
Abstract
The character of pure or mixed forest canopies and their litterfalls contribute to different forest-floor properties. These organic layers and also subjacent topsoil were studied at three study sites covered by mixed treatments such as beech–spruce, beech–fir, spruce–fir and two monospecific beech and spruce treatments. The age of the forest stands ranged from 11 to 15 years when sampled. All study sites were used as meadows when afforested; therefore, the forest floors were new, and the A-horizon topsoil properties were not influenced by older humus inherited from previous forest generations. The mineral soil was likely affected by different levels of former fertilization, which resulted in differences among the study sites. The early-developed forest floors showed differences between the treatments with beech and the others. The topsoil below beech with spruce had more nitrogen, oxidizable carbon and cations of exchangeable hydrogen as well as pH showing more acidic conditions and lower contents and saturation of base cations. Pure beech had more phosphorus. The nutrient pools did not differ among the treatments; significantly more matter was found below the oldest stands on the first afforested site, which also increased nutrient pools.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Forest soils are covered by accumulated organic layers that modify the microclimate, physical, chemical and biological properties of the soil (Binkley and Giardina 1998). These organic substances, which mainly come from litterfall, help develop forest floors. The decomposition of such layer releases nutrients bound in tissues (see Attiwill and Adams 1993). The thicknesses of forest floors differ according to the break-down conditions (Binkley and Giardina 1998). Soil below a newly established forest stand that has not been affected by a previous generation of forest shows a new forest-floor development. This, along with the presence of woody species, is the most apparent sign of a forest environment restoration. Replicated common-garden experiments have mostly been established under such conditions (Binkley 1995), because such former agricultural lands provide suitable conditions for investigating soil–plant relationships. For example, both significant and insignificant differences below beech and spruce were found in the forest floor and topsoil of former arable soil (Hagen-Thorn et al. 2004).
Turning away from growing monospecific stands is needed (Berger et al. 2010). However, this raises the issue of mixed-stand impacts on soil (see Dhiedt et al. 2021). Either tree species or their mixtures thrive differently, contributing to specific microclimate conditions, a mixture of litterfall and the presence of root biomass, which modifies the consumption of nutrients. As for beech and spruce, Finér et al. (2007) found greater fine-root biomass with beech than that of spruce. When mixed, spruce root biomass did not differ from the pure spruce stand, whereas the presence of beech in mixtures showed a significantly higher specific root length and specific surface area of fine roots compared to pure beech stands (Bolte and Villanueva 2006). In the Western Carpathians, Zielonka et al. (2021) found the highest levels of fine-root biomass below silver fir compared to Norway spruce and European beech. Despite the fact that beech stores less carbon in soil compared to maple and linden (Cukor et al. 2022), it is one of the more common species used for soil ameliorating in the Czech Republic. Spruce is a main Central European commercial species, whereas the most important conifer of the past—fir is very limited. These three trees accompany each other in different mixtures along a gradient from foothills to Alpine sites in Europe (Hilmers et al. 2020; Filipiak et al. 2021; Zielonka et al. 2021). As European foresters are facing a decline of monocultures, the issue of mixed-species forest restoration has been raised. In addition, more information on the interaction of such forests with soil, including the effects of the mixtures on soil, is needed.
The objective of this study was to compare the early development of forest floors in first-generation forest stand mixtures following afforestation; mixtures with spruce, beech and fir were compared with monospecific spruce and beech treatments at replicated experimental sites.
Materials and methods
Replicated row mixtures of beech with spruce and fir (Be_Sp; Be_Fi), spruce with fir (Sp_Fi) and also monospecific patches of beech (Be) and spruce (Sp) were established at the three formerly agricultural sites, Bystré—BY, Uhřínov—UH and Osečnice—OS (Table 1). The areas of each planted squared patch in BY, UH and OS were 0.022, 0.04 and 0.04 hectares, respectively. BY and OS were arable land until 1960, whereas tillage at parts of UH ceased a decade earlier. All study sites were meadows prior to planting. The rows of plants were spaced 1.6 m from each other, and their length was 15 m (BY) and 20 m (UH, OS). In mixed treatments, the patches were represented by five spruce rows and four rows of admixed species in BY and by 6 rows per species in the two other sites. The development of the stand treatment densities and mensuration characteristics are presented in Table 2. Beech had a lower survival rate at the UH site compared to the BY and OS sites.
