
Abstract
Understanding the growth efficiency of individual trees, or growth per unit of resource utilization, can inform silvicultural management strategies to maximize tree and stand growth. Stand structure—the size and spatial distributions of trees within the stand—strongly influences water, light, and nutrient availability, as well as the resource-use efficiency of each tree. Key silvicultural tools for stand management include manipulating tree density, size distribution, and arrangement by controlling natural regeneration, artificial seeding, planting seedlings, and/or subsequent thinning of established trees. We analyzed two sets of plots from even-aged stands of common coniferous species in central Spain, 106 pure Scots pine (Pinus sylvestris) and 92 pure Mediterranean pine (Pinus pinaster), to examine the dynamics of the dominance hypothesis, the relationship between stand structure and growth, and the relationship between structure and growth efficiency. Our main findings revealed a negative impact of size-class uniformity on stand growth in both Scots pine and Mediterranean pine, while the positive effect of tree size on growth efficiency was supported for Mediterranean pine stands but uncertain for Scots pine. At the operational level, our results highlight how thinning intensity is more important than the thinning method in Mediterranean pinewoods and how thinning can benefit the provision of multiple ecosystem services. We also recommend integrating dominance effects on growth into individual tree modelling.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Manipulating stand structure is fundamental to silvicultural control of net primary production in relation to three general stand attributes: light interception, as determined by total foliage mass and spatial distribution (Wang and Jarvis 1990; Maguire and Bennett 1996; Xu and Harrington 1998; Garber and Maguire 2005); hydraulic limitations on gas exchange imposed by tree height and branching pattern (Ryan and Yoder 1997; Ryan et al. 1997; Bond et al. 2007); and growth and maintenance respiration of various tree tissues (Maguire and Batista 1996; Ryan et al. 2004). In a mechanistic view of forest growth, foliage mass determines the amount of intercepted solar radiation, while the other attributes influence how efficiently solar energy converts CO2 and H2O into forest biomass. Structural differences imposed by silvicultural manipulation are therefore expected to induce differences in the efficiency of converting solar energy into total forest biomass and its allocations to harvestable tree components. Thinning, for example, has been shown to improve the water status of the remaining trees in water-limited forests (Moreno-Gutiérrez et al. 2011) and to increase growth and seed production (Ruano et al. 2013).
Silvicultural manipulation of the dimensions listed above is typically achieved by controlling tree density, size distribution and spatial arrangement (Asmann 1970; O’Hara 1988; O’Hara 1996; Pretzsch and Hilmers 2024). The implications of such activities for different types of growth efficiency at stand level and individual tree level have been explored in diverse forest compositions (Waring et al. 1980; Long and Smith 1990; Maguire et al. 1998). The most practical silvicultural definition of growth efficiency combines some measurement of above-ground net primary production (ANPP) with a measurement of growing space occupancy (GSO) (Mainwaring and Maguire 2004). Volume or mass are the most common surrogates for ANPP, partly because efficiency has often been viewed from the perspective of timber or merchantable biomass production. Biologically appealing surrogates for GSO would be the amount of foliage (Waring et al. 1980) or crown surface area (Hamilton 1969), given their implications for capturing primarily solar energy. The proportionality of these surrogates to the capture of below-ground resources (water and nutrients) remains unclear, though some evidence suggests that water uptake may be proportional to tree stem size (Fernández and Gyenge 2009). Operationally, the most convincing surrogates for GSO would translate directly into the ground area occupied by the tree and subsequently inform about spacing between trees. Measures of space occupancy have therefore also included crown projection area (e.g., O’Hara 1988) and area potentially available (e.g., Mainwaring and Maguire 2004). Similarly, growth efficiency in terms of stem volume production per unit leaf area has also been used to estimate vigor and pest resistance in trees and stands (Waring et al 1980) and study the relationship between forest stand structure and tree growth (O’Hara 1988; Smith and Long 1989; Sterba and Amateis 1998).
Silvicultural research on individual tree growth efficiency can help us identify the most efficient producers of stem wood volume in stands of varying structure (O’Hara 1988, 1996) and characterize stand-level growth efficiency patterns for diverse canopy structures (Smith and Long 1989). Leaf area (O’Hara 1996), stand density index (Cochran et al. 1994), and other GSO measurements can provide growth efficiency information that helps forest managers determine strategic allocation of growing space. Binkley et al. (2002) and Binkley (2004) developed a hypothesis to explain plantation productivity peaking, which implies growth-efficiency losses as size- class differentiation increases over the course of stand development. Those authors suggest that dominant trees capture the greatest amount of total site resources, but growth per unit resource uptake declines for the whole stand. While some evidence supports this hypothesis of increasing dominance as a mechanism for declining stand productivity soon after crown closure (Binkley et al. 2002), other research contradicts it (Fernández and Gyenge 2009). The theory has been tested by comparing stand productivity over time with concurrent degrees of size differentiation as inter-tree competition intensifies, and (to a lesser extent) by comparing stand structures at a specific moment of time (e.g., Binkley et al. 2002). Changes in stand structure over time combine two separate processes: (1) the increasing size and changing physiology of trees as they age; and (2) differences in degree of size-class differentiation and other aspects of stand structure among stands of a given age. Numerous mechanisms may contribute to the universal decline of even-aged stand productivity over time (Ryan et al. 1997, Binkley 2023). However, since stand structure can be readily manipulated by conventional silvicultural operations, the dominance hypothesis may suggest opportunities for translating the observed or hypothesized effects of vertical differentiation into guidelines for optimizing stand structure, growth efficiency, and total stand production.
In this study, we analyzed productivity and productivity distribution in different strata of forest stand to gain knowledge about how altering vertical stratification and spatial distribution affects stand productivity. Such changes can be induced through silvicultural practices, which could be customized for each stand depending on the ecosystem services demanded by society in response to social, economic, ecological or global changes. The research is founded on two established datasets for pure stands (198 initial plots) covering a wide range of age, density, and productivity for Scots pine (Pinus sylvestris L.) and Mediterranean pine (Pinus pinaster Aiton spp. mesogeensis). The first of these two species is of great interest for its high wood quality and the second for its adaptability and frugality.
