
Abstract
In some regions of the world, large carnivores, such as wolves, persist in landscapes with dense networks of paved roads. However, beyond the general impacts of roads on wildlife, we still lack information on carnivore responses to different types of roads and traffic volume levels. Using wolves in NW Spain as a case study, we show how wolves respond differently to paved road classes depending on road size, speed limit and traffic volume. All wolves evaluated (25 GPS collared wolves) crossed paved roads. Overall, during 3,915 sampling days, we recorded 29,859 wolf crossings. Wolf crossings of all paved road classes were recorded at a mean rate of 0.022 crossings/day/km (95% CI 0.016–0.027). Wolves crossed low speed and low traffic volume roads more frequently, and more often during the night, in order to lessen the chances of encountering traffic. We found mortality to be highest on roads with high speed and high traffic volume. How wolves interact with paved roads should be considered in landscape planning strategies in order to guarantee wolf long-term persistence in human-dominated landscapes. In our case, our results support an increasing focus on primary roads (class II) to identify segments of these roads where road mitigation efforts should be prioritised. Our study also highlights the importance of considering paved road classes when studying the impact of roads on wildlife.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
The magnitude and rate of change in land-use cover presents a challenge in endeavouring to fully understand the responses of wildlife to ever-encroaching human environments (Torres et al. 2016; Watson et al. 2016). While concern is rightly given on the conservation of roadless areas across the globe (Selva et al. 2011; Ibisch et al. 2016; Ascensão et al. 2018), there is increasing interest on how already existing and growing road networks may impact wildlife persistence (Bennett 2017). The expansion of roads in human-dominated landscapes can exacerbate habitat loss and fragmentation, lead to direct mortality by vehicle collisions and increase disturbance and human pressures on wildlife (e.g., Mech et al. 1988; Fahrig et al. 1995; Saunders et al. 2002; van Langevelde and Jaarsma 2004; Shephard et al. 2008; Ceia-Hasse et al. 2017; Laurance et al. 2017). By studying wildlife responses to growing road networks, conservationists are better able to delineate and prioritize effective mitigation measures to ensure the persistence of wildlife and functional ecosystems, in an increasingly human-dominated world (Forman 2000; Boutin and Hebert 2002; Dellinger et al. 2013; Wadey et al. 2018).
Evidence shows that large carnivores have an ability to persist in human-dominated landscapes, therefore indicating that human-large carnivore separation is not a necessary condition for their conservation (López-Bao et al. 2017). The impact of linear infrastructures on large carnivores, and their behavioural responses, has been the focus of attention in recent times (e.g., Whittington 2005; Llaneza et al. 2012; Basille et al. 2013; Dellinger et al. 2013; Boulanger and Stenhouse 2014; Ordiz et al. 2014; Riley et al. 2014; Ceia-Hasse et al. 2017; Find’o 2019; López-Bao et al. 2019; Zeller et al. 2021; Proctor et al. 2019). For example, roads decrease Amur tiger (Panthera tigris altaica) survivorship and reproductive success (Kerley et al. 2002), isolate lynx (Lynx lynx) habitat patches (Kramer-Schadt et al. 2004), and lead to grizzly bear (Ursus arctos) avoidance of landscapes with high road densities (Mace et al. 1996). Furthermore, studies have shown how carnivores can perceive risks of crossing roads, with black bears (Ursus americanus) having elevated heart rates when crossing high-traffic volume roads (Ditmer et al. 2018) or higher movement speeds when crossing roads than during other situations (Zeller et al. 2021), and grizzly bears (Ursus arctos horribilis) increasing their selection of roads closed to traffic (Whittington et al. 2019).
For species such as wolves (Canis lupus), accumulating evidence is leading an interesting debate on the levels of wolf resilience towards landscape transformations and human disturbance (e.g. Llaneza et al. 2012, 2016; Lesmerises et al. 2013; Ahmadi et al. 2014; Chapron et al. 2014; Bojarska et al. 2020); which may be highly context-dependent (e.g., Eurasia vs. North America; Chapron et al. 2014; Sazatornil et al. 2016; Muhly et al. 2019). With an estimated wolf density around 2.55 wolves/100 km2 (López-Bao et al. 2018), and a mean paved road density at 2.7 km/km2 (Llaneza et al. 2012), wolves in Galicia are expected to interact with roads frequently, as seen in other studies (e.g. Mladenoff et al. 1995; Whittington et al. 2004; Kaartinen et al. 2005; Lesmerises et al. 2013; Zimmerman et al. 2014; Ronnenberg et al. 2017). Forest roads and other trails can be advantageous to wolves, assisting movement through territories and consequently hunting and patrolling efficiency (Thurber et al. 1994; Musiani et al. 1998; James and Stuart-Smith 2000; Ciucci et al. 2003; Gurarie et al. 2011; Zimmerman et al. 2014; Dickie et al. 2017; Newton et al. 2017). On the other hand, roads and trails increase human accessibility (Thurber et al. 1994) and can lead to wolf mortality, either directly (i.e. vehicle collisions, Boyd and Pletscher 1999) or indirectly (i.e. hunting and poaching, Mech et al. 1988; Sazatornil et al. 2016; Suutarinen and Kojola 2018).
Several studies have suggested an ability of wolves to perceive mortality risks associated with humans, and therefore adjust, for instance, their use of space and time accordingly (Thurber et al. 1994; Merrill and Mech 2000; Habib and Khumar 2007; Ahmadi et al. 2014; Zimmerman et al. 2014). In regions where human presence is low, temporal use of roads can span both day and night (e.g., Thurber et al. 1994). However, in human-dominated landscapes, wolves become more cryptic and nocturnal, decreasing the probability of encountering humans (Vilà et al. 1995; Ciucci et al. 1997; Theuerkauf et al. 2003a; Kusak et al. 2005; Zimmerman et al. 2014). This evidence supports the idea that wolf responses to roads may be largely influenced by their use and the levels of traffic intensity, rather than the presence of the roads themselves (Hebblewhite and Merrill 2008; Kohn et al. 2009; Lesmerises et al. 2013). Since traffic intensity varies between different road types, presumably so too would wolf activity (Kohn et al. 2009).
Roads can be categorised into several classes depending on traffic volume, speed limit as well as physical attributes, such as number of lanes and fencing (van Langevelde and Jaarsma 2004; Colino-Rabanal et al. 2011). Consequently, distinguishing paved roads from one another would be a necessary step when studying wildlife responses to various road classes (Zeller et al. 2021). The Regional Government of Galicia has jurisdiction over most roads (although some of them are national jurisdiction), and they are classified into the following: (1) motorway (national and regional), (2) basic and complementary primary network (national and regional), (3) secondary network (regional) and (4) county roads network (regional). A number of studies investigating wolf, and large carnivore, movement patterns in relation to paved and forest roads grouped paved roads into one single class, namely primary roads, when analysing road network data (Theuerkauf et al. 2009; Gurarie et al. 2011; Zimmerman et al. 2014; Ordiz et al. 2015; López-Bao et al. 2019). Classification of paved road networks into more than one class has been described (e.g. primary and secondary roads; e.g., Theuerkauf et al. 2007; Ahmadi et al. 2014); but still the lack of differentiation between paved road classes can make wolf interactions to paved roads more difficult to interpret (see for example Find’o et al. 2019; Zeller et al. 2021).
