
Abstract
At the end of 2020, 18 European bison (Bison bonasus) were introduced to El Encinarejo estate, in the south of the Iberian Peninsula. This hunting-oriented estate has 956 hectares of dehesa, Mediterranean forest and scrubland in which about 400 red deer (Cervus elaphus) and 200 fallow deer (Dama dama) coexist with the bison. To know how trophic resources are used by these herbivore species, faecal samples were collected during the year 2021. The microhistological analysis technique was used to determine the percentage of epidermal fragments of the main functional groups (woody plants, graminoids, legumes and forbs) in each sample. The results showed significant differences between animal species and seasons of the year. Globally, woody plants are preferentially consumed in autumn and winter, graminoids in summer and legumes and forbs in spring. The woody component was the most abundant in the diet of the three species, accounting for 81% of the plant fragments present in red deer faeces, 52% in bison and 38% in fallow deer. This is the first study describing the dietary use of Pistacia lentiscus by European bison, the substantial use throughout all seasons indicates it as a very important diet source. In addition, the interspecific comparison showed that the bison significantly consumed more graminoids (21%), whereas legumes were more present in fallow deer diet (32%). Finally, red deer consumed less forbs than the other two species (8%). It is concluded that there is a certain distribution of trophic resources between the three species that may facilitate their coexistence.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Large herbivores consistently exert top-down regulatory influence on plant demography, species composition, and biomass within terrestrial ecosystems. Their enduring impact, spanning millions of years, has significantly shaped Earth’s ecosystems, influencing biogeochemistry, vegetation dynamics, ecological communities, and evolutionary processes (Trepel et al. 2024). These effects are intricately governed by the interplay between diverse species of large herbivores and their respective environments. Environmental, evolutionary, and historical factors collectively determine the coexistence patterns of these animals in a given area (Tokeshi 2009).
To comprehend the ecological adaptations and mechanisms that mitigate competition for resources and facilitate coexistence, it is imperative to conduct comparative studies on sympatric species (Zhou et al. 2013). Among the mechanisms enabling the coexistence of sympatric herbivores with similar resource requirements, the strategic distribution of trophic resources plays a pivotal role. It has been postulated that if multiple herbivores exhibit analogous spatial utilization and activity patterns, there would be differential consumption of plant species among these species (Li et al. 2022). This phenomenon is believed to have occurred in the Pleistocene in Europe, where certain fossil records indicate resource partitioning in distinct localities among large herbivores (Rivals and Lister 2016). Nevertheless, the diversity and abundance of European megaherbivores experienced a decline since the late Pleistocene, attributed to factors such as habitat loss, hunting, and displacement by livestock.
Among the surviving megaherbivores in Europe are the European bison (Bison bonasus), red deer (Cervus elaphus), and fallow deer (Dama dama). However, their cohabitation in contemporary habitats has become rare, likely influenced by human-induced factors such as habitat fragmentation, exploitation, and translocation (Ellis 2011; Baker et al. 2017; Queirós et al. 2019; Neov 2021). In the Iberian Peninsula, deer and fallow deer have historically shared territories for centuries. Presently, due to the decline in extensive livestock farming and the growing interest in rewilding pastures, there is a nascent introduction of the European bison in some private states and reserves. This development provides an opportunity for studies aimed at comprehending how ecosystems respond to the shifting populations of megaherbivores in an evolving environment.
In the beginning of the Holocene, the European bison was distributed throughout western, central, and southeastern Europe, where the Balkan and Caucasian ridges operated as southern borders, at around 40 degrees latitude (Neov et al. 2021). The species suffered from a gradual population decline since the Neolithic period, probably due to hunting (Patou-Mathis 2000; Ellis 2011), competition with livestock and loss of suitable habitats (Pucek 2004). The species survived in a few northern areas and eventually became extinct in the wild at the beginning of the 20th century. Current populations are the result of the recovery of a few zoo specimens that were carefully maintained and eventually reintroduced to the Polish side of the Białowieża Forest (Slatis 1960). The species is included in Appendix III of the Berne Convention, in Annexes II and IV of the Habitats Directive, and it has recently been reclassified as Near Threatened in the IUCN Red List of Threatened Species (Plumb et al. 2020) after recovering from Vulnerable and Endangered statuses since 1988. The global population at the end of 2020 counted 9100 individuals with over 6800 living in the wild (Olech and Perzanowski 2022).
The wide historical geographic distribution throughout the European continent of the European bison (Benecke 2005; Onar et al. 2017) as well as the success of ex situ populations in different European territories have revealed the capacity and plasticity of the species to adjust to new habitats and to a wide bioclimatic range (Hartvig et al. 2021; Kowalczyk et al. 2021). The creation of ex situ breeding sites in semi-free conditions has become a strategy to guarantee the survival of the species (Kaczmarek-Okrój and Olech 2022) and allows to apply its potential to restore and enhance the biodiversity of these areas (Kowalczyk et al. 2021; Dvorský et al. 2022). However, the introduction of the species in territories where there is no clear evidence of its presence in the past generates controversy. This is the case of the Iberian Peninsula (see, e.g., Massilani et al. 2016; Sorbelli et al. 2021), where the presence of the steppe bison (Bison priscus), the aurochs (Bos primigenius, ancestors of modern cattle) and Bison schoetensacki are well documented from fossil records, although, to date, not the presence of the European bison (but see Spassov and Stoytchev 2003).
Recent genetic analyses using ancient DNA records suggested that the European bison could be the result of a hybridization process between the steppe bison and the aurochs (Soubrier et al. 2016, but see Grange et al. 2018), and given that both species were widely represented in the pre-Neolithic Iberian plains, it would seem very plausible that this hybridization process also took place in the Iberian Peninsula, which would lead to a hypothetical presence of the European bison, despite no fossil records have been identified as such yet. On the other hand, and as an alternative hypothesis, genome data on the extinct B. schoetensacki (present in the Iberian fossil record) published in 2017 established it as a sister species of the European bison (Palacio et al. 2017) and provides genetic evidence supporting the continuous presence of B. schoetensacki up to the Upper Pleistocene. This publication (Palacio et al. 2017) assumes a possible evolution and recolonization of Europe from B. bonasus starting in Iberia and Southern France, where both species, B. bonasus, and B. priscus, were found living contemporaneously.