In the sampling years, spruce was the tallest and thickest species and was accompanied by both beech and fir at all study sites. Regardless of the age of all plots, beech with spruce showed the largest basal area, whereas pure beech showed the least basal area (Table 2). The basal areas of Sp patches in every plot were similar.
In the autumn of 2019, forest floor (L, F and H layers altogether) was sampled using a 625-cm2 iron frame in order to recalculate the dry matter enclosed within the frame to values per hectare. Topsoil was only partially taken from within the frame. Each treatment (Table 2) was sampled five times randomly inside the patches (excluding the inner boundary zones).
The forest-floor samples were dried and weighed (dry matter—DM). The DM was then sieved through a 2-mm mesh in order to separate coarse debris, thus allowing us to obtain its fine fraction (DM_Fine), which was then weighed again and analysed. Both the new organic layer and topsoil were analysed to get information on the contents of organic carbon (Cox), combustible matter (Comb_sum), Kjeldahl nitrogen (N), pH in water and KCl, Mehlich III plant-available nutrients (P, K, Ca and Mg) and base cation content (BCC), base saturation (BS), cation exchange capacity (CEC) and hydrogen cations (H = CEC–BCC) according to Kappen (1929).
Statistical analysis
The basic stand data are presented as average values per individual species and by stand treatment characteristics. Analyses were performed in the R statistical computing environment (4.0.3, R Core Team 2020). The principal component analysis (PCA) of the soil parameters of each horizon was computed using the FactoMineR package (Lê et al. 2008), and this served as the basal view of the data. The data on qualitative and quantitative (nutrient pools in the forest floor) soil properties were evaluated separately. The evaluated parameters were taken as input variables, and species treatment and the experimental plot were set as a factor. The results were visualized using the ggbiplot function (Vu® 2011).
The nutrient pools of every sample in the forest floor were computed using fine matter dry weights (DM_Fine). The nutrients in the fine matter fraction were considered to be released shortly. The properties of each soil layer were tested for differences among the treatments. The data for each variable were tested using the Shapiro–Wilk test for normality and by Levene’s test for homogeneity of variance across groups. In the forest-floor data, one strong outlier in dry weight, two in Cox and one in BCC were excluded from the analysis. Subsequently, ANOVA with a randomized block design was used:
where Yij is the response in treatment i for block j, µ is the mean for block j, αi represents the fixed treatment effects of the present species, β is the block effects of the experimental plot and εik is the normally distributed random errors. To satisfy the ANOVA assumptions when data were not normally distributed (more often in the A horizon) or when it was necessary (rarely) to stabilize variances, the Box–Cox transformation was used (Fox and Weisberg 2011). Orthogonal contrasts were defined for both effects. A linear model was computed using the lm function (statistics package), and type III ANOVA table outputs were evaluated. For subsequent post hoc mean separation, Tukey’s honestly significant difference test was used (statistics package). The normality of model residua was checked. The analysed differences were considered to be significant when p ≤ 0.05. Least-square means (LS) and standard errors (SEs) are presented. Pearson's product–moment correlation revealed relations between individual qualitative properties in humus and the A horizon.
Results
Forest floor
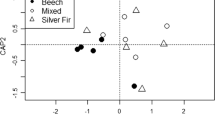
The first two axes of the PCA ordination diagram explained almost 75% of variability. The key trends were lower pH and base saturation below beech (Be) and lower nutrient contents in both coniferous variants (Sp, Sp_Fi); Sp showed the greatest variability of data (Fig. 1). General differences in the properties of forest floor were observed between the experimental plots; for example, BY samples tended to have lower pH, BS and partly also BCC and Ca concentrations, especially when compared to UH soil (Fig. 1).

An ordination diagram (PCA) of the analysed qualitative chemical properties of the forest floor according to the treatments (above) and the experimental plots (below); percentages express the variance explained by the two axes
Significant differences among the treatments were shown in all properties analysed with the exception of BCC, CEC and Ca (Table 3). The pure beech (Be) and also two other mixtures with beech (Be_Fi or Be_Sp) mostly differed from the coniferous treatments. DM_Fine below Be showed lower pH and base saturation (Table 4). The presence of beech increased combustible matter and Cox, K and Mg contents (Table 5). Similar to the PCA outputs, some differences among the three experimental sites were found. BY mostly differed in lower pH, BS, K and Mg contents, whereas it had a greater amount of hydrogen cations. The lowest phosphorus was found in the OS forest floor (Tables 4 and 5).
No differences in the DM_Fine nutrient pools of all treatments were found (Table 6). The oldest site, BY, showed more forest-floor biomass, which was also reflected in significantly more nutrients in the humus except for K and Mg (Table 7).
