
Abstract
Increased flux of carbon and nutrients from human activities in river basins were linked to acidification and deepwater hypoxia in estuaries and coastal areas worldwide. Annual loads (1995–2011) of suspended particulate matter (SPM), dissolved organic carbon (DOC), total nitrogen (TN) and total phosphorus (TP) were assessed at the Lake Ontario inlet of the St. Lawrence River (SLR) (7110 m3 s−1) and its estuarine outlet at Québec City (12,090 m3 s−1). Internal loads from the Ottawa River (1950 m3 s−1), seventeen other tributaries, urban wastewaters, atmospheric deposition and erosion were also estimated. Erosion (65% of SPM, 29% of TP), inflow from Lake Ontario (42% of DOC, 47% of TN) and Ottawa River (28% of DOC) contributed important flux to the estuary. Loads from other tributaries (20 and 27% of TN and TP at Quebec City) largely exceeded municipal sources (6% of exported TN and TP) and require future remediation. Aquatic plants fixed 277,000 t of C, 49,000 t of N and 7000 t of P (May–Sept.), delaying the nutrient flux to the estuary and turning the SLR into a nutrient sink over summers of lowest discharge. Degradation of exported organic C could consume 5.4–7.1 million t O2 year−1 in the estuary whereas SLR flux of N and P represent 31–47% and 7–14% of total annual estuarine flux, respectively. Carbon and Nitrogen flux from freshwaters partly explain the decline in pH and oxygen concentrations in deep estuarine waters thus highlighting the need to reduce diffuse sources of nutrients in the entire watershed.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Anthropogenic eutrophication of estuaries and coastal areas resulting from rising carbon and nutrient flux from freshwaters is an issue of increasing importance in many parts of the world (Gilbert et al. 2010; Sutton et al. 2011). Inflow of nutrients, especially nitrogen (N), stimulates primary production in the estuary, occasionally leading to a rise in frequency and intensity of harmful algal blooms (Weise et al. 2002; Bricker et al. 2008) which induce massive mortality of marine fish, birds and mammals (Starr et al. 2017). Organic carbon exported from the watershed and plankton produced within the estuary eventually sediments into deep estuarine waters, where enhanced microbial activity induces hypoxia and anoxia (Kemp et al. 2009). Median rates of oxygen decline are more severe in a 30 km band near the coast than in the open ocean (>100 km from the coast) (Gilbert et al. 2010) which also exerts a range of negative effects on marine coastal ecosystems (Diaz and Rosenberg 2008). Microbial degradation of organic matter generated by coastal eutrophication also amplifies acidification (Wallace et al. 2014). As with many coastal waters of the world’s ocean, the bottom layers of the St. Lawrence Estuary have shown declining oxygen concentrations (Gilbert et al. 2005) and concurrent rising acidification (Mucci et al. 2011).
Inflow of organic carbon or nutrients from terrestrial origins could thus partially account for increased O2 consumption in deep estuarine areas, either directly (through bacterial degradation of organic matter) or indirectly (through stimulation of estuarine primary productivity) (Thibodeau et al. 2006). Marine phytoplankton in temperate regions tend to be N-limited and suffer increasingly from cultural eutrophication as a result of N addition (Howarth et al. 1996). In contrast, phosphorus (P) supply limits primary productivity of most freshwater systems (Dillon and Rigler 1974). Many large rivers flowing to coastal waters are thus subjected to regulation that limits phosphorus inputs without concerns for the effects of uncontrolled N additions on estuarine areas located downstream. Riverine nutrient loads delivered to estuaries can be further modulated temporally by seasonal variations of river discharge and primary production processes (Hudon and Carignan 2008).
In sharp contrast with other large rivers, primary production in the St. Lawrence River (hereafter designated as SLR) is favoured by its unusually high water clarity (Secchi depth 1–10 m) and low suspended particulate matter (SPM) concentration (Meybeck and Ragu 1996). After exiting Lake Ontario, the SLR flows through a succession of broad (2–12 km-wide) and shallow (mean depth 3–7 m) fluvial lakes, which are extensively colonized by submerged aquatic vegetation (Vis et al. 2007; Lévesque et al. 2017). Little interest has been given to the seasonal variations in carbon and nutrient flux as a result of within-river biological processes such as carbon fixation, nutrient assimilation and transformation by bacteria and primary producers. Although geochemical and hydrological processes largely determine annual loads delivered by rivers to their estuary, we hypothesized that biological processes could modulate the seasonal carbon, nitrogen and phosphorus (hereafter referred to as C, N and P) flux exported from the SLR.
In the SLR, annual transport and loads of carbon (Pocklington and Tan 1987; Hélie and Hillaire-Marcel 2006), suspended sediment (Rondeau et al. 2000) and various contaminants (Carignan et al. 1994; Pham et al. 2000; Rondeau et al. 2005) have been previously documented. Over the last few decades, however, major changes in urban wastewater treatment (Marcogliese et al. 2014), farming practices (Clair et al. 2013), and temperature and hydrological regimes (Hudon et al. 2010) have occurred in the SLR watershed, most likely inducing changes in the C, N, and P flux to the estuary. Although degradation of the estuarine environment is increasingly attributed to river enrichment, riverine contribution must be assessed in comparison with oceanographic processes (e.g. upwelling). In addition, the relative importance of different anthropogenic sources of C, N, and P within the watershed, remains to be established in order to prioritize remediation actions.
The present study presents a mass-balance assessment of SPM and C, N, and P loads at the inflows (Lake Ontario, Ottawa River) and the outflow (Québec City) of the SLR (1995–2011). The contributions of internal sources were evaluated, including tributaries, aerial deposition, erosion and municipal wastewater treatment plants. The importance of anthropogenic sources was then contrasted with the magnitude of riverine primary production, as an ecosystemic service to within-river nutrient processing and water epuration. This information should help prioritize the interventions required in the watershed to alleviate the decline in O2 concentration, the increase of acidification and their impacts on fauna in the Laurentian channel of SLR estuary.
Study area
With a mean annual discharge of 12,090 m3 s−1 at Québec City (hereafter referred to as Québec), the SLR has the second largest discharge in North America, after the Mississippi. Over its 550 km course between Kingston (at Wolfe Island), at the outlet of Lake Ontario, and Québec, the SLR drains a watershed of over one million km2 (Table 1; Fig. 1). Lake Ontario outflow into the SLR represents, on average, 70% of total discharge at Québec. SLR discharge is regulated on a weekly basis at Cornwall through the operation of the Moses-Saunders dam (Fig. 1). Discharge of the Ottawa River (≈20% of SLR discharge at Québec) and other SLR tributaries (≈10% of SLR discharge at Québec) show strong seasonal variations, which largely drive the seasonal changes in SLR water level.

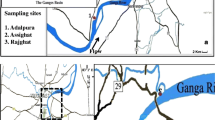
Map of the St. Lawrence River showing its two major inlets at Wolfe Island (Lake Ontario) and Carillon (Ottawa River) and outlet at Québec (stars), just upstream of the brackish water estuary (inset A). Most of the population is concentrated in the region between the greater Montreal area and Trois-Rivières (rectangle B). The outlets of the 6 largest physical–chemical WWTP are located at the eastern tip of the island of Montreal (circle). Major tributaries (circled numbers, see also Table 5), urban centers (dark grey), farmlands (light grey) and fluvial lakes Saint-François, Saint-Louis and Saint-Pierre (inset C) are shown. Inset C shows the tributaries flowing into Lake Saint-Pierre, the largest fluvial lake and wetland area (≈350 km2) of the St. Lawrence River. Colour version in pdf. (Color figure online)
The presence of the Laurentian Great Lakes (watershed: 737,000 km2), which are large (25,700–82,170 km2), deep lakes with very long residence times (2.6–191 years), at the head of SLR watershed results in properties that are unique for a river of this size. Waters originating from Lake Ontario are characterized by exceptionally low (<1 mg L−1) suspended particulate matter (SPM) and a high clarity (Secchi depth ≥10 m). In the SLR headwaters, reduction of phosphorus (P) loads following the Great Lakes Water Quality Agreement (IJC 1972; Stevens and Neilson 1987) coincided with declining TP concentrations in offshore Lake Ontario (25 to <10 μg P L−1 between 1976 and 1999, Nicholls et al. 2001). However, TP decline has not been observed in the littoral areas of Lake Ontario where the establishment of Dreissenid mussel population (1989–1991, Mills et al. 2003) and P inflow from tributaries (Makarewicz et al. 2012a) and urban sources have allowed a return to nearshore proliferation of Cladophora (Higgins et al. 2010). The 1995–2011 period covered by the present study thus coincides with substantial changes in nutrient dynamics in Lake Ontario.
Between Lake Ontario (Wolfe Island) and Québec, the river drains a watershed inhabited by 7 million people, about 4 million of whom live along its shores (Fig. 1, inset B) and discharge their wastewaters to the river following varying levels of treatment. In the greater Montreal area alone, the SLR receives the primary-treated municipal effluents of over 3 million inhabitants (Fig. 1, circle, Marcogliese et al. 2014). The Ottawa River (watershed: 146,334 km2) and other north shore tributaries (watershed: 63,790 km2) largely drain boreal forests growing on the Precambrian Shield, whereas south shore tributaries (watershed: 56,840 km2) originate from the Appalachian mountain range and mostly drain Ordovician and Silurian limestone (Douglas 1970). Fertile lowlands on both shores of the SLR valley are intensively farmed for corn, soy and hog production; farmlands occupy between <1 and 86% of watershed area of tributaries (Patoine 2017).
