
Abstract
The aim of this study was to determine the level of infection of Ixodes ricinus ticks with pathogens (Borrelia spp., Rickettsia spp., and Anaplasma spp.) collected from Lacerta agilis and Zootoca vivipara lizards in the urban areas of Wrocław (SW Poland). The study was carried out in July-August 2020. Lizards were caught by a noose attached to a pole or by bare hands, identified by species, and examined for the presence of ticks. Each lizard was then released at the site of capture. Ticks were removed with tweezers, identified by species using keys, and molecular tests were performed for the presence of pathogens. From 28 lizards (17 specimens of Z. vivipara and 11 specimens of L. agilis) a total of 445 ticks, including 321 larvae and 124 nymphs, identified as I. ricinus were collected. A larger number of ticks were obtained from L. agilis compared to Z. vivipara. Molecular tests for the presence of pathogens were performed on 445 specimens of I. ricinus. The nested PCR method for the fla gene allowed the detection of Borrelia spp. in 9.4% of ticks, and it was higher in ticks from L. agilis (12.0%) than from Z. vivipara (1.0%). The RFLP method showed the presence of three species, including two belonging to the B. burgdorferi s.l. complex (B. lusitaniae and B. afzelii), and B. miyamotoi. The overall level of infection of Rickettsia spp. was 19.3%, including 27.2% in ticks collected from Z. vivipara and 17.0% from L. agilis. Sequencing of randomly selected samples confirmed the presence of R. helvetica. DNA of Anaplasma spp. was detected only in one pool of larvae collected from L. agilis, and sample sequencing confirmed the presence of (A) phagocytophilum. The research results indicate the important role of lizards as hosts of ticks and their role in maintaining pathogens in the environment including urban agglomeration as evidenced by the first recorded presence of (B) miyamotoi and (A) phagocytophilum in I. ricinus ticks collected from L. agilis. However, confirmation of the role of sand lizards in maintaining (B) miyamotoi and A. phagocytophilum requires more studies and sampling of lizard tissue.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Infections with tick-borne pathogens (TBP) are noted not only in wild areas but also with increasing frequency in urban in suburban areas across the world (Uspensky 2014; Król et al. 2016). The risk assessment of tick-borne diseases requires comprehensive studies involving ticks, hosts, and reservoir animals that are responsible for the maintenance of zoonotic pathogens. In Central Europe, in wild, suburban, and urban areas, Ixodes ricinus is the main vector of TBP, however, also other tick species like Dermacentor reticulatus and I. hexagonus can support the circulation of pathogens in the environment (Uspensky 2014). In urban areas, as hosts of adult ticks can serve both wild animals like deer, lagomorphs, foxes, and raccoons, which easily penetrate into green spaces inside cities as well as domesticated animals including dogs, and cats which are often infested by female ticks (Uspensky 2014; Król et al. 2015, 2016; Kocoń et al. 2023). The important role as hosts of immature stages play rodents and birds, especially ground-feeding songbirds, which also can be reservoir hosts for several tick-borne pathogens (Mihalca and Sándor 2013; Rizzoli et al. 2014; Eisen 2023). As hosts of immature ticks can serve also lizards, which often inhabit urban areas. The important role of lizards as I. ricinus hosts was documented for example in Italy, where the comparable prevalence of infestation by I. ricinus larvae on lizards Podarcis muralis and mice Apodemus spp. was documented, and even more, the infestation level by nymphs were significantly greater on lizards (Amore et al. 2007). However, the role of lizards in maintaining tick population and tick-borne pathogens (TBP) has been still underestimated compared to that of mammals and birds (Ekner et al. 2011; Rizzoli et al. 2014).
Among bacterial TBP most significant and well-researched are Borrelia spp., Rickettsia spp., and Anaplasma spp. (Rochlin and Toledo 2020; Hauck et al. 2020a). Borrelia spirochetes transmitted by hard ticks are responsible for Lyme borreliosis (LB) being the most frequently reported vector-borne disease in the Northern Hemisphere and for tick-borne relapsing fever (TBRF). LB spirochaetes grouped into Borrelia burgdorferi sensu lato (s.l.) complex comprises over 20 named and proposed genospecies (Wolcott et al. 2021). Several of them are known to cause borreliosis in humans and animals (Sykes and Makiello 2017; Wolcott et al. 2021). In the relapsing fever group, an increasingly noted species is Borrelia miyamotoi, detected first in 1995 in ticks from Japan (Fraenkel et al. 2002). Disease caused by B. miyamotoi is similar to RF but can also bear similarities to borreliosis (like skin lesions) (Platonov et al. 2011). B. miyamotoi is also transmitted by Ixodes ticks but unlike.