With the information collected from these databases, we sought to gain insight into the following questions:
-
1.
Do changes in stand structure contribute to increased differences in resource-use efficiency between dominant and non-dominant trees and to decreased growth at stand-level? (Binkley dominance hypothesis)?
-
2.
Does stand-level wood production in Scots pine and Mediterranean pine stands in Spain decline with increasing differentiation in size class and vertical structure?
-
3.
Does tree-level stem growth decline with increasing size differentiation and structural complexity?
-
4.
Does tree-level growth efficiency in Scots pine and Mediterranean pine stands decline with increasing size differentiation and structural complexity?
Material and methods
Field work
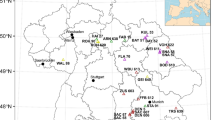
The datasets network used in this study was carefully designed to reflect the age distribution, densities and productivity levels of the study area (central Spain), based on previous Forest Service sampling. Two sets of plots were established between 2002 and 2004 for Pinus sylvestris L. (Scots pine) and Pinus pinaster Aiton ssp. mesogeensis (Mediterranean pine) (see Fig. 1). Individual tree data were available for 106 plots located in pure Scots pine stands and 92 plots in pure Mediterranean pine stands in central Spain (Tables 1 and 2). Stand age at time of plot establishment time ranged from 30 to 188 years for Scots pine and from 21 to 128 years for Mediterranean pine. The plots for both species represented the known range of site productivity, stand age and stand density (though only subsets of 50 and 37 Scots pine plots and 50 and 41 Mediterranean pine plots were remeasured in 2008 and 2013, respectively). None of the sampled stands showed unusual mortality events (tree mortality for all plots was only 5 trees in total) or had been thinned or tapped in the past 10 years. These dataset characteristics made them suitable for use in the study.

Distribution of Scot pine and Mediterranean pine plot dataset and location of the Central Spain area.
Each plot had a set of three nested, circular sub-plots with radii of 5, 10 and 15 meters. All trees with diameter at breast height (dbh) equal to or greater than 7.5, 12.5 and 22.5 cm, respectively, were tallied according to species for each plot and measurements were recorded for: dbh (nearest 0.1 cm); total height (ht) (nearest 0.1 m); height to crown base (hbc) (nearest 0.1 m); height to largest crown width (hlcw) (nearest 0.1 m); largest crown width (lcw), determined by measuring four perpendicular radii (nearest 0.1 m); and most recent five-year radial increment, determined by coring at breast height (nearest 1 mm) (Table 1).
Growth and stand structural computations
Our research questions focused on how growth efficiency is related to stand structure, and growth distribution among different-sized trees in the stand. Respiration capacity, space occupation, and leaf area index were chosen as key factors affecting productivity, represented by the surrogate variables of stem surface, crown projection and the ratio of crown surface to crown projection, respectively. In all cases, the form of the crown was established as an ellipsoid above the maximum crown width, with an inverted cone below.
Initial conditions and growth estimates were computed for three 5-years growth periods, including the 5-years period prior to plot establishment and the two 5-years growth periods following plot establishment. A reconstruction of the growth period just prior to plot establishment was included to cover the entire range of variability, as some plots could not be measured in subsequent growth periods due to mortality and technical impossibility. Variables for the initial 5-years period were computed from field measurements and pre-existing equations (e.g., site index curves, dominant height growth curves, bark thickness equations, as proposed by Hann and Hanus 2001). Back-dated tree dimensions provide unbiased estimates of permanent plot growth components (Wykoff 1990). They have been applied extensively to evaluate alternative measurements of competition (Biging and Dobbertin 1992, 1995; Bravo et al. 2001) and develop stand development models (Wensel et al. 1987; Wykoff 1990; Hann and Ritchie 1988; Hann and Larsen 1991). Variables for the two 5-years growth periods following plot installation were computed from the initial measurements and the two subsequent re-measurements. Plots with mortality between measurements were discarded. Stem volume (vol) and bole surface area (bsa) was computed from taper equations based on diameter and total height for each species (Lizarralde and Bravo 2003).
Productivity was considered constant throughout the study period and is closely related to the size of individual trees studied at different ages. Site Index (SI), calculated from curves previously developed for Scots pine (Rojo and Montero 1996; Bravo and Montero 2001) and Mediterranean pine (Bravo-Oviedo et al. 2004), was used as a proxy for productivity. After estimating the basic tree dimensions for the start of the initial 5-years growth period, additional tree and stand structural variables were calculated to describe initial conditions (Tables 1 and 2), including total basal area (BA), trees per ha (N), mean height (Hmean), and mean dbh (Dmean). Crown dimensions were assumed to have experienced no change over the course of the five-year growth period and were therefore the same at the start and finish of each initial growth period. Crown surface area (csa) was computed from basic crown measurements based on an ellipsoid above and a cone below maximum crown width, crown projected area (cpa) was computed based on a circle with lcw as diameter and leaf area index surrogate (treelai) was computed as the ratio between csa and cpa (Tables 3 and 4). Crown surface area was then expressed at stand level as the ratio of the summed surface area per unit of ground area (m2m−2; Tables 5 and 6).
For stand-level analyses, the key productivity variables (bole surface area, crown surface area and leaf area index surrogate) that were already calculated at the tree level had to be generalized to the forest stand level. Stand-level canopy cover was estimated by summing the crown projection areas for a plot and expressing the total as a proportion of the ground area covered by the crowns. Growth efficiency for individual trees was calculated as the ratio of periodic annual stem-volume growth to the crown projection area (GECPA).