Presumably, wolves persisting in close proximity to roads may be able to distinguish between different traffic intensities (Kaartinen et al. 2005; Whittington et al. 2005). With that in mind, we aimed to evaluate contrasting responses of Iberian wolves to different paved roads and traffic volume levels in a high road density region by employing data of 25 GPS collared wolves. Iberian wolves in NW Iberia have traditionally persisted in areas with high levels of human activity, and thus provide a good opportunity to investigate wolf responses to paved roads. We considered paved roads as separate road classes, according to their physical characteristics and traffic volume. Based on estimated crossing rates (which accounted for different lengths of road classes and wolf monitoring periods) made by GPS-collared wolves, we hypothesised that (1) wolves cross all paved road classes, (2) but crossing rates will be higher in low speed and low traffic volume roads compared with high speed and high traffic volume roads, (3) adults and sub-adults will cross roads more frequently than juveniles, due to adults and sub-adults moving further distances to hunt, patrol or disperse, and (4) wolves should cross all road classes more frequently during night hours, thereby reducing encounters with humans. We also hypothesised that the level of human activity, i.e. traffic volume, being different across road classes, influenced road use. We also expected mortality by vehicle collision to be highest on high-use road classes than on low-use classes, mainly because of the higher speed limit and traffic volume experienced on these road classes and therefore increasing the chances of a wolf being road-killed.
Methods
Study area
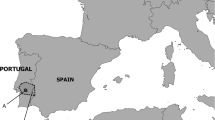
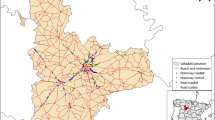
Fieldwork was carried out in Galicia (NW Spain, ca. 42.5°N 8.1°W; Fig. 1, highlighted in black box). The high human population in Galicia, mean population density of ca. 93 inhabitants/km2 (INE 2010), scattered in many settlements across the countryside, has resulted in a vast paved road network (mean paved road density 2.7 km/km2, Llaneza et al. 2012; Fig. 1b). There also exists a large network of dirt gravel roads (passable by 4 × 4 wheel drive, Fig. 1c), created to serve as access mainly for forestry and agricultural purposes. Consequently, the landscape of Galicia is heterogeneous and intensively managed by human activities comprised mainly of cropland (32%), scrubland (11%), and forestry plantations comprising of Eucalyptus spp. and Pinus spp. (43%). The last estimate of the status of wolves in Galicia was carried out between 2013 and 2015, resulting in the estimate of a continuous range distribution of wolves in the area, and 90 reproductive packs (Llaneza et al. 2015).

Galicia, north-western region of Iberian Peninsula, highlighted in black box. A Individual wolf home ranges used in this study (estimated as Minimum Convex Polygons using 100% of GPS locations; n wolves = 25. B Paved road network in Galicia, bold black lines representing motorways, national and regional roads, light roads representing secondary and county maintained roads. C Humanised landscapes of Galicia
The studied wolves
We investigated wolf response to paved roads by studying the spatial behaviour of 25 individuals equipped with GPS-GSM collars (Followit, Sweden). Between 2006 and 2013, wolves were captured with Belisle© leg-hold snares (Edouard Belisle, Saint Veronique, PQ, Canada) and chemically immobilised by intramuscular injection of medetomidine (Domitor®, Merial, Lyon, France; 0.10 mg/kg) using a blowpipe. All wolves were evaluated as clinically healthy at the moment of capture, and they only had minor lesions associated with trapping (i.e., skin abrasions). In Spain, wolves north of the Duero River are listed in Annex V of the European Habitats Directive (92/43/EEC). Fieldwork procedures were specifically approved by the Regional Government of Galicia. The wolves included in this study were captured under permits 19/2006, 71/2009, 86/2011, and 095/2013 from the Regional Government of Galicia (Spain). All fieldwork procedures adhered to the Spanish animal welfare regulations (Spanish Decree 53/2013).
Sex and age were determined in situ. Age was estimated by dental pattern and tooth wear (Gipson et al. 2000) and wolves were classified into three categories: juveniles (< 1 yr, n = 6), sub-adults (1–2 yrs, n = 10) and adults (> 2 yrs, n = 9). All wolves, except 4 pairs, belonged to different packs. We used the locations of GPS collars which were obtained every two hours for this study. Wolves were monitored an average of 156 days (range 23–390 days). For the purposes of this study, we estimated the home range of every individual as the Minimum Convex Polygon (MCP) using 100% of locations during the entire monitoring period for each wolf (Fig. 1a). Accordingly, the mean (± SD) wolf MCP of the 25 wolves studied was 262.05 ± 253.46 km2 (Appendix B).
The paved road network within wolf home ranges: Paved road classes
Firstly, we categorised paved roads into four classes depending on their purpose, speed limit and traffic volume (Table 1). We used official information on the Galician paved road network (information provided by the Regional Government of Galicia and Fomento Ministry of Government of Spain) to quantify the extent of the paved road network within every wolf home range (MCP). In addition, we also reviewed this paved road network within each wolf MCP by overlapping the official spatial information provided with high-resolution digital orthoimages, since some roads may not be included in the official data (Llaneza et al. 2018). All paved roads that were missing were added to the network manually using ArcGIS (ESRI, California, USA). Paved roads were classified in accordance to their purpose, speed limit and traffic volume, with number of lanes and fencing details given to describe wolf accessibility (Table 1). The length of each paved road class within each MCP was measured in km to calculate road class densities (km/km2).
Traffic flow
We obtained data of traffic flow from the official transport records of the Regional Government of Galicia (Autonomous Community maintained roads) and from the Spanish Ministry of Development (motorways and national roads, references in Appendix A). Traffic flow for motorways, national and regional roads, as well as a few secondary roads were obtained from those sources. No data on traffic flow was available for county roads and therefore this class was removed for this specific analysis. For the other three road types within wolf MCPs, traffic flow stations on roads crossed by wolves were utilised, and the mean traffic flow on a daily and every 2 h time interval were used to test the influence of traffic volume on wolf response to paved roads.