As evidence is non-conclusive yet and given the risk of introducing a potentially exotic wild species, current Spanish legislation only allows the presence of the European bison in fenced areas (Royal Decree 804/2011, of June 10, of the Spanish Government). Among the arguments put forward against the introduction of the European bison in the wild are doubts about its ability to adapt to Mediterranean environments and its possible resource competition with other large herbivores native to the Iberian Peninsula (Varillas 2013).
The objective of this work was to determine the seasonal diet composition of a herd of European bison introduced in a Mediterranean climate ecosystem located in the south of the Iberian Peninsula and compares it with the diet of sympatric populations of red deer and fallow deer, in order to detect their potential overlap or complementarity in the use of food resources.
Materials and methods
Study area and study populations
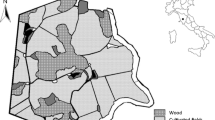
The study area is located at El Encinarejo estate (38o 9’4”N, 4o 0’3”W) in the province of Jaén (Spain), on the southern edge of the Sierra de Andújar Natural Park (Fig. 1). It is a hunting-oriented estate since the beginning of the 20th century, dedicated to hunting red deer, fallow deer and wild boar. The estate, like many other large hunting estates in the Iberian Peninsula, is fenced all around, permeable for wild boar but not for the three species under study. At the time of the study, the population of large herbivores was estimated at about 400 red deer and 200 fallow deer. In 2019 it was acquired by the current owners a herd of 18 individuals which was introduced at the end of 2020 in cooperation with the European Bison Conservation Centre—Spain, and under a legal status of zoological nucleus for the conservation of the endangered European bison.

Location of the El Encinarejo farm in the Iberian Peninsula. The perimeter of the property is shown in yellow
The study site in which all three study species roam covers 956 hectares of oak savanna, the so-called dehesa in southern Spain, with an orography that varies between 300 and 600 m asl. The soil is acidic and of granitic origin. Holm oak (Quercus ilex) is the dominant tree in this formation, maintaining a density of 20 to 50 oaks per hectare. The ground is covered by grass in more than 90% of the surface. Graminoids (grass-like species) dominate, and annual herbaceous plants are abundant, among which legumes of Trifolium genus stand out. The shrub layer covers a quarter of the surface and is dominated by Pistacia lentiscus and some labiates, such as Lavandula latifolia and Salvia rosmarinus.
One of the main limiting factors affecting the reproduction level of ex situ European bison populations is the availability of food resources. Thus, fodder supplements are sometimes necessary as an alternative to the natural migratory movements that bison may carry out in free-living conditions (Kowalczyk et al. 2013). At El Encinarejo estate, supplementation for the three animal species has been carried out throughout the year, except in spring, and mainly during the driest period, a usual management measure in large hunting estates in the Mediterranean region (Miranda et al. 2015). This contribution included the summer because that year the temperatures were very high (reaching 40ºC) and the grass withered very early. This supplementation consisted of vitamin-enriched concentrate compound pellets and trace element correctors provided in daily quantities of 60 kg distributed around the farm.
Sample collection
Five faecal samples per animal species (European bison, red deer and fallow deer) were collected in the field during the four seasons in 2021. Sample collection was undertaken at ground points where the presence of animals was previously observed, and whenever possible after observing an individual defecate, ensuring samples were as fresh as possible. Otherwise, faeces collected were those considered fresh based on their coloration and dampness, in order to ensure a deposition of no more than 12 h. Each sample contained about 100 g of faecal matter that was placed in individual plastic bags and frozen at -20ºC until processed in the laboratory. In total, 60 samples were analysed.
Microhistological analysis
For the analysis of the botanical composition of the diet, the microhistological technique was chosen since it has been used successfully, both in European bison (Forte et al. 2012), red deer (Maia et al. 2003; Krojerová-Prokešová et al. 2010) and fallow deer (Putman et al. 1993; Morse et al. 2009). The limitations of the method for accurate quantitative determination of species composition of ingested forage have been recognized by Holechek et al. (1982). As a rule, the digestion process has little effect on the epidermis of perennial plants (Storr 1961), but in annual plants, particularly forbs, the epidermal cells can be eroded to the extent that many fragments become unrecognizable (e.g. Stewart 1967). Despite this, many epidermal fragments of annual species can be identified at the group level (e.g. forbs or legumes). This technique is effective for determining the percentage of the botanical composition of the diet without attempting to quantify the biomass ingested by the animals, for which correction factors are required (Holechek et al. 1982).
After field collection and storage, frozen faeces were thawed and oven-dried at 60 °C for 24 h and then ground with a 1-mm pitch laboratory mill (Cyclotec 1093, FOSS Tecator, Höganäs, Sweden). Between 2 and 3 gr of each faecal sample was mixed with 3 ml of 65% concentrated HNO3 in test-tubes following the procedure used by Stewart (1967). These test-tubes were placed in a thermal bath at 80 °C for 2 min and then diluted into beakers with 200 ml of distilled water. This suspension was then passed through 1.00 mm and 0.125 mm filters. To blanch the epidermal fragments, the material collected on the 0.125 filter was then deposited in a test-tube with 3 ml of NaClO for 30 min. The contents of the tubes were emptied into beakers with 200 ml of distilled water and sieve again with a 0.125 mm filter. A subsample of the material obtained in the filter was homogeneously distributed onto three slides with 50% glycerine trying to avoid any fragment overlap. Finally, cover-slips (24 × 60 mm) were sealed with DPX resin (Herter Instruments, Barcelona, Spain). Preparations were observed under an optical microscope (Motic BA210, MoticEurope SLU, Barcelona, Spain) at 100 and 400 magnifications, making linear transects until a total of 200 epidermal fragments per sample were identified. Identification of epidermal fragments was based on the shape of epidermal cells, trichomes and stomata surrounding cells and a photographic repository of plant epidermis was used (available on the website: https://ddd.uab.cat/collection/atlepi?ln=es). When trichomes detached from the epidermis appeared in a field, all of them were counted as a single fragment. Epidermal fragments of floral parts, stems, petioles, and midribs were not registered due to difficulties in species identification. Recorded plants were pooled into four functional groups, namely: woody plants, graminoids, forbs and legumes. The two most abundant woody plants, Q. ilex and P. lentiscus, were also included in our analyses at the species level.