Topsoil
The first two PCA axes explained more than 80% of the data variability in the topsoil (Fig. 2). Be_Sp differed from the other treatments as it was higher in nitrogen, oxidizable carbon (Cox), the content of combustible matter (Comb_sum), the content of exchangeable hydrogen ions (H), and it also showed lower pH. As for the experimental sites, the conditions at the UH site were the most favourable, whereas the BY site exhibited the worst conditions, which generally matched the forest-floor analysis (Fig. 1).

An ordination diagram (PCA) of the analysed qualitative chemical properties in the A horizon according to the treatments (above) and the experimental plots (below). Percentages express the variance explained by the two axes
ANOVA confirmed the significant differences among all properties with the exception of K among the stand treatments and Cox among the study sites (Table 3). The trends were similar to the PCA findings; the lowest pH was found below Be_Sp, while the highest pH was found below the treatments with fir. The Be treatment showed the lowest BCC, CEC and BS, while the highest levels (only BS significantly) were found below the treatments with fir (Table 4). The poorest nutrient contents (except for P and N) were found below Be_Sp, which, however, showed the greatest amount of combustible matter (Table 5). The treatments with fir had more Ca and Mg; Be topsoil was the highest in P.
The acidity of the mineral topsoil along with BCC, CEC and BS decreased in the following order: UH > OS > BY (Table 4). This trend was also confirmed for Ca and Mg. Similar to the PCA results, the UH topsoil showed the most fertile conditions except for N (Table 5).
The whole dataset analysis confirmed highly significant correlations between forest floor and topsoil properties (for BCC p = 0.001, for CEC p = 0.004 and for others p < 0.001), with the exception of N and Ca, which showed independent patterns in their contents in both layers (for N p = 0.5, for Ca p = 0.2).
Discussion
Land-use history
Former agricultural soil reflects past practices. The impact of agricultural land-use history on soil properties results from operations such as ploughing, mowing, pasture and fertilization, and their legacy remains detectable in soil for a long time (e.g. Szujecki 1996; Koerner et al. 1997; Verheyen et al. 1999; Richter et al. 2000; Dupouey et al. 2002; Ritter et al. 2003; Wall and Hytönen 2005; Wall and Westman 2006). The legacy of fertilization was reported, for example, by Kacalek et al. (2011) for the same region in which the BY, OS and UH sites are situated. Despite similar ecosite classification, the soils in our study sites showed different chemical properties that cannot yet be attributed to plantation litterfall. Most of the analysed soil properties showed the following order UH > OS > BY, which likely reflects the fertility inherited from the previous land use. According to historical aerial orthophotographs, the BY and OS sites were used as arable land that had been tilled to at least the 1960s (CUZK 2022). The UH site is situated partly on a steeper slope (14°), which limited its historical use to meadow; arable use (which ceased in the 1950s) was likely applied on the plain land above and the small terrace below the slope. However, according to witnesses, infrequent manuring of the meadow above the afforested area was applied as late as in the 1980s. Manuring most likely enriched the study-site soil via surface and subsurface water flow, and both meadow plant vegetation and tree litterfall helped to recycle the nutrients (e.g. Stieglitz et al. 2003). This is a likely reason for the present higher fertility of UH compared to both other sites, as the BS values of UH > OS > BY show 88, 70 and 55%, respectively (see Table 5). The same site order was found more than a decade earlier (sampled in 2006) when the UH > OS > BY BS values were 87% > 57% > 45%, respectively (unpublished data). The most fertile UH site did not change too much over the two sampling campaigns; both the poorer OS and the poorest BY sites were higher in base nutrients compared to the samples from 2006.