Wolf road-kill dataset
We compiled official records on wolf mortality caused by vehicle collisions between 1991 and 2012 from the Regional Government of Galicia (Appendix F). The sex was known for all 54 dead wolves: 25 (46.3%) female and 29 (53.7%) male. Estimation of age was possible in 48 cases: 27 (50%) adults, 12 (22.2%) sub-adults and 9 (16.7%) juveniles. No age classification was given for 6 individuals (11.1%). Monthly distribution of vehicle collision mortalities was graphed (Appendix G).
Data analysis
For each individual, consecutive GPS locations were connected via straight-line segments, and these line segments were used to determine crossing rates, by overlapping them with the paved road network. Only line segments derived from effective locations every 2 h were considered, removing the small number of cases of missing locations. GPS success rate was very high in this area. For example, for 5 wolves in our dataset and 36,104 locations (range 3654–11,861 per wolf), GPS success rate was, on average, 98% per individual (Planella et al. 2016). When a line segment from the initial GPS location to the successive location was bisected by a road an intersect point was created using ArcGIS. Since we considered a 2 h time interval, our approach allowed us to derive a minimum number of crossings.
To test for differences in the frequency of wolf crossings in relation to the road class, we analysed the number of crossings on each road class by calculating an “Index Cross” for each wolf, accounting for sampling effort and road length. Thus, we took into account that the number of crosses depended on the number of days that each wolf was sampled (there was a significant and positive correlation between the number of crosses of paved roads and the number of sampling days, Pearson correlation analysis, rp = 0.856, P < 0.0001, n = 25). We also considered the length (km) of every road class within wolf MCPs (there was a significant and positive correlation between the number of crosses and the length of each paved road class, Pearson correlation, rp = 0.474, P = 0.017, n = 25).
The Index Cross for each paved road class within a wolf MCP was calculated as follows (Eq. 1):
where the number of crosses were standardised for all wolves by dividing this figure by the ratio between the number of wolf monitoring days and road length (km). We log-transformed the Index Cross for subsequent analysis. For each wolf, we also separated every Index Cross into three time periods within the circadian cycle. We split the 24-h period into: (i) night (only star and lunar light, i.e., difficult to make out horizon), (ii) twilight (the sun is 6 degrees below the horizon) and (iii) day, the remaining time. In order to account for wolf crepuscular activity (Merrill and Mech 2003; Theuerkauf 2009), we buffered twilight 2 h before sunrise and 2 h after sunset. Daylight Savings Time was also accounted for.
We built general linear mixed models (GLMMs) with Gaussian error distribution and identity link, to compare a set of seven competing models explaining wolf response to paved roads, using as a proxy the Index Cross (response variable), according to age, road class, time period and their interactions. Firstly, we considered (i) the null model, (ii) a model containing the variable wolf age (3 levels), (iii) a model considering the road class (4 levels, Table 1), and (iv) a model considering time period (3 levels). Three interaction models were also employed, including the interaction between road class and time period, in order to explore different temporal use of paved roads according to the road class, the interaction between age and road class and the interaction between age and time period. The interactions with age were considered to test for individual differences in crossing rates according to individual attributes. As repeated measurements of the wolf responses to different paved roads are unlikely to be independent for the same individual, the identity of the individual was treated as a random factor in these models. Sex was not considered in the set of competing models since we did not detect differences in crossing rates between the sexes (GLMM using Index Cross as response variable, sex as predictor, and wolf identity as random factor, P = 0.984).
In addition, for the small number of roads within wolf MCPs where data on traffic flow was available (n roads = 54), we built another GLMM with Negative Binomial error distribution and log link to evaluate how traffic flow (mean daily number of vehicles) influenced the number of crossings. We used as a response variable the mean daily number of crossings over a given paved road made by a wolf (i.e., count data, not distributed following a Poisson distribution). The identity of the individual was also treated as a random factor in this model. For this dataset, we compared this model against a model considering road class (i.e., the road class according to Table 1), and a model considering both predictors (traffic flow and road class), in order to get insights into the idea of whether traffic flow is more important than the type of road considered.
Akaike Information Criterion with a second order correction for small sample size (AICc) was used for model selection (Burnham and Anderson 2010). We also used the Akaike weights (wi) to determine the relative strength of support for each competing model (Burnham and Anderson 2010). We used the “glmmADMB” package for R software (Skaug et al. 2014) to run GLMMs, the ‘‘car” package (Fox et al. 2015) to evaluate the significance levels for model parameters, and the “bbmle” package to calculate Akaike weights (Bolker 2017). Additionally, we also correlated the mean hourly traffic flow against the number of wolf crossings, and the number of mortality events by type of paved road with the Index Cross using a Spearman’s Rank Correlation analysis. Finally, Kruskal–Wallis test was used to test for significant differences in the wolf response (i.e., Index Cross) to traffic flow across time periods. All statistical analyses were performed in R 3.0.2 (R Core Team 2015).
Results
On average, the studied wolves persisted in areas with densities of paved roads at the home range level (MCP) ranging from 0.75 to 2.95 km/km2, with a mean paved road density of 1.92 km/km2 (Appendix B). The majority of wolf MCPs (98%) contained primary and secondary roads (Appendix B), and eight out of twenty-five wolves (32%) had motorways within their MCPs, with 62% of these wolves having crossed them. Paved road length was positively and significantly correlated with MCP home range size (Spearman’s rank correlation, rs = 0.975, n = 25, P < 0.0001).
Overall, in 3915 sampling days, we recorded 29,859 wolf crossings over paved roads (mean number of crosses by wolf = 1194, range 43–4172) (Appendix C). Wolf crosses of all paved road classes were recorded at a mean rate of 0.022 crossings/day/km (95% CI 0.016–0.027). Considering this mean rate, and the minimum length of paved roads observed within a wolf home range (39 km; Appendix B), this means that the studied wolves were expected to cross a paved road at least once every day. Road class IV (county roads) and class II roads (primary roads) were crossed more frequently, followed by road class III (secondary roads) and road class I (motorways) (Fig. 2). The median index cross over class I roads was 0.001 crossings/day/km, 0.018 in class II roads, 0.010 in class III roads and 0.019 in class IV roads (Fig. 2). Crossings were most frequent at night (median, 0.014 crossings/day/km), then during dawn/dusk (0.003 crossings/day/km) and daylight (0.001 crossings/day/km) (Fig. 2). Adult (0.018 crossings/day/km) and sub-adult (0.028 crossings/day/km) wolves crossed roads more frequently than juveniles (0.009 crossings/day/km; Fig. 2), and all wolves crossed more frequently during night hours (Appendix D). The most parsimonious model explaining the variation in wolf crossings over paved roads was the model considering road class, time period and their interaction (wi = 1; Table 2). Wolf crossings over paved roads were significantly different across road types (Wald χ2 = 110.62, d.f. = 3, P < 0.0001; Appendix E); whereas time period and the interaction road class and time period did not show significant influence on wolf Index Cross (both P > 0.234; Appendix E).