In addition to the conventional stand-level variables listed above, two groups of indices were developed as measures of structural complexity. First, the Growth Dominance Coefficient (GDC) was calculated for each plot at the beginning of each 5-years growth period (West 2014). To construct the curves required for computing the GDC (see Fig. 2), the cumulative proportion of total stand volume growth (y-axis) was plotted against the cumulative initial stem volume of trees, ordered from lowest to highest initial volume (x-axis). If all trees grow proportional to their initial size, the curve is a straight diagonal line from (0,0) to (1,1). If smaller trees contribute a larger proportion to stand growth than their proportional contribution to initial stand volume, the curve moves above the diagonal. Conversely, if smaller trees contribute a smaller proportion to stand growth than their proportional contribution to initial stand volume, and if larger trees contribute a larger proportion, the curve falls below the diagonal. The GDC is the ratio of the area enclosed by the diagonal line of equality and the Lorenz curve to the total triangular area above or below the line of equality. If stem volume growth is perfectly proportional to initial stem volume, then the GDC is 0; it approaches 1 or −1 as proportional growth distribution departs from the initial proportional volume of individual trees (Table 6).

Development of the Growth Dominance Coefficient (GDC). Show the case of Dominance calculation and represent the Reverse Dominance line.
The second class of index characterized within-plot variability of tree dbh, tree height, crown projected area and crown surface area. Both the coefficient of variation (CV) and the Shannon-like index of diversity (H) were computed for these four attributes to determine if stand structural complexity contributed marginally to explaining differences in stand or tree productivity (Tables 5 and 6).
Statistical analyses
Testing the dominance hypothesis
The first step in exploring Binkley’s dominance hypothesis (Binkley et al. 2002; Binkley 2004) was to test the prevalence of growth dominance in the 198 plots. The Growth Dominance Coefficient (GDC) was calculated as previously described (West 2014). Figure 3 (Scots pine) and Fig. 4 (Mediterranean pine) show the curves used to construct the GDC for all plots, grouped by age.

GDC Graphs for Scots pine stands at different age ranges: (1) below 40 years, (2) between 40 and 70 years, (3) between 70 and 120 years, and (4) over 120 years.
GDC was then used to compute associated bootstrap errors (N = 2000 runs and α = 0.05) to test the null hypothesis that GDC equaled zero. We wanted to see if there was any relationship between this coefficient and the age, density or productivity of the stands studied, using graphical analysis that differentiated results that were significantly different from zero.

GDC Graphs for Mediterranean pine stands at different age ranges: (1) below 50 years, (2) between 50 and 70 years, (3) between 70 and 120 years, and (4) over 120 years.
Stand-level stem-volume production as a function of stand structure
To analyze volume growth at the stand level, we examined the relationship between volume growth and variables related to resource availability, then added in various structural variables and chose the two most significant. The base statistical model expressed periodic annual volume growth (m3 ha−1 years−1) as a function of surrogates for site productivity (SI, site index), respiratory load (total bole surface area), site occupancy (CANCOV, canopy cover), and photosynthetic potential or leaf area index (CSA, crown surface area). A mixed-effects model was fitted to the data to accommodate the repeated measurement structure (three 5-years growth periods for each plot) by considering growth period as a random effect. All models were fitted with PROC MIXED in SAS 9.4 (SAS Institute Inc, 2023). Akaike Information Criterion (AIC) was then applied to select the best log transformed and non-transformed models from those that contained statistically significant parameter estimates (α = 0.05), were biologically interpretable (sign and value of parameters) and met expected biological behavior. The resulting base model [1] for the two species was as follows:
where (VOLinc)ij is periodic annual volume increment of the ith plot in the jth growth period (m3ha−1 years−1), βls are parameters to be estimated from the data, and εij is the random error term for the ith plot in the jth growth period. The random effects were assumed to be normally distributed with mean zero and an unknown, unrestricted variance-covariance matrix. Therefore, the mixed model included six variance parameters for the repeated measurement factor, defined by the three five-year periods. These parameters were estimated with the Restricted Maximum Likelihood (REML) method and Akaike’s Criterion was used to select the best model. The R-square between observed and predicted values was calculated as a pseudo-measure of goodness of fit. The Kolmogorov–Smirnov test was used to check the normality of the studentized residuals. Logarithmic transformation of the dependent variable linearized the model and homogenized the variance to meet linear regression model assumptions.
To this model [1], we added a structure variable selected from the surrogates for size class or vertical differentiation (Model [2]). The structural variables tested (one each time) were Shannon-like diversity indices (H) or coefficients of variation (CV) for size variables (dbh, height, crown projection area and crown surface area) (Tables 5 and 6). The marginal effect of measures of structural diversity provides a test of the first hypothesis that stand-level productivity declines with increasing size differentiation, given a fixed total bole surface area, canopy closure, and LAI surrogate.
For each species, the selected base models for stand-level and tree-level growth (those with the smallest AIC) were compared with alternative expanded models to test for marginal effects of stand structure by using the percent reduction in mean squared error (MSE) relative to the base-model MSE.
Tree-level stem growth as a function of stand structure
To better understand growth response to structural changes, we analyzed volume growth at the tree level first using a base model for each species, then included variables related to structure and selected the two most significant variables.
In the base model (Eq. 3), the dependent variable was the logarithm of the periodic annual volume increment of the tree (dm3 years−1), and the explanatory variables were the surrogate variables for productivity (SI), tree size (volume), growing stock (bsa), occupancy (crown surface area) and leaf area index surrogate (treelai). A mixed-effect model similar to the previous one was fitted to the data to include the repeated measurement structure, considering growth period as a random effect, but here we added another random effect that considers trees within plots. We then used the same procedure to select the models, including log transformation as needed or dropping a variable if we were unable to achieve parameter significance.
This new model structure implies two random error effects for structure, \(\delta_{ij}\) for the i-plot at the five-year j-period, and \(\varepsilon_{ijk}\) for the tree k in the i-plot at the five-year j-period. These include nine variance parameters each: three for the random effect of the plot and six for the repeated measurements factor reflecting the three five-year periods.