Impacts of tree species
The effects of tree species can be expected mainly in a forest floor (Ritter et al. 2003) and/or both organic layers and mineral topsoil (Vesterdal et al. 2008; Cremer et al. 2016). A new forest floor is the main C sink over the three decades following afforestation under temperate conditions (Mayer et al. 2020). The positive impact of afforestation on the physical properties of topsoil and upper subsoil has also been reported (e.g. by Vopravil et al. 2021). Below-canopy nutrient inputs are attributable to the species-specific nature of tree species canopies, as beech-dominated, spruce-dominated and mixed canopies show different patterns of litterfall and throughfall nutrient inputs (Hojjati et al. 2009). These authors reported a lower Mg input via litterfall below spruce canopy compared to beech. Our monospecific spruce and mixtures with spruce also showed a forest floor lower in Mg compared to monospecific beech. Soil differences between spruce and beech have been compared many times. Binkley (1995) pointed out that spruce was considered a site-deteriorating species in Europe, whereas beech was considered either a “mother of the forest” or a producer of the worst-type humus. The worst characteristics attributable to spruce—topsoil acidification (Augusto et al. 2003; Hagen-Thorn et al. 2004; Podrázský and Remeš 2008)—were not observed in our study. Among the other negative characteristics of the impact of spruce on soil, various effects such as a retardation of organic matter decomposition (Albers et al. 2004), a loss of leached base cations due to shallow rooting (Berger et al. 2006), foliage lower in nutrients (Berger et al. 2009), a deeper mineral (B) soil horizon higher in SO42− in formerly air-polluted mountains (Tejnecký et al. 2013) and a lower soil sorption capacity (Oulehle et al. 2016) have been reported. As compared to beech treatments, the respective negative impact of spruce was not significantly supported in our study. However, spruce might not have only soil-deteriorating effects as, for example, more porous soil was found below spruce compared to other trees such as European beech, English oak, sycamore maple and small-leaved lime (Cukor et al. 2022). The inner-rhizosphere topsoil was also enriched with base cations such as K, Ca and Mg below both spruce and beech (Collignon et al. 2011). Silver fir’s ability to use nutrients is not well known (Dušek et al. 2020), but according to Třeštík and Podrázský (2017), the soil-improving effects of silver fir do not differ from that of Norway spruce. Some shifts were, however, found in the carbon contents of soluble compounds in slow-evolved fir and fast-turnover beech organic matter (Pizzeghello et al. 2006).
The pure beech treatment that showed lower pH levels, base saturation and more exchangeable H+ compared to pure spruce in our study. This is not an extraordinary result for undecomposed plant matter because the shallowest part of the beech forest floor was found more acidic than that under spruce (Trum et al. 2011). Also, when deeper layers of organic origin are developed, beech has less acidic or the same conditions as spruce (Trum et al. 2011). Sometimes differences of microbial C, N and P contents were found at higher levels in forest floors under beech than under spruce (Zederer et al. 2017). However, when recalculated to pools, mean forest-floor stocks of microbial P and total P did not differ between these two tree species, due to the increased organic matter accumulation in the forest floor under spruce (Zederer et al. 2017). This is what we also observed in our study, where total dry matter, fine dry matter (sieved through a 2-mm mesh) and nutrient stocks did not differ among the treatments; the dry matter of all treatments was significantly higher only at the oldest BY site. Hansen et al. (2009) also previously found similar total litterfall among tree species such as beech, oak, Douglas fir, Norway spruce and Sitka spruce, but site factors affected the litterfall significantly. Hou et al. (2020) concluded, similarly to our findings, that soil organic carbon sequestration rates in both deciduous broadleaf and evergreen conifer forests were comparable.
Mixture-specific impacts
Hojjati et al. (2009) reported a lower Mg input via litterfall below a spruce canopy compared to a spruce–beech canopy. In our study, the forest floor with spruce needles was also lower in Mg compared to the beech with fir mixture. Mixed stands with beech (especially Be_Sp) had higher tree densities and larger G. The mixed beech–spruce litters showed the greatest richness of fungi and also similar or higher mass loss compared to pure litter types (Kubartová et al. 2009). Beech–conifer (Douglas fir and Norway spruce) mixtures were also reported to have higher carbon stocks in the forest floor compared to pure beech; the mixture forest floor C was lower or similar to pure conifer stands (Cremer et al. 2016). However, our early-developed forest floor showed no differences among the treatments. The beech–conifer mixtures were found capable of increasing soil pH and base saturation; both mixtures maintained soil fertility better than pure conifer stands (Cremer and Prietzel 2017). As for the basic cation stocks, Cremer and Prietzel (2017) found intermediate stocks of Ca and Mg below the mixture of beech and spruce, whereas pure beech had more Ca and Mg compared to pure conifers. The early impacts following the afforestation of the BY, OS and UH sites showed no difference among the stocks of these nutrients; concentrations of Ca did not differ; and Mg concentrations showed the following order (Be; Be_Fi) > (Be_Sp; Sp; Sp_Fi). Spruce seemed to be the main species driving the difference under the analysed conditions.