The microhistological analysis was performed at the animal production laboratory of the Veterinary Faculty of the Universitat Autònoma de Barcelona (UAB). This laboratory is part of the ASFAC-LAB quality program, consisting of an exercise of intercomparison of laboratories performing analysis of feed and raw materials for animal feed (http://www.asfaclab.com/en/).
Statistical analysis
To determine diet composition differences between the study species, a parametric ANOVA test was carried out through a standard least square fit model. Factor variables were Species, Season and Species x Season, and response variables were the proportion of the four functional plant groups and the two most abundant woody plants present in the study area (see above). Arcsine square root transformations were made to accomplish normality and the significance level was established at 0.5. The statistical program used was JMP 8 for MacOS.
An index of diet overlap (Os) between animal species (s) was calculated using the following formula:
where Dis is the proportion of one plant resource (i) in one animal species (s).
The overlap of one plant resource (Oi) in the compared diets of a pair of animal species can then be written as:
This is equivalent to the commonly used Kulczynsky similarity index (Oosting 1956).
Results
Differences in fragment composition according to animal species and season were highly significant for all plant components (Table 1). The interaction between animal species and season was significant in the categories of forbs, P. lentiscus and Q. ilex.
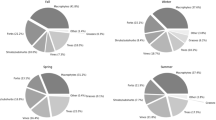
Figure 2 shows the seasonal diet composition of the three studied herbivores, estimated from the microhistological analysis of their faeces. In general, graminoids were more consumed by bison (bison 21%, red deer 5% and fallow deer 5%, on average), woody plants by red deer (red deer 81%, bison 52% and fallow deer 38% on average), and legumes by fallow deer (fallow deer 32%, red deer 6% and bison 4% on average). The woody component in the diets presented its highest values in autumn and winter for the three animal species, however red deer consumed significantly more than the other herbivores in summer and autumn when its diet almost solely consisted of woody plants. During these two seasons there were no differences between the European bison and fallow deer, but in winter the latter consumed significantly less woody plants than the other two herbivores. In the specific case of Q. ilex the pattern was maintained in summer and autumn, when red deer is the main consumer, but it differed in spring, when red deer and bison consumption was very similar, and in winter, when bison consumption was significantly higher than that of red deer. As for the consumption of P. lentiscus, a unique pattern was observed for each species. In summer it was clearly more consumed by red deer and in autumn by bison. Fallow deer was the one with the lowest values regarding this plant species, which was significant in winter and spring. The European bison showed a similar pattern in terms of in which season which proportion was consumed of both woody species; highest in autumn and winter, lowest in spring and summer. The graminoids presented the highest consumption values in summer for the three species, but consumption by bison was significantly higher during most of the year and this was especially the case in summer. Consumption of this resource by both deer species remained consistently low throughout the year.

Average seasonal percentage of plant fragments identified in faecal samples of sympatric European bison, red deer and fallow deer in El Encinarejo estate (southern Spain). Different letters indicate significant differences between animal species and seasons
The consumption of legumes characterized the diet of fallow deer, which was significantly higher than any of the other two herbivores throughout the entire year, with peaks in winter and spring. For this plant type, no differences were observed between bison and red deer. Forbs were significantly more consumed by fallow deer and bison, most notably in spring and summer. Red deer showed the lowest values for this resource during all four seasons of the year.
In relation to the overlap index of diets between the different animals (Os), Table 2 shows relatively high values between the European bison and the two species of deer only during spring. However, the annual averages show moderate overlap in the three pairs of comparisons.
The overlap for each type of plant resource is shown in Table 3. The highest values of the overlap index (Oi) for the three pairs of comparisons occur in the case of Q. ilex. Forbs show a high overlap between European bison and fallow deer, P. lentiscus between European bison and red deer and graminoids between red deer and fallow deer. The rest of the comparisons show low or moderate overlap values.
Discussion
The European bison, Europe’s largest terrestrial wild mammal, can play an important role in the management and conservation of pastures, being a keystone species like its counterpart, the American bison (Bison bison), in North American prairies (Knapp et al. 1999). That is why understanding the European bison foraging habits, particularly their herbivory impact on grasslands and other functional plant groups, is crucial to understand their ecological role in habitats where it coexists with other large herbivores, such as deer. The results obtained in this study show that, in a Mediterranean environment, European bison vary their diet seasonally, adapting to resource availability, a trend observed in studies carried out in other European territories hosting herds of this species (Cromsigt et al. 2018; Zielke et al. 2019). Red deer and fallow deer would adapt their diet to the presence of the European bison, differing in the consumption of certain resources that satisfy their needs and at the same time allow them to coexist in sympatry. In general, the three species consumes more woody plants in autumn and winter and more herbaceous plants in spring and summer, when the availability of forbs is higher. This feeding pattern is consistent with Hoffman’s classification of intermediate feeders for these three species (Hofmann 1989) and also with previous diet studies (Spitzer et al. 2020). However, in this classification the species of the Bovine subfamily would be the most grazers, although the bison is the least. This is in line with Mendoza and Palmqvist (2008) who postulate that the European bison might have evolved in grasslands or mixed habitats. This group of animals maintains a preference for graminoids when they are available. This could explain that although its diet is dominated by woody plants, the fraction of graminoids is higher than in the other two species (both belonging to the Cervidae family) and the level of overlap for this resource is low. In this sense, the proportions of the components of bison diet differ notably with respect to those of both deer species. The importance of graminoids in the diet of the European bison contrasts with the relative low frequency of this resource observed in the diet of the American bison according to Bergmann et al. (2015), although it coincides with former bison studies carried out in other North American territories (Reynolds et al. 1978; Coppedge et al. 1998). In line with Zielke et al. (2019), in winter the European bison behaves like a browser, with a predominant consumption of woody plants. In our study this was like red deer, with the particularity that the former consumes more Q. ilex in that particular season. In summer and autumn however, the proportion of Q. ilex is much higher in red deer diet. This plant species is widely used by large herbivores in the Mediterranean region (e.g., Díaz et al. 1997) and this would explain the high degree of overlap observed in the use of this resource by the three species of ungulates. The European bison is known to forage from Quercus sp. (Krasińska and Krasiński 2013; Hartvig et al. 2021; Kowalczyk et al. 2019), therefore it is no surprise that it utilizes this food resource in the Mediterranean dehesa as well, as the species is known for its high plasticity (Kowalczyk et al. 2019). The European bison, on the other hand, is considered much more of a browser than the American bison (Kowalczyk et al. 2011; Merceron et al. 2014), but also in the latter a temporary browsing behaviour has been observed (Waggoner and Hinkes 1986; Painter and Ripple 2012; Hecker et al. 2021). Craine et al. (2015) postulates that when the American bison is subjected to greater nutritional stress in warmer climates, where grass protein concentrations are reduced, it may have to attempt to compensate by browsing more at the expense of grazing. This strategy could be even higher in the European bison in the Mediterranean environment, and this would explain the fact that more than half of their diet is made up of the woody fraction. Interestingly, during summer, the European bison diet is quite evenly distributed by the proportions of graminoids, woody plants and forbs. Contrary to our results, a previous study on European bison diet, carried out in the National Park Zuid-Kennemerland in the Netherlands, showed an expected much larger proportion of graminoids and, though substantial, a smaller proportion of woody species, throughout seasons (Cromsigt et al. 2018). However, in this area the vegetation is dominated by dry and wet grasslands, and shrub land characterized by a continuous grass layer that do not suffer the water stress of the Mediterranean summer. These differences would indicate that the European bison adapts to very different habitats, varying the composition of its diet depending on the availability of resources, which in the case of the south of the Iberian Peninsula would be the sclerophyllous woody species.