To test the marginal effect of stand structure, the coefficient of variation and Shannon-like diversity index (CVs, and Hs) were considered as additional explanatory variables in the selected base model for periodic, annual, tree-volume growth (Eq. 4).
Tree-level stem-volume growth efficiency as a function of stand structure
To further explore the response of growth efficiency to structural changes, a tree-level analysis was designed using the same method as in the previous section, with a base model and an expanded model that included variables related to structure, from which the two most significant variables were selected.
We adjusted the mixed model to include growth efficiency as a response variable in the base equation: GECPA (Eq. 5).
To test the marginal effect of stand structure on growth efficiency after selecting the best-fitting variable transformations, we added the coefficient of variation, the Shannon-like diversity index (CVs, and Hs) and relative tree diameter or tree height to the base model as additional explanatory variables, to obtain the expanded model (Eq. 5):
Results
Test of the dominance hypothesis
Nearly 20% (21 of 107 plots) of the Scots pine dataset had a Growth Dominance Coefficient (GDC) significantly different from zero in growth from backdating to the first inventory, and between the first and second inventories in two plots. Mediterranean pine datasets had 47.83% non-zero GDC (44 out of 92 plots) in growth from backdating to the first inventory.
The graph depicting GDC versus Age (Fig. 5a) shows no clear evolution of dominance with age in any of the sampled stands, though the 4 plots presenting dominance were over 100 years old. However, most plots with significant GDC exhibited reverse dominance across the entire age range. Similar behavior was observed for density (Fig. 5b) and productivity (Fig. 5c), which presented no clear trend.

Growth Dominance Coefficient represented against main stand characteristics: (1) Age, (2) Stand Density Index (SDI) and (3) productivity (SI). In grey plots with values not significantly different from 0 after bootstrapping
Stand-level stem volume production
With this analysis, we wanted to assess whether increased structural diversity negatively affected wood production. The results indicated that volume growth slowed as structural diversity (represented by the Shannon-like index) increased, although the effect was more relevant in Scots pine stands, especially for the diversity of crown surface size. Extended models were also adjusted for the coefficient of variation (CV), but the results were worse than for the Shannon-like diversity index.
Explanatory variables for describing stand-level stem-volume growth were similar for both species, i.e., total bole surface area per hectare (log transformed), canopy cover (log transformed) and LAI surrogate (log transformed for Mediterranean pine stands) (Table 7). In these base models, stand-level growth increased with increasing bole surface area and average crown surface area but declined with increasing canopy cover (see mixed-model results in Supplementary Table S1).
To see the effect of structure on stand, the models were expanded with variables for structural diversity (H) and structural variability (CV). From the adjusted variables, the two expansions that gave the best result, or the lowest AIC, were selected.
The expanded models with better statistically significant structural variables for Scots pine included the Shannon-like diversity index for the two variables of crown size (HCSA) and height (HH), which reduced the MSE by 19% and 5% respectively (Table 7).
In Scots pine stands, growth declined with increasing size class uniformity, as indicated by the greater Shannon-like diversity index for crown surface (HCSA) and total height (HH). See marginal effects in Fig. 6.

Plot volume increment for a Scots pine and for b Mediterranean pine by mean initial height and site index. Each graph represents a different level of other covariates, including initial bole surface area (m2m−2), canopy coverage, and crown surface area (m2m−2). Plot volume increment for Scots pine by mean initial height and site index. Each graph represents a different level of other covariates, including initial basal area (m2m−2), canopy coverage, and crown surface area (m2m−2).
Introduction of stand structural variables reduced MSE much less for Mediterranean pine (3-4%; Table 7). The Shannon-like index for height (HH) and for crown surface area (HCSA) proved to be the best structural variables for this species. Stand growth declined with increasing height and crown-size diversity in Mediterranean pine stands (Fig. 6).
Tree-level models for stem-volume growth
This tree-level analysis assessed whether timber production decreased when size diversity and structural complexity increased, as occurs at stand level. The results revealed that volume growth decreased as height increased in Scots pine stands, indirectly leading to an increase in structural diversity, while increased crown-size diversity in Mediterranean pine stand, as represented by the Shannon-like index, also negatively affected wood productivity. The other structural variables analyzed were statistically less significant or not significant.
The Explanatory variables selected to describe tree-level stem growth in the base model were slightly different for each species. For Mediterranean pine stands, we followed the same stand-level analysis scheme, including productivity (SI), respiration surrogate (bsa), crown cover (csa), and LAI surrogate variables (Table 3). For Scots pine stands, however, we included volume instead of LAI surrogate as response variable because the latter lacked significance. Site index (SI) and volume (for Scots pine stands only and log transformed) were significant as standard variables in the base model. Bole surface area was selected as the respiration surrogate, (log transformed for both species), and crown surface area as the occupation surrogate (log transformed for Mediterranean pine stands). The leaf area index surrogate (log transformed) was included only for Mediterranean pine stands. Mixed base-model results can be found in Supplementary Tables S3 and S4.
To analyze the effect of structure on growth at the individual tree level, variables representing structure were added to the base model in a way that was analogous to the procedure used at the stand level.
The most suitable structural variable for Scots pine stands was maximum stand height, which reduced MSE by almost 28%, followed by average mean height (log transformed), which also reduced MSE by more than 26% (Table 8).
The variable selected for Mediterranean pine stands was relative dbh (ratio of dbh vs. maximum dbh), which reduced MSE by over 9%. Including the Shannon-like crown-surface diversity (HCSA) reduced MSE by over 7% (Table 8).
The results seem to indicate that structural diversity variables have a greater effect on volume growth in Scots pine stands compared to Mediterranean pine stands. This result seems consistent with the greater dependence of the first species on light resources, while the second species is more dependent on soil resources (see Fig. 7).