Bar charts of median index cross (crossings/day/km) by road class, solar time, and wolf age with inter-quartile range displayed as error bars
Traffic flow influenced the number of crossings registered by wolves. We detected a negative and marginal significant effect of traffic flow on the mean daily number of wolf crossings (Wald χ2 = 3.38, d.f. = 1, P = 0.066). The model containing traffic flow showed the highest support (wi = 0.50), compared to the model including road class (wi = 0.26) and the model with both predictors (wi = 0.25) (road class was not significant in both models (P > 0.382). The median response of mean traffic flow differed across time periods (Kruskal–Wallis, H = 367.50, d.f. = 11, P = 0.0001). Interestingly, when traffic flow and number of crossings were examined on a two hour basis, we observed a significant and negative correlation between mean traffic flow and the mean number of crossings (Spearman’s rank correlation, rs:− 0.385, n = 708, P = 0.0001; Fig. 3).

Mean number of crossings versus mean traffic across time (2-h interval)
Out of the 54 records of wolves being road-killed in the area between 1991 and 2012, the majority of cases occurred on primary roads (class II) (61%), followed by secondary roads (class III) (28%), county roads (class IV) (9%) and motorways (class I) (2%) (Appendix F). We did not detect a significant correlation between the mean Index Cross by road class and the number of road kills (Spearman’s rank correlation, rs = 0.634, n = 4, P = 0.365; Fig. 4). But, the mean Index Cross for primary roads (class II) (0.026 crossings/day/km), and the number of road-killed wolves was the highest (Fig. 4). By looking at the monthly distribution of vehicle collision mortalities, it was observed that most collisions occurred in the early spring and late autumn months, with peaks in February and October (Appendix G).

Bar chart of mortality numbers against the Index Cross for each road class
Discussion
Iberian wolves in Galicia live in a highly human-dominated landscape, with road densities (mean paved road density in wolf home ranges -MCPs-: 1.92 km/km2) well above road thresholds reported in most previous studies (between 0.23 km/km2 and 1.12 km/km2; e.g., Mech et al. 1988, 1989; Mladenoff et al. 1995, 1997, 1999; Theuerkauf et al. 2003a; Jȩdrzejewski et al. 2005; Kaartinen et al. 2005; Potvin et al. 2005; Mladenoff et al. 2009; Mattisson et al. 2013; Ronneberg et al. 2017). As expected, wolves crossed all four classes of paved roads in our study area; although further research is needed to explore in depth avoidance behaviours towards roads or particular landscape characteristics associated with crossing sites (see for example Baigas et al. 2017; Find’o et al. 2019; Zeller et al. 2021). Canada lynx (Lynx canadiensis) displayed no avoidance behaviour and were found to cross 2-lane highways every other day (Baigas et al. 2017). Elsewhere, wolves have been recorded crossing many paved roads, especially during dispersal events (e.g., Mech et al. 1995; Blanco and Cortés 2007). Since in this study we considered a 2 h time interval to calculate the number of wolf crossings, our approach allowed us to derive a minimum number of crossings.
Among road classes, wolves crossed more frequently low speed and low traffic volume roads, when compared with high speed and high traffic volume roads. County roads (class IV) had higher crossing rates in comparison to motorways (class I), presumably because pavement width is shorter, speed limit is slower and traffic volume is lower (see Skuban et al. 2017 for similar responses by brown bears). The higher density of county roads (class IV) within wolf MCPs (Appendix B) would also account for the higher crossing rates, as the probability of a wolf encountering this road class is greater. With the exception of two wolves, primary roads (class II) and secondary roads (class III) were present in all wolf home ranges (Appendix B). Interestingly, primary roads (class II) were the second most frequently crossed road class followed by secondary roads (class III). This road class has a wider pavement width and experiences higher speed limits and traffic volume compared with secondary roads (class III), which may lead to higher mortality rates, as discussed further (Fig. 4). The lower crossing rates shown by juveniles, may be related to the fact that wolves select breeding sites away from paved roads (Theuerkauf et al. 2003b; Iliopoulos et al. 2014; Sazatornil et al. 2016; Llaneza et al. 2018).
Of the wolves that had a section of motorway (class I) within their home range (n = 8), five crossed this road class and more than once for some individuals. However, we were unable to determine whether the five wolves that crossed a section of motorway used passages or crossed the motorway itself frequently. Crossings made by wolves on motorways have been described elsewhere (Licht and Fritts 1994; Kohn et al. 2000; Merrill and Mech 2000; Blanco et al. 2005; Ciucci et al. 2009; Lesmerises et al. 2013). Wolves were reported to cross a motorway after a period of local disturbance (i.e. roadwork completion, Lesmerises et al. 2013). In contrast, an increasing human presence on motorways has resulted in fewer crossings made by wolves in other regions (Alexander and Waters 2000; Musiani et al. 2010). As well as traffic, fencing can also affect wolf response to motorways. Fencing along a motorway can act as a barrier and lead to fewer crossings (Paquet and Callaghan 1996), while an unfenced motorway in Wisconsin had no”barrier effect” on wolf movements (Kohn et al. 2000). Remarkably, fenced motorways have not hindered the expansion of wolves in Europe (Blanco et al. 2005; Ciucci et al. 2009; Reinhardt et al. 2019). Overall, wolves crossed roads mainly during the night given that traffic volume increases during daylight hours. Similar patterns of nocturnal use of trails and paved roads were reported before (Vilà et al. 1995; Ciucci et al. 1997; Theuerkauf et al. 2003a; Blanco et al. 2005; Kusak et al. 2005; Hebblewhite and Merrill 2008; Houle et al. 2010; Gurarie et al. 2011; Zimmerman et al. 2014). In contrast, wolves living in areas with low traffic volume may not display such preferences for nocturnal activity (Thurber et al. 1994).
One of the major causes of death for wildlife in human-dominated landscapes is traffic mortality (Jaarsma et al. 2006). Over twenty-one years, the majority of wolves found road-killed were adults with mortalities occurring in the late winter/early spring (mating and breeding season) and early autumn months (dispersal). In comparison, high road mortality was observed for younger wolves in Italy but similarly to this study, adult mortality on roads occurred during the winter months (Lovari et al. 2007). We found wolf mortality to be highest on primary roads (class II), with the least number of mortalities occurring on motorways (class I). The high speed limit and traffic volume experienced on a primary road (class II) would certainly result in a higher probability of a wolf being road-killed. However, by contrast, wolf mortality was highest on motorways elsewhere in Spain, with fencing being reported as a key predictor in roadkill (Colino-Rabanal et al. 2011). Whilst traffic volume is an obvious explanation of mortality, the effects of fencing may not be immediately apparent, as they are usually thought of as an effective mitigation tool in reducing roadkill (Spanowicz et al. 2020). However, if a wolf enters a motorway through gaps in fencing, they may be forced to spend more time on the road searching for an exit, thus heightening their chances of being killed.