As for red deer, previous diet studies indicated a higher proportion of graminoids compared to our findings. Specifically, the review carried out by Gebert and Verheyden (2008) showed that graminoids represent almost a third of the diet of the red deer in all the different habitats studied. These authors suggested that even if graminoids are not the most important resource, their availability is not limiting for red deer. This type of animal avoids highly fibrous plants, such dry graminoids in summer, as long and as much as possible. This explains that in conditions of sympatry the red deer barely uses this resource, much more exploited by the European bison. In the case of fallow deer, a study carried out by Jackson (1977) in New Forest (Hampsire, UK), where they live in sympatry with other ungulates, showed that graminoids constituted most of the diet (50–75%) during spring and summer, while woody species dominated the rest of the year. It should be noted that most of these woody species are dwarf shrubs, such as Calluna vulgaris, which indicates that this animal, even in forest habitats, prefers to graze in the lower strata of the vegetation. A later study in the same region, carried out by Putman (1986), concluded that fallow deer were primarily grazers, with a 60% of graminoids in their diets. The dominance of graminoids in the diet of fallow deer has also been observed in the forests of Poland (Borkowski and Obidziński 2003). Also in Poland, Obidziński et al. (2013) showed a much higher proportion of graminoids in autumn and winter diets of fallow deer compared to red deer, and, in line with our findings, a substantial lower proportion of browse. In our case, the presence of the European bison, as the largest consumer of graminoids, could explain the low consumption of these species by deer, in accordance with the low overlap in the use of this resource in their diets. Our results also show that fallow deer and European bison consume practically the same proportion of forbs throughout the year, which results in a high degree of overlap, and in greater quantities than red deer. A comparison study by Garcia-Gonzalez and Cuartas (1992) in a nearby region (Sierra de Cazorla, Spain) showed a similar seasonal variation of forbs for fallow deer, and a similarly much lower proportion of this forage for red deer. One of the aspects to highlight about our work is that the fallow deer has a higher proportion of legumes in its diet showing the lowest values of overlap with the two species for this resource. This would indicate that the fallow deer, under conditions of sympatry, can take better advantage of a good quality resource than the other two species, such as legumes in general.
In a previous study comparing the diet of sympatric red deer and fallow deer in the same mountain range (Sierra de Andújar), Azorit et al. (2012) also indicated the significance of woody plants in their diets, although their results, based on macroscopic plant features obtained from rumen samples, showed that grasses constituted the main component in the annual diet of both deer species. In addition, they pointed out that red deer consumed more woody plants than fallow deer, as also has been observed by García-González and Cuartas (1992) and summarized by Spitzer et al. (2020). Bearing this in mind, our results allow us to postulate the hypothesis that the introduction of the European bison into the same landscape might accentuate the browsing versus grazing behaviour of, respectively, red and fallow deer, resembling the intermediate role played by bison in Holocene times, as it was detected in Early Holocene sites in northern Europe by Bocherens et al. (2015), who found that the European bison diet can be situated between that of a typical grazing herbivore, the aurochs (Bos primigenius), and a typical browser, the moose (Alces alces).
Among the woody plants consumed by the three studied herbivores, it is worth highlighting the presence of P. lentiscus. This is the first study describing the dietary use of P. lentiscus by European bison, a plant species found in the Mediterranean and Middle East regions, also known as mastic, a shrub rich in secondary compounds, mainly tannins (Landau et al. 2010), which appears in relatively large quantities in both European bison and red deer diet. In this case, both species have the highest overlap value recorded by any other resource. This shrub appears in the autumn and winter diet of the European bison with a consumption of 30%, also reaching that figure in red deer winter and summer diet. The substantial proportion of that plant species indicate high dietary adaptability of European bison especially considering that the species is known for its limited selectivity, compared to red deer which is more of a selective feeder (Krasińska and Krasiński 2013). It is known that (mule) deer produce a tannin-binding protein in their saliva for overcoming adverse effects of tannins (Austin et al. 1989). As for the European bison, to the best of our knowledge such a protein has not been described, and different postulations can be put forward to explain their tolerance to tannins: (a) the production of a tannin-binding protein, (b) a natural tolerance to these secondary compounds, or (c) their consumption as a self-medication behaviour, as it has been evidenced in other herbivore species (Lisonbee et al. 2009). Fallow deer, however, incorporates this shrub only in a small proportion in its diet throughout the year. As this is the first diet study for the European bison in the south of Iberian Peninsula, this finding adds to the general understanding of the feeding capabilities of the species when found in a Mediterranean environment occupied by other large herbivores. In line with other scientific studies that have provided a wider understanding about European bison diet and habitat use in the past decade, our study shows that the species is able to cope with other conditions than previously widely recognized as being suitable for the species, which has important implications for the species further conservation (Kerley et al. 2012).
In the context of interspecific competition, a pronounced dietary congruence between the European bison and cervids has been observed exclusively during the spring season. It is noteworthy, however, that a substantial dietary overlap does not inherently signify competitive interactions. Conversely, during periods of resource abundance, such as in the spring, the observed overlap is more indicative of the equitable distribution of available resources among these species (Azorit et al. 2012).