B. burgdorferi s.l. it can be transmitted not only in a transstadial way but also via transovarial transmission (Rollend et al. 2013). Rickettsia species are grouped into four main groups: spotted fever group (SFG), typhus group (TG), transitional group (TRG), and ancestral group (AG) (Kim 2022). The SFG of Rickettsia frequently infects ticks and causes tick-transmitted rickettsioses (Portillo et al. 2015). In Central Europe, SFG of Rickettsia which are potentially able to affect humans include among other Rickettsia, which are transmitted by I. ricinus like R. helvetica, and R. monacensis (Karbowiak et al. 2016a). Anaplasma spp., similar to Rickettsia, are intracellular obligate pathogens that can be transmitted by vector (Bakken et al. 2008). From a medical point of view, the most important is Anaplasma phagocytophilum, an agent of human granulocytic anaplasmosis (HGA) (Dumler et al. 2005). The infection with A. phagocytophilum is also detected in several mammalian species, including domesticated animals like ruminants, horses, dogs, and cats (Karbowiak et al. 2016b). The current study was undertaken to evaluate the prevalence of Borrelia spp., A. phagocytophilum, and Rickettsia spp. in ticks collected from lizards in urban areas of Wrocław, SW Poland.
Materials and methods
Ticks collection
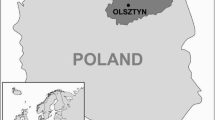
The study was carried out in July-August 2020 in a mid-forest meadow in the urban forest (Rędziński Forest) in the northern part of Wrocław, SW Poland (Fig. 1). The Rędziński Forest is located alongside two rivers: Oder and Widawa, and it is second in size forest within the city borders, covering an area of 400 ha. Part of the area belongs to a protected area designated as a Natura 2000 site under the name of Widawa Valley (Filipiak and Zaręba 2014). This study area is an important part of city green areas popular for recreation (Filipiak and Zaręba 2014; Turzanska and Turowicz 2014). The Rędziński Forest is inhabited by valuable species of wild animals, and it is also frequently visited by dogs walked by their owners. Two species of co-existing lizards: the sand lizard Lacerta agilis and the common or viviparous lizard Zootoca vivipara were caught by a noose pole or by bare hands (Bennett et al. 2001). Captured lizards were examined for the presence of ticks, which were removed with forceps and stored in 70% ethanol. After tick collection lizards were released back into the habitat. Collected ticks were identified to the species level by the use of the key to Ixodida identification (Estrada-Peña et al. 2017). The study was carried out under a permit from the Polish Authorities No. WPN.6401.212.2019.MH.1.

Lizards catching place located in Rędziński Forest, Wrocław (SW Poland)
Molecular methods
All collected nymphs of I. ricinus were tested separately. In the case of I. ricinus larvae, a total of 107 pools containing three specimens, were prepared, including 23 pools of larvae collected from Z. vivipara, and 84 pools of larvae obtained from L. agilis. DNA extraction from ticks was done with the ammonium method by Guy and Stanek (1991). Homogenized ticks (nymphs separately, and larvae pooled by 3 specimens) were placed in 100 µl of 0,65 M of ammonium hydroxide (NH4OH) and heated for 20 min in 100℃ in closed tubes, followed by 10 min in 100 °C in open tubes. After the extraction tubes were centrifuged at 11 000 rpm for 3 min, the supernatant was stored at -20℃ until molecular methods. For detection of Borrelia spp. Anaplasma spp, and Rickettsia spp. the specific nested PCR were used (Table 1). Borrelia spp. detection targeted flaB gene encoding flagellin (Wodecka et al. 2009), Anaplasma spp. detection targeted 16 S rRNA (Massung et al. 1998), and Rickettsia spp. detection targeted gltA gene, encoding citrate synthase (Prakash et al. 2012; Mitková et al. 2015). All nested PCR were carried out with the use of PCRMix Plus (A&A Biotechnology) and Thermal Cycler T100 (BioRad T100™ Thermal Cycler), reactions were followed by electrophoresis in 1,5% agarose gel (30 min, 100 V), and results were observed under UV light. Randomly selected nested PCR positive samples, including 3 samples of Rickettsia, 4 samples of Borrelia and the only Anaplasma detected, were prepared with the use of EPPiC kit (A&A Biotechnology) and sent for sequencing to MACROGEN (Netherlands). Obtained sequences were compared to existing ones in GenBank with the use of BLAST.
For differentiation genospecies of Borrelia spp. nested PCR positive samples were analysed by RFLP with the use of restriction enzyme HpyF3I (FastDigest HpyF3I, ThermoScientific), which recognises CTNAG sequence (Wodecka 2011). Restriction patterns obtained after electrophoresis in 3% agarose gel (75 min, 65 V) were observed under UV light and assigned to appropriate genospecies.
Statistical analysis
To assess the significance of obtained results χ2 test with p < 0,05 and 95% confidence intervals (95% CI) was used. To assess the discrepancy in the number of ticks parasitising certain lizard species, the standard deviation (σ) was calculated, based on the average number of parasitising ticks. To assess the level of infection in larvae ticks, Minimal Infection Rate (MIR) was calculated, assuming that one positive larvae gives a positive pool, and in the positive pool, there was only one positive larvae.
