for the animal feed industry and potential for EU produced alternatives. OCL 21:D407. https://doi.org/10.1051/ocl/2014021
Zanotto S, Palmé A, Helgadóttir Á, Daugstad K, Isolahti M, Öhlund L, Marum P, Moen MA, Veteläinen M, Rognli OA, Ergon Å (2021) Trait characterization of genetic resources reveals useful variation for the improvement of cultivated Nordic red clover. J Agron Crop Sci 207:492–503. https://doi.org/10.1111/jac.12487
Acknowledgements
We are grateful for advice on red clover crossing from Petter Marum and Helga Amdahl in Graminor AS and for the technical assistance from Øyvind Jørgensen, Liv Berge, Karin Svinnset and Anne Langerud.
Funding
Open access funding provided by Norwegian University of Life Sciences. This study was funded by Norwegian Research Council Project No. 225330 (Agropro).
Author information
Authors and Affiliations
Contributions
ÅE conceived the idea for the study and collected survivors from Ås. AKB collected survivors from Stjørdal. ÅE and AKB designed the experiments. ÅE conducted the crossings, the growth and freezing experiments, analyzed the data and wrote the manuscript. AKB critically revised the manuscript and approved the final version.
Corresponding author
Ethics declarations
Conflict of interests
The authors have no relevant financial or non-financial interests to disclose.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Ergon, Å., Bakken, A.K. Breeding for intercropping: the case of red clover persistence in grasslands. Euphytica 218, 98 (2022). https://doi.org/10.1007/s10681-022-03051-7
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s10681-022-03051-7