
Abstract
There is little knowledge how ungulate pressure on forest regeneration may be mitigated by silvicultural methods. The knowledge is especially needed for artificially regenerated, deciduous tree species. We studied factors affecting browsing incidence by deer in the Pisz Forest District in Poland, an area where 10,000 ha of forest was damaged by a 2002 hurricane. In 2006, we established three experimental plots (in total, 22.6 ha), in which the main species was Scots pine (Pinus sylvestris) admixed with pedunculate oak (Quercus robur). The data on browsing were collected in 2008–2015. In general, oak browsing incidence was unrelated to oak planting density. On a plantation scale, it was significantly affected by the pine age. Although in each variant all the oaks were browsed for four consecutive years (2009–2012), in 2013 browsing incidence began to decrease. When the pines grew higher and formed a physical barrier, it was harder for deer—roe deer (Capreolus capreolus), red deer (Cervus elaphus) and moose (Alces alces)—to move through and locate the oaks. Moreover, within plantations, oak browsing incidence was higher in the patches with shorter pines. Browsing of individual saplings or small groups of saplings was also negatively affected by the height of neighbouring pine saplings. Oak density influenced deer selectivity depending on the tree height. In a low oak tree density, browsing incidence was unrelated to oak height, while in higher tree density, deer selected oaks of the height between 40 and 100 cm. We postulate that deciduous admixture in a coniferous (unattractive) stand can be planted with a few year delay. Older coniferous trees should impede locating of attractive tree species by deer and the browsing incidence.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Recent years have witnessed a growing interest in uneven-aged (Kuuluvainen et al. 2012) and mixed forest stands (Laiho et al. 2011). Mixing of complementary tree species may increase stand productivity, mitigate the effects of drought and other risks, and increase the stability and the efficiency of resource use in forest production systems in response to climate change (Pretzsch et al. 2015). Kelty (2006) and Zhang et al. (2012) observed that mixtures are often much more productive than monocultures. However, there is a general scarcity of systematic empirical studies investigating the effects of mixing commercially important and widespread tree species. The genus oak (Quercus spp.) is possible for admixture in Europe. Besides commercial aspects, oaks are important in nature conservation, and they provide habitats for various groups of organisms (Lindbladh and Foster 2010; Pasinelli 2000; Ranius et al. 2008; Ranius and Jansson 2000; Thor et al. 2010).
Large herbivores, in particular several species of deer (especially roe deer, red deer and moose), cause significant disturbance in European forestry through browsing and bark stripping (Gill 1992; Conover 1997; Reimoser and Putman 2011). These animals generally prefer deciduous over coniferous trees, and oaks are particularly frequently browsed (Götmark et al. 2005; Bergquist et al. 2009), which is why tree admixtures may be difficult to manage in the presence of cervids. Deer density is not a simple determinant of forest damage (Reimoser and Putman 2011), but there is evidence that sport hunting could significantly reduce the negative impacts of browsing (Hothorn and Müller 2010). However, the existing game management systems are, in general, not sufficiently effective to prevent the increase in deer populations in Europe (Apollonio et al. 2010) and North America (Brown et al. 2000). Therefore, attempts to mitigate the negative effects of deer pressure on forest regeneration (both natural and artificial) should be an important part of forest management strategies, even though their effectiveness may be limited by high deer densities (Horsley et al. 2003; Tremblay et al. 2007). Natural forest regeneration has been investigated by several authors (Heuze et al. 2005; Barrett and Schmitz 2013), but there is a great demand for information on the relationship between forest management and deer pressure in man-made plantations. In general, forests managed by clear-cutting are more susceptible to browsing than forests under close-to-nature management (Reimoser and Gossow 1996). This study tested 4 hypotheses relating to the effectiveness of various treatments for managing forests with deciduous admixtures, in particular different planting scenarios, to mitigate deer browsing. The admixed deciduous species was pedunculate oak (Quercus robur)—as mentioned before one of the most attractive tree species for deer. If the tested hypotheses are validated for oak, the relevant findings should also apply to other deciduous tree species.
Deer browsing has been found to be inversely correlated with tree density (Lyly and Saksa 1992; Reimoser et al. 2009; Gerhardt et al. 2013). Therefore, we hypothesised that the percentage of browsed oak saplings will be inversely correlated with oak tree density (Hypothesis 1).
Browsing on deciduous admixtures in coniferous plantations can be regarded as foraging in a feeding station in a plant community (sensu Senft et al. 1987). The pressure on plants (in this case, oak trees) in a feeding station is determined by the matrix of surrounding plants which may form biotic refuges. A biotic refuge for plants physically impedes herbivore foraging by limiting herbivore access and, consequently, herbivore pressure (Milchunas and Noy-Meir 2002). Various studies have demonstrated that protective shrubs (Jensen et al. 2012) and coarse woody debris (Smit et al. 2012) can reduce oak browsing. Therefore, we hypothesised (Hypothesis 2) that pines (Pinus silvestris) as the main species surrounding oak saplings will affect browsing incidence. Specifically, we predicted (P 2.1) that oak browsing will be influenced by the age of the pine plantation and that browsing incidence will decrease as pine trees grow taller. We also predicted (P 2.2) that the incidence of oak browsing will be higher in forest patches with shorter pines where oak saplings are easier to locate. Finally, we predicted (P 2.3) that in a pre-thicket stage, also on a single tree/nest scale (for nest definition see below), browsing incidence will be negatively affected by the number and height of the pine trees located in the direct vicinity.