The SLR comprises four large (150–350 km2), shallow (mean depth 3–7 m) and broad (2–12 km wide) fluvial lakes, which are bisected by a central navigation channel (≥11.3 m deep). Fluvial lakes St. Lawrence, Saint-François, Saint-Louis and Saint-Pierre together support >200 km2 of emergent wetlands (Hudon et al. 2006), in addition to extensive riverbed colonization by submerged aquatic macrophytes (Lévesque et al. 2017). In Lake Saint-Pierre alone (LSP, ≈350 km2, Fig. 1, inset C), overall primary production (2000–2001) was estimated at about 30 × 103 tonnes of carbon per year (105 g C m−2 year−1), distributed among phytoplankton (29-38%), epiphytes (14-19%), submerged (23%) and emergent (25–29%) macrophytes (Vis et al. 2007).
Methods
Water discharge
Daily water discharge data were obtained for the St. Lawrence River at Cornwall Moses Saunders dam (Environment Canada 2012a, Daily St. Lawrence River Discharge at Cornwall (1995–2011). Great Lakes—St. Lawrence Regulation Office, Cornwall, Ontario. Rob Caldwell, P. Eng, Great Lakes—St. Lawrence Regulation Office, 111 Water St E, Cornwall, ON K6H 6S2, personal communication) and the Ottawa River at the Carillon dam (Hydro-Québec 2012). Daily St. Lawrence River discharge at Québec was obtained from the sum of measured discharge at LaSalle (near Montreal, Fig. 1) with that of major tributaries between Montreal and Québec, including a correction for ungauged watersheds (Environment Canada 2012b, Daily St. Lawrence River Discharge at Québec (1995–2012). Sylvain Martin, Environmental Data Management, Hydrology and Ecohydraulic Section, Meteorological Service of Canada, Environment Canada, 801–1550, avenue d’Estimauville, Québec, QC G1 J 0C3, personal communication).
Water quality sampling
Water quality variables were measured at the outlet of Lake Ontario at Wolfe Island (Hill 2013), Carillon, and the city of Lévis (across from Québec) (Environment Canada MSQE-BOATL Divisions) (Fig. 2). Nitrogen data (total nitrogen, nitrites-nitrates, ammonium) were obtained for five additional sites near Québec (Lauzon, Sainte-Foy and 3 sites in the navigation channel south of île d’Orléans) from the Quebec Ministry of the Environment (MDDELCC, Suivi de la Qualité des Rivières). Depending on site, measurements were available either on a year-round, weekly basis (Wolfe Island, Carillon, Lévis) or monthly, for the ice-free months (Lauzon, Sainte-Foy, navigation channel). Sites near Québec (Lévis, Lauzon, Ste-Foy and the three channel sites, hereafter designated together as Québec) were sampled under ebbing tide conditions to ensure samples were representative of outflowing river water (Rondeau 1998). When more than one measurement of a given variable was available on a given day (for example, at the 6 Québec-area sites), an average daily concentration was calculated.

Measurements available over the span of the study period (1995–2011) for each water quality variable at the outflow of Lake Ontario to the St. Lawrence River (Wolfe Island, grey lines), at the mouth of the Ottawa River (Carillon, fine black lines) and at SLR outlet to the estuary (Québec City, bold black lines)
At each site, water samples were collected from water intake pipes (3–10 m-depth) for measurements of suspended particulate matter (SPM, APHA 1995), total nitrogen (TN, method CPQ123E, Environment Canada 2014) and total phosphorus (TP, Stainton et al. 1977). At Carillon and Québec, duplicate samples were filtered for the measurement of dissolved organic carbon (DOC, Shimadzu TOC-5000), total dissolved phosphorus (TDP), total dissolved nitrogen (TDN), total Kjeldahl Nitrogen (TKN, method NLET 7010, Environment Canada 2014), nitrites-nitrates and ammonia (NO2 +NO3 and NH3, Lachat autoanalyzer methods number 10–107–04–1–B and 11–104–03–1–B). At Wolfe Island, water samples for nutrient assessment were not filtered (with the exception of TDP and DOC) owing to the very low particulate matter concentration (<1 mg L−1). Total dissolved organic nitrogen (DON) was calculated at Wolfe Island (DON = TKN–NH3) and at Québec (DON = TDN–NO2–NO3–NH3), but could not be assessed at Carillon (no TDN or TKN measurements). Living phytoplankton abundance (μg chlorophyll a L−1) was determined spectrophotometrically after filtration (Whatman GF/C) of 300–1000 mL of surface water. Filters were ground and extracted in 9:1 V/V acetone and the optical densities of the extract was measured at 663, 645 and 630 nm (Wetzel and Likens 1991).
Assessment of urban sources of C, N, and P
Urban sources of SPM, C, N, and P were assessed for the greater Montreal area down to Trois-Rivières (Fig. 1, inset B) which comprises the major urban centers (population ≈3.2 millions) discharging their treated wastewaters directly into the SLR. Mean annual concentrations (SPM, TP) and SPM loads in treated waters for the year 2010 were obtained for each wastewater treatment plant (hereafter designated as WWTP) subjected to mandatory annual performance assessment (Moreira 2011). Since chemical phosphorus removal is also carried out during summer, additional values of annual TP loads (2010–2012) were obtained in the treated effluents of the 50 municipal WWTP.
Concentrations of substances other than SPM and TP, for which measurements were not mandatory for WWTP (DOC, TKN, NH3, NO2–NO3), were estimated separately for large physical–chemical (N = 6) and small (N = 44) WWTP using other methods of water treatment. For the large WWTP, average concentrations measured in the treated effluent of the City of Montreal were multiplied by total discharge (36.7 m3 s−1) from the six physical–chemical WWTP. Taken together, these six plants accounted for 80% of the population and 87% of treated effluent discharge to the St. Lawrence River between Coteau Landing and Trois-Rivières (Fig. 1), which justified the separate assessment of their effluent. Large WWTP included City of Montreal (1.78 million persons, 28.8 m3 s−1), Longueuil-Centre d’épuration Rive Sud (350 000 persons, 3.7 m3 s−1), Laval-La Pinière (271 630 persons, 3.2 m3 s−1), Laval-Fabreville (59,000 persons, 0.5 m3 s−1), Repentigny (69,955 persons, 0.3 m3 s−1) and Rosemère (27,000 persons, 0.2 m3 s−1). DOC and NO2–NO3 concentrations were estimated from sporadic point measurements made between May and October 2012 at the treated effluent of the City of Montreal WWTP (Ville de Montréal 2012). High frequency measurements of TKN (202 measurements between June and Dec. 2012) and NH3 (1015 measurements taken bi-weekly in 2000–2011) were obtained for the same effluent since these components represent the bulk of N originating from urban sources. Annual loads of DOC, NO2–NO3 and TKN outflowing from the City of Montreal WWTP were calculated from the product of mean discharge and concentration. Annual load of NH3 was calculated using the discharge-weighted concentration method (Raymond et al. 2013).
For the 44 smaller WWTP using treatments other than physical–chemical (activated sludge, aerated ponds, biofiltration, other treatments), mean concentrations of SPM, NH3, NO2–NO3 and TKN in treated outflow were assessed using 2001 data for 13 WWTP sampled during both cold (winter) and warm (summer) conditions (MEQ-EC 2001). Owing to the scarcity of published concentrations and in the absence of high frequency measurements for individual WWTP, mean annual loads originating from the 44 small municipal WWTP were assessed from the product of mean concentrations derived above and the sum of mean daily discharge multiplied by 365 days and were maintained constant for the entire 1995–2011 period. Our use of recent data (2010–2012) to estimate load values for the largest WWTP should represent the upper range of potential urban contributions to total SLR loads (1995–2011). Total annual loads and error values (95% confidence interval) for the 6 large (physical–chemical treatment) and the 44 small (other treatment methods) WWTP were obtained from the sum of individual values without transformation.
Loads from tributaries and diffuse sources
The contribution of 17 other significant (other than the Ottawa River) SLR tributaries (Fig. 1) and of the sum of smaller (watershed <1000 km2) north and south shore tributaries as internal sources of SPM, TN and TP was obtained from recent (2009–2012) load estimations (Patoine 2017). Concentration of each substance was measured at least at a monthly frequency near the mouth of each river; mean daily discharge was estimated from the nearest gauging sites using a kriging approach. Annual loads of each substance in each tributary were calculated with the software Flux32 v.3.32 [Regression Method 6 described in Walker (1996)]. For DOC, data for January 2009 to December 2012 were used to derive a flow-concentration relationship (N = 34–42) for each river except for the Sainte-Anne (2005–2012, N = 79) and Yamaska (2008–2012, N = 50) rivers for which additional measurements from other years were included to improve the relationship. Data was stratified (January to April, May to August, September to December) when necessary, to eliminate seasonal patterns. For small tributaries (watershed <1000 km2, see list as a footnote to Table 5), DOC loads were calculated the same way using 27 (2009–2012) to 68 (2007–2012) data points for each river; total annual loads and error values (95% confidence interval) for the north and south shores were obtained from the sum of individual values without transformation.
Shoreline and riverbed erosion (Rondeau et al. 2000) and atmospheric deposition (Ouimet and Duchesne 2009; Mladenov et al. 2012) were obtained from literature data. Erosion of the bed and banks of the St. Lawrence River was previously estimated in the order of 65% of the suspended sediments measured at Québec (Rondeau et al. 2000). The TP load resulting from eroded glacial clays from the St. Lawrence River bed was assessed from the P content of fine-grained inorganic sediments predating 1950 (~950 mg P kg−1, Carignan et al. 1994) applied to the estimation of eroded material. Atmospheric deposition of N on the river wetted surface (2637 km2 between Wolfe Island and Québec) was estimated from a mean ± SD value of 5.8 ± 1.8 kg N ha−1 year−1 for southern Québec (Ouimet and Duchesne 2009). In the absence of recent regional estimates, atmospheric deposits of C (7 kg C ha−1 year−1) and P (0.15 kg P ha−1 year−1) from the Colorado Rocky Mountains were used (Mladenov et al. 2012).