The conversion of a spruce monoculture into a mixture of spruce and beech was found to increase the invertebrate decomposer’s potential attributable to the quality of the beech litter (Elmer et al. 2004). In addition, beech leaves were more favourable to microorganisms than spruce needles, and the decomposition rates showed the following order: beech > mix > spruce (Albers et al. 2004). On the other hand, Andivia et al. (2016) found that the carbon stock in the forest floor reflected a proportion of spruce needles in litterfall. This relation did not yet apply to our young stands. Studies in mature forest stands have reported higher litter amounts accumulated below spruce. According to Berger and Berger (2014), these higher amounts are not attributable to the inherent recalcitrance of needles. Forest-floor nitrogen in our study was comparable among pure beech and both coniferous treatments, which differed significantly from both mixtures of beech with evergreen conifers. As for the oxidizable carbon proportion/share, only monospecific beech differed from the mixture of spruce and fir significantly, while the pools were not different. Rehschuh et al. (2021) concluded that adding conifers to beech increases carbon accumulation in soils; it, however, did not apply to our early stage of forest cover restoration on former agricultural sites, and further changes can be expected both as the stands use nutrients to grow and also as litterfall is returned onto the soil.
Conclusions
Analyses of the impact of a new forest canopy on former agricultural land showed that even young (at an age of 12–18 years) tree species and their mixtures are capable of distinctively impacting topsoil chemical properties. The pattern of changes, however, differs from the published findings from studies conducted in older forests. Only qualitative properties manifested significant differences among the treatments in our study; dry matter amounts and also nutrient pools were similar. Particularly, it can be concluded that:
-
Monospecific beech and mixtures with beech showed the lower pH levels and base saturation compared to coniferous treatments.
-
Dry mass and fine dry mass of the forest floor did not differ among the treatments and nutrient pools were also comparable.
-
The nutrient contents in the forest floor were not entirely reflected in the nutrients of the topsoil. The contents of N and Ca showed fully independent patterns in both layers.
-
Mineral topsoil below mixed beech with spruce (Be_Sp) was higher in nitrogen, oxidizable carbon, the content of combustible matter and the content of exchangeable hydrogen ions, and showed also lower pH levels and base saturation.
-
In spite of a similar ecosite classification, differences in the forest floor were observed between the analysed study sites. The land-use history seems to have long-term impact on the nutrient cycling of a new forest.
During the study period, every analysed treatment formed a forest-floor layer. Soil nutrient consumption by roots and nutrient return through litterfall and its decomposition also changed the properties of the topsoil. The diverse patches of monospecific and mixed stands are expected to affect forest soil properties related to the specific nutrient cycling, thus also supporting forest biodiversity.
References
Albers D, Migge S, Schaefer M, Scheu S (2004) Decomposition of beech leaves (Fagus sylvatica) and spruce needles (Picea abies) in pure and mixed stands of beech and spruce. Soil Biol Biochem 36:15–164. https://doi.org/10.1016/j.soilbio.2003.09.002
Andivia E, Rolo V, Jonard M, Formánek P, Ponette Q (2016) Tree species identity mediates mechanisms of top soil carbon sequestration in a Norway spruce and European beech mixed forest. Ann for Sci 73:437–447. https://doi.org/10.1007/s13595-015-0536-z
Attiwill PM, Adams MA (1993) Tansley review No. 50 nutrient cycling in forests. New Phytol 124:561–582
Augusto L, Dupouey J-L, Ranger J (2003) Effects of tree species on understory vegetation and environmental conditions in temperate forests. Ann for Sci 60:823–831. https://doi.org/10.1051/forest:2003077
Berger TW, Berger P (2014) Does mixing of beech (Fagus sylvatica) and spruce (Picea abies) litter hasten decomposition? Plant Soil 377:217–234. https://doi.org/10.1007/s11104-013-2001-9
