
Abstract
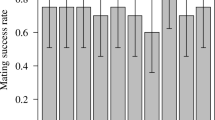
Generally, males increase their reproductive success by mating with as many females as possible, whereas females increase their reproductive success by choosing males who provide more direct and indirect benefits. The difference in reproductive strategy between the sexes creates intense competition among males for access to females, therefore males spend much energy and time for competition with rival males for their reproduction. However, if they do not need to engage themselves into male competition and females are in no short supply, how many females can a male mate with and fertilize? We address this question in the two-spotted spider mite, Tetranychus urticae Koch. In this study, we investigated how many females a young, virgin male mated in 3 h, and checked whether the mated females were fertilized. We found that on average males mated with 12–13 females (range: 5–25). As latency to next mating did not change with the number of matings, the males are predicted to engage in even more matings if the mating trial were continued beyond 3 h. Copulation durations decreased with the number of matings and typically after 11 copulations with females any further copulations did not lead to fertilization, suggesting that males continued to mate with females even after sperm depletion. We discuss why spider mite males continue to display mating and copulation behaviour even after their sperm is depleted.




Similar content being viewed by others


References
Abe J (2019) Sperm-limited males continue to mate, but females cannot detect the male state in a parasitoid wasp. Behav Ecol Sociobiol 73:52. https://doi.org/10.1007/s00265-019-2653-4
Andersson M (1994) Sexual selection. Princeton University Press, Princeton
Andersson M, Simmons LW (2006) Sexual selection and mate choice. Trends Ecol Evol 21:296–302. https://doi.org/10.1016/j.tree.2006.03.015
Bateman AJ (1948) Intra-sexual selection in Drosophila. Heredity 2:349–368. https://doi.org/10.1038/hdy.1948.21
Brooks ME, Kristensen K, van Benthem KJ, Magnusson A, Berg CW, Nielsen A, Skaug HJ, Maechler M, Bolker BM (2017) glmmTMB: Balances speed and flexibility among packages for zero-inflated generalized linear mixed modeling. R J 9:378–400
Cone WW (1985) Mating and chemical communication. In: Helle W, Sabelis MW (eds) Spider mites: their biology, natural enemies and control, vol 1A. Elsevier, Amsterdam, pp 243–252
Damiens D, Boivin G (2005) Male reproductive strategy in Trichogramma evanescens: sperm production and allocation to females. Physiol Entomol 30:241–247. https://doi.org/10.1111/j.1365-3032.2005.00453.x
Damiens D, Boivin G (2006) Why do sperm-depleted parasitoid males continue to mate? Behav Ecol 17:138–143. https://doi.org/10.1093/beheco/arj009
Emlen DJ (2008) The evolution of animal weapons. Annu Rev Ecol Evol Syst 39:387–413. https://doi.org/10.1146/annurev.ecolsys.39.110707.173502
Garcia-Gonzalez F (2004) Infertile matings and sperm competition: the effect of “nonsperm representation” on intraspecific variation in sperm precedence patterns. Am Nat 164:457–472. https://doi.org/10.1086/423987
Golizadeh A, Ghavidel S, Razmjou J et al (2017) Comparative life table analysis of Tetranychus urticae Koch (Acari: Tetranychidae) on ten rose cultivars. Acarologia 57:607–616. https://doi.org/10.24349/acarologia/20174176
Gross MR (1996) Alternative reproductive strategies and tactics: diversity within sexes. Trends Ecol Evol 11:92–98. https://doi.org/10.1016/0169-5347(96)81050-0
Helle W (1967) Fertilization in the two-spotted spider mite (Tetranychus urticae: Acari). Entomol Exp Appl 10:103–110. https://doi.org/10.1111/j.1570-7458.1967.tb00049.x
Jacob S, Boivin G (2004) Insemination potential of male Trichogramma evanescens. Entomol Exp Appl 113:181–186. https://doi.org/10.1111/j.0013-8703.2004.00221.x
Kelly CD, Jennions MD (2011) Sexual selection and sperm quantity: meta-analyses of strategic ejaculation. Biol Rev 86:863–884. https://doi.org/10.1111/j.1469-185X.2011.00175.x