Periodic annual tree volume increment for: a Scots pine by mean height and site index; and b Mediterranean pine by mean dbh and site index. Each graph represents a different level of other covariates, including bole surface area (m2m−2), canopy coverage, and crown surface area (m2 m−2). It compares effect of two structure variable.
Tree-level models for growth efficiency
With this tree-level analysis, we wanted to determine if growth efficiency decreased with increasing size diversity and structural complexity. The results indicate that volume growth efficiency decreased as crown surface variability increased in Scots pine stands, and growth efficiency decreased with increasing height in Mediterranean pine stands. The other structural variables analyzed were statistically less significant or not significant.
Base equations to analyze growth efficiency included the same variables for both species: site index, volume, bole surface area, crown surface area, and tree leaf-area index surrogate (See details in supplementary Tables S5 y S6).
For Scots pine stands, increasing crown-size variability was associated with lower growth efficiency and reduced MSE by more than 18%. Including relative diameter reduced MSE by more than 19% but with a positive effect of increasing growth efficiency as dbh increased. For Mediterranean pine stands, stand average and maximum height variables representing structure negatively affected growth efficiency, reducing MSE by over 24% and 22%, respectively (see Table 9).
Crown-size differences negatively impacted tree growth efficiency for Scots pine while maximum stand high had similar effect in Mediterranean pine stands (Fig. 8). Both effects (total height increase and crown heterogeneity) had the expected effect on productivity (stands with higher site index grew faster than stands with lower site index)

Plot for growth efficiency for Scots pine and Mediterranean pine. It compares the alternative effect of two structure variables
Discussion
Size-class uniformity impacts negatively on growth of Scots pine and Medirrenean pine. Similar outcome was reported by Bravo and Guerra (2002) analyzing an intensive sampling plot in a Pinus pinaster stand in Central Spain. There was no clear support for Binkley’s hypothesis (Binkley et al. 2002 and Binkley 2004) that growth efficiency decreases at the stand level when size-class differentiation increases. Our results do not clearly show that dominant trees capture most of the total site resources as growth efficiency declines for the stand as a whole.
Instead, the analysis outcomes indicated that tree-size differentiation or initial inequality (expressed as dominance and reverse dominance) at the beginning of the growth period impacted stand growth. However, Forrester (2019) indicated that size inequality may be correlated with other variables that influence growth more strongly, such as seasonal productivity or stand density. Our findings support the idea that productivity modulates the impact of size inequalities on stand growth, but this may be affected by the negative relation between size inequality and productivity reported in experimental studies (Sun et al. 2018). Because no natural mortality events or thinning were included in our sampling of these observational studies, we could not analyze self-thinning and density regulation effects on size inequalities in the stand. Size inequality is more frequent in mixed-species stands with stratified canopy structures, potentially leading to higher tree densities and higher yield than monocultures of the overstory species (Pretzsch and Schütze 2015). The origin of the unequal size distribution may be correlated with the uneven growth distribution among trees of different species or among individual trees of the same species (Metsaranta and Lieffers 2007). The interaction between size distribution and stand productivity may make it difficult to understand the impact of size inequality on forest growth, which is more pronounced in years of low growth. Lin et al. (2021) found that forest structure is more important than site productivity for tree growth. In this study, the oldest stands on the best-quality sites were not fully represented because most had reached rotation age and been harvested prior to our sampling. The high-quality Mediterranean pine stands were also influenced to an unknown degree by resin tapping approximately 50 years ago.
The growth dominance effect observed in most of our plots gave larger trees a disproportionate growth and competition advantage (Nord-Larsen et al. 2006). This effect can be modulated through operational thinning, promotion of mixed-species stands, and forest manipulation to enhance size and structural complexity. However, Moore et al. (1994) found that thinning and fertilization did not change the relationship between tree size and tree growth; larger trees continued to capture more of the site resources.
Most definitions of growth efficiency assume that measurement of the growing space occupied can serve as a surrogate for resources captured by the tree: most commonly light but also soil water and nutrients. Comparing growth efficiency in different tree sizes can increase our understanding of the factors contributing to tree growth allocation within the stand and inform the design of thinning regimes to induce the desired growth response patterns. In Mediterranean forests, growth may be more limited by water availability than light availability, so an appropriate measure of growing space should ideally be correlated with the size of the root system. Thus, it remains unclear whether leaf area, crown projection area, or area potentially available (Asmann 1970) would be the most appropriate measure, because root systems overlap considerably more than tree crowns. In this regard, resource-use efficiency (Binkley et al. 2004) is a more precise concept because it directly measures the stem-volume growth per unit of consumed resource. However, our capacity for directly measuring resource use by trees is limited.
Forests typically grow until one or more of the factors necessary for growth become limiting, e.g., light, water, soil nutrients, oxygen, or carbon dioxide. “Growing space” ultimately measures net resource availability and the growth potential of the associated tree. Conversely, tree growth a is a measure of a tree’s ability to occupy a part of the growing space and associated site resources. The actual growth rate of the tree is determined by the total amount of resources captured and how efficiently those resources are used to produce biomass or stem volume (Asmann 1970). Knowledge of resource-use efficiency is a key factor for silvicultural manipulation of site resources and for assessing potential adaptation to climate change and associated changes in total resource availability, including annual and seasonal fluctuations. Growth efficiency has been assessed in many managed and unmanaged North American forest ecosystems (Waring et al 1981; Smith and Long 1989; Maguire et al. 1998; Mainwaring and Maguire 2004). However, analysis of growth efficiency in plantation ecosystems (Stoneman and Whitford 1995) and studies on forest growth efficiency in northern Europe (O’Hara et al. 2001) are less common. The relationship between forest growth efficiency and water availability (positive relationship) or temperature increase (positive relationship while water is available) has been documented for multiple ecosystems (Collalti et al. 2020). In our case, canopy variables act as a surrogate for light capture, while the thinning that impacts tree-size distribution in the stands also modifies site occupancy and transpiration.