Berger TW, Swoboda S, Prohaska T, Glatzel G (2006) The role of calcium uptake from deep soils for spruce (Picea abies) and beech (Fagus sylvatica). For Ecol Manag 229:234–246. https://doi.org/10.1016/j.foreco.2006.04.004
Berger TW, Untersteiner H, Toplitzer M, Neubauer C (2009) Nutrient fluxes in pure and mixed stands of spruce (Picea abies) and beech (Fagus sylvatica). Plant Soil 322:317–342. https://doi.org/10.1007/s11104-009-9918-z
Berger TW, Inselsbacher E, Zechmeister-Boltenstern S (2010) Carbon dioxide emissions of soils under pure and mixed stands of beech and spruce, affected by decomposing foliage litter mixtures. Soil Biol Biochem 42:986–997. https://doi.org/10.1016/j.soilbio.2010.02.020
Binkley D (1995) The influence of tree species on forest soils: processes and patterns. In: Mead DJ, Cornfort IS (eds) Proceedings of the trees and soil workshop. Lincoln University 28 February–2 March 1994. Agronomy Society of New Zealand Special Publication No. 10., Lincoln University Press, Canterbury, pp 1–33
Binkley D, Giardina C (1998) Why do tree species affect soils? The warp and woof of tree–soil interactions. Biogeochem 42:89–106. https://doi.org/10.1023/A:1005948126251
Bolte A, Villanueva I (2006) Interspecific competition impacts on the morphology and distribution of fine roots in European beech (Fagus sylvatica L.) and Norway spruce (Picea abies (L.) Karst.). Eur J for Res 125:15–26. https://doi.org/10.1007/s10342-005-0075-5
CGS (2019) Geoscience maps [online]. https://mapy.geology.cz/geo/. Accessed 14 June 2022
Collignon C, Calvaruso C, Turpault M-P (2011) Temporal dynamics of exchangeable K, Ca and Mg in acidic bulk soil and rhizosphere under Norway spruce (Picea abies Karst.) and beech (Fagus sylvatica L.) stands. Plant Soil 349:355–366. https://doi.org/10.1007/s11104-011-0881-0
Cremer M, Prietzel J (2017) Soil acidity and exchangeable base cation stocks under pure and mixed stands of European beech, Douglas fir and Norway spruce. Plant Soil 415:393–405. https://doi.org/10.1007/s11104-017-3177-1
Cremer M, Kern NV, Prietzel J (2016) Soil organic carbon and nitrogen stocks under pure and mixed stands of European beech, Douglas fir and Norway spruce. For Ecol Manag 367:30–40. https://doi.org/10.1016/j.foreco.2016.02.020
Cukor J, Vacek Z, Vacek S, Linda R, Podrázský V (2022) Biomass productivity, forest stability, carbon balance, and soil transformation of agricultural land afforestation: A case study of suitability of native tree species in the submontane zone in Czechia. CATENA 210:105893. https://doi.org/10.1016/j.catena.2021.105893
CUZK (2022) Land Survey Office [online maps]. https://ags.cuzk.cz/archiv/. Accessed 14 June 2022
Dhiedt E, Verheyen K, De Smedt P, Ponette Q, Baeten L (2021) Early tree diversity and composition effects on topsoil chemistry in young forest plantations depend on site context. Ecosystems 24:1638–1653. https://doi.org/10.1007/s10021-021-00605-z
Dupouey JL, Dambrine E, Laffite JD, Moares C (2002) Irreversible impact of past land use on forest soils and biodiversity. Ecology 83:2978–2984. https://doi.org/10.2307/3071833
Dušek D, Kacálek D, Novák J, Slodičák M (2020) Foliar nutrients in current and one-year-old needles of naturally-regenerated Norway spruce and silver fir saplings. Rep for Res - Zpr Lesnického Výzkumu 65:146–152 ([in Czech with English abstract and Summary])
Elmer M, La France M, Förster G, Roth M (2004) Changes in the decomposer community when converting spruce monocultures to mixed spruce/beech stands. Plant Soil 264:97–109. https://doi.org/10.1023/B:PLSO.0000047776.86805.0f
Filipiak M, Gubański J, Jaworek-Jakubska J, Napierała-Filipiak A (2021) The strong position of silver fir (Abies alba Mill.) in fertile variants of beech and oak-hornbeam forests in the light of studies conducted in the Sudetes. Forests 12:1203. https://doi.org/10.3390/f12091203
Finér L, Helmisaari H-S, Lõhmus K, Majdi H, Brunner I, Børja I, Eldhuset T, Godbold D, Grebenc T, Konôpka B, Kraigher H, Möttönen M-R, Ohashi M, Oleksyn J, Ostonen I, Uri V, Vanguelova E (2007) Variation in fine root biomass of three European tree species: Beech (Fagus sylvatica L.), Norway spruce (Picea abies L. Karst.), and Scots pine (Pinus sylvestris L.). Plant Biosyst 141:394–405. https://doi.org/10.1080/11263500701625897