Browsing damage to coniferous and broad-leaved seedlings increases with tree height up to a certain threshold defined by deer sizes (Pépin et al. 2006). Götmark et al. (2005) also noted that seedlings (<20 cm) were less frequently browsed than saplings (20–130 cm). In general, browsing affects trees at different height classes. In Sitka spruce (Picea sitchensis), which is browsed by red deer and roe deer, the relevant height interval was 30–60 cm (Staines and Welch 1984; Welch et al. 1991). Motta (2003) reported intervals of 20–160 cm and 20–200 cm for Norway spruce and rowan, respectively. However, the cited studies covered a short period of time or as in case of that of Motta (2003) several mountain sites where height intervals were affected by snow depth. Long-term studies investigating the relationship between browsing incidence and height intervals are rare. Moreover, the height of saplings is determined by age; therefore, the effect of height may be difficult to distinguish from the effect of age in naturally regenerated trees of different age. For instance, wood density influences browsing incidence (Boulanger et al. 2009), whereas wood density tends to change with seedling/sapling age (Domec and Gartner 2002). This paper presents data from an 8-year study of browsing incidence in even-aged pine–oak mixtures. We hypothesised (Hypothesis 3) that the width of the browsing interval will change with plantation age and tree density. We predicted (P 3.1) that in a young plantation, deer will be relatively less selective in terms of tree height than in older plantations where trees will be selected at an optimal height range. Renaud et al. (2003) demonstrated in a cafeteria test that tamed red deer with a shoulder height of 80–105 cm selected willow bunches positioned at a height of 85–115 cm. We also predicted (P 3.2) that deer will be more selective in the case of higher oak tree density since high tree availability will enable them to feed on preferred height classes, while low tree availability will not ensure them such a possibility.
Oak stands can be established among others by cluster planting (Saha et al. 2012 and citations therein). Two cluster planting methods are used to establish oak stands: nest planting and group planting—the size of a nest is remarkably smaller than that of a group (Saha et al. 2012 and citations therein). In general, group planting (seedling spacing −1 × 1 m; 19–27 trees per group) is more effective than nest planting (seedlings spacing—0.2 × 0.2 m; 21 trees per nest), whereas nest planting as oak admixtures has been rarely studied. The nest planting approach has been co-developed by Szymański (1983) (see Saha et al. 2012 for the history of the method) who postulated that it should be used mainly for planting oak admixtures in Scots pine plantations. According to Szymański (1983), nests also offer protection against browsing. When nest oaks develop strong root systems, they can grow out of deer reach, and they do not require protection against browsing (Szymański 1983). However, deer densities have increased considerably since the time of Szymański’s study, and the performance of nest clusters under higher browsing pressure remains generally unknown. A nest cluster should feature a central oak which will achieve high final quality due to the positive influence of the surrounding trees. Therefore, it is also essential to know how the location of a tree in the nest influences its browsing by deer. We hypothesised that browsing pressure on nest cluster trees will be determined by their location within the cluster (Hypothesis 4), and we predicted that trees growing in outer portions of the nest will be browsed more intensely than trees with a more central position. The tested hypotheses/predictions are summarised in Table 1.
Study area and experimental design
Study area
The study was conducted in the Pisz Forest District which is a part of the Piska Forest in north-eastern Poland. The study site is a lowland area with a mean annual temperature of 7 °C and a mean annual precipitation of 541 mm (mostly rainfall in the summer months). The growing season is relatively short and lasts 190–200 days. The Pisz Forest District is characterised by a predominance of unfertile sandy soils, and Peucedano-Pinetum is the main forest community. The predominant tree species is Scots pine with an admixture of auxiliary species such as silver birch (Betula pendula), Norway spruce (Picea abies) and black alder (Alnus glutinosa) in more humid sites. In 2002, a local storm event caused significant damage to forests in north-eastern Poland. The most affected area was the Piska Forest, in particular the Pisz Forest District. More than 30,000 hectares of forest was affected. In the Pisz Forest District, the storm damaged 10,000 hectares of forest area. Nearly the entire damaged area was cleared. All trees were cut, wood was logged, and the area was replanted, mainly with Scots pines and admixtures of Norway spruce, silver birch and to a lesser extent small-leaved lime (Tilia cordata) and pedunculate oak. Scots pines were planted in rows, whereas admixed tree species were planted singly. Deer density was moderate during the studied period at 3–4, 3–3.5 and 0.2–0.5 individuals per 100 ha for red deer, roe deer and moose, respectively (deer density data were obtained from the Pisz Forest District Headquarters).
Experimental design
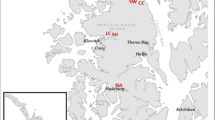
The experiment was established in 2006 in a post-disturbance area. Three different experimental plots were selected (Fig. 1). Each experimental plot was split into two, approximately equal subplots. The Scots pine was planted in one subplot and sown in the other subplot. The first attempt at sowing Scots pine was mostly unsuccessful and was repeated in the spring of 2007. Additional pines were planted where necessary. Since one-year-old bare-rooted pines (and oaks—see below) were planted, sown pine was shorter and younger than the planted one (and the difference was even more pronounced in places where sawing was unsuccessful and repeated a year later). The initial spacing in the planted segment of the pine plantation was 1.5 m between rows and 0.8 m between trees. The seedlings of both species were grown in a local nursery.

Location of the Pisz Forest District (left side) and experimental plots within the study area (right side). Shaded is the area of the Pisz Forest District affected by the hurricane in 2002
Here, pedunculate oak was the only admixed species in the planted and sown segments of all plots. In Variant 1 (total area of 8.88 ha), oaks were planted singly (as every 10th tree in a row) with a density of 900 trees per ha. In Variants 2 and 3, oaks were planted in nests (1 m2, 21 oaks per nest, spacing—0.25 × 0.25 m), and the variants differed in nest spacing. In Variant 2 (6.15 ha), 130 clusters (2835 oak trees) were planted per ha, and in Variant 3 (7.58 ha), 230 clusters (4830 oak trees) per ha. Nest spacing was 8 × 8 m in Variant 2 and 6 × 6 m in Variant 3.