Another aspect to consider is the ecosystem services that large herbivores provide. In this sense, Spain is projected to experience the greatest agricultural land abandonment in the European Union with more than 1 million ha over the course of 2015–2030 (Castillo et al. 2018). Van Wieren (1995) highlighted the potential of grazing and trampling by large herbivores as a source of reduction in fire risk for abandoned, dry territories in southern Europe. Lasanta et al. (2018) reported on the positive impact that extensively managed livestock together with shrubland clearing had in La Rioja, northeastern Spain, in regard to prevention and control of wildfires. According to Valdés-Correcher et al. (2018), European bison could be a substitute for cattle to use in natural management to keep encroachment in check. Also, recently the contribution of deer in fuel reduction has been highlighted by controlling tree and shrub densities (Hanberry and Faison 2023). A comparison by Kowalczyk et al. (2021) on the impact of large wild herbivores including red deer shows that the European bison is most effective in keeping encroachment limited. In our opinion, given the ecological similarities between the European bison and domestic cattle looking at the plant group level (Cromsigt et al. 2018), the former may well play a crucial role into the conservation of dehesas in the Iberian Peninsula, currently threatened due to the decline in extensive livestock farming (Pinto-Correia and Godinho 2013). Recently, Wang et al. (2023) highlighted the benefits of rewilding over afforestation of abandoned farmland, a topic that certainly applies to the Mediterranean landscapes and specifically Spain. The presence of the European bison, a species never domesticated, fits well in this rewilding frame. Also, in regard to the risk of wildfires, the potential of wild herbivores, such as the European bison and deer, in a rewilding approach should not be ignored (Rouet-Leduc et al. 2021). Since P. lentiscus is a flammable species, our results indicate that both red deer and European bison might have a positive effect in decreasing the risk of fire, especially in summer and autumn, in line with findings of Mancilla-Leytón et al. (2021) in Doñana Natural Park, where herbivory control the structure of the shrub land.
As a conclusion, this study shows the European bison is capable to adjust its foraging behaviour to a Mediterranean ecosystem, given there is a variety of trophic resources available, both spatially as seasonally. In addition, when added to a species assemblage that include red deer and fallow deer, or potentially species that fall into the same feeding types as these two species, coexistence is to be expected though species specific foraging behaviours might get more accentuated into browsing (red deer), and grazing (fallow deer). The moderate annual values of overlap observed between the diets of the three species would fit with a situation of coexistence with resource sharing. Beyond the fact that the European bison inhabited the Iberian Peninsula in the past or not, the time has come to listen to the words of Davis et al. (2011) saying that it is time to implement effective conservation measures, and primarily focus on ecosystem functions of species instead of on their origin.
References
Austin PJ, Suchar LA, Robbins CT, Hagerman AE (1989) Tannin-binding proteins in saliva of deer and their absence in saliva of sheep and cattle. J Chem Ecol 15(4):1335–1347. https://doi.org/10.1007/bf01014834
Azorit C, Tellado S, Oya A, Moro J (2012) Seasonal and specific diet variations in sympatric red and fallow deer of southern Spain: a preliminary approach to feeding behaviour. Anim Prod Sci 52(8):720–727. https://doi.org/10.1071/AN12016
Baker K, Gray H, Ramovs, Mertzanidou D, Akın Pekşen Ç, Bilgin CC, Sykes N, Hoelzel AR (2017) Strong population structure in a species manipulated by humans since the neolithic: the European fallow deer (Dama dama dama). Heredity 119:16–26. https://doi.org/10.1038/hdy.2017.11
Benecke N (2005) The Holocene distribution of European bison - the archaeozoological record. MUNIBE (Anthropologia-Arkeologia) 57:421–428 Brehm-Bücherei 105. Leipzig
Bergmann GT, Craine JM, Robeson IIMS, Fierer N (2015) Seasonal shifts in Diet and Gut Microbiota of the American Bison (Bison bison). PLoS ONE 10(11):e0142409. https://doi.org/10.1371/journal
Bocherens H, Hofman-Kamińska E, Drucker DG, Schmölcke U, Kowalczyk R (2015) European Bison as a Refugee species? Evidence from Isotopic Data on Early Holocene Bison and other large herbivores in Northern Europe. PLoS ONE 10(2):e0115090. https://doi.org/10.1371/journal.pone.0115090
Borkowski J, Obidziński A (2003) The composition of the autumn and winter diets in two Polish populations of fallow deer. Acta Theriol 48:539–546. https://doi.org/10.1007/bf03192499
Castillo CP, Kavalov B, Diogo V, Jacobs-Crisioni C, e Silva FB, Lavalle C (2018) Agricultural land abandonment in the EU within 2015–2030. JRC Working Paper JRC113718, Joint Research Centre
Coppedge BR, Leslie DM, Shaw JH (1998) Botanical composition of bison diets on tallgrass prairie in Oklahoma. J Range Manag 51:379–382. https://doi.org/10.2307/4003321
Craine JM, Towne EG, Miller M, Fierer N (2015) Climatic warming and the future of bison as grazers. Sci Rep 5(1):16738. https://doi.org/10.1038/srep16738
Cromsigt JP, Kemp YJ, Rodriguez E, Kivit H (2018) Rewilding Europe’s large grazer community: how functionally diverse are the diets of European bison, cattle, and horses? Restor Ecol 26(5):891–899. https://doi.org/10.1111/rec.12661