King BH (2000) Sperm depletion and mating behavior in the parasitoid wasp Spalangia cameroni (Hymenoptera: Pteromalidae). Great Lakes Entomol 33:117–127
Koveos DS, Kroon A, Veerman A (1993) Geographic variation of diapause intensity in the spider mite Tetranychus urticae. Physiol Entomol 18:50–56. https://doi.org/10.1111/j.1365-3032.1993.tb00448.x
Krainacker DA, Carey JR (1989) Reproductive limits and heterogeneity of male twospotted spider mites. Entomol Exp Appl 50:209–214. https://doi.org/10.1111/j.1570-7458.1989.tb01194.x
Ludecke D, Ben-Shachar MS, Patil I, Waggoner P, Nakowski D (2021) performance: an R package for assessment, comparison and testing of statistical models. J Open Source Softw 6:3139. https://doi.org/10.21105/joss.03139
Macke E, Magalhães S, Bach F, Olivieri I (2011) Experimental evolution of reduced sex ratio adjustment under local mate competition. Science 334:1127–1129. https://doi.org/10.1126/science.1212177
Macke E, Magalhães S, Do-Thi Khanh H et al (2012) Mating modifies female life history in a haplodiploid spider mite. Am Nat 179:E147–E162. https://doi.org/10.1086/665002
Morita A, Ullah MS, Sera T, Gotoh T (2020) Effectiveness of second mating in sperm-depleted females in the two-spotted spider mite, Tetranychus urticae (Acari: Tetranychidae). Syst Appl Acarol. https://doi.org/10.11158/saa.25.9.4
Pérez-Lachaud G (2010) Reproductive costs of mating with a sibling male: sperm depletion and multiple mating in Cephalonomia hyalinipennis. Entomol Exp Appl 137:62–72. https://doi.org/10.1111/j.1570-7458.2010.01037.x
Potter DA, Wrensch DL (1978) Interrupted matings and the effectiveness of second inseminations in the twospotted spider mite. Ann Entomol Soc Am 71:882–885. https://doi.org/10.1093/aesa/71.6.882
Potter DA, Wrensch DL, Johnston DE (1976a) Aggression and mating success in male spider mites. Science 193:160–161. https://doi.org/10.1126/science.193.4248.160
Potter DA, Wrensch DL, Johnston DE (1976b) Guarding, aggressive behavior, and mating success in male twospotted spider mites. Ann Entomol Soc Am 69:707–711. https://doi.org/10.1093/aesa/69.4.707
Puurtinen M, Fromhage L (2017) Evolution of male and female choice in polyandrous systems. Proc R Soc B 284:20162174. https://doi.org/10.1098/rspb.2016.2174
R Core Team (2020) R: a language and environment for statistical computing. Foundation for Statistical Computing, Vienna
Ramadan MM, Wong TTY, Wong MA (1991) Influence of parasitoid size and age on male mating success of Opiinae (Hymenoptera:Braconidae), larval parasitoids of fruit flies (Diptera:Tephritidae). Biol Control 1:248–255. https://doi.org/10.1016/1049-9644(91)90074-A
Rodrigues LR, Figueiredo ART, Van Leeuwen T et al (2020) Costs and benefits of multiple mating in a species with first-male sperm precedence. J Anim Ecol 89:1045–1054. https://doi.org/10.1111/1365-2656.13171
Sato Y, Sabelis MW, Egas M, Faraji F (2013) Alternative phenotypes of male mating behaviour in the two-spotted spider mite. Exp Appl Acarol 61:31–41. https://doi.org/10.1007/s10493-013-9673-y
Sato Y, Sabelis MW, Egas M (2014) Alternative male mating behaviour in the two-spotted spider mite: dependence on age and density. Anim Behav 92:125–131. https://doi.org/10.1016/j.anbehav.2014.03.032
Sato Y, Rühr PT, Schmitz H et al (2016) Age-dependent male mating tactics in a spider mite—a life-history perspective. Ecol Evol 6:7367–7374. https://doi.org/10.1002/ece3.2489
Satoh Y, Yano S, Takafuji A (2001) Mating strategy of spider mite, Tetranychus urticae (Acari: Tetranychidae) males: postcopulatory guarding to assure paternity. Appl Entomol Zool 36:41–45. https://doi.org/10.1303/aez.2001.41
Schausberger P, Sato Y (2019) Parental effects of male alternative reproductive tactics (ARTs) on ARTs of haploid sons. Funct Ecol 33:1684–1694. https://doi.org/10.1111/1365-2435.13385