Results
From 28 lizards (17 specimens of Zootoca. vivipara and 11 specimens of Lacerta agilis) a total of 445 ticks, including 321 larvae and 124 nymphs(Table 2). All ticks collected from these lizards were identified as Ixodes ricinus. A larger number of ticks were obtained from L. agilis (342 ticks, including 252 larvae, and 90 nymphs) compared to Z. vivipara (103 ticks, including 69 larvae and 34 nymphs). The average number of ticks infested L. agilis was 31.1, and Z. vivipara 6.1 with the highest number of 90 ticks on a single L. agilis and 19 ticks on Z. vivipara.
Total infection rate of Borrelia spp. for all 445 I. ricinus ticks, calculated for larvae and nymphs together (assuming that one positive larvae gives a positive pool, and in the positive pool there was only one positive larvae) was 9.4% (42/445, 95% CI: 6.7–12.2) (Table 3), and it was higher in ticks from L. agilis (12.0%, 41/342, 95% CI: 8.6–15.4) than from Z. vivipara (1.0%, 1/103, 95% CI: 0-2.9). MIR of Ixodes ricinus larvae collected from both species of lizards was 4.0%, however, spirochaetes were only present in larvae from L. agilis (MIR = 5.2%). The total infection rate of nymphs (collected from both species of lizards) with Borrelia spp. was 23.4% (29/124, 95% CI: 15.9–30.8). There was a statistically higher infection rate in nymphs from L. agilis (31.1%, 28/90, 95% CI: 21.6–40.7) than in nymphs from Z. vivipara (2.9%, 1/34, 95% CI: 0-8.6) (χ2 = 10,93, df = 1, p < 0,001). RFLP analysing of 42 positive products of nested PCR allows the identification of 3 restriction patterns, including two from B. burgdorferi s.l. complex (B. lusitaniae and B. afzelii), and B. miyamotoi (Table 4). Sequencing four randomly selected samples identified using RFLP as B. lusitaniae confirmed their species identification. The dominant species of Borrelia was B. lusitaniae identified in 88.0% samples, including 69.0% mono-infections and 19.0% coinfection with B. miyamotoi. B. lusitaniae was the only Borrelia species detected in nymphs (38.0%) of both lizard species. B. afzelii was detected only in one nymph (2.4%) collected from L. agilis. B. miyamotoi was detected only in nymphs collected from L. agilis both as single specimens of Borrelia (9.5%) and coinfection with B. lusitaniae (19.0%).
The total infection rate of Rickettsia spp. was 19.3% (86/445, 95% CI: 15.7–23.0), including 27.2% (28/103, 95% CI: 18.6–35.8) in ticks collected from Z. vivipara and 17.0% (58/342, 95% CI: 13.0-20.9) from L. agilis. MIR for I. ricinus larvae collected from both lizard species was 10,5% and was higher for Z. vivipara (23,2%) than for L. agilis (7,1%) (χ2 = 13.557, df = 1, p = 0.0002). The prevalence for nymphs was 42.0% (52/124, 95% CI: 33.3–50.6), including 35.3% (12/34, 95% CI: 19.2–51.4) for nymphs collected from Z. vivipara and 44.4% (40/90, 95% CI: 34.2–54.7) from L. agilis, however the difference in the prevalence of Rickettsia spp. detected in nymphs was insignificant (χ2 = 0.849, df = 1, p = 0.849). Sequencing of three randomly selected samples confirmed the presence of R. helvetica.
DNA of Anaplasma spp. was detected only in one pool of larvae collected from L. agilis. Sample sequencing confirmed the presence of A. phagocytophilum.