Fox J, Weisberg S (2011) An R companion to applied regression. Sage Publications, Los Angeles
Hagen-Thorn A, Callesen I, Armolaitis K, Nihlgård B (2004) The impact of six European tree species on the chemistry of mineral topsoil in forest plantations on former agricultural land. For Ecol Manag 195:373–384. https://doi.org/10.1016/j.foreco.2004.02.036
Hansen K, Vesterdal L, Schmidt IK, Gundersen P, Sevel L, Bastrup-Birk A, Pedersen LB, Bille-Hansen J (2009) Litterfall and nutrient return in five tree species in a common garden experiment. For Ecol Manag 257:2133–2144. https://doi.org/10.1016/j.foreco.2009.02.021
Hilmers T, Biber P, Knoke T, Pretzsch H (2020) Assessing transformation scenarios from pure Norway spruce to mixed uneven-aged forests in mountain areas. Eur J for Res 139:567–584. https://doi.org/10.1007/s10342-020-01270-y
Hojjati SM, Hagen-Thorn A, Lamersdorf NP (2009) Canopy composition as a measure to identify patterns of nutrient input in a mixed European beech and Norway spruce forest in central Europe. Eur J for Res 128:13–25. https://doi.org/10.1007/s10342-008-0235-5
Hou G, Delang CO, Lu X, Gao L (2020) Grouping tree species to estimate afforestation-driven soil organic carbon sequestration. Plant Soil 455:507–518. https://doi.org/10.1007/s11104-020-04685-z
Kacalek D, Dusek D, Novak J, Slodicak M, Bartos J, Cernohous V, Balcar V (2011) Former agriculture impacts on properties of Norway spruce forest floor and soil. For Syst 20:437–443. https://doi.org/10.5424/fs/20112003-11042
Kappen H (1929) Die Bodenazidität. Springer, Berlin
Koerner W, Dupouey JL, Dambrine E, Benoît M (1997) Influence of past land use on the vegetation and soils of present day forest in the Vosges mountains, France. J Ecol 85:351–358
Kubartová A, Ranger J, Berthelin J, Beguiristain T (2009) Diversity and decomposing ability of saprophytic fungi from temperate forest litter. Microb Ecol 58:98–107. https://doi.org/10.1007/s00248-008-9458-8
Lê S, Josse J, Husson F (2008) FactoMineR: an R package for multivariate analysis. J Stat Softw 25:1–18. https://doi.org/10.18637/jss.v025.i01
Mayer M, Prescott CE, Abaker WEA, Augusto L, Cécillon L, Ferreira GWD, James J, Jandl R, Katzensteiner K, Laclau J-P, Laganière J, Nouvellon Y, Paré D, Stanturf JA, Vanguelova EI, Vesterdal L (2020) Tamm review: influence of forest management activities on soil organic carbon stocks: a knowledge synthesis. For Ecol Manag 466:118127. https://doi.org/10.1016/j.foreco.2020.118127
Opletal M, Domečka K (ed) (1983) Synoptic geological map of the Orlické hory Mts. Scale 1:100 000. Ústřední ústav geologický, Praha
Oulehle F, Růžek M, Tahovská K, Bárta J, Myška O (2016) Carbon and nitrogen pools and fluxes in adjacent mature Norway spruce and European beech forests. Forests 7:282. https://doi.org/10.3390/f7110282
Pizzeghello D, Zanella A, Carletti P, Nardi S (2006) Chemical and biological characterization of dissolved organic matter from silver fir and beech forest soils. Chemosphere 65:190–200. https://doi.org/10.1016/j.chemosphere.2006.03.001
Podrázský V, Remeš J (2008) Rapidity of forest soil character restoration of afforested localities of the Orlické hory Mts. Rep for Res - Zpr Lesnického Výzkumu 53:89–93 ([in Czech with English abstract and Summary])
R Core Team (2020) R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna
Rehschuh S, Jonard M, Wiesmeier M, Rennenberg H, Dannenmann M (2021) Impact of European beech forest diversification on soil organic carbon and total nitrogen stocks—a meta-analysis. Front for Glob Chang 4:606669. https://doi.org/10.3389/ffgc.2021.606669
Richter DD, Markewitz D, Heine PR, Jin V, Raikes J, Tian K, Wells CG (2000) Legacies of agriculture and forest regrowth in the nitrogen of old-field soils. For Ecol Manag 138:233–248. https://doi.org/10.1016/S0378-1127(00)00399-6
Ritter E, Vesterdal L, Gundersen P (2003) Changes in soil properties after afforestation of former intensively managed soils with oak and Norway spruce. Plant Soil 249:319–330. https://doi.org/10.1023/A:1022808410732¨
Stieglitz M, Shaman J, McNamara J, Engel V, Shanley J, Kling GW (2003) An approach to understanding hydrologic connectivity on the hillslope and the implications for nutrient transport. Glob Biogeochem Cycles 17(4):1105. https://doi.org/10.1029/2003GB002041