Methods
Data collection
Data on browsing incidence were collected every year in spring 2008–2015, except for the year 2010. Data collection methods differed across the experimental variants (because of the way the oaks were planted—see Experimental design). In Variant 1, the number of browsed trees was counted along 20 randomly mapped transects (10 in the sown and 10 in the planted segments of the plot). Each transect was composed of 10 nearest oak trees in the same or neighbouring rows. The height of every oak tree was measured. In Variants 2 and 3, the data on browsing incidence were collected in 50 randomly selected nests (25 in the sown and 25 in the planted segments of each plot). In each nest, the total number of trees, the number of browsed trees and the height of every tree were recorded. Each oak tree was assigned to one of the three zones within the nest: central (close to the centre of the nest), internal and peripheral (close to the nest boundary) zones. Every zone had the shape of a circle at a width was approximately equal to one-third of the circle’s (nest’s) radius (Fig. 2).

Division of a nest into three zones (EX external zone, IN internal zone, C central zone)
The number and height of trees growing within a radius of 5 m from a sapling/nest were recorded to estimate the effect of a sapling’s/nest’s neighbourhood on browsing incidence. Approximately 75 randomly selected saplings/nests were measured in each experimental variant. In Variant 1, data on the influence of neighbouring trees were gathered in 2013 and 2015, and in Variants 2 and 3—in 2012 and 2014. Browsing incidence in saplings/nests and the height of oak trees were also recorded.
Statistical analysis
The influence of plantation age, experimental variants (oak density), pine regeneration method and sapling/nest surroundings on browsing intensity were analysed using a generalised linear model (GLM) with a logit link function. The same method was also used for the analysis of browsing incidence in relation to oak location within the nest. Statistical hypotheses were verified by the Wald test with approximately Chi-square distribution.
Tree selection based on height was analysed by calculating the degrees of freedom (df) in the Chi-square test depending on the number of height classes in each experimental variant. All calculations were performed using STATISITCA software (StatSoft Inc. 2014).
Results
The size of oak tree samples for evaluating browsing incidence differed across the experimental variants and years of the study (Table 2). In Variant 1, the number of trees was constant at 200 in every year of the study. In Variant 2, the number of trees ranged from 728 to 1100 across years, and in Variant 3, the respective range was 813–1034.
Influence of oak density, plantation age and sapling/nest surroundings on browsing incidence
In the simplest model testing, the influence of plantation age, experimental variant, pine regeneration method and the interactions between these variables on oak browsing, age (p < 0.001) and experimental variant (p = 0.002) were significant variables (Table 3). The year x variant interaction was also significant (p < 0.001). The pine regeneration method and other interactions were non-significant (p > 0.05 in all cases). Although oak browsing incidence differed significantly across the experimental variants (with different sapling density), the noted effect was significant only in the first and in the last three years of the study (Fig. 3). Moreover, the results were inconsistent: in the first year, oak browsing was lowest in Variant 2, whereas in the last year of the study, Variant 2 was characterised by the highest browsing incidence. Therefore, Hypothesis 1 postulating the influence of oak density on browsing incidence was rejected.

Oak browsing intensity depending on plantation age and variant of experiment
As predicted (P 2.1), plantation age significantly influenced oak browsing incidence. In 3-year-old plantations (the first year of data collection), the incidence of oak browsing ranged from 60 to 80% across the experimental variants (Fig. 3). All oak trees were browsed in 4–5- and 7-year-old plantations. As mentioned before, in 2010 (6-year-old plantations), the data were not collected; however, since two years earlier and one year later, browsing concerned all the oak trees, we assume that in 2010 browsing incidence was also equal 100%. In 8-year-old plantations, oak browsing incidence decreased to around 60%. In 9-year-old plantations, browsing incidence further decreased to 20–25%. Finally, in the 10th year, oak browsing incidence remained similar, apart from Variant 2 where it increased to 40% (Fig. 3). As mentioned earlier (refer to Study area), the pine regeneration method (planting vs. sowing) contributed to differences in age and growth rates, and planted pines were older and taller than sown pines. The influence of the pine regeneration method on oak browsing was not significant; therefore, Prediction 2.2 was rejected. When the analysis was repeated for the last three years of the study (oak browsing was below 100%), the simplest model revealed the same explanatory variables, i.e. plantation age (p < 0.001), experimental variant (p < 0.001) and the year x variant interaction (p = 0.005).
The data collected in the last three years of the study were analysed again, but the analysis was limited to oaks from the height interval selected by deer (40–100 cm) (refer to the next section “Oak browsing with regard to tree height” for details). This approach confirmed the importance of the same explanatory variables (Table 4); however, the model also revealed a significant influence of the pine regeneration method (p = 0.046). Oak browsing incidence was higher in subplots where pines were sown (i.e. with smaller trees) than in the subplots where pines were planted (Fig. 4). Therefore, we sustained the prediction (P 2.2) concerning the influence of pine tree height growing in the matrix on oak browsing incidence, but only for the oak height interval selected by deer.