Our results do not strongly support the Binkley hypothesis about growth efficiency increasing with tree size (reflected by relatively low GDC), which suggests that the dominance hypothesis may not be especially applicable to Mediterranean pine stands in central Spain. Similarly, Fernandez Tschieder et al. (2012) found that even with high stand density and competition, Pinus ponderosa tree growth in Patagonia (Argentina) was almost always proportional to tree size, implying no strong dominance pattern.
In the Mediterranean forests of southern Europe, stand-level growth of both Scots pine and Mediterranean pine increase with increasing crown surface area, but at a rate that indicate a decline in efficiency (growth per unit crown surface area) as crown surface area increase. As crown surface area increases in larger trees, the degree of self-shading typically increases if the larger crown size is not associated with a higher crown position (Riofrío et al. 2017). From a silvicultural perspective, growth per unit of occupied stand area is the most relevant type of growth efficiency and probably best estimated as area potentially available or as a multiple of crown projection area. Growth dominance and growth efficiency metrics are based on assumptions and simplifications (such as the allocation of trees into size classes) that may not capture the variability of tree growth patterns within stands (Fernandez Tschieder et al. 2012).
Limitations or considerations associated with our results (and those of other studies, such as Fernandez Tschieder et al. 2012; Cordonnier and Kunstler 2015; Navarro-Cerrillo et al. 2016; or Liang et al. 2023) include the following: (1) model simplifications such as additivity in the effects considered and variable transformations; (2) limitations associated with the data sample size and representation of temporal and spatial scale; (3) unique species-specific responses; (4) methodological capability for model selection and interpretation of the results due to complexity, the structure of the variance associated with the random factors, and the inclusion of numerous potentially intercorrelated variables; (5) potential confounding factors not considered, including the microsite mosaic and variation introduced by subtle differences in spatial distributions and related tree interactions; and (6) assumptions about the role of competition between trees (e.g. the relative importance of symmetric versus asymmetric competition) that influence the selection of competition variables. Taken together, these factors may not reliably reflect the complexity of interactions among trees in a stand, thereby limiting generalization and application of the results to other forestry contexts. Additionally, the inclusion of additional structure variables like Gini coefficient, already tested by Pretzsch and Hilmers (2024) and Pretzsch et al (2024), or the use of variable transformations, like adding quadratic terms in the structure variables, could improve our results.
To sum up, this empirical study indicates that: (1) stand growth declined with increasing size-class uniformity (increasing Shannon-like diversity index for crown size) in Scots pine stands and with increasing height in Mediterranean pine stands; (2) Tree size differentiation or initial inequality (expressed as dominance and reverse dominance) at the beginning of the growth period impacted stand growth; and (3) the Binkley hypothesis about growth efficiency increasing with tree size (as reflected by relatively low GDC) was not strongly supported, suggesting that the dominance hypothesis may not adequately describe Mediterranean pine stands in central Spain.
Our findings have three significant implications for Mediterranean pinewood silviculture: (1) thinning improves stand level growth, but the thinning intensity is more important than the method (systematic, from below, from above); (2) thinning, which increases water yield (Sun et al. 2015) while concentrating growth among a reduced number of trees, can generate multiple ecosystem services (water yield, biomass production, carbon sequestration) simultaneously; and (3) our results improve individual tree modelling by integrating dominance effects into the explanatory variables for growth. Such information can enhance the modelling and simulation of mixed and complex stands (Bravo et al. 2019) to facilitate locally adapted silvicultural designs that provide various ecosystem services.
This approach makes it possible to expand our understanding of individual tree interactions and the growth mechanisms that drive size distribution, which is especially interesting for mixed, structurally complex stands. Further research on such stands would increase our knowledge of how size inequalities impact tree growth.
References
Asmann E (1970) The principles of forest yield study. Pergamon Press, Oxford
Biging GS, Dobbertin M (1992) A comparison of distance-dependent competition measures for height and basal area growth of individual conifer trees. For. Sci. 38:695–720
Biging GS, Dobbertin M (1995) Evaluation of competition indices in individual tree growth models. For. Sci. 41:360–377
Binkley D (2004) A hypothesis about the interaction of tree dominance and stand production through stand development. For. Ecol. & Manage. 190:265–271
Binkley D (2023) Acorn review: The persistent mystery of declining growth in older forests. For Ecol Manage 538:121004. https://doi.org/10.1016/j.foreco.2023.121004
Binkley D, Stape JL, Ryan MG, Barnard HR, Fownes J (2002) Age-related decline in forest ecosystem growth: an individual-tree, stand-structure hypothesis. Ecosystems 5:58–67
Binkley D, Stape JL, Ryan MG (2004) Thinking about efficiency of resource use in forests. For Ecol Manage 193:5–16
Bond BJ, Czarnomski NM, Cooper C, Day ME, Greenwood MS (2007) Developmental decline in height growth in Douglas-fir. Tree Phys 27:441–453
Bravo F, Guerra B (2002) Forest structure and diameter growth in Maritime Pine in a Mediterranean Area. In: von Gadow K, Nagel J, Saborowski J (eds) Continuous cover forestry. Managing forest ecosystems, vol 4. Springer, Dordrecht. https://doi.org/10.1007/978-94-015-9886-6_10
Bravo F, Montero G (2001) Site index estimation in Scots pine (Pinus sylvestris L.) stands in the High Ebro Basin (northern Spain) using soil attributes. Forestry 74(4):395–406
Bravo F, Hann D, Maguire DA (2001) Impact competitor species composition on predicting diameter growth and survival rates of Douglas-fir trees in southwest Oregon. Can. J. For. Res. 31:2237–2247
Bravo F, Fabrika M, Ammer C, Barreiro S, Bielak K, Coll L, Fonseca T, Kangur A, Löf M, Merganičová K, Pach M, Pretzsch H, Stojanović D, Schuler L, Peric S, Rötzer T, del Río M, Dodan M, Bravo-Oviedo A (2019) Modelling approaches for mixed forests dynamics prognosis. Research gaps and opportunities. For Syst 28(1):eR002. https://doi.org/10.5424/fs/2019281-14342
Bravo-Oviedo A, Del Rio M, Montero G (2004) Site index curves and growth model for Mediterranean maritime pine (Pinus pinaster Ait.) in Spain. For Ecol Manag 201:187–197
Cochran PH, Geist JM, Clemens DL, Clausnitzer RR, Powell DC (1994) Suggesting stocking levels for forest stands in northeastern Oregon and southeastern Washington. Research Paper - US Department of Agriculture, Forest Service.