Davis MA, Chew MK, Hobbs RJ, Lugo AE, Ewel JJ, Vermeij GJ., Brown JH, Rosenzweig ML, Gardener MR, Carroll SP, Thompson K, Pickett STA, Stromberg JC, Tredici PD, Suding KN, Ehrenfeld JG, Grime JP, Mascaro J, Briggs JC (2011) Don’t judge species on their origins. Nature, 474(7350), 153–154. https://doi.org/10.1038/474153a. Mascaro J, Briggs JC
Díaz M, Campos Palacín P, Pulido FJ (1997) The Spanish dehesas: a diversity in land-use and wildlife. In: Pain D, Pienkowski M (eds) Farming and birds in Europe. Academic, London, UK, pp 178–209
Dvorský M, Mudrák O, Doležal J, Miloslav J (2022) Reintroduction of large herbivores restored plant species richness in abandoned dry temperate grassland. Plant Ecol 223:525–535. https://doi.org/10.1007/s11258-022-01225-w
Ellis EC (2011) Anthropogenic transformation of the terrestrial biosphere. Philosophical Trans Royal Soc A 369:1010–1035. https://doi.org/10.1098/rsta.2010.0331
Forte C, Diaferia M, Manganelli E, Piergili Fioretti D, Trabalza Marinucci M, Olivieri O (2012) The European Bison living in the National Park of Poloniny, Slovakia: dietary habits and dynamics of endoparasite population. Biosférické Rezeervácie na Slovesku IX. (edit. R. Midriak) Zborník referátov z 9. národnej konferencie o biosférických rezerváciách SR. konanej, Stakčíne, pp 50–57
Garcia-Gonzalez R, Cuartas P (1992) Food habits of Capra pyrenaica, Cervus elaphus and Dama dama in the Cazorla Sierra (Spain). Mammalia 56(2):195–202. https://doi.org/10.1515/mamm-1992-0203
Gebert C, Verheyden H (2008) Variations of diet composition of red deer (Cervus elaphus L.) in Europe. Mammal Rev 31:189–201. https://doi.org/10.1111/j.1365-2907.2001.00090.x
Grange T, Brugal J-P, Flori L, Gautier M, Uzunidis A, Geigl E-M (2018) The evolution and population diversity of bison in Pleistocene and Holocene Eurasia: sex matters. Diversity 10:65. https://doi.org/10.3390/d10030065
Hanberry BB, Faison EK (2023) Re-framing deer herbivory as a natural disturbance regime with ecological and socioeconomic outcomes in the eastern United States. Sci Total Environ 868:161669. https://doi.org/10.1016/j.scitotenv.2023.161669
Hartvig I, Howe AG, Schmidt ENB, Pertoldi C, Lund Nielsen J, Buttenschoen RM (2021) Diet of the European bison (Bison bonasus) in a forest habitat estimated by DNA barcoding. Mammal Res 66:123–136. https://doi.org/10.1007/s13364-020-00541-8
Hecker LJ, Edwards MA, Nielsen SE (2021) Assessing the nutritional consequences of switching foraging behavior in wood bison. Ecol Evol 11(22):16165–16176. https://doi.org/10.1002/ece3.8298
Hofmann RR (1989) Evolutionary steps of ecophysiological adaptation and diversification of ruminants: a comparative view of their digestive system. Oecologia 78:443–457. https://doi.org/10.1007/BF00378733
Holechek JL, Vavra M, Pieper RD (1982) Botanical Composition determination of Range Herbivore diets: a review. J Range Manag 35:309–315
Jackson J (1977) The annual diet of the fallow deer (Dama dama) in the New Forest, Hampshire, as determined by rumen content analysis. J Zool 181:465–473. https://doi.org/10.1111/j.1469-7998.1977.tb03257.x
Kaczmarek-Okrój M, Olech W (2022) Reproduction parameters of wisent in ex situ condition. Eur Bison Conserv Newsl 14:29–42
Kerley GIH, Kowalczyk R, Cromsigt JPGM (2012) Conservation implications of the refugee species concept and the European bison: king of the forest or refugee in a marginal habitat? Ecography 35:519–529. https://doi.org/10.1111/j.1600-0587.2011.07146.x
Knapp AK, Blair JM, Briggs JM, Collins SL, Hartnett DC, Johnson LC et al (1999) The keystone role of bison in north American tallgrass prairie. Bioscience 49:39–50. https://doi.org/10.2307/1313492
Kowalczyk R, Taberlet P, Coissac E, Valentini A, Miquel C, Kamiński T, Wojcik JM (2011) Influence of management practices on large herbivore diet—case of European bison in Białowieża Primeval Forest (Poland). For Ecol Manage 261:821–828. https://doi.org/10.1016/j.foreco.2010.11.026
Kowalczyk R, Krasińska M, Kamiński T, Górny M, Struś P, Hofman-Kamińska E, Krasiński ZA (2013) Movements of European bison (Bison bonasus) beyond the Białowieża Forest (NE Poland): range expansion or partial migrations? Acta Theriol 58:391–401. https://doi.org/10.1007/s13364-013-0136-y
Kowalczyk R, Wójcik JM, Taberlet P, Kamiński T, Miquel C, Valentini A, Craine JP, Coissac É (2019) Foraging plasticity allows a large herbivore to persist in a sheltering forest habitat: DNA metabarcoding diet analysis of the European bison. For Ecol Manag 449:117474. https://doi.org/10.1016/j.foreco.2019.117474
Kowalczyk R, Kamiński T, Borowik T (2021) Do large herbivores maintain open habitats in temperate forests? For Ecol Manag 494:119310. https://doi.org/10.1016/j.foreco.2021.119310
Krasińska M, Krasiński Z (2013) European Bison: The Nature Monograph. https://doi.org/10.1007/978-3-642-36555-3
Krojerová-Prokešová J, Barančeková M, Šustr P, Heurich M (2010) Feeding patterns of red deer Cervus elaphus along an altitudinal gradient in the Bohemian Forest: effect of habitat and season. Wildl Biology 16(2):173–184. https://doi.org/10.2981/09-004
Landau S, Azaizeh H, Muklada H, Glasser T, Ungar ED, Baram H, Abbas N, Markovics A (2010) Anthelmintic activity of Pistacia lentiscus foliage in two middle eastern breeds of goats differing in their propensity to consume tannin-rich browse. Vet Parasitol 173(3–4):280–286. https://doi.org/10.1016/j.vetpar.2010.07.006
Lasanta T, Khorchani M, Pérez-Cabello F, Errea P, Sáenz-Blanco R, Nadal-Romero E (2018) Clearing shrubland and extensive livestock farming: active prevention to control wildfires in the Mediterranean mountains. J Environ Manage 227:256–266. https://doi.org/10.1016/j.jenvman.2018.08.104