Oak browsing intensity depending on pine regeneration and variant of experiment
Similarly, in the scale of individual sapling/nest, oak browsing incidence was affected by surrounding pines, although the relation was modified by the variant of the study. The analysis was performed separately for oaks from all tree height intervals and for oaks in the 40–100 cm interval. For all oak heights in Variant 1, browsing incidence was negatively influenced by pine height (p = 0.045), whereas the number of pine trees had no influence on oak browsing incidence (p > 0.05) (Table 5). In Variants 2 and 3, oak browsing incidence was negatively influenced by pine height (p < 0.001 in both cases), and although the influence of the number of trees was significant (p < 0.001 in both cases), the relation was positive (Table 5). An analysis of oak height intervals selected by deer (40–100 cm) revealed that the influence of pines surrounding oak saplings/nests on browsing incidence was similar to that noted for all height intervals. In Variant 1, oak browsing was negatively affected by pine height (p = 0.035) and unrelated to the pine number (p > 0.05). In Variants 2 and 3, oak browsing incidence was negatively influenced by pine height and positively by pine number (p < 0.001 in all cases) (Table 5). Prediction P 2.3 (regarding the influence of pine height) was only partially confirmed because the influence of pine height and the number of pines differed between experimental variants (refer to Discussion for an explanation for the positive influence of the number of surrounding pines). Nevertheless, taking into account all predictions, Hypothesis 2 postulating the influence of pine trees on browsing incidence in the oak admixture was confirmed. Therefore, since the trends for pine height and number differed, we have only partly sustained Prediction P 2.3 (namely its part regarding the influence of the pine height). Taking into account all the predictions, we sustained Hypothesis 2 referring to the influence of pine trees on the browsing incidence of oak admixture.
Oak browsing with regard to tree height
The analysis of the influence of oaks’ height on browsing was performed only for the years of the study when browsing incidence was below 100%. In general, browsing incidence in relation to oak height varied across the experimental variants (Fig. 5). In Variant 1, oak browsing was proportional to the availability of height classes (p > 0.05 in all analysed years). Therefore, no changes in the selection of tree height were observed in the analysed period in Variant 1. In Variants 2 and 3, oak browsing differed from the availability in the height classes (p < 0.001 in all cases). However, trees were much shorter in the third year of the plantation’s life; therefore, browsing incidence related to height was difficult to compare with years 8–10. Nevertheless, deer appeared to avoid the shortest trees (0–20 and 20–40 cm) in both experimental variants in the third year (Fig. 5). The same trend was noted in both variants in years 8–10. In the third year of the study, in Variants 2 and 3, browsing incidence on trees from 40 to 60 cm and 60–80 cm height classes significantly exceeded their availability. The above classes as well as the 80–100 cm height class (Variants 2 and 3) and the 100–120 cm height class (year 8, Variant 2) were also selected in the second half of the experiment, excluding the last year in Variant 3 when all trees taller than 40 cm were browsed proportionally to their availability (Fig. 5). However, we were unable to reject or confirm Prediction 3.1 due to the long period of browsing on all oak trees.

Oak browsing intensity in relation to tree height depending on different years and variants of the study
As mentioned before, deer did not exhibit a clear selection of any height class in any of the studied years in Variant 1 (p > 0.05 in all cases). In Variants 2 and 3, deer expressed clear selection for certain height classes (p < 0.001 in all cases). Therefore, Prediction 3.2 was sustained with a restriction for low (Variant 1) versus higher (Variants 2 and 3) densities. No significant differences between moderate (Variant 2) and high (Variant 3) densities were observed.
The influence of oak location in the nest on browsing incidence
The location of oak trees in the nest significantly influenced browsing incidence, and oaks located in the central zone of the nest were least browsed (Fig. 6). Browsing incidence in the internal zone was higher than in the central zone, but the noted difference was not significant (p > 0.05). Browsing incidence was highest in the peripheral zone, and it was significantly higher than in the internal zone and the central zone (p < 0.001 in both cases). Therefore, Hypothesis 4 was sustained.

Oak browsing intensity depending on the location within a nest (results for Variants 3 and 4)
Discussion
Hypothesis 1 postulating that the density of oak admixtures influences browsing incidence was rejected. The absence of such a relation could seem surprising because a negative correlation between tree density and browsing incidence was reported in several studies (Lyly and Saksa 1992; Reimoser et al. 2009; Gerhardt et al. 2013). For instance, a systematic literature review by Gerhardt et al. (2013) revealed that naturally regenerated forests tend to be less vulnerable to browsing than artificially regenerated stands because natural forests are characterised by higher tree density than artificial stands. However, other authors examined the relation between forest regeneration and deer damage in relatively large spatial scales. Our study was performed in three experimental plots (total area of 22.6 ha) and therefore in relatively small spatial scale. Moreover, we did not study the main tree species in stands but only the admixed species (which further limits the number of the analysed trees). The examined habitats were relatively poor (refer to Study area) with very few deciduous species, mostly birch. The majority of young plantations with oak admixtures were fenced off to protect trees against browsing by deer. Selected pine plantations were also fenced off. In such habitats, oaks were highly attractive for deer, as demonstrated by the fact that 100% of oaks were browsed for several consecutive years in all experimental plots representing three oak density variants. Tree species with high nutrient content are usually browsed more intensely than trees with lower nutritional value (Gerhardt et al. 2013). In our study, despite generally very high levels of deer pressure on oak admixtures, the density of browsed trees would modify to some extent deer impact in particular in the study period when browsing incidence had decreased (refer to Discussion).