Calculation of annual river loads and uncertainty
Loads originating from Wolfe Island (outlet of Lake Ontario), Carillon (Ottawa River mouth) and Québec were calculated using the most appropriate method to address the different patterns of temporal variations characterizing each water quality variable at each site. Each variable sampled at each site was assigned to one of five different patterns of temporal variations, which are presented below in decreasing order of parsimony: (1) no systematic temporal variation: daily flux were calculated as the product of mean daily discharge and mean concentration; (2) concentrations related to discharge: daily flux were calculated as the product of daily discharge and predicted concentration from log–log regression with daily discharge; (3) consistent among-year seasonal (day-of-year) pattern in the concentrations: fluxes were calculated as the product of smoothed concentration pattern and daily discharge; (4) no persistent seasonal pattern, but inter-annual variations in concentrations: fluxes were estimated as the product of annual mean concentrations and daily discharge; (5) both persistent seasonal and inter-annual variations in concentrations: the two effects were added and multiplied with daily discharge values (Foglia et al. 2007; Ye et al. 2008).
The fit of each temporal model to each measurement series (5 temporal models described above × 9 water quality variables × 3 sites = 135 assessments) was evaluated both statistically and graphically. Statistical fit was measured with the corrected Akaike information criterion (AICC, Burnham and Anderson 2002) and residuals were examined visually for lack of randomness. In addition, temporal model selection took into account consistency with known environmental effects (hydrology, climate, biology), minimal AICC and error values, and under equivalent performance, the most parsimonious models (no temporal trend, discharge) were preferred (Foglia et al. 2013). Since nutrient concentrations and dynamics of chemical species may differ considerably among sites according to watershed morphology, models were selected independently for each variable at each site. Long-term (1995–2011) temporal trends in C, N, and P concentrations were tested using Spearman rank-order correlation with year (SAS, PROC CORR).
A mass balance approach was used to determine whether the SLR acted as a source or a sink of SPM, C, N, and P for the estuary over the seasonal cycle (between months) and under different hydrological conditions (between years). For this, weekly loads measured upstream (Wolfe Island and Carillon) and downstream (Québec) were calculated for the entire time series. Weekly loads were then summed up on a monthly basis which were used to calculate the net monthly flux by difference (load at Québec − (∑ loads at Wolfe Island + Carillon)). A monthly temporal scale was selected to minimize the uncertainty and integrate the cumulative effects and seasonal variations of water parcel travel time downstream. Average water travel time between the outlet of Lake Ontario (Wolfe Island) and Québec (550 km) was estimated to be in the order of one to 2 weeks for an average speed between 0.5 and 1 m s−1 in the main river channel over the range of seasonal discharge conditions. The annual balance (and associated error) of incoming C, N, and P loads over the Québec portion of the watershed [load at Québec − (∑ loads at Wolfe Island + Carillon)] was compared with the sum of loads and error values (95% confidence interval) for the quantifiable internal sources (erosion, municipal WWTP, tributaries and atmospheric deposition). Uncertainty on load estimations was assessed using the standard error and 95% confidence interval associated with the mean load value for different data subsets as described in supplementary material (.csv files). Data files included in the Supplementary Material provide (1) a detailed comparison of the fit of each potential model (AICC) (File Supplementary Material—Model Fit), (2) an assessment of annual loads and errors for each substance at the three sites for each year (File Supplementary Material—Annual Loads) and (3) the evaluation of errors associated to the estimation of monthly surplus load at Québec (Output–Inputs) (File Supplementary Material—Québec Surplus). For example, comparison of the performance of the 5 models fitted for NO3 loads at Québec yielded AICC criteria ranging from 785 (Year, poorest model fit) to 129 (Season; Flow, best model fit); taken on their own, models accounting for Season (AICC = 176) and for Flow (AICC = 487) were markedly less efficient (Supplementary Material Model Fit). Corresponding estimates of annual NO3 loads at Québec ranged from 123,568 (no model) to 134,208 t N year−1 (Year; Season), representing an overall variation of <10% (File Supplementary Material—Annual Loads). On a monthly basis, net (mean difference ± standard error) monthly DIN load (constituted of about 90% NO3 at Québec) outflowing to the SLR estuary ranged 9-fold between August (1097 ± 335 t N) and April (6853 ± 746 t N) and among years (722 ± 586 t N in 2010 and 6341 ± 455 t N in 2009) (File Supplementary Material—Quebec Surplus). The consistency of error components estimated above gave us confidence in the robustness of the results and major conclusions of our study.
Contribution of primary producers to carbon fixation and nutrient assimilation
We assessed the importance of annual primary production in the SLR to compare the relative magnitude of C fixation and assimilation of N and P by aquatic plants in comparison with other processes. In Lake Saint-Pierre, over years of average (2000) and low (2001) water levels, total annual primary production of phytoplankton (29–38%), epiphytes (14–19%), submerged (23%) and emergent (25–29%) macrophytes was estimated at about 30 × 103 tonnes of carbon per year, yielding a mean areal C fixation of 105 g C m−2 year−1 (Vis et al. 2007). Using the Redfield (molar) ratio for C:N:P (106:16:1), we estimated that such carbon fixation (105 g C m−2 year−1) would require a corresponding assimilation of N (18.5 g N m−2 year−1) and P (2.6 g P m−2 year−1). The total annual contribution of primary producers to C fixation and nutrient assimilation was then estimated by applying these values to the entire wetted area of SLR bottom (2637 km2) between Wolfe Island and Québec. This estimation was deemed conservative since both the biomass (Lévesque et al. 2017) and maximum depth of colonization (i.e. water transparency, see Hudon et al. 2000) of submerged aquatic vegetation are lower in Lake Saint-Pierre than for fluvial lakes located upstream.
Results
Hydrological conditions and water quality
Mean annual SLR outflow at Québec averaged 12,090 m3 s−1 for the 1995–2011 period which is very close to the value of 11,880 m3 s−1 obtained for the same period using the alternative method of Bourgault and Koutitonsky (1999). Mean monthly discharge showed strong seasonal variations with maximal values in the spring (April–May) and minimum discharge in summer (August–September) (Fig. 3). Lake Ontario discharge to the SLR showed little seasonal (e.g. monthly) variability and, on an annual basis, constituted the bulk of total freshwater outflow at Québec (Fig. 3b). In contrast, Ottawa River discharge exhibited high inter-annual and seasonal variability. Seasonal variations in SLR discharge at Québec were largely driven by the Ottawa River (min–max daily discharge over the sampling period: 1520–2360 m3 s−1) and other tributaries which together accounted for 28% (August) to 55% (April) of total discharge at Québec.

Comparison of a mean annual (1995–2011, dotted line indicates overall mean) and b mean monthly discharge (m3 s−1) at Wolfe Island (squares), Carillon (diamonds) and Québec (circles). Overall (1995–2011) mean (±SD, full lines) monthly discharges are contrasted with years of high (2008, medium dashed lines) and low discharge (2010, dotted lines)
Waters originating from Lake Ontario exhibited very low concentrations of SPM, DOC, TP and chlorophyll a (Table 1). In comparison, consistently higher concentrations of all substances (except DON and NO2–NO3) were recorded at Québec, including SPM (30-fold), TP (2-fold), TDP (2-fold), NH3 (3-fold), DOC (1.3-fold) and chlorophyll a (2.6-fold). High concentrations measured at the mouth of the Ottawa River partially explained the recorded increases at Québec (Table 1).
Temporal variations of C, N, and P concentrations
The temporal variations of concentrations of each water quality variable differed markedly among sites as a function of the morphology of their watershed. At Wolfe Island, none of the water quality variables showed a correlation with discharge owing to the very stable outflow of Lake Ontario. Instead, concentrations of most variables fluctuated on a seasonal and/or inter-annual basis (Table 2) consistent with the cycle of vertical mixing/stratification and year-to-year variations of Lake Ontario waters. In contrast, river discharge and seasonal variations were strong drivers of SPM, DOC and nutrient concentrations in the Ottawa River and at Québec, indicative of the influence of seasonal variations in runoff and temperature over the watershed. Accounting for differences in temporal variations allowed us to select the optimal model to calculate annual loads of each substance at each site.
Time series of NO2–NO3 and TP over several years exemplify the contrast in seasonal/inter-annual patterns at our three sites (Fig. 4; Table 2). At the outlet of Lake Ontario, concentrations of both NO2–NO3 and TP showed seasonal variations with minimum concentrations over summer months and maximum concentrations in winter. In addition, winter and spring (February to May) maxima tended to rise over the last years of the NO2-NO3 and TP series at Wolfe Island, justifying our use of season and year to model their loads at the Lake Ontario outlet (Table 2). For the 1995–2001 period, mean monthly NO2–NO3 concentrations were consistently <0.4 mg N L−1 whereas concentrations >0.4 mg N L−1 occurred for 10 of the 15 months between January 2010 and March 2011(Fig. 4a). For TP, mean monthly concentrations remained <10 µg P L−1 for the 2001–2004 period but rose >20 µg P L−1 for 12 of the 15 months between January 2010 and March 2011 (Fig. 4b). At the mouth of the Ottawa River (Carillon), discharge explained most of the variations in SPM (Spearman r = 0.51, N = 882, p < 0.0001), DOC (r = 0.44, N = 172, p < 0.0001), and TN (r = 0.36, N = 43, p = 0.02) concentrations, whereas concentrations of dissolved inorganic nitrogen species (NO2–NO3 Fig. 4c, and NH3) and chlorophyll a fluctuated seasonally (Table 2). Neither TP nor TDP followed any specific seasonal or long-term (1995–2011) temporal trend (Table 2).