Li Z, Wang J, Khattak RH, Han X, Liu P, Liu Z, Teng L (2022) Coexistence mechanisms of sympatric ungulates: behavioral and physiological adaptations of blue sheep (Pseudois nayaur) and red deer (Cervus elaphus alxaicus) in Helan Mountains, China. Front Ecol Evol 10:925465. https://doi.org/10.3389/fevo.2022.925465
Lisonbee LD, Villalba JJ, Provenza FD, Hall JO (2009) Tannins and self-medication: Implications for sustainable parasite control in herbivores. Behavioural Processes, 82(2), 184–189. https://doi.org/10.1016/j.beproc.2009.06.009
Maia MJ, Rego F, Sousa F (2003) Determining Optimal Sampling schemes to study red deer diets by fecal analysis. Silva Lusitana 11:91–99
Mancilla-Leytón JM, Hernando C, Cambrollé J, Muñoz-Vallés S, Pino-Mejías R, Vicente ÁM (2021) Can Shrub Flammability be affected by Goat Grazing? Flammability Parameters of Mediterranean Shrub Species under Grazing. Sustainability 13(3):1555. https://doi.org/10.3390/su13031555
Massilani D, Guimaraes S, Brugal J-P, Bennett EA, Tokarska M, Arbogast R-M, Baryshnikov G, Boeskorov G, Castel J-C, Davydov S, Madelaine S, Putelat O, Spasskaya NN, Uerpmann H-P, Grange T, Geigl E-M (2016) Past climate changes, population dynamics and the origin of Bison in Europe. BMC Biol 14:93. https://doi.org/10.1186/s12915-016-0317-7
Mendoza M, Palmqvist P (2008) Hypsodonty in ungulates: an adaptation for grass consumption or for foraging in open habitat? J Zool Vol 274. https://doi.org/10.1111/j.1469-7998.2007.00365.x
Merceron G, Hofman-Kamińska E, Kowalczyk R (2014) 3D dental microwear texture analysis of feeding habits of sympatric ruminants in the Białowieża Primeval Forest, Poland. For Ecololy Manage 328:262–269. https://doi.org/10.1016/j.foreco.2014.05.041
Miranda M, Cristóbal I, Díaz L, Sicilia M, Molina-Alcaide E, Bartolomé J, Fierro Y, Cassinello J (2015) Ecological effects of game management: does supplemental feeding affect herbivory pressure on native vegetation? Wildl Res 42(4):353–361. https://doi.org/10.1071/WR15025
Morse BW, McElroy ML, Miller KV (2009) Seasonal diets of an Introduced Population of fallow deer on Little St. Simons Island, Georgia. Southeast Nat 8(4):571–586. https://doi.org/10.1656/058.008.0401
Neov B, Spassov N, Hristova L, Hristov P, Radoslavov G (2021) New data on the evolutionary history of the European bison (Bison bonasus) based on subfossil remains from Southeastern Europe. Ecol Evol. https://doi.org/10.1002/ece3.7241
Obidziński A, Kiełtyk P, Borkowski J, Bolibok L, Remuszko K (2013) Autumn-winter diet overlap of fallow, red, and roe deer in forest ecosystems, Southern Poland. Open Life Sci 8(1). https://doi.org/10.2478/s11535-012-0108-2
Olech W, Perzanowski K (2022) European Bison (Bison bonasus) Strategic species Status Review 2020. IUCN SSC Bison Specialist Group and European Bison Conservation Center, Warsaw, pp 1–138
Onar V, Soubrier J, Toker NY, van Loenen A, Llamas B, Siddiq AB, Pasicka E, Tokarska M (2017) Did the historical range of the European bison (Bison bonasus L.) extend further south? A new finding from the Yenikapı Metro and Marmaray excavation. Turk Mammal Res 62(1):103–109. https://doi.org/10.1007/s13364-016-0299-4
Oosting HJ (1956) The study of plant communities: an introduction to plant ecology. WH Freeman and Company, San Francisco, p 440
Painter LE, Ripple WJ (2012) Effects of bison on willow and cottonwood in northern Yellowstone National Park. For Ecol Manage 264:150–158. https://doi.org/10.1016/j.foreco.2011.10.010
Palacio P, Berthonaud V, Guérin C, Lambourdière J, Maksud F, Philippe M et al (2017) Genome data on the extinct Bison schoetensacki establish it as a sister species of the extant European bison (Bison bonasus). BMC Evol Biol 17(1):1–11. https://doi.org/10.1186/s12862-017-0894-2
Patou-Mathis M (2000) Neanderthal subsistence behaviors in Europe. Int J Osteoarchaeology 10:379–395. https://doi.org/10.1002/1099-1212(200009/10)10:5%3C379::aid-oa558%3E3.0.co;2-4
Pinto-Correia T, Godinho S (2013) Changing agriculture-changing landscapes: what is going on in the high valued montado. In: Ortiz-Miranda D, Moragues-Faus A, Arnalte-Alegre E (eds) Agriculture in Mediterranean Europe: between old and new paradigms (research in rural sociology and development, vol 19. Emerald Group Publishing Limited, Bingley, pp 75–90. https://doi.org/10.1108/S1057-1922(2013)0000019006
Plumb G, Kowalczyk R, Hernandez-Blanco JA (2020) Bison bonasus. The IUCN Red List of Threatened Species 2020: e.T2814A45156279. https://doi.org/10.2305/IUCN.UK.2020-3.RLTS.T2814A45156279.en
Pucek Z (2004) European Bison. Status survey and conservation action plan. IUCN/SSC Bison specialist Group. IUCN, Gland, Switzerland and Cambridge, UK
Putman RJ (1986) Competition and coexistence in a multi-species grazing system. Acta Theriol 31:271–291. https://doi.org/10.4098/at.arch.86-26
Putman RJ, Culpin S, Thirgood SJ (1993) Dietary differences between male and female fallow deer in sympatry and in allopatry. J Zool 229(2):267–275. https://doi.org/10.1111/j.1469-7998.1993.tb02635.x
Queirós J, Acevedo P, Santos JPV, Barasona J, Beltran-Beck B, González-Barrio D, Armenteros JA, Diez-Delgado I, Boadella M, Fernandéz de Mera I, Ruiz-Fons JF, Vicente J, de la Fuente J, Gortázar C, Searle JB, Alves PC (2019) Red deer in Iberia: molecular ecological studies in a southern refugium and inferences on European postglacial colonization history. PLoS ONE 14(1):e0210282. https://doi.org/10.1371/journal.pone.0210282
Reynolds H, Hansen R, Peden D (1978) Diets of the Slave River lowland bison herd, Northwest territories, Canada. J Wildl Manag 42:581–590. https://doi.org/10.2307/3800821
Rouet-Leduc J, Pe’er G, Moreira F, Bonn A, Helmer W, Shahsavan Zadeh SAA, Zizka A, van der Plas F (2021) Effects of large herbivores on fire regimes and wildfire mitigation. J Appl Ecol. https://doi.org/10.1111/1365-2664.13972