Collalti A, Ibrom A, Stockmarr A, Cescatti A, Alkama R, Fernández-Martínez M, Matteucci G, Sitch S, Friedlingstein P, Ciais P, Goll DS, Nabel JEMS, Pongratz J, Arneth A, Haverd V, Prentice IC (2020) Forest production efficiency increases with growth temperature. Nat Commun 11:5322. https://doi.org/10.1038/s41467-020-19187-w
Cordonnier Georges, Kunstler T (2015) The gini index brings asymmetric competition to light perspectives in plant. Ecol Evol Systemat 17(2):107–115. https://doi.org/10.1016/j.ppees.2015.01.001
Fernández ME, Gyenge J (2009) Testing Binkley’s hypothesis about the interaction of individual tree water use efficiency and growth efficiency with dominance patterns in open and close canopy stands. For Ecol Manag 257:1859–1865
Forrester D (2019) Linking forest growth with stand structure: Tree size inequality, tree growth or resource partitioning and the asymmetry of competition. For Ecol Manag. https://doi.org/10.1016/J.FORECO.2019.05.053
Fernández Tschieder E, Fernández ME, Schlichter TM, Pinazo MA, Crechi EH (2012) Influence of growth dominance and individual tree growth efficiency on Pinus taeda stand growth. A contribution to the debate about why stands productivity declines. Forest Ecol Manag 277:116–123. https://doi.org/10.1016/j.foreco.2012.04.025
Garber SM, Maguire DA (2005) The response of vertical foliage distribution to spacing and species composition in mixed conifer stands in central Oregon. For Ecol Manag 211:341–355
Hamilton GJ (1969) The Dependence of Volume Increment of Individual Trees on Dominance Crown Dimensions and Competition. Forestry 42(2) 133–144. https://doi.org/10.1093/forestry/42.2.133
Hann DW, Larsen DR (1991) Diameter growth equations for fourteen tree species in southwest Oregon. In: Forest Research Laboratory, Oregon State University, Corvallis
Hann DW, Hanus ML (2001) Enhaced mortality equations for trees in the mixed conifer zone of Southwest Oregon. Forest Research Lab. Oregon State University, Corvallis
Hann DW, Ritchie MW (1988) Height growth rate of Douglas-fir: a comparison of model forms. For. Sci. 34:165–175
Liang R, Xie Y, Sun Y, Wang B, Ding Z (2023) Temporal changes in size inequality and stand growth partitioning between tree sizes under various thinning intensities in subtropical Cunninghamia lanceolata plantations. For Ecol Manage 547. https://doi.org/10.1016/j.foreco.2023.121363
Lin S, Li Y, Chen Q, Wang Q, He K (2021) Influence of tree size local forest structure topography and soil resource availability on plantation growth in Qinghai Province. China Ecol. Indic. 120:106957. https://doi.org/10.1016/j.ecolind.2020.106957
Lizarralde I, Bravo F (2003) Crown and taper equations for Scots pine (Pinus sylvestris L.) in Northern Spain. In: Proceedings: decision support for multiple purpose forestry. Wien.
Long JN, Smith FW (1990) Determinants of stemwood production in Pinus contorta var. latifolia forests: the influence of site quality and stand structure. J Appl Ecol 27:847–856
Maguire DA, Bennett WS (1996) Pattern in vertical distribution of foliage in young coastal Douglas-fir. Can J For Res 26:1991–2005
Maguire DA, Batista JLF (1996) Sapwood taper models and implied sapwood volume and foliage profiles for coastal Douglas-fir. Can. J. For. Res. 26:849–863. https://doi.org/10.1139/x26-093
Maguire DA, Brissette JC, Gu L (1998) Crown structure and growth efficiency of red spruce in uneven-aged, mixed-species stands in Maine. Can J For Res 28:1233–1240
Mainwaring DB, Maguire DA (2004) The effect of local stand structure on growth and growth efficiency in heterogeneous stands of ponderosa pine and lodgepole pine in central Oregon. Can J For Res 34(11):2217–2229
Metsaranta J, Lieffers V (2007) Inequality of size and size increment in Pinus banksiana in relation to stand dynamics and annual growth rate. Ann Bot 101:561–571. https://doi.org/10.1093/aob/mcm320
Moore JA, Zhang L, Newberry JD (1994) Effects of intermediate silvicultural treatments on the distribution of within-stand growth. Can J For Res 24(2):398–404. https://doi.org/10.1139/x94-053
Moreno-Gutiérrez C, Barberá G, Nicolás E, Luis M, Castillo V, Martínez-Fernández F, Querejeta J (2011) Leaf δ18O of remaining trees is affected by thinning intensity in a semiarid pine forest. Plant Cell Environ 34(6):1009–1019. https://doi.org/10.1111/j.1365-3040.2011.02300.x
Navarro-Cerrillo RM, Sánchez-Salguero R, Herrera R, Ceacero Ruiz CJ, Moreno-Rojas JM, Manzanedo RD, López-Quintanilla J (2016) Contrasting growth and water use efficiency after thinning in mixed Abies pinsapo-Pinus pinaster-Pinus sylvestris forests. J For Sci 62(2):53–64. https://doi.org/10.17221/104/2015-JFS
Nord-Larsen T, Damgaard C, Weiner J (2006) Quantifying size-asymmetric growth among individual beech trees. Can. J. For. Res. 36:418–425
O’Hara KL (1988) Stand structure and growing space following thinning in an even-aged Douglas-fir stand. Can J For Res 18:859–866
O’Hara KL, Lähde E, Laiho O, Norokorpi Y, Saksa T (2001) Leaf area allocation as guide to stocking control in multi-aged, mixed-conifer forests in southern. Finland For 74(2):171–185