Borrelia and Rickettsia coinfection was found in 13 cases (2 pools of larvae and 11 nymphs) of ticks collected only from L. agilis. Among Borrelia identified in coinfection were B. lusitaniae (2 pools of larvae and 9 nymphs), B. lusitaniae and B. miyamotoi (one nymph), and B. afzelii (one nymph),
Discussion
In Poland, Lacerta agilis and Zootoca vivipara are among the most numerous and frequently occurring reptile species (Ekner et al. 2008; Borczyk et al. 2022; Jurczyk and Borczyk 2022). Lacerta agilis can be found in a wide range of habitats and occur in western, central, and eastern Europe as well as in western and central Asia, while the range of Z. vivipara covers most of Europe and northern Asia. However, especially in Western Europe, both species are endangered and declining (Ekner et al. 2008). Our study confirms the occurrence of L. agilis and Z. vivipara in urban areas of Wrocław (SW Poland) and indicates their important role as hosts of immature stages of Ixodes ricinus. The role of L. agilis and Z. vivipara in maintaining I.ricinus population in urban areas was also documented in western Poland (Gwiazdowicz et al. 2020). In our study, we find that the importance of L. agilis in maintaining the tick population seems to be much higher compared to Z. vivipara. The greater number of ticks infesting L. agilis compared to Z. vivipara (in our study we collected an average of 31.1 ticks per one infested specimen of L. agilis and 6.1 ticks per Z. vivipara), may be related to greater mobility of L. agilis than Z. vivipara (Ekner et al. 2008) as well as in the differences in microhabitat preferences between the species (Borczyk et al. 2022). In contrast, Gwiazdowicz et al. (2020) found that females of L. agilis had lower infestation than Z. vivipara. The sex of the lizards may have a significant impact on the level of infestation because Dudek et al. (2016) found that males of L. agilis lizards are more frequently parasitized by ticks than females. Wieczorek et al. (2020), on the other hand, analyzing ticks attacking L. agilis on the outskirts of the city (Zielona Góra, Poland) did not confirm the correlation between the sex of lizards and the level of infestation by ticks, but they draw attention to body mass. The significant role of L. agilis in maintaining I. ricinus population in Poland was reported previously (Gryczyńska-Siemiątkowska et al. 2007; Gwiazdowicz et al. 2020; Ekner et al. 2011). L. agilis as a host of I. ricinus was also found in other European countries, including Hungary (Földvári et al. 2009), the Netherlands (Tijsse-Klasen et al. 2010), Germany (Richter and Matuschka 2006), Sweden (Olsson et al. 2000).
In our study, DNA of Borrelia spp. was detected in I. ricinus collected from both lizard species, however, a higher percentage of infected ticks was found in L. agilis than Z. vivipara, and from L. agilis both larvae and nymphs were infected, while from Z. vivipara only one nymph. Among identified Borrelia the dominant genospecies was Borrelia lusitaniae (88.0% positive samples). Borrelia lusitaniae was also the only identified Borrelia species in nymph from Z. vivipara and in larvae from L. agilis. A greater diversity of Borrelia was found in nymphs from L. agilis, because in addition to B. lusitaniae, also B. afzelii and B. miyamotoi were detected. Moreover, B. miyamotoi occurred in 28.5% of cases with Borrelia, including 9.5% single Borrelia infection and 19.0% co-infection with B. lusitaniae. According to our knowledge, this is the first report of the presence of B. miyamotoi in ticks collected from L. agilis. In an earlier study conducted in the area of Ranzenberg Mountain, Germany by Richter and Matuschka (2006), the occurrence of B. miyamotoi was observed in one nymph obtained from a common wall lizard (Podarcis muralis). However, the presence of B. miyamotoi in nymphs collected from lizards may be the result of acquiring the pathogen from the blood of the first host on which the tick was parasitized in the larva stage. It cannot also be excluded, that B. miyamotoi was transovarially to larvae and from larvae to nymphs transstadially transmitted, because B. miyamotoi, unlike B. burgdorferi, is capable to transovarial transmission in ticks of the Ixodes species (Rollend et al. 2013). Confirmation of the role of sand lizards in maintaining B. miyamotoi acquired more studies and sampling of lizard tissue.
The presence of B. lusitaniae in I. ricinus collected from L. agilis is detected also in other studies. Ekner et al. (2011) during a study carried out in the years 2008–2009 in the Barycz Valley, in Polan found DNA of B. burgdorferi s.l. in 4.1% ticks, including predominant B. lusitaniae and less frequently B. burgdorferi s.s. Gryczyńska-Siemiątkowska et al. (2007) in 2002–2003 in the Mazurian Lake region (Northeastern Poland) detected in 4.7% ticks parasitized sand lizards DNA of B. burgdorferi s.l., including B. afzelii, B. garinii and B. burgdorferi s.s. In the area near the town Odolanów, SW Poland, prevalence reached 6% with B. lusitaniae as the dominant genospecies and less frequently B. valaisiana (Majláthová et al. 2008). In Slovakia, the only detected genospecies in nymphs were B. lusitaniae, and B. lusitaniae and B. afzelii in larvae (Majláthová et al. 2008). While in the Netherlands, B. burgdorferi s.s and Borrelia afzelii were identified in ticks collected from sand lizards and no B. lusitaniae was detected (Tijsse-Klasen et al. 2010).