As hypothesised (Hypothesis 2), browsing incidence on oak admixtures was influenced primarily by the characteristics of the main tree species in the analysed plantations. It should be noted that the main tree species was Scots pine, and coniferous trees are generally far less browsed by deer than deciduous trees, in particular oaks (Kullberg and Bergström 2001). In our study, browsing incidence on pines was negligible, whereas all oak trees (at least for some years) were browsed. In the pines studied by us, browsing incidence generally did not exceed 10% (Borkowski et al. unpublished data). If the main species had been more attractive for deer than pine, probably it would not have offered effective protection for the oak admixture. As predicted (P 2.1), the age of the plantation significantly influenced browsing incidence. In 3-year-old plantations, the percentage of browsed trees ranged from 60 to 80% across the experimental variants. All oaks were browsed in the following several years, and browsing incidence began to gradually decrease in year 8. The plantations were highly accessible to deer when pine trees were young, relatively short and flexible, but they were more difficult to access when pine stands grew taller and thicker, and the canopy began to close. As a result, deer movement was probably limited, and the animals probably found it more difficult to locate oak trees. The mitigating influence of naturally regenerated, unattractive tree species (balsam fir) on the browsing incidence of more attractive species (Eastern hemlock) has been described by Borgmann et al. (1999) for naturally regenerated trees. The authors concluded that “balsam fir patches are typically quite dense, hard to manoeuvre through, and difficult to visually inspect, thus creating a physical barrier for white-tailed deer”. In the literature, other types of biotic (browsing) refuges were also found to reduce herbivore pressure on target trees, including shrubs with spines (Barbosa et al. 2009), forest understorey (Heuze et al. 2005; Jensen et al. 2012), woody debris (Smit et al. 2012) and slash on clear-cuts (Bergquist and Örlander 1998). Our study provides evidence that also in artificially regenerated stands, browsing on attractive admixed tree species can be reduced by a browsing refuge created by a less attractive main tree species.
It is theoretically possible that the decrease in browsing incidence observed after the 7th year in the studied plantations was induced by other factors. Firstly, it is possible that most oaks grew to a height that was not accessible to deer. However, this explanation seems improbable. In the selected oak height classes (40–100 cm), browsing incidence in 8-year-old plantations ranged from 60 to 86% across the experimental variants (Table 6). In the following years, browsing incidence in the selected height classes decreased to 20-40%, apart from 10-year-old-plantations in Variant 2 where 55-66% of oak saplings were browsed. Additionally, deer density is another factor influencing browsing incidence (Gerhardt et al. 2013). However, in our study, changes in browsing incidence were correlated with neither the density of red deer (Pearson correlation: r = −0.14, p = 0.73) nor the combined densities of moose, red deer and roe deer (Pearson correlation: r = −0.29, p = 0.56) during the study period. Note that the data on deer density were obtained from the forest district and one may question the reliability of density estimates. Deer numbers were estimated by the drive count method (J. Buszyniewicz, Pisz Forest District, pers. comm.). If well performed, this method provides reliable estimates on population size (Borkowski et al. 2011). The estimated red deer density are in line with other estimates in north-eastern Poland (2.6–7.6 ind/100 ha, Borowik and Jędrzejewska 2017). To sum up, the biotic refuge explanation seems to be most plausible.
Pine characteristics influenced oak browsing incidence not only in the plantation scale, but also in the patch scale (P 2.2 was sustained). When oak trees from the entire height range in all the years of the study were considered, the height of pine trees in the analysed patches did not affect oak browsing incidence. Similarly, there was no effect when the entire height range of oak trees in the last three years (when browsing incidence decreased) was considered. However, an analysis involving only the oaks from the height class selected by deer (40–100 cm) in the last three years of the study revealed higher browsing incidence in plots with shorter pines. Jensen et al. (2012) investigated the incidence of oak browsing in relation to protection provided by shrubs and found that browsing incidence increased when seedlings became taller than the shrub height. However, in our study, deer manoeuvrability was probably even a more important factor than pine height. As mentioned before, pine trees regenerated by sowing were younger and shorter than planted pines; therefore, deer found it easier to move and locate oak saplings/nests. In 8-year-old plantations, the mean height of sown pines was 186 cm, whereas the mean height of planted pines (303 cm) was nearly 120 cm greater (Borkowski et al. unpublished data).
The relation between browsing incidence and the features of the pine trees growing in the direct vicinity of individual saplings/nests was also examined (P 2.3 was only partially sustained; see Discussion). A negative relation between oak browsing incidence and the height of pine trees surrounding saplings/nests was observed in all three variants. However, similarly to the discussed influence of pine characteristics on the level of browsing in a patch scale, also in the scale of individual saplings/nests, besides the pine height, the age of pine probably also played an important role. Saplings/nests surrounded by shorter and younger (more flexible) pines were easier to access than those surrounded by taller and older (more rigid) pines. However, this explanation should be treated with caution. It is possible that the effect of direct neighbourhood on oak sapling/nest browsing is an artefact, and that saplings/nests were browsed more frequently in plots of sown than planted pines. However, the annually inspected saplings/nests were selected randomly (and their location was not recorded); therefore, we were unable to verify this assumption.
Surprisingly, the number of pines in the direct vicinity was positively related to nest browsing incidence in Variants 2 and 3. A reverse relation was anticipated because oaks surrounded by a higher number of pines should be less accessible and, consequently, exposed to lower deer pressure. This observation could also be linked with the fact that sown pines were more numerous than planted pines, but they were younger, shorter and, consequently, they provided easier access to oak nests. It has been reported that trees surrounded by fewer neighbours are more susceptible to browsing (Gerhardt et al. 2013).
Interestingly, browsing incidence was influenced not only by pine trees surrounding the oak admixture, but also by the characteristics of oak trees. The height of oak trees was an important factor, but not (for obvious reasons) in 4–5- and 7-year-old plantations where all trees were browsed (most possibly, it concerned also 6-year-old plantations, when the data were not collected). In the remaining years of the study, the correlations between browsing incidence and tree height varied across the experimental variants. In Variant 1, oak browsing was independent of tree height, and all height classes were browsed to a similar extent. In Variants 2 and 3, deer avoided the shortest trees (up to 40 cm) in both young (3 year old) and older plantations (second half of the study). Similar results were reported by other authors (Kullberg and Bergström 2001; Palmer and Truscott 2003; Götmark et al. 2005; Smit et al. 2012). In relatively young saplings, the above could be attributed to ground vegetation which protects short saplings, since browsing incidence increases when sapling height exceeds that of ground vegetation (Palmer and Truscott 2003). In this study, the above explanation could not be applied to plantations older than 8 years when pine trees formed thickets. The shortest trees in nests had already been negatively affected by competition with taller trees which were in better condition. According to the plant vigour hypothesis (Price 1991), herbivores feed preferentially on vigorous, large and fast-growing plants or plant parts. The above hypothesis probably explains why small saplings were avoided by deer in the second half of the experiment.