Temporal trends in mean monthly concentrations of NO2–NO3 (left) and TP (right) measured at Wolfe Island (a, b, top), Carillon (c, d, middle) and Québec (e, f, bottom). Axes are on the same scale for each nutrient
At the SLR outlet at Québec, positive correlations between river discharge and DOC (Spearman r = 0.59, N = 406, p < 0.0001), and TN concentrations (r = 0.51, N = 35, p < 0.0001) were observed, whereas models including discharge and season accounted for most of the temporal variations of SPM, TP, TDP and NO2–NO3 (Table 2). Overall, regardless of major hydrological and morphological differences among sites, all dissolved nitrogen forms, including NH3 (Table 2) and NO2–NO3 (Fig. 4e), followed clear seasonal variations with highest concentrations observed over the coldest months and lowest concentrations in late summer. In contrast, chlorophyll a concentrations were maximal during the spring and summer at all sites. No significant long-term trend was observed for any of the water quality variables (Spearman rank-order correlation).
C, N, and P loads from urban sources
Wastewaters from the 6 largest physical–chemical WWTP represented the treated domestic effluents from about 2.6 million persons—about 4 times the population whose wastewaters were treated in the 44 small WWTP using other treatment methods (Table 3). Treated wastewater discharge and annual loads of SPM, TKN and DOC from the 6 large physical–chemical WWTP were about 2–10 times higher than for the 44 small WWTP. Large physical–chemical plants were more effective than small WWTP in removing TP, whose load was only twice that originating from the 44 small plants. As expected, TKN (i.e. organic N + NH3) represented the bulk of nitrogen loads in treated municipal waters whereas NO2–NO3 only represented a minor component (Table 3).
Loads at St. Lawrence River inlets and outlet
Calculation of annual flux of SPM, C, N, and P from Wolfe Island and Carillon further emphasized the major differences in morphology and watershed characteristics between Lake Ontario and the Ottawa River. In spite of a discharge 3.6 times higher at Wolfe Island than at Carillon, the SPM load originating from Lake Ontario was 4 times smaller than for the Ottawa River (Table 4). Loads of DOC, TP, TDP, NH3 and chlorophyll a originating from the Great Lakes were only about twice those of the Ottawa River. In contrast, NO2–NO3 loads from Lake Ontario were 5 times higher than from the Ottawa River, contributing 58% of the load at Québec. Loads originating from the 50 municipal WWTP were generally small in comparison with those from the main river stem and the Ottawa River with the notable exception of NH3, which represented 51% of the load measured at Québec (Table 4).
At Québec, SPM exhibited a 10-fold increase in comparison with inflowing sources, whereas DOC (1.4-fold), TN (1.5-fold) and TP (2-fold) showed a lesser enrichment (Table 4). Loads of planktonic chlorophyll a were 2.3-fold higher at Québec than at the two major inlets. This information indicates that over its 550 km-long course, the St. Lawrence is enriched by diffuse nutrient sources, including tributaries, erosion and atmospheric sources, which reflect land-use through their individual and area/specific loads.
Diffuse sources and area-specific C, N, and P loads
In spite of the large population inhabiting the Laurentian Great Lakes watershed (≈34 million persons) and intensive land-use, area-specific loads of SPM (0.13 t km−2 year−1), C, N, and P (750, 165 and 4 kg km−2 year−1, respectively) flowing into the SLR at Wolfe Island were very low, reflecting the large proportion of the watershed covered by the lakes themselves acting as a sedimentation basin for particles. Ottawa River area-specific output was about 2-3-fold higher than the Great Lakes for DOC (2650 kg C km−2 year−1), TN (236 kg N km−2 year−1) and TP (11 kg P km−2 year−1) but about 20-fold higher for SPM (2.99 t km−2 year−1, Table 5). Area-specific TN and TP measured at Québec (260 and 12 kg km−2 year−1, respectively) were slightly higher than output values for the Ottawa River watershed.
Load values from Wolfe Island and Carillon contrasted sharply with the inflow from tributaries draining the north and south shores of the SLR (Patoine 2017, Table 5). Taken together, these tributaries annually brought large loads of SPM (about 1.9 million t), DOC (0.46 million t), TN (53,000 t) and TP (3300 t) to the SLR. Area-specific carbon loads between 2.36 (Richelieu River) and 5.90 (Jacques-Cartier River) t C km2 year−1 reflected the proportion of forested watershed and land use. N/C ratio also provided a distinction between largely forested (N/C < 0.1) and intensively farmed watersheds (N/C > 0.1). Farming induced important differences in area-specific loads of SPM and nutrients among tributaries. South shore tributaries flowing through the fertile, intensively farmed (cultivated watershed area: 22–61%, Patoine 2017) St. Lawrence River lowlands contributed about 3 times the loads of TN as those draining less intensively farmed (cultivated watershed area: <1–17%, Patoine 2017) watersheds from the north shore. Of all the tributaries documented, the Yamaska (55% cultivated area) exhibited the highest area-specific output of SPM (42 t SPM km−2 year−1), TN (1579 kg N km−2 year−1) and TP (90 kg P km−2 year−1). Most major south shore tributaries (all but the Richelieu River) exhibited elevated area-specific output of SPM (>20 t SPM km−2 year−1), TN (>500 kg N km−2 year−1) and/or TP (>40 kg P km−2 year−1). In comparison, only two north shore tributaries (Du Nord and L’Assomption rivers) were similarly enriched (Table 5). Noticeably, small tributaries (<1000 km2) from both shores of the SLR carried the highest area-specific loads of SPM, C, N, and P (Table 5) since those watersheds were located almost entirely on the SLR lowlands and supported the highest proportion of farmlands (up to 82% of watershed area).
Annual atmospheric deposition of C, N, and P over the wetted St. Lawrence River surface from Wolfe Island to Québec (2637 km2) were estimated to be in the order of 4480, 1530 and 40 tonnes of C, N, and P, respectively (Table 6), assuming a high retention rate of nutrients and carbon into seldom flooded watershed area. Given a 64% NO3 composition (Ouimet and Duchesne 2009), atmospheric N deposition translated into a direct addition of about 980 tonnes of NO3 to the SLR (0.7% of annual NO2–NO3 load at Québec).
Mass-balance assessment and relative importance of individual sources compared to within-river C, N, and P processing by primary producers
The assessment of mean annual loads originating from various sources (Table 6) allowed us to determine their relative contribution to the total annual flux of SPM, C, N, and P into the SLR estuary at Québec which we present in decreasing order of magnitude. Shoreline and riverbed erosion accounted for the largest part of SPM (65%) and TP (29%) flux to the SLR estuary. By virtue of its large discharge, inflow from Lake Ontario carried 42% of DOC and 47% of TN loads at Québec. The Ottawa River was also a large source of SPM and DOC (8 and 28% of load at Québec, respectively). Although they represented much smaller cumulated watershed areas than the Ottawa River, tributaries draining the north and south shore (6% of watershed area each) contributed equivalent or higher proportions of SPM (8 and 25%, respectively), TN (7 and 13%) and TP (9 and 18%) as originated from the Ottawa River. Treated effluents from 50 municipal WWTP outflowing directly into the SLR contributed a small fraction of DOC (3%), TN (6%) and TP (6%) loads at Québec. Atmospheric deposition represented <1% of total annual C, N, and P flux at Québec. Unaccounted loads represented −4% of SPM, 5% of DOC, 11% of TN and −17% of TP load at Québec (Table 6, line K), revealing a higher level of uncertainty for the nutrient budgets than for SPM and DOC. The negative balance observed for SPM (−4%) and TP (−17%) suggested an overestimation of internal sources, possibly linked to our use of a fixed proportion (65% of SPM load at Québec) for erosion. The positive balance for DOC (5%) and TN (11%) was indicative of unaccounted internal sources such as small and ungauged tributaries, unconnected residences and industries, urban sewer storm overflow and/or other sources.
Annual C, N, and P turnover through primary production of aquatic plants (phytoplankton, epiphytes, submerged and emergent macrophytes) estimated for the entire wetted SLR surface represented a substantial fraction of the overall flux at Québec. Annual aquatic plant production fixed 277,000 t of C and assimilated 49 000 t of N and 7 000 t of P (Table 6, Line L), representing 20, 18 and 58% of annual loads at Québec, respectively. These percentages reveal that riverine plant production taking place over the growth season (May–Sept.) can significantly modify the seasonal nutrient and organic carbon flux to the estuary.
The observation that seasonal growth of aquatic plants could modify the seasonal flux of C, N, and P from the SLR to its estuary was examined further through the monthly SPM, C, N, and P net flux at Québec (Fig. 5) which revealed the major influence of seasonal and inter-annual discharge variations. All water quality variables exhibited peak loads in April, coincident with the spring freshet, followed by receding values during the summer growth season down to a minimum in September which coincides with the time of minimum discharge. Net flux of all substances rose again in the fall corresponding to rising discharge and reduced biological activity. Strong inter-annual variability was also observed for all substances with lower than average net flux over years of low discharge (2010) and maximum flux during high flow years (2008) (Fig. 5). For both TP and DIN, negative net monthly flux were observed for summer months (June–September) over years of low discharge, indicating the SLR acted as a nutrient sink under lowest water level conditions.

Net monthly flux (×1000 tonnes) of a SPM, b DOC, c TP and d DIN exiting from the SLR [monthly load at Québec—(monthly loads at Wolfe Island + Carillon)] to the estuary. Mean 1995–2011 (±SD) net flux are compared to monthly flux recorded in 2008 (high discharge) and 2010 (low discharge). The river acts as a net source when the outflow at Québec exceeds the documented sum of inflows from Wolfe Island and Carillon. Negative net monthly loads (hatched area) are identified as sink
Discussion
Annual and seasonal SPM, C, N, and P loads
Our study yielded an assessment of annual and seasonal (monthly) loads of SPM, dissolved C, total N and total P at SLR inlets (Wolfe Island + Carillon) and outlet (Québec). For SPM, mean annual loads of 1.02 × 105, 4.37 × 105 and 56.04 × 105 tonnes (at Wolfe Island, Carillon and Québec, respectively) from the present study were consistent with earlier values of 1.99, 4.35 and 69 × 105 tonnes SPM year−1 (1989–1993, Rondeau et al. 2000). The importance of erosion as a factor contributing SPM to the SLR was supported by the strong correlation between discharge and the seasonal pattern of monthly SPM flux from the SLR.