Xenodiagnosis, being the strongest method used for estimating vector competence, confirms the reservoir competence of Psammodromus algirus for B. lusitaniae (Dsouli et al. 2006; Wolcott et al. 2021). However, reservoir competence for B. lusitaniae seems to have more lizard species, including L. agilis, and the role of L. agilis in maintaining B. lusitaniae is evidenced by lizard studies. In Poland, DNA of B. lusitaniae was detected in 1.2% of L. agilis collected in the Barycz Valley (Ekner et al. 2011). In Hungary, B. lusitaniae was found in tissue samples of green lizards Lacerta viridis, Balkan wall lizards Podarcis taurica, and L. agilis, although the presence of Borrelia in ticks collected from L. agilis has not been confirmed (Földvári et al. 2009). Biopsy specimens from L. agilis positive for B. lusitaniae were also found in 45% of the sand lizards from Slovakia and 57% from Romania, although the same studies have not confirmed the presence of B. burgdorferi s.l. in samples of L. agilis from Poland (Majláthová et al. 2008). Many studies confirmed, however, the presence of B. lusitaniae both in lizard and parasiting ticks. Borrelia both in L. agilis and I. ricinus was found in Slovakia and in Romania (Majláthová et al. 2008), in the Czech Republic in Lacerta viridis and I.ricinus (Musilová et al. 2022), in Italy, in Podarcis muralis and I. ricinus (Amore et al. 2007), in Slovakia in Lacerta viridis and I. ricinus (Majláthová et al. 2006), in Italy in Podarcis siculus and I. ricinus (Mendoza-Roldan et al. 2019).
The most common among the bacterial pathogens detected in our study was Rickettsia spp. because 19.3% of ticks (both larvae and nymphs) were infected, including 27.2% ticks from Z. vivipara (23.2% larvae, 35.3% nymphs) and 17.0% from L. agilis (7.1% larvae, 44.4% nymphs). Rickettsia spp. found in ticks collected from sand lizards in the Netherlands were also identified as R. helvetica (Tijsse-Klasen et al. 2010) and in Slovakia in L. viridis (Václav et al. 2011). A high level of Rickettsia infection in ticks from lizards was also found in the Iberian Peninsula, where Kubelová et al. (2015) detected rickettsial DNA in 47% of nymphs and 31.6% of larvae collected from Iberian lizard Lacerta schreiberi. However, taking into account that most Rickettsia species can be transmitted transovarially, the infection in larvae may occur without the need to parasitize on the reservoir hosts.
Unexpectedly, we detected DNA of Anaplasma phagocytophilum in I. ricinus larvae collected from Lacerta agilis. However recently, the possibility of transovarial transmission of A. phagocytophilum is being considered because A. phagocytophilum was found in questing I. ricinus larvae (Jahfari et al. 2014, Hauck et al. 2020b). It can suggest that transovarial transmission in ticks might occur, however with low efficiency. The possibility of the occurrence of Anaplasma in L. agilis is also supported by molecular studies conducted by Ekner et al. 2011), who found DNA of Anaplasmataceae both in I. ricinus collected from L. agilis, and additional, for the first time, in L. agilis. The occurrence of Anaplasma/Ehrlichia spp. was also found in ticks from sand lizards in the Netherlands. however, the prevalence was significantly lower than in questing ticks (Tijsse-Klasen et al. 2010). Whereas, a relatively high percentage of infection with A. phagocytophilum of ticks collected from L. viridis found Václav et al. (2011) in Slovakia, who estimated the infection rate at 13.1% for nymphs and 8.7% for larvae.
Knowledge of transmission ways is crucial for understanding the complete lifecycle of A. phagocytophilum, including the possible role of lizard in A. phagocytophilum maintenance.
Conclusion
Lacerta agilis and Zootoca vivipara in peri-urban areas are among the important host of immature stages of Ixodes ricinus. However, the role of L. agilis seems to be greater compared to Z. vivipara. The high levels of B. lusitaniae infections in ticks that had fed on L. agilis lizards may point to implicates this species of lizard in the transmission cycle of B. lusitaniae. However, the presence of B. miyamotoi and A. phagocytophilum detected for the first time in ticks collected from this lizard species requires further research to confirm their participation in the transmission cycle of these pathogens.