In Variants 2 and 3, deer not only avoided shortest saplings, but also showed a clear selection for certain oak height classes. In plantations older than 7 years, browsing intensity was highest in oaks with the height of 40–100 cm. Browsing generally affects trees at the intermediate level between the ground and the full reach of the deer species (Gill 1992). The browsing range noted in the present study is consistent with that reported by other authors (Palmer and Truscott 2003; Renaud et al. 2003). However, we were unable to reject or confirm Prediction 3.1. In a sense, the browsing range got narrow (after the period when all oak trees were browsed); however, we did not expect all oaks to be browsed; therefore, this is not exactly the scenario we had predicted.
According to Prediction 3.2, the browsing range was negatively influenced by tree density; however, this difference was observed only between Variant 1 and the remaining experimental variants (with no differences between them). As mentioned before, in Variant 1 where oaks were planted individually, deer expressed no selection towards oak heights, and all height classes were browsed proportionally to their availability. Since, as already discussed, no differences in browsing incidence were noted between the variants, the consequences of the deer pressure for oak growth in Variant 1 were probably more serious than in the variants with higher oak tree densities. Dobrowolska et al. (in prep.) found that oak trees in Variant 1 were shorter than in the remaining variants. As a result, in Variant 1, not only did the deer pressure concentrate on a smaller tree number, but deer were also responsible for the lower height of oaks and the lower diversity of height classes, which reduced their choice of trees in a given height range. In the remaining variants, deer pressure concerned a considerably higher number of trees, which, despite browsing, grew faster and were more diversified in height. The observed diversity in height was probably also increased by intraspecific competition between oaks because competition within nests tends to be relatively more intense than in other oak regeneration methods (Saha et al. 2014). For this reason, Hypothesis 3 was sustained only in the part regarding tree density.
Hypothesis 4 regarding the relation between browsing incidence and the location of trees inside a nest was sustained. As predicted, peripheral oak trees were browsed more frequently than oaks in the internal zone. Our findings are consistent with the observations made by Szymański (1986) who reported that peripheral oaks provide browsing protection for internal trees. Despite higher browsing incidence, peripheral oaks in our nests were taller than those in the internal zone (Dobrowolska, in prep.). The discussed oak regeneration method became popular at the end of the twentieth century in Germany and France (Saha et al. 2012), but very little is known about browsing incidence in trees planted in nests. Nest planting cannot be directly compared with other regeneration methods (such as single oak planting) based on our findings because the analysed plantations also differed considerably in oak density. Saha et al. (2012) did not study browsing incidence, but their meta-analysis revealed that browsing has far greater consequences (growth and quality) for oaks that are planted in nests than for trees that are planted in rows. Further research on our plantations should provide more information on oak performance in nests, such as the information on the quality of oaks. This point is of additional importance since the majority of oak trees were exposed to severe browsing during several consecutive years.
Conclusions
At relatively poor forest sites, oak seedlings and saplings (as probably other highly attractive tree species) may suffer from intense browsing for several consecutive years. Increasing the planting density of admixed trees may not help to limit the browsing incidence. It has been postulated that regeneration success should be measured by the number of unbrowsed rather than of browsed trees (Reimoser and Gossow 1996). In general, it is obviously true, however, that at poor sites, literally all the deciduous trees may be browsed, and it may be hard to find any unbrowsed ones. If oak is to serve primarily as a biocenotic admixture (i.e. its timber quality is not a primary goal), browsing may be of lesser importance if at least some saplings will survive.
A large number of studies on browsing are short term in character and therefore not always helpful for forest management (Senn and Suter 2003). Our study shows that within a relatively short period, browsing incidence can change remarkably. For instance, browsing survey done in the seventh year of the plantation would show 100% of oak browsing, while two years later, it would be by nearly 80% lower. Long-term research on the consequences of severe browsing on tree development and quality is clearly required.
Although browsing has an enormous impact on forestry, unsatisfying achievements of silviculture in relation to browsing pressure may come not only from the intensity of damage by herbivores, but also from unsuitable silviculture practices (Heuze et al. 2005). Moreover, regulation of herbivore populations may not have a substantial effect on the level of browsing (e.g. Månsson 2009, but see Hothorn and Müller 2010). It is therefore essential, as mentioned in Introduction, to develop new forest management methods which may reduce the effects of large herbivores on forest regeneration. The issue is especially important for artificially regenerated forests, since trees in man-made stands are usually substantially more exposed to damage than their naturally regenerated counterparts (Beguin et al. 2016). However, as already mentioned in Introduction, one should keep in mind that the option to reduce the negative effects of deer browsing on tree regeneration by silvicultural measures may become rather limited if the deer density is high (Horsley et al. 2003; Tremblay et al. 2007). Therefore, silvicultural measures cannot replace appropriate deer management and should be seen as a supplementary option. Our study suggests that browsing incidence of admixed tree species may depend not only on their age, but also on the age of plantations’ “matrix species”. Incidence of oak browsing significantly decreased when the pine trees came to closure and formed a thicket. In such conditions, deer had difficulties to move within the thickets and locate the oak trees. In most cases, main and admixed species are planted at the same time. However, if a plantation’s main species is relatively unattractive for herbivores, the admixed species planted several years (e.g. 5–7) after the main species should be much less vulnerable to browsing. Obviously, the admixed trees might then be planted in sites ensuring them enough space and light to grow. One of the potential problems arising when applying such an approach is a competition between the main and relatively much younger admixed species. We believe, however, this can be solved to a remarkable extent, by promoting the admixed species’ individuals/nests when cleaning (successively removing competitors from the main species) and providing a growing space for the admixed trees. Probably, seedlings used for the delayed planting should be a few years old (older than 1 year).