For TP, the estimated annual load of 1260 tonnes TP year−1 we used (Table 5; Patoine 2017) for the three major tributaries of Lake Saint-Pierre (Richelieu, Saint-Francois and Yamaska rivers for 2009–2012) compared well with that of Hudon and Carignan (2008) (1201 tonnes TP year−1 for 2000–2005). Overall, our estimations of annual loads of DOC, total dissolved P and inorganic N fractions at Wolfe Island and Carillon coincided within <10% with results from an earlier study (Hudon and Carignan 2008). Larger discrepancies in estimated loads of the present study and those of Hudon and Carignan (2008) were noticeable for SPM (30–100%) and TP (25%), which is understandable given the notoriously high variability of riverine particulate fractions.
Results from the present study yield area-specific annual N delivery of 165, 235 and 260 kg N km−2 year−1 for the outlet of Lake Ontario (Wolfe Island), the Ottawa River Basin (Carillon) and the entire Great Lakes—St. Lawrence River watershed at its outlet (Québec), respectively. Global N output at Québec (260 kg N km−2 year−1) from our study was higher than the 211.8 kg km−2 year−1 value reported for the SLR watershed (Clair et al. 2013), but was comparable to N inflows to the Baltic Sea from the Gulf of Finland (271 kg N km−2 year−1) (Sutton et al. 2011). Denitrification by microbial activity is increasingly invoked to explain the comparatively lower N exportation rate from the St. Lawrence River in comparison with other densely populated agricultural river systems (Clair et al. 2013). This process is enhanced by the long water residence time and vertical stratification in Lake Ontario in addition to a number of factors related to hydrology and physical characteristics of the watercourse (Seitzinger et al. 2006). In the SLR, summers of low discharge coincide with warm water temperature and reduced current speed in the shallow littoral areas where submerged vegetation proliferates (Hudon et al. 2010). Taken together, these conditions promote anoxia at the sediment interface, which stimulates bacterial denitrification and explains the high retention of nitrogen within the SLR during summers of reduced discharge (Hudon and Carignan 2008).
Our study of nutrients highlights the major effect of the Ottawa River and of nutrient-enriched tributaries. Rivers flowing on the south shore of the SLR drained only 6% of the watershed area at Québec, yet contributed 13 and 18% of the annual flux of TN and TP to the estuary respectively (Table 6). Diffuse anthropogenic sources of TP represented 64–99% of annual TP loads (Patoine 2017) originating from small tributaries (<1000 km2) draining the St. Lawrence River lowlands. Annual TP loads from the larger tributaries flowing into rural Lake Saint-Pierre (see Fig. 1 inset C) were primarily derived from diffuse anthropogenic (farming) sources (65–84% of TP loads) rather than from urban point sources (4–30%), with the only exception of the Saint-Francois River (22% farming and 38% urban sources)(Patoine 2017). Similarly, estimates of nutrient production in sub-catchments of the Red River (Manitoba) indicated that agricultural activities, particularly through synthetic fertilizer application, were by far the largest potential source of anthropogenic nutrients whereas residential sources in rural setting accounted for <1% of the total load of nutrients (Yates et al. 2012).
Area-specific N loads for SLR tributaries (Patoine 2017, Table 5) spanned a wide range of values and showed the expected relation between proportion of farmlands and N/C and N/P ratio. Nitrogen export rate for the 99% forested Saint-Maurice River (166 kg N km−2 year−1) was well below that reported for the coast of western Norway (332 kg N km−2 year−1) whereas the N exportation rate for heavily farmed (54.5% of area) Yamaska River (1579 kg N km−2 year−1) was above that documented for the East English and Scottish Coast (1500 kg N km−2 year−1) to the North Sea (Sutton et al. 2011). Overall, nitrogen and phosphorus contributions from tributaries of the SLR largely exceeded municipal sources and should be the focus of future remedial actions.
Our estimates of domestic wastewater loads did not include (1) inflow of raw sewage (storm overflows) from older combined urban systems and (2) populations not connected to a municipal sewer system. Domestic loads originating from the (largely rural and semi-urban) population scattered throughout the SLR watershed outside of the greater Montreal metropolitan area (about 4 million people, Institut de la Statistique du Québec 2016) were mostly included in tributary loads (Table 5; Patoine 2017). Conversely, our use of the most recent years (2009–2012) to assess mean annual contribution of metropolitan urban centers against 1995–2011 SLR loads likely represented upper values of urban loads for that period given the overall trends of rising urban population and nutrient concentrations in municipal effluents. Management interventions and improvement of infrastructures to reduce episodes of raw sewer overflow must be undertaken to improve future SLR water quality in urban areas. Such measures are warranted in light of the (unquantified albeit most likely) significant contribution of overflows to C, N, and P loads and their anticipated rise with extreme rain events under climate change. Far from being local, both the chemical signature and the ecosystemic effects of the plume of the City of Montreal treated wastewater effluent were detectable several km downstream of its release point (Marcogliese et al. 2014).
With respect to C, our estimate of area-specific C outflow from the entire SLR watershed (1350 kg C km−2 year−1 for DOC only without including inorganic C) was higher than the 1202 kg C km−2 year−1 value reported by Clair et al. (2013). That value integrated the relatively low area-specific C flux for the Great Lakes (750 kg C km−2 year−1 at Wolfe Island) as well as high exportation rates for the Ottawa River (2650 kg C km−2 year−1 at Carillon) and individual tributaries (up to 5900 kg C km−2 year−1 for the 98% forested Jacques-Cartier River, Patoine 2017). High area-specific C export values (>4000 kg C km−2 year−1) are typical of humic waters of large northern Québec rivers draining the boreal forest on the Pre-Cambrian Shield (Hudon et al. 1996; Clair et al. 2013), but may also be observed in heavily farmed watersheds (this study). At Québec, mean DOC annual exports (1.39 × 106 t C year−1, Table 4) from our study were about 4 times smaller than dissolved inorganic carbon (DIC) flux (5.94–6.85 × 106 t C year−1 (1998 data, Hélie et al. 2002); our focus on the DOC compartment was justified in view of its eventual degradation in deep estuarine waters.
Our results also support the earlier finding that the N/C ratio of exportations from different rivers reflects watershed land use and the origin of riverine production (Khalili et al. 2010; Clair et al. 2013). Low N/C ratio (<0.10) reflected low N exportations relative to high C outputs from forested and wetland areas (Hudon et al. 1996) typically encountered for north shore tributaries draining the Pre-Cambrian Shield such as the Jacques Cartier (N/C = 0.05) and Saint-Maurice (N/C = 0.04) rivers (Table 5). Values of N/P ratio reflected the relative importance of different sources of N and P with the highest values of N/P recorded for the outflow of Lake Ontario (N/P = 86) and atmospheric deposition (N/P = 85). Heavily farmed watersheds (>40% of farmed areas: Chateauguay, Richelieu, Yamaska, Nicolet Sud-Ouest, Nicolet rivers) showed more variable N/P ratio values (24–51), indicative of different rates of fertilizer applications and/or different N and P leaching rate from soils under various agricultural settings.
Within-river processes mediated by primary producers
Overall, our results reveal that biological activity and hydrological conditions sharply modulate the seasonal flux of C, N, and P to the SLR estuary. Our estimation that biological plant production represented 20, 18 and 58% of C, N, and P loads (respectively) at Québec is a strong indication that, during the growth season, St. Lawrence River aquatic ecosystems assimilate and process anthropogenic nutrient inputs, effectively acting as a very large tertiary water treatment plant. Phosphorus inputs from tributaries and wastewaters stimulate growth of algae and vascular plants, leading to enhanced atmospheric carbon fixation and high primary production within the SLR. At the time of plant decay, carbon and nutrients that were fixed/assimilated during summer are eventually released, either in the water (as SPM or dissolved substances) to be flushed downstream towards the SLR estuary or lost to the atmosphere through respiration (CO2) and denitrification (N2).
At the outlet of Lake Ontario, the SLR carries low concentrations of SPM but relatively high concentrations of dissolved substances compared to other major world rivers (Meybeck and Ragu 1996). The high ratio (≈17) of mean annual DOC to POC documented for SLR (Rondeau et al. 2000) justified our focussing on DOC. The low contribution of POC to the SLR estuary is reflected by the similarity between mean DOC annual exports (1.39 × 106 t C year−1, Table 4) derived from our study and the mean TOC flux of 1.24 × 106 t C year−1 estimated by Rondeau et al. (2000). In contrast with the eutrophic River Loire in which phytoplankton represented the most important primary producer (Meybeck et al. 1998), annual production of rooted vascular plants and associated epiphytes in the SLR constitutes ≈66% of total autochthonous production (Vis et al. 2007). Given the predominance of benthic primary producers along the 550-km-long course of the SLR (Lévesque et al. 2017), it is most likely that plant decay takes place, at least partially, locally (yielding DOC) rather than being entirely exported as detritus (POC) to the estuary. The isotopic signature of organic carbon fractions indeed showed that aquatic primary producers contributed a significant proportion of total organic carbon year-round and that degradation of aquatically-produced POC into DOC was most important during summer months throughout the SLR system (Hélie and Hillaire-Marcel 2006). Exportation of the entire annual riverine primary production to the SLR estuary as POC (0.28 million t C year−1, Table 7) would increase our estimate of annual C exportation by 20%, but would only represent 5% of the total SPM (mineral and organic) load exported at Québec.