Road density may not greatly influence wolf distribution at a landscape level, rather it may be on a finer, home range level, that the presence of public-paved roads can have a negative influence on wolf movements in some regions (Ciucci et al. 2003; Zlatanova and Popova 2013). The variations seen in response to roads, and other human structures, can depend on many factors, including the rate of disturbance and persecution experienced (Ciucci et al. 2003; Sazatornil et al. 2016). Wolf persistence in human-dominated landscapes is supported by a number of behavioural adaptations to deal with risk and allow persistence (Theuerkauf et al. 2003a; Kusak et al. 2005; Ahmadi et al. 2014; Zimmerman et al. 2014; Llaneza et al. 2016). In Iberian Peninsula, wolves do not have a lot of options in inhabiting remote regions when available land is located at short distances from roads (Torres et al. 2016). Thus roads will continue to pose risks to wolves (Merrill and Mech 2000) but by selecting to cross roads with shorter pavement widths, slower speeds and lower traffic volume, predominately during night hours, wolves decrease the probability of being injured or killed by a vehicle.
Factors predicting wolf mortality on Galician roads should be considered in future landscape planning strategies in order to guarantee the conditions for long-term wolf persistence. An increasing focus should be allocated to identify drivers of wolf mortality on primary roads (class II) in our study area (Fig. 4), in order to identify segments of roads where mitigation efforts should be prioritised (e.g., crossing structures such as bridges, underpasses, or fencing; Grilo et al. 2010; Van der Ree et al. 2015; Spanowicz et al. 2020). Research has indicated that wildlife-vehicle collisions are not random events, but clustered (Clevenger et al. 2003; Ramp et al. 2006; Morelle et al. 2013; Keken et al. 2019). Models will be useful in determining these factors (Litvaitis and Tash 2008), and road-related predictors could include traffic volume, road alignment and road-side topography (Clevenger et al. 2003; van Langevelde and Jaarsma 2004; Barrientos and Bolonio 2009), as well as incorporating wolf movement/crossing patterns (this study). A predictive model in wolf-vehicle collision risk may be able to inform decision-makers on the exact location of where fencing, guard-rails or a crossing structure may be required (e.g., Malo et al. 2004). If fencing of particular segments along primary roads (class II) is to be implemented as a tool for reducing collision risk, funnelling animals towards suitable crossing structures should be prioritized (Huijser et al. 2016). Our study also highlights the importance of considering different paved road classes when studying the impact of roads on wildlife.
References
Ahmadi M, López-Bao JV, Kaboli M (2014) Spatial heterogeneity in human activities favours the persistence of wolves in agroecosystems. PLoS ONE 9:e108080
Alexander SM, Waters NM (2000) The effects of highway transportation corridors on wildlife: a case study of Banff National Park. Transp Res Part C 8:307–320
Ascensão F, Fahrig L, Clevenger AP, Corlett RT, Jaeger JA, Laurance WF, Pereira HM (2018) Environmental challenges for the belt and road initiative. Nat Sustain 1:206–209
Baigas PE, Squires JR, Olsen LE, Ivan JS (2017) Using environmental features to model highway crossing behaviour of Canada lynx in the Southern Rocky Mountains. Landsc Urban Plan 157:200–213
Barrientos R, Bolonio L (2009) The presence of rabbits adjacent to roads increases polecat road mortality. Biodivers Conserv 18:405–418
Basille M, Van Moorter B, Herfindal I, Martin J, Linnell JDC, Odden J, Andersen R, Gaillard JM (2013) Selecting habitat to survive: the impact of road density on survival in a large carnivore. PLoS ONE 8:e65493
Bennett VJ (2017) Effects of road density and pattern on the Conservation of species and biodiversity. Curr Landsc Ecol Rep 2:1–11
Blanco JC, Cortés Y (2007) Dispersal patterns, social structure and mortality of wolves living in agricultural habitats in Spain. J Zool 273:114–124
Blanco JC, Cortés Y, Virgós E (2005) Wolf response to two kinds of barriers in an agricultural habitat in Spain. Can J Zool 83:312–323
Bojarska K, Sulich J, Bachmann S, Okarma H, Theuerkauf J, Gula R (2020) Opportunity and peril: how wolves use a dense network of forest roads. Mamm Biol 100:203–211
Bolker, B. (2017). Package ‘bbmle’ for R.
Boulanger J, Stenhouse GB (2014) The impact of roads on the demography of grizzly bears in Alberta. PLoS ONE 9:e115535
Boutin S, Hebert D (2002) Landscape ecology and forest management: developing an effective partnership. Ecol Appl 12:390–397
Boyd DK, Pletscher DH (1999) Characteristics of dispersal in a colonizing wolf population in the Central Rocky Mountains. J Wildl Manag 63:1094–1108
Burnham KP, Anderson DR (2010) Model selection and multimodel inference. Springer, New York, p 516
Ceia-Hasse A, Borda-de-Água L, Grilo C, Pereira HM (2017) Global exposure of carnivores to roads. Glob Ecol Biogeogr 26:592–600
Chapron G, Kaczensky P, Linnell JDC, von Arx M, Huber D, Andrén H, López-Bao JV, Adamec M, Álvares F, Anders O, Balčiauskas L, Balys V, Bedő P, Bego F, Blanco JC, Breitenmoser U, Brøseth H, Bufka L, Bunikyte R, Ciucci P, Dutsov A, Engleder T, Fuxjäger C, Groff C, Holmala K, Hoxha B, Iliopoulos Y, Ionescu O, Jeremić J, Jerina K, Kluth G, Knauer F, Kojola I, Kos I, Krofel M, Kubala J, Kunovac S, Kusak J, Kutal M, Liberg O, Majić A, Männil P, Manz R, Marboutin E, Marucco F, Melovski D, Mersini K, Mertzanis Y, Mysłajek RW, Nowak S, Odden J, Ozolins J, Palomero G, Paunović M, Persson J, Potočnik H, Quenette PY, Rauer G, Reinhardt I, Rigg R, Ryser A, Salvatori V, Skrbinšek T, Stojanov A, Swenson JE, Szemethy L, Trajçe A, Tsingarska-Sedefcheva E, Váňa M, Veeroja R, Wabakken P, Wölfl M, Wölfl S, Zimmermann F, Zlatanova D, Boitani L (2014) Recovery of large carnivores in Europe’s modern human-dominated landscapes. Science 346:1517–1519
Ciucci P, Boitani L, Francisci F, Andreoli G (1997) Home range, activity and movements of a wolf pack in Central Italy. J Zool 243:803–819