Our study was performed with oak, but probably such a delayed admixture planting might be applied to the majority of attractive admixed species. Concerning the main species, coniferous trees seem to be more effective in a protection of delayed admixture regeneration. When they come to closure, their branches tend to spread more in comparison with the majority of deciduous species and secondly, the branches are also present in the lower part of the stem, which is not the case in deciduous species. As a result, it may be harder for deer to move and visually inspect a coniferous plantation than a deciduous one.
The height of the main tree species growing in the vicinity of the admixed trees was an important factor influencing the browsing incidence in our study. This finding has at least two important implications for the planting of admixed trees—planting them in the patches with slower-growing main tree species may require: (1) further delay in the planting time (until the main tree species reaches satisfactory height) or (2) protection of the patches with shorter main trees by fencing or individual tree protection methods. Our study suggests that also in artificial forest regeneration, there is a space to modify silviculture methods to mitigate the impact of large herbivores on young plantations. It needs to be emphasised that, although the strong point of our study is its length, its weakness is the lack of replications, which additionally stresses its preliminary character. Definitely, further research is needed on a proposed delayed admixture planting, testing particular methods of planting, suitable tree species used (both main and admixed ones) and the development of the admixed trees under such a treatment.
References
Apollonio M, Andersen R, Putman R (2010) European ungulates and their management in the 21st century. Cambridge University Press, Cambridge
Barbosa P, Hines J, Kaplan I, Martinson H, Szczepaniec A, Szendrei Z (2009) Associational resistance and associational susceptibility: having right or wrong neighbors. Annu Rev Ecol Evol Syst 40:1–20
Barrett KJ, Schmitz OJ (2013) Effects of deer settling stimulus and deer density on regeneration in a harvested southern New England forest. Int J For Res. doi:10.1155/2013/690213
Beguin J, Tremblay JP, Thiffault N, Pothier D, Côté SD (2016) Management of forest regeneration in boreal and temperate deer—forest systems: challenges, guidelines, and research gaps. Ecosphere 7(10):e01488. doi:10.1002/ecs2.1488
Bergquist J, Örlander G (1998) Browsing damage by roe deer on Norway spruce seedlings planted on clearcuts of different ages 1. Effect of slash removal, vegetation development, and roe deer density. For Ecol Manag 105:283–293
Bergquist J, Löf M, Örlander G (2009) Effects of roe deer browsing and site preparation on performance of planted broadleaved and conifer seedlings when using temporary fences. Scand J For Res 24:308–317
Borgmann KL, Waller DM, Rooney TP (1999) Does balsam fir (Abies balsamea) facilitate the recruitment of eastern hemlock (Tsuga canadensis)? Am Midl Nat 141:391–397
Borkowski J, Palmer SCF, Borowski Z (2011) Drive counts as a method of estimating ungulate density in forests: mission impossible? Acta Theriol 56:239–253
Borowik T, Jędrzejewska B (2017) Heavier females produce more sons in a low-density population of red deer. J Zool 302:57–62
Boulanger V, Baltzinger C, Said S, Ballon P, Picard JF, Dupouey JL (2009) Ranking temperate woody species along a gradient of browsing by deer. For Ecol Manag 258:1397–1406
Brown T, Decker DJ, Riley SJ, Enck JW, Lauber TB, Curtis PD, Mattfeld GF (2000) The future of hunting as a mechanism to control white-tailed deer populations. Wildl Soc Bull 28:797–807
Conover MR (1997) Monetary and intangible evaluation of deer in the United States. Wildl Soc Bull 25:298–305
Domec JC, Gartner BL (2002) Age- and position-related changes in hydraulic versus mechanical dysfunction of xylem: inferring the design criteria for Douglas fir wood structure. Tree Physiol 22:91–104
Gerhardt P, Arnold JA, Hackländer K, Hochbichler E (2013) Determinants of deer impact in European forests—a systematic literature analysis. For Ecol Manag 310:173–186
Gill RMA (1992) A review of damage by mammals in north temperate forests. 1. Deer. Forestry 65:145–169
Götmark F, Berglund Å, Wiklander K (2005) Browsing damage on broadleaved trees in semi-natural temperate forest in Sweden, with a focus on oak regeneration. Scand J For Res 20:223–234
Heuze P, Schnitzler A, Klein F (2005) Is browsing the major factor of silver fir decline in the Vosges Mountains of France?. For Ecol Manag 217:219–228
Horsley SB, Stout SL, DeCalesta DS (2003) White-tailed deer impact on the vegetation dynamics of a northern hardwood forest. Ecol Appl 13:98–118
Hothorn T, Müller J (2010) Large-scale reduction of ungulate browsing by managed sport hunting. For Ecol Manag 260:1416–1423
Jensen AM, Götmark F, Löf M (2012) Shrubs protect oak seedlings against ungulate browsing in temperate broadleaved forests of conservation interest: a field experiment. For Ecol Manag 266:187–193
Kelty MJ (2006) The role of mixtures in plantation forestry. For Ecol Manag 33:195–204