O’Hara KL (1996) Dynamics and stocking-level relationships of multi-aged ponderosa pine stands. For Sci Monograph 33
Pretzsch H, Schütze G (2015) Effect of tree species mixing on the size structure, density, and yield of forest stands. Eur J For Res 135:1–22. https://doi.org/10.1007/s10342-015-0913-z
Pretzsch H, Hilmers T, del Río M (2024) The effect of structural diversity on the self-thinning line, yield level, and density-growth relationship in even-aged stands of Norway spruce. For Ecol Manag 556:121736
Pretzsch H, Hilmers T (2024) Structural diversity and carbon stock of forest stands. Tradeoff as modified by silvicultural thinning. Eur J For Res (submitted)
Riofrío J, Del Río M, Bravo F (2017) Mixing effects on growth efficiency in mixed pine forests. Forestry 90:381–392
Rojo A, Montero G (1996) El pino silvestre en la Sierra de Guadarrama, Madrid
Ruano I, Rodríguez-García E, Bravo F (2013) Effects of pre-commercial thinning on growth and reproduction in post-fire regeneration of Pinus halepensis Mill. Ann For Sci 70:357–366. https://doi.org/10.1007/s13595-013-0271-2
Ryan MG, Yoder BJ (1997) Hydraulic limits to tree height and tree growth. BioScience 47:235–242
Ryan MG, Binkley D, Fownes JH (1997) Age-related decline in forest productivity: pattern and process. Adv Ecol Res 27:213–262
Ryan MG, Binkley D, Fownes JH, Giardina CP, Senock RS (2004) An experimental test of the causes of forest growth decline with stand age. Ecol Monogr 74:393–414
Smith FW, Long JN (1989) The influence of canopy architecture on stemwood production and growth efficiency of Pinus contorta var. latifolia. J Appl Ecol 26:681–691
Sterba H, Amateis RL (1998) Crown efficiency in a loblolly pine (Pinus taeda) spacing experiment. Can J For Res. 28:1344–1351
Stoneman GL, Whitford K (1995) Analysis of the concept of growth efficiency in Eucalyptus marginata (jarrah) in relation to thinning, fertilising and tree characteristics. For Ecol Manag 76:47–53
Sun G, Caldwell P, McNulty S (2015) Modelling the potential role of forest thinning in maintaining water supplies under a changing climate across the conterminous United States. Hydrol Process 29:5016–5030. https://doi.org/10.1002/hyp.10469
Sun H, Diao S, Liu R, Forrester D, Soares Á, Saito D, Dong R, Jiang J (2018) Relationship between size inequality and stand productivity is modified by self-thinning, age, site and planting density in Sassafras tzumu plantations in central China. For Ecol Manag. https://doi.org/10.1016/J.FORECO.2018.02.003
Wang YP, Jarvis PG (1990) Influence of crown structural properties on PAR absorption, photosynthesis, and transpiration in Sitka spruce: application of a model (MAESTRO). Tree Phys 7:297–316
Waring RH, Thies WG, Muscato D (1980) Stem growth per unit of leaf area: a measure of tree vigor. For Sci 36:1077–1104
Waring RH, Newman K, Bell J (1981) Efficiency of tree crowns and stemwood production of different canopy leaf densities. Forestry 54:129–137
Wensel LC, Meerschaert WJ, Biging GS (1987) Tree height and diameter growth models for northern California conifers. Hilgardia 55:1–20
West PW (2014) Calculation of a growth dominance statistic for forest stands. For Sci 60(6):1021–1023. https://doi.org/10.5849/forsci.13-186
Wykoff WR (1990) A basal area increment model for individual conifers in the northern Rocky Mountains. For. Sci. 36:1077–1104
Xu M, Harrington TB (1998) Foliage biomass distribution of loblolly pine as affected by tree dominance, crown size, and stand characteristics. Can J For Res 28:887–889
Acknowledgements
To the two external reviewers and editor who provided insightful comments. This research was funded by the following grants: ‘CLU-2019-01-iuFOR Institute Unit of Excellence’ of the University of Valladolid, funded by the Junta de Castilla and co-financed by the European Union (ERDF “Europe drives our growth”); the IMFLEX Grant PID2021-126275OB-C22 funded by MCIN/AEI/https://doi.org/10.13039/501100011033; and “ERDF A way of making Europe,” financed by the Spanish Ministry for Science, Innovation and Universities and the European Union.
Funding
Open Access funding provided thanks to the CRUE-CSIC agreement with Springer Nature.
Author information
Authors and Affiliations
Contributions
D.A.M. and F.B. and C.O. wrote the main manuscript text D.A.M. and F.B. and C.O. prepared and designed the study D.A.M. prepared data calculus and desing of variables of interest. V.P. guided statistical analysis, wrote statistical methods and prepared SAS scripts. C.O. prepare data, tables and figures. All authors reviewed the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Communicated by Hans Pretzsch.
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Ordóñez, C., Maguire, D.A., Pando, V. et al. Stand structural effects on growth distribution and growth efficiency in Scots pine and Mediterranean pine in Spain. Eur J Forest Res (2024). https://doi.org/10.1007/s10342-024-01698-6
Received:
Revised:
Accepted:
Published:
DOI: https://doi.org/10.1007/s10342-024-01698-6