Ciucci P, Masi M, Boitani L (2003) Winter habitat and travel route selection by wolves in the Northern Apennines, Italy. Ecography 26:223–235
Ciucci P, Reggioni W, Maiorano L, Boitani L (2009) Long-distance dispersal of a rescued wolf from the Northern Apennines to the Western Alps. J Wildl Manag 73:1300–1306
Clevenger AP, Chruszcz B, Gunson KE (2003) Spatial patterns and factors influencing small vertebrate fauna road-kill aggregations. Biol Conserv 109:15–26
Colino-Rabanal VJ, Lizana M, Peris SJ (2011) Factors influencing wolf Canis lupus roadkills in Northwest Spain. Eur J Wildl Res 57:399–409
Dellinger JA, Proctor C, Steury TD, Kelly MJ, Vaughan MR (2013) Habitat selection of a large carnivore, the red wolf, in a human-altered landscape. Biol Conserv 157:324–330
Dickie M, Serrouya R, McNay RS, Boutin S (2017) Faster and farther: wolf movement on linear features and implications for hunting behaviour. J Appl Ecol 54:253–263
Ditmer M, Rettler S, Fieberg J, Iaizzo P, Laske T, Noyce K, Garshelis D (2018) American black bears perceive the risks of crossing roads. Behav Ecol 29:667–675
Fahrig L, Pedlar JH, Pope SE, Taylor PD, Wegner JF (1995) Effect of road traffic on amphibian density. Biol Conserv 73:177–182
Find’o S, Skuban M, Kajba M, Chalmers J, Kalaš M (2019) Identifying attributes associated with brown bear (Ursus arctos) road-crossing and roadkill sites. Can J Zool 97:156–164
Forman RTT (2000) Estimate of the area affected ecologically by the road system in the United States. Conserv Biol 14:31–35
Fox J, Weisberg S, Adler D, Bates D, Baud-Bovy G, Ellison S, Heiberger R (2015) Package ‘car.’ R Foundation for Statistical Computing, Vienna
Gipson PS, Ballard WB, Nowak RM, Mech LD (2000) Accuracy and precision of estimating age of grey wolves by tooth wear. J Wildl Manag 64:752–758
Grilo C, Bissonette JA, Cramer PC (2010) Mitigation measures to reduce impacts on biodiversity. In: Jones SR (ed) Highways: construction, management, and maintenance. Nova Science Publisher, New York, pp 73–114
Gurarie E, Suutarinen J, Kojola I, Ovaskainen O (2011) Summer movements, predation and habitat use of wolves in human modified boreal forests. Oecologia 165:891–903
Habib B, Kumar S (2007) Den shifting by wolves in semi-wild landscapes in the Deccan Plateau, Maharashtra, India. J Zool 272:259–265
Hebblewhite M, Merrill E (2008) Modelling wildlife-human relationships for social species with mixed-effects resource selection models. J Appl Ecol 45:834–844
Houle M, Fortin D, Dussault C, Courtois R, Ouellet JP (2010) Cumulative effects of forestry on habitat use by grey wolf (Canis lupus) in the boreal forest. Landsc Ecol 25:419–433
Huijser MP, Fairbanks ER, Camel-Means W, Graham J, Watson V, Basting P, Becker D (2016) Effectiveness of short sections of wildlife fencing and crossing structures along highways in reducing wildlife-vehicle collisions and providing safe crossing opportunities for large mammals. Biol Conserv 197:61–68
Ibisch PL, Hoffmann MT, Kreft S, Pe’er G, Kati V, Biber-Freudenberger L, DellaSala DA, Vale MM, Hobson PR, Selva N (2016) A global map of roadless areas and their conservation status. Science 354:1423–1427
Iliopoulos Y, Youlatos D, Sgardelis S (2014) Wolf pack rendezvous site selection in Greece is mainly affected by anthropogenic landscape features. Eur J Wildl Res 60:23–34
INE (2010) Censo de población y vivienda. Instituto Nacional de Estadística de España, Madrid
Jaarsma CF, van Langevelde F, Botma H (2006) Flattened fauna and mitigation: traffic victims related to road, traffic, vehicle, and species characteristics. Transp Res Part D 11:264–276
James ARC, Stuart-Smith AK (2000) Distribution of caribou and wolves in relation to linear corridors. J Wildl Manag 64:154–159
Jȩdrzejewski W, Niedzialkowska M, Mysiajek RW, Nowak S, Jedrzejewska B (2005) Habitat selection by wolves Canis lupus in the uplands and mountains of southern Poland. Acta Theriol 50:417–428
Kaartinen S, Kojola I, Colpaert A (2005) Finnish wolves avoid roads and settlements. Acta Zool Fennica 42:523–532
Keken Z, Sedoník J, Kušta T, Andrášik R, Bíl M (2019) Roadside vegetation influences clustering of ungulate vehicle collisions. Transp Res D 73:381–390
Kerley LL, Goodrich JM, Miquelle DG, Smirnov EN, Quigley HB, Hornocker MG (2002) Effects of roads and human disturbance on Amur tigers. Conserv Biol 16:97–108
Kohn BE, Frair JL, Unger DE, Gehring TM, Shelley DP, Anderson EM, Keenlance PW (2000) Impacts of the US highway expansion project on wolves in north-western Wisconsin–final report. Wisconsin Department of Natural Resourses, Winconsin
Kohn BE, Anderson EM, Thiel RP (2009) Wolves, roads, and highway development. Chapter 14. In: Wydeven AP, Van Deelen TR, Heske EJ (eds) Recovery of Grey Wolves in the Great Lakes Region of the United States: an endangered species success story. Springer, New York
Kramer-Schadt S, Revilla E, Wiegand T, Breitenmoser U (2004) Fragmented landscapes, road mortality and patch connectivity: modelling influences on the dispersal of Eurasian lynx. J Appl Ecology 41:711–723
Kusak J, Skrbinšck AM, Huber D (2005) Home ranges, movements, and activity of wolves (Canis lupus) in the Dalmatian part of Dinarids, Croatia. Eur J Wildl Res 51:254–262
Laurance WF, Campbell MJ, Alamgir M, Mahmoud MI (2017) Road expansion and the fate of Africa’s tropical forests. Front Ecol Evol 5:75
Lesmerises F, Dussault C, St. Laurent MH (2013) Major road work impacts the space use behaviour of grey wolf. Landsc Urban Plan 112:18–25
Licht DS, Fritts SH (1994) Grey wolf (Canis lupus) occurrences in the Dakotas. Am Midl Nat 132:74–81
Litvaitis JA, Tash JP (2008) An approach toward understanding wildlife-vehicle collisions. Environ Manag 42:688–697
Llaneza L, López-Bao JV, Sazatornil V (2012) Insights into wolf presence in human-dominated landscapes: the relative role of food availability, humans and landscape attributes. Divers Distrib 18:459–469
Llaneza, L., López-Bao, J.V., Palacios, V., García, E.J. (2015). Wolf monitoring in Galicia, 2013–2014. Report to Spanish Ministry of Agriculture, Food, and Environment.