In addition, the large difference we observed between mean TP (15 µg P L−1) and TDP (6 µg P L−1) concentrations at the outlet of Lake Ontario likely results from the predominance of nutrient-rich phytoplankton cells rather than mineral particles within SPM. Although measurements of chlorophyll a concentration have been interrupted since 2004, TN and TP (and SPM) concentrations at Wolfe Island appear to be rising since 2005, suggesting that this site could reflect the changing water quality from littoral areas of Lake Ontario as well as that from the more stable oligotrophic central lake (Makarewicz et al. 2012b). Rising nutrient concentrations at Wolfe Island could therefore reflect the rising inflow of nutrients from tributaries and urban sources in the nearshore lake area, inducing the recent resurgence of Cladophora (Higgins et al. 2010; Makarewicz et al. 2012a).
On a seasonal basis, nutrient concentrations at Wolfe Island were highest in the winter and spring and declined over the course of the summer, likely reflecting the mixing-stratification cycle of central Lake Ontario waters as well as biological activity nearshore. In the Laurentian Great Lakes, rising water temperature was shown to increase water column stability and the duration of summer stratification (Magnuson et al. 1997). These physical changes lead to rising primary productivity and more severe epilimnetic DIN deficit (Makarewicz et al. 2012b). At Carillon and Québec, the occurrence of similar monthly cycles in NO3 concentrations suggests that assimilation by benthic primary producers (rooted vegetation and epiphytes) and bacterial denitrification also play a significant role in shallow and/or clear large rivers.
Calculation of net monthly flux (outflow-inflow) of the different forms of C, N, and P revealed the magnitude of biological processes taking place within the riverine segment which modulate the seasonal inflow of C, N, and P from the river to its estuary. At Québec, the low summer values of N and P flux to the SLR estuary coincided with aquatic plant growth, low river discharge and warm water temperatures (Hudon and Carignan 2008; Hudon et al. 2010). The sink effect for N and P was particularly important under low discharge summer conditions during which water retention time is maximized in the shallow, warm, well-illuminated littoral areas. Low SLR water levels were indeed shown to induce substantial shifts in the distribution and relative importance of primary producers although total river primary production remained stable between years (Vis et al. 2007). The nutrient sink effect of Lake Saint-Pierre for SPM, DOC, NO2-NO3, TP and TDP during summer months was previously documented Hudon and Carignan (2008). Bacterial denitrification taking place at the sediment–water interface of the 110 km2 sector of southern Lake Saint-Pierre alone could consume about 1.5 t N per day (NO3) during summer (Hudon and Carignan 2008). Extensive, shallow SLR wetlands areas (i.e. fluvial lakes St. Lawrence, Saint-François, Saint-Louis and Saint-Pierre) represent complex ecosystems where significant nutrient processing (assimilation-retention-transformation) take place. In a nutshell, biological activity within SLR ecosystem provides significant ecosystemic services, assimilating nutrients, reducing nitrogen output to the estuary and improving water quality during summer months. In the upper Mississippi River, biological processes were also shown to play a large role in the seasonal dynamics of N and P, taking place particularly in the backwaters and impounded areas colonized by large beds of aquatic macrophytes during summer (Houser and Richardson 2010). Minimum TN concentrations in late summer and fall in the upper Mississippi were attributed to low N inputs via precipitation and runoff and relatively high rates of N removal via bacterial denitrification owing to warm water and sediment temperatures (Strauss et al. 2006).
Contribution of freshwater C, N, and P flux to hypoxia and acidification in the Laurentian Channel
Annual flux of C, N, and P from the SLR potentially contributes to hypoxia and acidification of the lower estuary through two major processes: (1) the direct consumption of O2 for decomposition of riverine organic C and (2) the stimulation of estuarine C production through the injection of riverine N and P. For C, we combined our measurement of annual DOC flux (1.39 × 106 t C year−1) with estimations of particulate organic C (POC) for four scenarios ranked from highly conservative (presuming that phytoplankton is the major source of exported POC, ≈0.1% of SPM flux at Québec) to less conservative (exportation as detritus of all riverine C fixed annually by primary producers, ≈5% of SPM flux at Québec) (Table 7). In comparison, Pocklington and Tan (1987) estimated that POC represented 3–14% of TOC in the SLR (1981–1985). In our study, the mean DOC concentration we measured at Québec (Table 1, 3.68 ± 0.30 mg C L−1 as DOC, 1995–2011) was very close to that reported by Pocklington and Tan (1987) (3.8 mg C L−1 DOC, 1981–1985) and to the TOC concentration estimated by Clair et al. (2013) (3.7 mg C L−1 TOC, 2000–2007).
The loads of total organic C (TOC) estimated for the four scenarios ranged from 1.40 to 1.67 million t C year−1, all of which were lower than the 1.83 million t C year−1 value reported by Pocklington and Tan (1987). In turn, based on the Redfield ratio (117 mol C:170 mol O2) and assuming that all C entering the estuary was degraded within a year (Tremblay and Gagné 2009), we calculated that between 5.42 and 7.09 million t O2 would potentially be consumed annually in the estuary for the degradation of organic C exported from the SLR (Table 7). The fraction of this O2 consumption actually taking place in the bottom waters of the lower estuary, thus directly contributing to deep-layer hypoxia, however remains to be assessed.
For P and N, the relative magnitude of riverine flux was compared with values from previous studies and used to calculate the proportion of SLR flux to the total NP flux to the estuarine waters, including the contribution of upwelling and vertical mixing (Table 8). In comparison with earlier studies, annual dissolved P flux from the present study were low; depending on the estimates, P flux from the SLR represented between 7% (this study, as TDP) and 19% (Coote and Yeats 1979, as PO4) of total estuarine flux. This decrease in P flux over time might reflect remediation efforts focused on P within the SLR watershed. In contrast, our estimation of N flux was higher than those from earlier studies (Table 8); estimates of NOx flux from SLR relative to the total estuarine NOx flux range from 18% (Savenkoff et al. 2001) to 31% (this study). Discrepancies in total P and N flux between studies may stem from differences in annual discharge, the predominant use of summer measurements and/or long-term drop (P) or rise (N) in concentrations. For both P and N, however, it could be argued that the overall flux of these nutrients (as TP and TN) from the SLR to the estuary should be considered rather than focusing only on their readily available forms (PO4 and NOx) for primary producers. This argument is justifiable given the high rate of mineralization taking place within the estuary (Tremblay and Gagné 2009) and the inclusion of organic N and P fractions highly valuable as substrate for microbial heterotrophs whose respiration further consumes O2. Primary production within the turbid transition zone of the upper estuary is light-limited (Levasseur et al. 1984) and supports a rich heterotrophic microbiota (Frenette et al. 1995). Calculation of the % contribution of SLR to total estuarine flux using TP (14%) and TN (47%) markedly increased the importance or river enrichment to estuarine microbial production, making them all the more significant.
Downstream from Québec, tidal mixing of C, N, and P-rich fresh water with saltwater stimulates microbial production and induces the flocculation of terrigenous dissolved and colloidal organic matter. A significant fraction of the organic material brought from upstream eventually settles in the deep Laurentian channel (Annane et al. 2015) where rising organic content of sediment and dropping O2 concentrations have been reported (Gilbert et al. 2005; Thibodeau et al. 2006). In turn, mineralization of organic matter into CO2 contributes to estuarine acidification (Mucci et al. 2011). Understanding the magnitude of freshwater C, N, and P inputs from the SLR is the first step in the assessment of the impact of human activities in the watershed leading to the eutrophication, hypoxia and acidification hundreds of kilometers downstream into its estuary.
Conclusions
Our study showed that annual flux and seasonal dynamics of SPM and nutrients differ among the three sites we studied along the SLR, largely reflecting the natural setting and anthropogenic activities in the watershed. At the outlet of Lake Ontario, SPM and nutrients loads are low for a river of this size, owing to their low concentrations in this large lacustrine environment. Erosion of riverbed and shorelines along the 550-km-long river course accounted for most of the SPM load at Québec. Inflow of the Ottawa River (13 and 14% for N and P loads respectively at Québec) and from tributaries (20 and 27% of N and P loads respectively, at Québec) draining farmlands accounted for a disproportionately large fraction and highest area-specific nutrient loads to the SLR estuary, largely exceeding municipal sources (6% for both N and P). Direct atmospheric deposition represented a comparatively minor source (<1%) of nutrient enrichment to the SLR estuary. Carbon fixation and nutrient assimilation/transformation by aquatic primary producers (emergent and submerged macrophytes, epiphyton and phytoplankton) represented significant internal processes modulating the seasonal flux of C, N, and P to the SLR estuary. Net monthly SPM, C, N, and P loads from the SLR to its estuary were highest during the spring but were sharply reduced during the period of plant growth, turning the SLR into a temporary sink over summers of lowest river discharge. Outflow of C, N, and P from freshwaters to the estuary likely contributed significantly to deep water hypoxia and acidification, either directly through carbon mineralization (whole water column O2 consumption = 5.4 − 6.5 × 106 t O2 year−1) or indirectly, by stimulation of estuarine productivity through increased N supply (31–47%). Improved monitoring and reduction of episodes of raw sewer overflow must be achieved to further control urban point sources of nutrients. Reduction of diffuse nutrient sources, focusing on small (watersheds <1000 km2) and large south shore tributaries draining farmlands, represent the most important remedial actions to improve SLR water quality and alleviate the negative impacts to its estuary.