References
Amore G, Tomassone L, Grego E, Ragagli C, Bertolotti L, Nebbia P, Rosati S, Mannelli A (2007) Borrelia lusitaniae in immature Ixodes ricinus (Acari: Ixodidae) feeding on common wall lizards in Tuscany, central Italy. J Med Entomol 44(2):303–307. https://doi.org/10.1603/0022-2585(2007)44[303:bliiir]2.0.co;2
Bakken JS, Dumler S (2008) Human granulocytic anaplasmosis. Infect Dis Clin North Am 22(3):433–448. https://doi.org/10.1016/j.idc.2008.03.011
Bennett D, Hampson K, Yngente V (2001) A noose trap for catching a large arboreal lizard, Varanus olivaceus. Herpet Rev 32:167–168
Borczyk B, Gottfried I, Urban RG, Kania J (2022) Great capricorn beetle-created corridors as refuges for lizards. Herpetozoa 35:59–63. https://doi.org/10.3897/herpetozoa.35.e81190
Dsouli N, Younsi-Kabachii H, Postic D, Nouira S, Gern L, Bouattour A (2006) Reservoir role of lizard Psammodromus algirus in transmission cycle of Borrelia burgdorferi Sensu Lato (Spirochaetaceae) in Tunisia. J Med Entomol 43(4):737–742. https://doi.org/10.1603/0022-2585(2006)43[737:rrolpa]2.0.co;2
Dudek K, Skórka P, Sajkowska ZA, Ekner-Grzyb A, Dudek M, Tryjanowski P (2016) Distribution pattern and number of ticks on lizards. Ticks Tick Borne Dis 7(1):172–179. https://doi.org/10.1016/j.ttbdis.2015.10.014
Dumler JS, Choi KS, Garcia-Garcia JC, Barat NS, Scorpio DG, Garyu JW, Grab DJ, Bakken JS (2005) Human granulocytic anaplasmosis and Anaplasma phagocytophilum. Emerg Infect Dis 11(12):1828–1834. https://doi.org/10.3201/eid1112.050898
Eisen L (2023) Rodent-targeted approaches to reduce acarological risk of human exposure to pathogen-infected Ixodes ticks. Ticks Tick Borne Dis 14(2):102119. https://doi.org/10.1016/j.ttbdis.2023.102119
Ekner A, Majláth I, Majláthová V, Hromada M, Bona M, Antczak M, Bogaczyk M, Tryjanowski P (2008) Densities and morphology of two co-existing lizard species (Lacerta Agilis and Zootoca vivipara) in extensively used farmland in Poland. Folia Biol (Krakow) 56(3–4):165–171. https://doi.org/10.3409/fb.56_3-4.165-171
Ekner A, Dudek K, Sajkowska Z, Majláthová V, Majláth I, Tryjanowski P (2011) Anaplasmataceae and Borrelia burgdorferi Sensu Lato in the sand lizard Lacerta Agilis and co-infection of these bacteria in hosted Ixodes ricinus ticks. Parasit Vectors 4:182. https://doi.org/10.1186/1756-3305-4-182
Estrada-Peña A, Michalca AD, Peteney TN (2017) Ticks of Europe and North Africa. A guide to specimen identification. Springer Int Publ. https://doi.org/10.1111/mve.12410
Földvári G, Rigó K, Majláthová V, Majláth I, Farkas R, Pet’ko B (2009) Detection of Borrelia burgdorferi sensu lato in lizards and their ticks from Hungary. Vector Borne Zoonotic Dis 9(3):331–336. https://doi.org/10.1089/vbz.2009.0021
Filipiak K, Zaręba A (2014) Lasy podmiejskie Wrocławia: Las Rędziński. Acta Sci Pol 13(4): 57–70
Fraenkel CJ, Garpmo U, Berglund J (2002) Determination of novel Borrelia genospecies in Swedish Ixodes ricinus ticks. J Clin Microbiol 40(9):3308–3312. https://doi.org/10.1128/jcm.40.9.3308-3312.2002
Gryczyńska-Siemiątkowska A, Siedlecka A, Stańczak J, Barkowska M (2007) Infestation of sand lizards [Lacerta Agilis] resident in the Northeastern Poland by Ixodes ricinus [L.] ticks and their infection with Borrelia burgdorferi sensu lato. Acta Parasitol 52:165–170
Guy EC, Stanek G (1991) Detection of Borrelia burgdorferi in patients with Lyme disease by the polymerase chain reaction. J Clin Pathol 44(7):610–611
Gwiazdowicz DJ, Gdula AK, Kurczewski R, Zawieja B (2020) Factors influencing the level of infestation of Ixodes ricinus (Acari: Ixodidae) on Lacerta Agilis and Zootoca vivipara (Squamata: Lacertidae). Acarologia 60(2):390–397. https://doi.org/10.24349/acarologia/20204372
Hauck D, Springer A, Chitimia-Dobler L, Strube C (2020b) Two-year monitoring of tick abundance and influencing factors in an urban area (city of Hanover, Germany). Ticks Tick Borne Dis 11(5):101464. https://doi.org/10.1016/j.ttbdis.2020.101464
Hauck D, Jordan D, Springer A, Schunack B, Pachnicke S, Fingerle V, Strube C (2020a) Transovarial transmission of Borrelia spp., Rickettsia spp. and Anaplasma phagocytophilum in Ixodes ricinus under field conditions extrapolated from DNA detection in questing larvae. Parasit Vectors 13(1):176. https://doi.org/10.1186/s13071-020-04049-7