Schausberger P, Gotoh T, Sato Y (2019) Spider mite mothers adjust reproduction and sons’ alternative reproductive tactics to immigrating alien conspecifics. R Soc Open Sci 6:191201. https://doi.org/10.1098/rsos.191201
Scranton K, Stavrinides M, Mills NJ, de Valpine P (2013) Small-scale intraspecific life history variation in herbivorous spider mites (Tetranychus pacificus) is associated with host plant cultivar. PLoS ONE 8:e72980. https://doi.org/10.1371/journal.pone.0072980
Silk MJ, Harrison XA, Hodgson DJ (2020) Perils and pitfalls of mixed-effects regression models in biology. PeerJ 8:e9522. https://doi.org/10.7717/peerj.9522
Steiner S, Henrich N, Ruther J (2008) Mating with sperm-depleted males does not increase female mating frequency in the parasitoid Lariophagus distinguendus. Entomol Exp Appl 126:131–137. https://doi.org/10.1111/j.1570-7458.2007.00641.x
Yano S, Takafuji A (2002) Variation in the life history pattern of Tetranychus urticae (Acari: Tetranychidae) after selection for dispersal. Exp Appl Acarol 27:1–10. https://doi.org/10.1023/A:1021518221031
Acknowledgements
We thank students who joined the practice courses in 2020: ‘Laboratory and field studies in biodiversity of model organism’ and ‘Advanced field course in the ecology of selected model organisms’, Ms. Yukine Kawamura, Ms. Nanako Yasuhara, Ms. Misaki Ono, Mr. Yakumo Watanabe and Mr. Takunori Yasuda in University of Tsukuba for their help in mating behaviour observation in T. urticae. We thank Mr. Gomei Yoda, Mr. Naoki Matsumoto and Mr. Shota Konaka for their help in the research project. We also thank Drs. Yosuke Degawa, Natsumi Kanzaki and Yoshiaki Iwamoto for their cooperation in the research project. This research was supported in part by JSPS KAKENHI Grant Number 20K06810 (Grant-in-Aid for Scientific Research C to YS).
Author information
Authors and Affiliations
Contributions
YS and HK contributed to the study conception and design. Material preparation, data collection and analysis were performed by HK and YS. The first draft of the manuscript was written by YS and ME and all authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Conflict of interest
The authors have no conflict of interest to declare.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Supplementary file1 (TIFF 879 kb)
Figure S1 Copulation duration (a) and latency to mate (b) along sequential number of matings (mating order) in each male of Tetranychus urticae. Results from the same male are connected by grey dotted lines along time (h). Bold lines and filled areas along the lines in (b) indicate the predicted copulation durations and the 95% confidence interval by the models for series A and B in Table 2b, c. In (a) no line is shown because mating order did not have a significant effect on latency to mate
Supplementary file2 (TIFF 879 kb)
Figure S1 Copulation duration (a) and latency to mate (b) along sequential number of matings (mating order) in each male of Tetranychus urticae. Results from the same male are connected by grey dotted lines along time (h). Bold lines and filled areas along the lines in (b) indicate the predicted copulation durations and the 95% confidence interval by the models for series A and B in Table 2b, c. In (a) no line is shown because mating order did not have a significant effect on latency to mate
Supplementary file3 (TIFF 879 kb)
Figure S2 Female ratio in offspring along sequential number of matings (mating order) in each male of Tetranychus urticae. Results from the same male are connected by grey dotted lines along time (h). The bold solid line and filled area along the line indicate the predicted female fertilization status and the 95% confidence interval by the model in Table 4c
Rights and permissions
About this article

Cite this article
Kobayashi, H., Sato, Y. & Egas, M. Males mate with females even after sperm depletion in the two-spotted spider mite. Exp Appl Acarol 86, 465–477 (2022). https://doi.org/10.1007/s10493-022-00706-x
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10493-022-00706-x