Llaneza L, García EJ, Palacios V, Sazatornil V, López-Bao JV (2016) Resting in risky environments: the importance of cover for wolves to cope with exposure risk in human-dominated landscapes. Biodivers Conserv 25:1515–1528
Llaneza L, Sazatornil V, López-Bao JV (2018) The importance of fine-scale breeding site selection patterns under a landscape-sharing approach for wolf conservation. Biodivers Conserv 27:1239–1256
López-Bao JV, Bruskotter J, Chapron G (2017) Finding space for large carnivores. Nat Ecol Evol 1:140
López-Bao JV, Godinho R, Pacheco C, Lema FJ, García E, Llaneza L, Palacios V, Jiménez J (2018) Toward reliable population estimates of wolves by combining spatial capture-recapture models and non-invasive DNA monitoring. Sci Rep 8:2177
López-Bao JV, Aronsson M, Linnell JD, Odden J, Persson J, Andrén H (2019) Eurasian lynx fitness shows little variation across Scandinavian human-dominated landscapes. Sci Rep 9:1–10
Lovari S, Sforzi A, Scala C, Fico R (2007) Mortality parameters of the wolf in Italy: does the wolf keep himself from the door? J Zool 272:117–124
Mace RD, Waller JS, Manley TL, Lyon LJ, Zuuring H (1996) Relationship among grizzly bears, roads and habitat in the Swan Mountains Montana. J Appl Ecol 33:1395–1404
Malo JE, Suárez F, Díez A (2004) Can we mitigate animal-vehicle accidents using predictive models? J Appl Ecol 41:701–710
Mattisson J, Sand H, Wabakken P, Gervasi V, Liberg O, Linnell JDC, Rauset GR, Pedersen HC (2013) Home range size variation in a recovering wolf population: evaluating the effect of environmental, demographic, and social factors. Oecologia 173:813–825
Mech LD (1989) Wolf population survival in an area of high road density. Am Midl Nat 121:387–389
Mech LD, Fritts SH, Radde GL, Paul WJ (1988) Wolf distribution and road density in Minnesota. Wildl Soc Bull 16:85–87
Mech LD, Fritts SH, Wagner D (1995) Minnesota wolf dispersal to Wisconsin and Michigan. Am Midl Nat 133:368–370
Merrill S, Mech LD (2000) Details of extensive movement by Minnesota wolves (Canis lupus). Am Midl Nat 144:428–433
Merrill S, Mech LD (2003) The usefulness of GPS telemetry to study wolf circadian and social activity. Wildl Soc Bull 31:947–960
Mladenoff DJ, Sickley TA, Haight RG, Wydeven AP (1995) A regional landscape analysis and prediction of favourable grey wolf habitat in the Northern Great Lakes Region. Conserv Biol 9:279–294
Mladenoff DJ, Haight RG, Sickley TA, Wydeven AP (1997) Causes and implications of species restoration in altered ecosystems: a spatial landscape projection of wolf population recovery. Bioscience 47:21–31
Mladenoff DJ, Sickley TA, Wydeven AP (1999) Predicting grey wolf landscape recolonisation: logistic regression models vs. new field data. Ecol Appl 9:37–44
Mladenoff DJ, Clayton MK, Pratt SD, Sickley TA, Wydeven AP (2009) Change in occupied wolf habitat in the northern Great Lakes Region. Chapter 8. In: Wydeven AP, Van Deelen TR, Heske EJ (eds) Recovery of Grey Wolves in the Great Lakes Region of the United States: an endangered species success story. Springer, New York
Morelle K, Lehaire F, Lejeune P (2013) Spatio-temporal patterns of wildlife-vehicle collisions in a region with a high-density road network. Nat Conserv 5:53–73
Muhly TB, Johnson CA, Hebblewhite M, Neilson EW, Fortin D, Fryxell JM, Latham ADM, Latham MC, McLoughlin PD, Merrill E, Paquet PC, Patterson BR, Schmiegelow F, Scurrah F, Musiani M (2019) Functional response of wolves to human development across boreal North America. Ecol Evol 9:10801–10815
Musiani M, Okarma H, Jȩdrzejewski W (1998) Speed and actual distances travelled by radiocollared wolves in Białowieża Primeval Forest (Poland). Acta Theriol 43:409–416
Musiani M, Anwar SM, McDermid GJ, Hebblewhite M, Marceau DJ (2010) How humans shape wolf behaviour in Banff and Kootenay National Parks, Canada. Ecol Model 221:2374–2387
Newton EJ, Patterson BR, Anderson ML, Rodgers AR, Vander Vennen LM, Fryxell JM (2017) Compensatory selection for roads over natural linear features by wolves in northern Ontario: implications for caribou conservation. PLoS ONE 12:e0186525
Ordiz A, Kindberg J, Sæbø S, Swenson JE, Støen OG (2014) Brown bear circadian behaviour reveals human environmental encroachment. Biol Conserv 173:1–9
Ordiz A, Milleret C, Kindberg J, Mansson J, Wabakken P, Swenson JE, Sand H (2015) Wolves, people, and brown bears influence the expansion of the recolonising wolf population in Scandinavia. Ecosphere 6:1–14
Paquet, P.C., Callaghan, C. (1996) Effects of linear developments on winter movements of gray wolves in the bow river valley of Banff National Park, Alberta. In: Evink, G.L., Garrett, P., Zeigler, D., Berry, J. (Eds.) Trends in addressing transportation related wildlife mortality, proceedings of the transportation related wildlife mortality seminar. FL, USA.