Jahfari S, Coipan EC, Fonville M, van Leeuwen AD, Hengeveld P, Heylen D, Heyman P, van Maanen C, Butler CM, Földvári G, Szekeres S, van Duijvendijk G, Tack W, Rijks JM, van der Giessen J, Takken W, van Wieren SE, Takumi K, Sprong H (2014) Circulation of four Anaplasma phagocytophilum ecotypes in Europe. Parasit Vectors 15;7:365. https://doi.org/10.1186/1756-3305-7-365
Jurczyk K, Borczyk B (2022) Body size structure and sex ratio in a population of the common lizard Zootoca vivipara (Lichtenstein, 1823) from SW Poland. Folia Biologica (Kraków) 70:107–112. https://doi.org/10.3409/fb_70-3.12
Karbowiak G, Biernat B, Stańczak J, Szewczyk T, Werszko J (2016a) The role of particular tick developmental stages in the circulation of tick-borne pathogens affecting humans in Central Europe. 3. Rickettsiae. Ann Parasitol 62(2):89–100. https://doi.org/10.17420/ap6202.38
Karbowiak G, Biernat B, Stańczak J, Werszko J, Wróblewski P, Szewczyk T, Sytykiewicz H (2016b) The role of particular ticks developmental stages in the circulation of tick-borne pathogens in Central Europe. 4. Anaplasmataceae. Ann Parasitol 62(4):267–284. https://doi.org/10.17420/ap6204.62
Kim HK (2022) Rickettsia-host-tick interactions: knowledge advances and gaps. Infect Immun 90(9):e0062121. https://doi.org/10.1128/iai.00621-21
Kocoń A, Nowak-Chmura M, Asman M, Kłyś M (2023) Review of ticks attacking domestic dogs and cats, and their epidemiological role in the transmission of tick-borne pathogens in Poland. Ann Agric Environ Med 30(1):22–30. https://doi.org/10.26444/aaem/161552
Król N, Kiewra D, Szymanowski M, Lonc E (2015) The role of domestic dogs and cats in the zoonotic cycles of ticks and pathogens. Preliminary studies in the Wrocław Agglomeration (SW Poland). Vet Parasitol 214(1–2):208–212. https://doi.org/10.1016/j.vetpar.2015.09.028
Król N, Obiegala A, Pfeffer M, Lonc E, Kiewra D (2016) Detection of selected pathogens in ticks collected from cats and dogs in the Wrocław Agglomeration, South-West Poland. Parasit Vectors 9(1):351. https://doi.org/10.1186/s13071-016-1632-0
Kubelová M, Papoušek I, Bělohlávek T, de Bellocq JG, Baird SJ, Široký P (2015) Spotted fever group rickettsiae detected in immature stages of ticks parasitizing on Iberian endemic lizard Lacerta Schreiberi Bedriaga, 1878. Ticks Tick Borne Dis 6(6):711–714. https://doi.org/10.1016/j.ttbdis.2015.06.003
Majláthová V, Majláth I, Derdáková M, Víchová B, Pet’ko B (2006) Borrelia lusitaniae and green lizards (Lacerta Viridis), Karst Region, Slovakia. Emerg Infect Dis 12(12):1895–1901. https://doi.org/10.3201/eid1212.060784
Majláthová V, Majláth I, Hromada M, Tryjanowski P, Bona M, Antczak M, Víchová B, Dzimko Š, Mihalca A, Peťko B (2008) The role of the sand lizard (Lacerta Agilis) in the transmission cycle of Borrelia burgdorferi sensu lato. Internat J Med Microbiol 298 S1:161–167. https://doi.org/10.1016/j.ijmm.2008.03.005
Massung RF, Slater K, Owens JH, Nicholson WL, Mather TN, Solberg VB, Olson JG (1998) Nested PCR assay for detection of granulocytic ehrlichiae. J Clin Microbiol 36:1090–5. https://doi.org/10.1128/JCM.36.4.1090-1095.1998
Mendoza-Roldan JA, Colella V, Lia RP, Nguyen VL, Barros-Battesti DM, Iatta R, Dantas-Torres F, Otranto D (2019) Borrelia burgdorferi (Sensu Lato) in ectoparasites and reptiles in southern Italy. Parasit Vectors 12(1):35. https://doi.org/10.1186/s13071-019-3286-1
Mihalca AD, Sándor AD (2013) The role of rodents in the ecology of Ixodes ricinus and associated pathogens in Central and Eastern Europe. Front Cell Infect Microbiol 3:56. https://doi.org/10.3389/fcimb.2013.00056
Miková K, Berthová L, Kalúz S, Kazimírová M, Burdová L, Kocianová E (2015) First detections of Rickettsia helvetica and R. monacensis in ectoparasitic mites (Laelapidae and Trombiculidae) infesting rodents in south-western Slovakia. Parasitol Res 114(7):2465–2472. https://doi.org/10.1007/s00436-015-4443-x
Musilová L, Kybicová K, Fialová A, Richtrová E, Kulma M (2022) First isolation of Borrelia lusitaniae DNA from green lizards (Lacerta Viridis) and Ixodes ricinus ticks in the Czech Republic. Ticks Tick Borne Dis 13(2):101887. https://doi.org/10.1016/j.ttbdis.2021.101887