Szujecki A (1996) Ekologiczne aspekty odtwarzania lasu na glebach porolnych. Prace IBL, Ser B 27:47–55 ([in Polish])
Tejnecký V, Bradová M, Borůvka L, Němeček K, Šebek O, Nikodem A, Zenáhlíková J, Rejzek J, Drábek O (2013) Profile distribution and temporal changes of sulphate and nitrate contents and related soil properties under beech and spruce forests. Sci Total Environ 442:165–171. https://doi.org/10.1016/j.scitotenv.2012.10.053
Třeštík M, Podrázský V (2017) Soil improving role of the silver fir (Abies alba Mill.): a case study. Rep for Res - Zpr Lesnického Výzkumu 62:182–188 ([in Czech with English abstract and Summary])
Trum F, Titeux H, Ranger J, Delvaux B (2011) Influence of tree species on carbon and nitrogen transformation patterns in forest floor profiles. Ann for Sci 68:837–847. https://doi.org/10.1007/s13595-011-0080-4
Verheyen K, Bossuyt B, Hermy M, Tack G (1999) The land use history (1278–1990) of a mixed hardwood forest in western Belgium and its relationship with chemical soil characteristics. J Biogeogr 26:1115–1128. https://doi.org/10.1046/j.1365-2699.1999.00340.x
Vesterdal L, Schmidt IK, Callesen I, Nilsson LO, Gundersen P (2008) Carbon and nitrogen in forest floor and mineral soil under six common European tree species. For Ecol Manag 255:35–48. https://doi.org/10.1016/j.foreco.2007.08.015
Vopravil J, Formánek P, Heřmanovská D, Khel T, Jacko K (2021) The impact of agricultural land afforestation on soil water content in Central Bohemia. J for Sci 67:512–521. https://doi.org/10.17221/108/2021-JFS
Vu VQ (2011) Ggbiplot. A tool for visualizing the results of PCA—a ggplot2 based biplot. https://github.com/vqv/ggbiplot. Accessed 6 March 2021
Wall A, Hytönen J (2005) Soil fertility of afforested arable land compared to continuously forested sites. Plant Soil 275:247–260. https://doi.org/10.1007/s11104-005-1869-4
Wall A, Westman CJ (2006) Site classification of afforested arable land based on soil properties for forest production. Can J for Res 36:1451–1460. https://doi.org/10.1139/x06-0
Zederer DP, Talkner U, Spohn M, Joergensen RG (2017) Microbial biomass phosphorus and C/N/P stoichiometry in forest floor and A horizons as affected by tree species. Soil Biol Biochem 111:166–175. https://doi.org/10.1016/j.soilbio.2017.04.009
Zielonka A, Drewnik M, Musielok L, Dyderski MK, Struzik D, Smułek G, Ostapowicz K (2021) Biotic and abiotic determinants of soil organic matter stock and fine root biomass in mountain area temperate forests—examples from cambisols under European beech, Norway spruce, and silver fir (Carpathians, Central Europe). Forests 12:823. https://doi.org/10.3390/f12070823
Acknowledgements
The study was supported by the Ministry of Agriculture of the Czech Republic, institutional support MZE-RO0118 and project QK1810126.
Funding
Open access publishing supported by the National Technical Library in Prague.
Author information
Authors and Affiliations
Contributions
J.B. and D.K. designed the experiment and sampled soils, O.Š. analysed the data, prepared tables and figures and wrote body of the manuscript, D.K. lead writing of the discussion, J.L. supervised the research. All authors reviewed the manuscript.
Corresponding author
Ethics declarations
Conflict of interest
The authors have no relevant financial or nonfinancial interests to disclose and no competing interests to declare that are relevant to the content of this article. All authors certify that they have no affiliations with or involvement in any organization or entity with any financial interest or nonfinancial interest in the subject matter or materials discussed in this manuscript. The authors have no financial or proprietary interests in any material discussed in this article.
Additional information
Communicated by Agustin Merino.
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Špulák, O., Kacálek, D., Bartoš, J. et al. The early impact of mixed canopies with Norway spruce, European beech and silver fir on a new forest floor. Eur J Forest Res 142, 851–863 (2023). https://doi.org/10.1007/s10342-023-01557-w
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10342-023-01557-w