References
American Public Health Association (APHA) (1995) Standard methods for the examination of water and wastewater, 19th edn. American Public Health Association, Washington, DC
Annane S, St-Amand L, Starr M, Pelletier E, Ferreyra GA (2015) Contribution of transparent exopolymeric particles (TEP) to estuarine particulate organic carbon pool. Mar Ecol Prog Ser 259:17–34. doi:10.3354/meps11294
Bourgault D, Koutitonsky V (1999) Real-time monitoring of the freshwater discharge at the head of the St. Lawrence Estuary. Atmosphere-Ocean 37:203–220. doi:10.1080/07055900.1999.9649626. http://ogsl.ca/fr/debits/donnees/tableaux.html
Bricker SB, Longstaff B, Dennison W, Jones W, Boicourt K, Wicks C, Woerner J (2008) Effects of nutrient enrichment in the nation’s estuaries: a decade of change. Harmful Algae 8:21–23. doi:10.1016/j.hal.2008.08.028
Burnham KP, Anderson DR (2002) Model selection and multimodel inference: a practical information-theoretic approach, 2nd edn. Springer, New York
Carignan R, Lorrain S, Lum K (1994) A 50-yr record of pollution by nutrients, trace metals, and organic chemicals in the St. Lawrence River. Can J Fish Aquat Sci 51:1088–1100. doi:10.1139/f94-108
Clair TA, Dennis IF, Bélanger S (2013) Riverine nitrogen and carbon exports from the Canadian landmass to estuaries. Biogeochemistry 115:195–211. doi:10.1007/s10533-013-9828-2
Coote A, Yeats P (1979) Distribution of nutrients in the Gulf of St. Lawrence. J Fish Res Board Can 36:122–131. doi:10.1139/f79-021
Diaz RJ, Rosenberg R (2008) Spreading dead zones and consequences for marine ecosystems. Science 321:926–929. doi:10.1126/science.1156401
Dillon PJ, Rigler FH (1974) The phosphorus-chlorophyll relationship in lakes. Limnol Oceanogr 19:767–773. doi:10.4319/lo.1974.19.5.0767
Douglas RJW (1970) Geology and economic minerals of Canada. Geological Survey of Canada, Department of Energy, Mines and Resources, Ottawa, Ontario, Economic Report No. 1
Environment Canada (2014) Manuel des méthodes d’analyses (annexe B). Environment Canada, Québec Region, Scientific and Technical Services Section. St. Lawrence Centre, Montreal
Foglia L, Mehl SW, Hill MC, Perona P, Burlando P (2007) Testing alternative ground water models using cross-validation and other methods. Ground Water 45:627–641. doi:10.1111/j.1745-6584.2007.00341.x
Foglia L, Mehl SW, Hill MC, Burlando P (2013) Evaluating model structure adequacy: the case of the Maggia Valley groundwater system, southern Switzerland. Water Resour Res 49:260–282. doi:10.1029/2011WR011779
Frenette J-J, Vincent WF, Dodson JJ, Lovejoy C (1995) Size-dependent variations in phytoplankton and protozoan community structure across the St. Lawrence River transition region. Mar Ecol Prog Ser 120:99–110. doi:10.3354/meps120099
Gilbert D, Sundby B, Gobeil C, Mucci A, Tremblay G-H (2005) A seventy-two-year record of diminishing deep-water oxygen in the St. Lawrence estuary: the northwest Atlantic connection. Limnol Oceanogr 50:1654–1666. doi:10.4319/lo.2005.50.5.1654
Gilbert D, Rabalais NN, Diaz RJ, Zhang J (2010) Evidence for greater oxygen decline rates in the coastal ocean than in the open ocean. Biogeosciences 7:2283–2296. doi:10.5194/bg-7-2283-2010
Greisman P, Ingram RG (1977) Nutrient distribution in the St. Lawrence Estuary. J Fish Res Board Can 34:2117–2123. doi:10.1139/f77-278
Hélie J-F, Hillaire-Marcel C (2006) Sources of particulate and dissolved organic carbon in the St Lawrence River: isotopic approach. Hydrol Process 20:1945–1959. doi:10.1002/hyp.5962
Hélie J-F, Hillaire-Marcel C, Rondeau B (2002) Seasonal changes in the sources and fluxes of dissolved inorganic carbon through the St. Lawrence River—isotopic and chemical constraint. Chem Geol 186:117–138. doi:10.1016/S0009-2541(01)00417-X
Higgins SN, Pennuto CM, Howell ET, Lewis TW, Makarewicz JC (2010) Urban influences on Cladophora blooms in Lake Ontario. J Great Lakes Res 38:116–123. doi:10.1016/j.jglr.2011.11.017
Hill RB (2013) St. Lawrence River/Wolfe Island water quality monitoring program 1989/90—2006/06. Concentrations and trends. Environment Canada. Water quality monitoring and surveillance—Ontario. 40 pages + app
Houser JN, Richardson WB (2010) Nitrogen and phosphorus in the Upper Mississippi River: transport, processing, and effects on the river ecosystem. In: Romano SP, Ickes B (eds). Upper Mississippi River Research Synthesis: Forty Years of Ecological Research. Hydrobiologia 640: 71–88. doi:10.1007/s10750-009-0067-4
Howarth RW, Billen G, Swaney D, Townsend A, Jaworski N, Lajtha K, Downing JA, Elmgren R, Caraco N, Jordan T, Berendse F, Freney J, Kudeyarov V, Murdoch P, Zhao-liang Z (1996) Regional nitrogen budgets and riverine inputs of N and P for the drainages to the North Atlantic Ocean: natural and human influences. Biogeochemistry 35:75–139. doi:10.1007/978-94-009-1776-7_3
Hudon C, Carignan R (2008) Cumulative impacts of hydrology and human activities on water quality in the St. Lawrence River (Lake Saint-Pierre, Québec, Canada). Can J Fish Aquat Sci 65:1165–1180. doi:10.1139/F08-069
Hudon C, Morin R, Bunch J, Harland R (1996) Carbon and nutrient output from the Great Whale River (Hudson Bay) and a comparison with other rivers around Québec. Can J Fish Aquat Sci 53:1513–1525. doi:10.1139/f96-080
Hudon C, Lalonde S, Gagnon P (2000) Ranking the effects of site exposure, plant growth form, water depth and transparency on aquatic plant biomass. Can J Fish Aquat Sci 57:31–42
Hudon C, Wilcox D, Ingram J (2006) Modeling wetland plant community response to assess water level regulation scenarios in the Lake Ontario—St. Lawrence River Basin. Environ Monitor Assess 113:303–328. doi:10.1007/S10661-005-9086-4
Hudon C, Armellin A, Gagnon P, Patoine A (2010) Variations of water temperature and level in the St. Lawrence River (Québec, Canada): effects on three common fish species. Hydrobiologia 647:145–161. doi:10.1007/s10750-009-9922-6
Hydro-Québec (2012) Daily Ottawa River Discharge at Carillon (1995–2011). Prévision et qualité des données hydroélectriques, Division Production Hydro-Québec, Montréal, Québec. Samer Alghabra, Prévision et qualité des données hydroélectriques, Division Production Hydro-Québec, 75 boul. René-Lévesque Ouest, 9e étage, Montréal, QC Québec H2Z 1A4, personal communication
International Joint Commission (IJC) (1972) Great Lakes Water Quality. Agreement, with Annexes and Texts and Terms of Reference, between the United States of America and Canada signed at Ottawa April 15, 1972 with Appendix dated November 1973
Institut de la statistique du Québec (2016) Population du Québec, 1971–2015. http://www.stat.gouv.qc.ca/statistiques/population-demographie/structure/qc_1971-20xx.htm
Kemp WM, Testa JM, Conley DJ, Gilbert D, Hagy JD (2009) Temporal responses of coastal hypoxia to nutrient loading and physical controls. Biogeosciences 6:2985–3008. doi:10.5194/bg-6-2985-2009
Khalili MI, Temnerud J, Fröberg M, Karltun E, Weyhenmeyer GA (2010) Nitrogen and carbon interactions between boreal soils and lakes. Glob Biogeochem Cycl 24:GB4011. doi:10.1029/2009GB003668
Levasseur M, Therriault JC, Legendre L (1984) Hierarchical control of phytoplankton succession by physical factors. Mar Ecol Prog Ser 19:211–222
Lévesque D, Hudon C, James P, Legendre P (2017) Environmental factors structuring benthic primary producers at different spatial scales in the St. Lawrence River (Canada). Aquat Sci 79:345–356. doi:10.1007/s00027-016-0501-4
Magnuson JJ, Assel RA, Bouser CJ, Dillon PJ, Eaton JG, Evans HE, Fee EJ, Hall RI, Mortsch LR, Schindler DW, Quinn FH, Webster KH (1997) Potential effects of climate change on aquatic systems: laurentian Great Lakes and Precambrian shield region. Hydrol Process 11:825–872. doi:10.1002/(SICI)1099-1085(19970630)11:8<825:AID-HYP509>3.0.CO;2-G
Makarewicz JC, Booty WG, Bowen GSGL (2012a) Tributary phosphorus loading to Lake Ontario. J Great Lakes Res 38:14–20. doi:10.1016/j.jglr.2012.08.001
Makarewicz JC, Lewis TW, Boyer GL (2012b) Nutrient enrichment and depletion on the shoreside of the spring thermal front. J Great Lakes Res 38:72–77. doi:10.1016/j.jglr.2011.12.004
Marcogliese DJ, Blaise C, Cyr D, de Lafontaine Y, Fournier M, Gagné F, Gagnon C, Hudon C (2014) Effects of a major municipal effluent on the St. Lawrence River : a case study. Ambio 44:257–274. doi:10.1007/s13280-014-0577-9
MEQ-EC-Ministère de l’Environnement du Québec et Environnement Canada (2001) Évaluation du potentiel toxique des effluents des stations d’épuration municipales du Québec – Rapport final. Saint-Laurent Vision 2000, Phase III – Volet Industriel et Urbain. Rapport (136 pages) et annexes (222 pages)
Meybeck M, Ragu A (1996) River discharges to the oceans. An assessment of suspended solids, major ions, and nutrients. Environment Information and Assessment Report. UNEP, Nairobi