Olsson M, Wapstra E, Madsen T, Silverin B (2000) Testosterone, ticks and travels: a test of the immunocompetence–handicap hypothesis in free-ranging male sand lizards. Proc Biol Sci 267(1459):2339–2343. https://doi.org/10.1098/rspb.2000.1289
Platonov AE, Karan LS, Kolyasnikova NM, Makhneva NA, Toporkova MG, Maleev VV, Fish D, Krause PJ (2011) Humans infected with relapsing fever spirochete Borrelia miyamotoi, Russia. Emerg Infect Dis 17(10):1816–1823. https://doi.org/10.3201/eid1710.101474
Portillo A, Santibáñez S, García-Álvarez L, Palomar AM, Oteo JA (2015) Rickettsioses in Europe. Microbes Infect 17(11–12):834–838. https://doi.org/10.1016/j.micinf.2015.09.009
Prakash J, Sohan Lal T, Rosemol V, Verghese V, Pulimood S, Reller M, Dumler J (2012) Molecular detection and analysis of spotted fever group Rickettsia in patients with fever and rash at a tertiary care centre in Tamil Nadu, India. Pathog Glob Health 106(1):40–45. https://doi.org/10.1179/2047773212Y.0000000001
Richter D, Matuschka FR (2006) Perpetuation of the Lyme Disease spirochete Borrelia lusitaniae by lizards. Appl Environ Microbiol 72(7):4627–4632. https://doi.org/10.1128/AEM.00285-06
Rizzoli A, Silaghi C, Obiegala A, Rudolf I, Hubálek Z, Földvári G, Plantard O, Vayssier-Taussat M, Bonnet S, Spitalská E, Kazimírová M (2014) Ixodes ricinus and its transmitted pathogens in urban and peri-urban areas in Europe: new hazards and relevance for public health. Front Public Health 2:251. https://doi.org/10.3389/fpubh.2014.00251
Rochlin I, Toledo A (2020) Emerging tick-borne pathogens of public health importance: a mini-review. J Med Microbiol 69(6):781–791. https://doi.org/10.1099/jmm.0.001206
Rollend L, Fish D, Childs J-E (2013) Transovarial transmission of Borrelia spirochetes by Ixodes scapularis: a summary of the literature and recent observations. Ticks Tick Borne Dis 4(1–2):46–51. https://doi.org/10.1016/j.ttbdis.2012.06.008
Sykes RA, Makiello P (2017) An estimate of Lyme borreliosis incidence in Western Europe†. J Public Health (Oxf) 39(1): 74–81. https://doi.org/10.1093/pubmed/fdw017
Tijsse-Klasen E, Fonville M, Reimerink JHJ, van der Spitzen A, Sprong H (2010) Role of sand lizards in the ecology of Lyme and other tick-borne diseases in the Netherlands. Parasit Vectors 3:42. https://doi.org/10.1186/1756-3305-3-42
Uspensky I (2014) Tick pests and vectors (Acari: Ixodoidea) in European towns: introduction, persistence and management. Ticks Tick Borne Dis 5(1):41–47. https://doi.org/10.1016/j.ttbdis.2013.07.011
Václav R, Ficová M, Prokop P, Betáková T (2011) Associations between coinfection prevalence of Borrelia lusitaniae, Anaplasma sp., and Rickettsia sp. in hard ticks feeding on reptile hosts. Microb Ecol, 61(2)L 245–253. https://doi.org/10.1007/s00248-010-9736-0
Wieczorek M, Rektor R, Najbar B, Morelli F (2020) Tick parasitism is associated with home range area in the sand lizard, Lacerta Agilis. Amphib Reptil 41(4):479–488. https://doi.org/10.1163/15685381-bja10018
Wodecka B (2011) flaB gene as a molecular marker for distinct identification of Borrelia species in environmental samples by the PCR-Restriction fragment length polymorphism method. Appl Environ Microbiol 77(19):7088–7092. https://doi.org/10.1128/AEM.05437-11
Wodecka B, Rymaszewska A, Sawczuk M, Skotarczak B (2009) Detectability of tick-borne agents DNA in the blood of dogs, undergoing treatment for borreliosis. Ann Agric Environ Med 16(1):9–14
Wolcott KA, Margos G, Fingerle V, Becker NS (2021) Host association of Borrelia burgdorferi sensu lato: a review. Ticks Tick Borne Dis 12(5):101766. https://doi.org/10.1016/j.ttbdis.2021.101766
Author information
Authors and Affiliations
Contributions
D.D, A.K. and D.K. wrote the main manuscript text. D.D., A.K., B.B. and A.K. collected data. D.D., A.K. and D.K analyzed the data. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Dyczko, D., Krysmann, A., Kolanek, A. et al. Bacterial pathogens in Ixodes ricinus collected from lizards Lacerta agilis and Zootoca vivipara in urban areas of Wrocław, SW Poland– preliminary study. Exp Appl Acarol 93, 409–420 (2024). https://doi.org/10.1007/s10493-024-00927-2
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10493-024-00927-2