Planella A, Palacios V, García EJ, Llaneza L, García-Domínguez F, Muñoz-Igualada J, López-Bao JV (2016) Influence of different GPS schedules on the detection rate of wolf feeding sites in human-dominated landscapes. Eur J Wildl Res 62:471–478
Potvin MJ, Drummer TD, Vucetich JA, Beyer DE, Peterson RO, Hammill JH (2005) Monitoring and habitat analysis for wolves in upper Michigan. J Wildl Manag 69:1–10
Proctor MF, McLellan BN, Stenhouse GB, Mowat G, Lamb CT, Boyce MS (2019) Effects of roads and motorized human access on grizzly bear populations in British Columbia and Alberta, Canada. Ursus 30e2:16–39
R Core Team (2015) R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna
Ramp D, Wilson VK, Croft DB (2006) Assessing the impacts of roads in peri-urban reserves: road-based fatalities and road usage by wildlife in the Royal National Park, New South Wales, Australia. Biol Conserv 129:348–359
Reinhardt I, Kluth G, Nowak C, Szentiks CA, Krone O, Ansorge H, Mueller T (2019) Military training areas facilitate the recolonization of wolves in Germany. Conserv Lett 12:e12635
Riley SP, Serieys LE, Pollinger JP, Sikich JA, Dalbeck L, Wayne RK, Ernest HB (2014) Individual behaviours dominate the dynamics of an urban mountain lion population isolated by roads. Curr Biol 24:1989–1994
Ronnenberg K, Habbe B, Gräber R, Strauß E, Siebert U (2017) Coexistence of wolves and humans in a densely populated region (Lower Saxony, Germany). Basic Appl Ecol 25:1–14
Saunders SC, Mislivets MR, Chen J, Cleland DT (2002) Effects of roads on landscape structure within nested ecological units of the Northern Great Lakes Region, USA. Biol Conserv 103:209–225
Sazatornil V, Rodríguez A, Klaczek M, Ahmadi M, Álvares F, Arthur S, Blanco JC, Borg BL, Cluff D, Cortés Y, García EJ, Geffen E, Habib B, Iliopoulos Y, Kaboli M, Krofel M, Llaneza L, Marucco F, Oakleaf JK, Person DK, Potočnik H, Ražen N, Rio-Maior H, Sand H, Unger D, Wabakken P, López-Bao JV (2016) The role of human-related risk in breeding site selection by wolves. Biol Conserv 201:103–110
Selva N, Kreft S, Kati V, Schluck M, Jonsson BG, Mihok B, Okarma H, Ibisch PL (2011) Roadless and low-traffic areas as conservation targets in Europe. Environ Manag 48:865
Shepard DB, Kuhns AR, Dreslik MJ, Phillips CA (2008) Roads as barriers to animal movement in fragmented landscapes. Animl Conserv 11:288–296
Skaug, H., Fournier, D., Nielsen, A., Magnusson, A., Bolker, B. (2014) glmmADMB: generalized linear mixed models using AD Model Builder. R package version 0.8.0.
Skuban M, Finďo S, Kajba M, Koreň M, Chalmers J, Antal V (2017) Effects of roads on brown bear movements and mortality in Slovakia. Eur J Wildl Res 63:82
Spanowicz A, Teixeira F, Jaeger J (2020) An adaptive plan for prioritizing road sections for fencing to reduce animal mortality. Conserv Biol. https://doi.org/10.1111/cobi.13502
Suutarinen J, Kojola I (2018) One way or another: predictors of wolf poaching in a legally harvested wolf population. Anim Conserv 21:414–422
Theuerkauf J (2009) What drives wolves: fear or hunger? Humans, diet, climate and wolf activity patterns. Ethology 115:649–657
Theuerkauf J, Jȩdrzejewski W, Schmidt K, Gula R (2003a) Spatiotemporal segregation of wolves from humans in the Białowieża forest (Poland). J Wildl Manag 67:706–716
Theuerkauf J, Rouys S, Jedrzejewski W (2003b) Selection of den, rendezvous, and resting sites by wolves in the Białowieża forest, Poland. Can J Zool 81:163–167
Theuerkauf J, Gula R, Pirga B, Tsunoda H, Eggermann J, Brzezowska B, Rouys S, Radler S (2007) Human impact on wolf activity in the Bieszczady Mountains, SE Poland. Acta Zool Fennica 44:225–231
Thurber JM, Peterson RO, Drummer TD, Thomasma SA (1994) Grey wolf response to refuge boundaries and roads in Alaska. Wildl Soc Bull 22:61–68
Torres A, Jaeger AG, Alonso JC (2016) Assessing large-scale wildlife responses to human infrastructure development. PNAS 113:8472–8477
van der Ree R, Smith DJ, Grilo C (2015) The ecological effects of linear infrastructure and traffic: challenges and opportunities of rapid global growth. Chapter 1. In: van der Ree R, Smith DJ, Grilo C (eds) Handbook of road ecology. Wiley Blackwell, Hoboken
van Langevelde F, Jaarsma CF (2004) Using traffic flow theory to model traffic mortality in mammals. Landsc Ecol 19:895–907
Vilà C, Urios V, Castroviejo J (1995) Observations on the daily activity patterns in the Iberian wolf. In: Carbyn LN, Fritts SH, Seip DR (eds) Ecology and conservation of wolves in a changing world. Canadian Circumpolar Institute, Occasional Publication, Edmonton, pp 335–340
Wadey J, Beyer HL, Saaban S, Othman N, Leimgruber P, Campos-Arceiz A (2018) Why did the elephant cross the road? The complex response of wild elephants to a major road in Peninsular Malaysia. Biol Conserv 218:91–98
Watson JE, Shanahan DF, Di Marco M, Allan J, Laurance WF, Sanderson EW, Mackey B, Venter O (2016) Catastrophic declines in wilderness areas undermine global environment targets. Curr Biol 26:2929–2934
Whittington J, St. Clair CC, Mercer G (2004) Path tortuosity and the permeability of roads and trails to wolf movement. Ecol Soc 9:1
Whittington J, St. Clair CC, Mercer G (2005) Spatial responses of wolves to roads and trails in mountain valleys. Ecol Appl 15:543–553
Whittington J, Low P, Hunt B (2019) Temporal road closures improve habitat quality for wildlife. Sci Rep 9:3772
Zeller KA, Wattles DW, Conlee L, Destefano S (2021) Response of female black bears to a high-density road network and identification of long-term road mitigation sites. Anim Conserv. https://doi.org/10.1111/acv.12621
Zimmermann B, Nelson L, Wabakken P, Sand H, Liberg O (2014) Behavioral responses of wolves to roads: scale-dependent ambivalence. Behav Ecol 25:1353–1364
Zlatanova D, Popova E (2013) Habitat variables associated with wolf (Canis lupus, L.) distribution and abundance in Bulgaria. Bulg J Agric Sci 19:262–266
Acknowledgements
JVLB was supported by a Ramon & Cajal research contract (RYC-2015-18932) from the Spanish Ministry of Economy, Industry and Competitivenes. We are in debt to the staff of the Regional Government of Galicia. We thank Victor Sazatornil, Emilio J. García and Vicente Palacios for wolf collaring and field assistance. Spatial information from some wolves in the study area comes from research projects funded by DESA S.L. and GAMESA S.L. This is scientific paper no. from the Iberian Wolf Research Team (IWRT).
Funding
Open Access funding provided thanks to the CRUE-CSIC agreement with Springer Nature.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Additional information
Communicated by Adeline Loyau.
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Dennehy, E., Llaneza, L. & López-Bao, J.V. Contrasting wolf responses to different paved roads and traffic volume levels. Biodivers Conserv 30, 3133–3150 (2021). https://doi.org/10.1007/s10531-021-02239-y
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10531-021-02239-y