Meybeck M, Cauwet G, Dessery S, Somville M, Gouleau D, Billen G (1998) Nutrients (organic C, P, N, Si) in the eutrophic River Loire (France) and its estuary. Estuar Coast Shelf Sci 27:595–624. doi:10.1016/0272-7714(88)90071-6
Mills EL, Casselman JM, Dermott R, Fitzsimons JD, Gal G, Holeck KT, Hoyle JA, Johannsson OE, Lantry BF, Makarewicz JC, Millard ES, Munawar IF, Munawar M, O’Gorman R, Owens RW, Rudstam LG, Schaner T, Stewart TJ (2003) Lake Ontario: food web dynamics in a changing ecosystem (1970–2000). Can J Fish Aquat Sci 60:471–490. doi:10.1139/f03-033
Mladenov N, Williams MW, Schmidt SK, Cawley K (2012) Atmospheric deposition as a source of carbon and nutrients to an alpine catchment of the Colorado Rocky Mountains. Biogeosciences 9:3337–3355. doi:10.5194/bg-9-3337-2012
Moreira J (2011) Ouvrages de surverse et stations d’épuration. Évaluation de performance des ouvrages municipaux d’assainissement des eaux pour l’année 2010. Rapport du Ministère des Affaires municipales, des Régions et de l’Occupation du territoire (MAMROT). http://www.mamrot.gouv.qc.ca/pub/infrastructures/suivi_ouvrages_assainissement_eaux/eval_perform_rapport_2010.pdf
Mucci A, Starr M, Gilbert D, Sundby B (2011) Acidification of Lower St. Lawrence Estuary bottom waters. Atmosphere-Ocean 49:206–218. doi:10.1080/07055900.2011.599265
Nicholls KH, Hopkins GJ, Standke SJ, Nakamoto L (2001) Trends in total phosphorus in Canadian near-shore waters of the Laurentian Great Lakes: 1976–1999. J Great Lakes Res 27:402–422. doi:10.1016/S0380-1330(01)70656-9
Ouimet R, Duchesne L (2009) Dépôts atmosphériques dans les forêts au Québec: retombées actuelles et tendances au cours des 20 à 30 dernières années. Le naturaliste canadien 133:56–64
Patoine, M (2017) Charges de phosphore, d’azote et de matières en suspension à l’embouchure des rivières du Québec – 2009 à 2012, Québec, Ministère du Développement durable, de l’Environnement et de la Lutte contre les changements climatiques, Direction générale du suivi de l’état de l’environnement, ISBN 978-2-550-77490-7 (PDF), 25 pages and 11 app. [On line]. http://www.mddelcc.gouv.qc.ca/eau/eco_aqua/phosphore/charge-phosphore-azote-mes2009-2012.pdf
Pham TT, Rondeau B, Sabik H, Proulx S, Cossa D (2000) Lake Ontario: the predominant source of triazine herbicides in the St. Lawrence River. Can J Fish Aquat Sci 57(S1):78–85. doi:10.1139/f99-233
Pocklington R, Tan FC (1987) Seasonal and annual variations in the organic matter contributed by the St Lawrence River to the Gulf of St. Lawrence. Geochim Cosmochim Acta 51:2579–2586. doi:10.1016/0016-7037(87)90308-5
Raymond S, Moatar F, Meybeck M, Bustillo V (2013) Choosing methods for estimating dissolved and particulate riverine fluxes from monthly sampling. Hydrol Sci J 58:1326–1339. doi:10.1080/02626667.2013.814915
Rondeau B (1998) Validation d’une station de référence pour le suivi de la qualité des eaux dans le Saint-Laurent à Québec. Environnement Canada – Région du Québec, Conservation de l’environnement, Centre Saint-Laurent. Rapport scientifique et technique ST-175
Rondeau B, Cossa D, Gagnon P, Bilodeau L (2000) Budget and sources of suspended sediments transported in the St. Lawrence River, Canada. Hydrol Proc 1:21–36. doi:10.1002/(SICI)1099-1085(200001)14:1<21:AID-HYP907>3.0.CO;2-7
Rondeau B, Cossa D, Gagnon P, Pham TT, Surette C (2005) Hydrological and biogeochemical dynamics of the minor and trace elements in the St. Lawrence River. Appl Geochem 20:1391–1408. doi:10.1016/j.apgeochem.2005.02.011
Savenkoff C, Vézina A, Smith P, Han G (2001) Summer transports of nutrients in the Gulf of St. Lawrence estimated by inverse modelling. Estuar Coast Shelf Science 52:565–587. doi:10.1006/ecss.2001.0774
Seitzinger S, Harrison JA, Böhlke JK, Bouwman AF, Lowrance R, Peterson B, Tobias C, Drecht GV (2006) Denitrification across landscapes and waterscapes: a synthesis. Ecol Appl 16:2064–2090. doi:10.1890/1051-0761(2006)016[2064:DALAWA]2.0.CO;2
Stainton MP, Capel MJ, Armstrong FAJ (1977) The chemical analysis of freshwater, 2nd ed. Can Fish Mar Set Misc Spec Publ
Starr M, Lair S, Michaud S, Scarratt M, Quilliam M, Lefaivre D, Robert M, Wotherspoon A, Michaud R, Ménard N, Sauvé G, Lessard S, Béland P, Measures L (2017) Multispecies mass mortality of marine fauna linked to a toxic dinoflagellate bloom. PLoS ONE 12(5):e0176299. doi:10.1371/journal.pone.0176299
Stevens RJJ, Neilson MA (1987) Response of Lake Ontario to reductions in phosphorus load, 1967–82. Can J Fish Aquat Sci 44:2059–2068. doi:10.1139/f87-255
Strauss EA, Richardson WB, Cavanaugh JC, Bartsch LA, Kreiling RM, Standorf AJ (2006) Variability and regulation of denitrification in an Upper Mississippi River backwater. J North Am Benthol Soc 25:596–606
Sutton MA, Howard CM, Erisman JW, Billen G, Bleeker A, Grennfelt P, van Grinsven H, Grizzetti B (2011) European nitrogen assessment: sources, effects and policy perspectives, Chap 13, pp 271–297. In: Nitrogen flows from European regional watersheds to coastal marine waters. Lead author: Gilles Billen Cambridge University Press, Cambridge, US
Thibodeau B, de Vernal A, Mucci A (2006) Recent eutrophication and consequent hypoxia in the bottom waters of the Lower St. Lawrence Estuary: micropaleontological and geochemical evidence. Mar Geol 231:37–50. doi:10.1016/j.margeo.2006.05.010
Tremblay L, Gagné JP (2009) Organic matter distribution and reactivity in the waters of a large estuarine system. Mar Chem 116:1–12. doi:10.1016/j.marchem.2009.09.006
Ville de Montréal (2012) Concentrations of DOC, TKN, NO2-NO3 (2012) and NH3 (2000-2012) measured in the treated effluent of the City of Montreal WWTP. Carole Fleury and Tony di Fruscia, Service de l’eau - Ville de Montréal, Direction de l’épuration des eaux usées, 12001 Maurice-Duplessis, Montréal, QC H1C 1V3, Canada, personal communication
Vis C, Hudon C, Carignan R, Gagnon P (2007) Spatial analysis of production by macrophytes, phytoplankton and epiphyton in a large river system under different water-level conditions. Ecosystems 10:293–310. doi:10.1007/s10021-007-9021-3
Walker, WW (1996) Simplified procedures for eutrophication assessment and prediction: User manual. Instruction Report W-96-2. (Updated in April 1999). U.S. Army Engineer Waterways Experiment Station Vicksburg, MS. https://wiki.umn.edu/pub/Nieber/WebHome/FLUX-Bathtub_MANUAL_IR-W-96_2.pdf
Wallace RB, Baumann H, Grear JS, Aller RC, Gobler CJ (2014) Coastal ocean acidification: the other eutrophication problem. Estuar Coast Shelf Sci 148:1–13. doi:10.1016/j.ecss.2014.05.027
Weise AM, Levasseur M, Saucier FJ, Senneville S, Bonneau E, Roy S, Sauvé G, Michaud S, Fauchot J (2002) The link between precipitation, river runoff, and blooms of the toxic dinoflagellate Alexandrium tamarense in the St. Lawrence. Can J Fish Aquat Sci 59:464–473. doi:10.1139/f02-024
Wetzel RA, Likens GE (1991) Limnological analyses, 2nd edn. Springer, New-York
Yates AG, Culp JM, Chambers PA (2012) Estimating nutrient production from human activities in subcatchments of the Red River, Manitoba. J Great Lakes Res 38:106–114. doi:10.1016/j.jglr.2011.04.009
Ye M, Meyer PD, Neuman SP (2008) On model selection criteria in multimodel analysis. Water Resour Res 44:W03428. doi:10.1029/2008WR006803
Acknowledgements
We thank François Boudreault for drafting the map and the staff from the Environment and Climate Change Canada Laboratoire des Essais Environnementaux (ECCC-LEEQ) for their help with water quality data acquisition. Samer Alghabra (Hydro-Québec), Rob Caldwell and Sylvain Martin (ECCC) kindly supplied Ottawa River and SLR discharge data. The assessment of inflow from treated urban sources would not have been possible without the help of staff members from the City of Montreal, particularly Carole Fleury, Tony Di Fruscia and Guy Deschamps. Manon Harwood and Shirley-Anne Smyth from ECCC supplied data for small wastewater treatment plants. We thank M. Duchemin, M. Gélineau and D. Labrie (MDDELCC) for supplying municipal TP loads corrected for seasonal dephosphatation. The constructive comments of Yves de Lafontaine and four anonymous referees on earlier versions of the manuscript are acknowledged with thanks. This study is a contribution to the St. Lawrence Action Plan (Canada-Québec).
Author information
Authors and Affiliations
Corresponding author
Additional information
Responsible Editor: Sujay Kaushal.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.
About this article

Cite this article
Hudon, C., Gagnon, P., Rondeau, M. et al. Hydrological and biological processes modulate carbon, nitrogen and phosphorus flux from the St. Lawrence River to its estuary (Quebec, Canada). Biogeochemistry 135, 251–276 (2017). https://doi.org/10.1007/s10533-017-0371-4
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10533-017-0371-4