Rivals F, Lister AM (2016) Dietary flexibility and niche partitioning of large herbivores through the Pleistocene of Britain. Q Sci Rev 146:116–133. https://doi.org/10.1016/j.quascirev.2016.06.007
Slatis (1960) An analysis of inbreeding in the European bison. Genetics 45:275–287. https://doi.org/10.1093/genetics/45.3.275
Sorbelli L, Alba DM, Cherin M, Moullé P-É, Brugal J-P, Madurell-Malapeira J (2021) A review on Bison schoetensacki and its closest relatives through the early-middle pleistocene transition: insights from the Vallparadís Section (NE Iberian Peninsula) and other European localities. Q Sci Rev 261:106933. https://doi.org/10.1016/j.quascirev.2021.106933
Soubrier J, Gower G, Chen K, Richards SM, Llamas B, Mitchell KJ, Ho SYW, Kosintsev P, Lee MSY, Baryshnikov G, Bollongino R, Bover P, Burger J, Chivall D, Crégut-Bonnoure E, Decker JE, Doronichev VB, Douka K, Fordham DA, Fontana F, Fritz C, Glimmerveen J, Golovanova LV, Groves C, Guerreschi A, Haak W, Higham T, Hofman-Kamińska E, Immel A, Julien MA, Krause J, Krotova O, Langbein F, Larson G, Rohrlach A, Scheu A, Schnabel RD, Taylor JF, Tokarska M, Tosello G, van der Plicht J, van Loenen A, Vigne JD, Wooley O, Orlando L, Kowalczyk R, Shapiro B, Cooper A (2016) Early cave art and ancient DNA record the origin of European bison. Nat Commun 7:13158. https://doi.org/10.1038/ncomms13158
Spassov N, Stoytchev T (2003) On the origin of the wisent, Bison bonasus (Linnaeus, 1758): presence of the wisent in the Upper Palaeolithic art of Eurasia. Advances in Vertebrate Paleontology Hen to Panta, papers in honour of C. Radulescu and P.M. Samson. Bucharest: 125–130
Spitzer R, Felton A, Landman M, Singh NJ, Widemo F, Cromsigt JP (2020) Fifty years of European ungulate dietary studies: a synthesis. Oikos 129(11):1668–1680. https://doi.org/10.1111/oik.07435
Stewart DRM (1967) Analysis of plant epidermis in faeces: a technique for studying the food preferences of grazing herbivores. J Appl Ecol 4:83–111. https://doi.org/10.2307/2401411
Storr GM (1961) Microscopic Analysis of Faeces, a technique for ascertaining the Diet of Herbivorous, mammals. Australian J Biol Sci 14(1):157–164
Tokeshi M (2009) Species coexistence: ecological and evolutionary. Perspectives. JohnWiley & Sons, Hoboken, NJ
Trepel J, le Roux E, Abraham AJ, Buitenwerf R, Kamp J, Kristensen JA, Tietje M, Erick J, Lundgren EJ, Svenning JC (2024) Meta-analysis shows that wild large herbivores shape ecosystem properties and promote spatial heterogeneity. Nature Ecology & Evolution. https://doi.org/10.1038/s41559-024-02327-6
Valdés-Correcher E, Rodriguez E, Kemp YJM, Wassen MJ, Cromsigt JPGM (2018) Comparing the impact of a grazing regime with European bison versus one with free-ranging cattle on coastal dune vegetation in the Netherlands. Mammal Res. https://doi.org/10.1007/s13364-018-0373-1
Van Wieren SE (1995) The potential role of large herbivores in nature conservation and extensive land use in Europe. Biol J Linn Soc 56:11–23. https://doi.org/10.1111/j.1095-8312.1995.tb01114.x
Varillas B (2013) Vuelven Los grandes herbívoros salvajes que Alimentaron Al Hombre De Atapuerca. Dendra médica. Revista De Humanidades 12:192–210
Waggoner V, Hinkes M (1986) Summer and fall browse utilization by an alaskan bison herd. J Wildl Manage 50:322–324. https://doi.org/10.2307/3801921
Wang L, Pedersen PBM, Svenning JC (2023) Rewilding abandoned farmland has greater sustainability benefits than afforestation. Npj Biodivers 2:5. https://doi.org/10.1038/s44185-022-00009-9
Zhou Q, Wei H, Tang H, Huang Z, Krzton A, Huang C (2013) Niche separation of sympatric macaques, Macaca assamensis and M. Mulatta, in limestone habitats of Nonggang, China. Int J Primatol 55:125–137. https://doi.org/10.1007/s10329-013-0385-z
Zielke L, Wrage-Mönnig N, Müller J, Neumann C (2019) Implications of spatial habitat diversity on diet selection of European Bison and Przewalski’s horses in a Rewilding Area. Diversity 11(4):63. https://doi.org/10.3390/d11040063
Acknowledgements
Field work was possible at El Encinarejo estate thanks to its owners, the Hohne and Murdoch families, who provided all the facilities and support. We thank the Polish State Forest and the European Bison Friends Society for their great contribution in providing the E. bison, and Mossy Earth for providing additional monitoring equipment for the project. We are also grateful to Francisco Ceacero for his recommendations and to Andrea Roverso for his help in field work. Finally, we would like to thank the technicians from the Animal Production Laboratory of the Autonomous University of Barcelona, Cristina Xufre, Carme Martínez and Anna Garrit, for their work in preparing and preserving field samples.
Funding
H.M. received a traineeship grant from the Erasmus + program. Laboratory costs were assumed by the Department of Animal and Food Science (Universitat Autònoma de Barcelona).
Open Access Funding provided by Universitat Autonoma de Barcelona.
Author information
Authors and Affiliations
Contributions
Conceptualization and experimental design, J.B.F., F.M., Y.K., J.C. Field work, Y.K., Laboratory work, J.B.F., H.M., A.G. Data analysis, J.C. Writing and editing, J.B.F., F.M., Y.K., J.C. All authors reviewed the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Communicated by Nigel Stork.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Filella, J.B., Morán, F., Kemp, Y.J.M. et al. Diet comparison between sympatric European bison, red deer and fallow deer in a Mediterranean landscape. Biodivers Conserv 33, 1775–1791 (2024). https://doi.org/10.1007/s10531-024-02832-x
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10531-024-02832-x