Kullberg Y, Bergström R (2001) Winter browsing by large herbivores on planted deciduous seedlings in southern Sweden. Scand J For Res 16:371–378
Kuuluvainen T, Tahvonen O, Aakala T (2012) Even-aged and uneven-aged forest management in boreal Fennoscandia: a review. Ambio 41:720–737
Laiho O, Lahde E, Pukkala T (2011) Uneven- vs even-aged management in Finnish boreal forests. Forestry 84:547–556
Lindbladh M, Foster DR (2010) Dynamics of long-lived foundation species: the history of Quercus in southern Scandinavia. J Ecol 98:1330–1345
Lyly O, Saksa T (1992) The effect of stand density on moose damage in young Pinus sylvestris stands. Scand J For Res 7:393–403
Månsson J (2009) Environmental variation and moose Alces alces density as determinants of spatio-temporal heterogeneity in browsing. Ecography 32(4):601–612
Milchunas DG, Noy-Meir I (2002) Grazing refuges, external avoidance of herbivory and plant diversity. Oikos 99:113–130
Motta R (2003) Ungulate impact on rowan (Sorbus aucuparia L.) and Norway spruce (Picea abies (L.) Karst.) height structure in mountain forests in the Eastern Italian Alps. For Ecol Manag 181:139–150
Palmer SCF, Truscott AM (2003) Browsing by deer on naturally regenerating Scots pine (Pinus sylvestris L.) and its effects on sapling growth. For Ecol Manag 182:31–47
Pasinelli G (2000) Oaks (Quercus sp.) and only oaks? Relations between habitat structure and home range size of the middle spotted woodpecker (Dendrocopos medius). Biol Conserv 93:227–235
Pépin D, Renaud PC, Boscardin Y, Goulard M, Mallet C, Anglard F, Ballon P (2006) Relative impact of browsing by red deer on mixed coniferous and broad-leaved seedlings—an enclosure-based experiment. For Ecol Manag 222:302–313
Pretzsch et al (2015) Growth and yield of mixed versus pure stands of Scots pine (Pinus sylvestris L.) and European beech (Fagus sylvatica L.) analysed along a productivity gradient through Europe. Eur J For Res 134(5):927–947
Price PW (1991) The plant vigor hypothesis and herbivore attack. Oikos 62:244–251
Ranius T, Jansson N (2000) The influence of forest regrowth, original canopy cover and tree size on saproxylic beetles associated with old oaks. Biol Conserv 95:85–94
Ranius T, Eliasson P, Johansson P (2008) Large-scale occurrence patterns of redlisted lichens and fungi on old oaks are influenced both by current and historical habitat density. Biodivers Conserv 17:2371–2381
Reimoser F, Gossow H (1996) Impact of ungulates on forest vegetation and its dependence on the silvicultural system. For Ecol Manag 88:107–119
Reimoser F, Putman RJ (2011) Impact of large ungulates on agriculture, forestry and conservation habitats in Europe. In: Putman RJ, Apollonio M, Andersen R (eds) Ungulate management in Europe: problems and practices. Cambridge University Press, Cambridge, pp 144–191
Reimoser S, Partl E, Reimoser F, Vospernik S (2009) Roe-deer habitat suitability and predisposition of forest to browsing damage in its dependence on forest growth—model sensitivity in an alpine forest region. Ecol Model 220:2231–2243
Renaud PC, Verheyden-Tixier H, Dumont B (2003) Damage to saplings by red deer (Cervus elaphus): effect of foliage height and structure. For Ecol Manag 181:31–37
Saha S, Kuehne C, Kohnle U, Brang P, Ehring A, Geisel J, Leder B, Muth M, Petersen R, Peter J, Ruhm W, Bauhus J (2012) Growth and quality of young oaks (Quercus robur and Quercus petraea) grown in cluster plantings in central Europe: a weighted meta-analysis. For Ecol Manag 283:106–118
Saha S, Kuehne C, Bauhus J (2014) Intra- and interspecific competition differently influence growth and stem quality of young oaks (Quercus robur L. and Quercus petraea (Mattuschka) Liebl.). Ann For Sci 71:381–393
Senft RL, Coughenour MB, Bailey DW, RittenhouseLR Sala OE, Swift DM (1987) Large herbivore foraging and ecological hierarchies. Bioscience 37:789–799
Senn J, Suter W (2003) Ungulate browsing on silver fir (Abies alba) in the Swiss Alps: beliefs in search of supporting data. For Ecol Manag 181:151–164
Smit C, Kuijper DPJ, Prentice D, Wassen MJ, Cromsigt JPGM (2012) Coarse woody debris facilitates oak recruitment in Białowieża Primeval Forest, Poland. For Ecol Manag 284:133–141
Staines BW, Welch D (1984) Habitat selection and impact of red (Cervus elaphus L.) and roe (Capreolus capreolus L.) deer in a Sitka spruce plantation. Proc R Soc Edinb 82B:303–319
StatSoft Inc. (2014) STATISTICA (data analysis software system), version 12. www.statsoft.com
Szymański S (1983) Results of 30 years experimentation on the group plantation of oak. (in Polish with English summary). Sylwan 27(9–10):109–114
Szymański S (1986) Die Begründung von Eichenbeständen in Nest-Kulturen. Forst Holzwirtsch 41:3–7
Thor G, Johansson P, Jönsson MT (2010) Lichen diversity and red-listed lichen species relationships with tree species and diameter in wooded meadows. Biodivers Conserv 19:2307–2328
Tremblay J-P, Huot J, Potvin F (2007) Density-related effects deer browsing on the regeneration dynamics of boreal forests. J Appl Ecol 44:552–562
Welch D, Staines BW, Scott D, French DD, Catt DC (1991) Leader browsing by red and roe deer on young Sitka spruce trees in western Scotland: I. Damage rates and the influence of habitat factors. Forestry 64(1):61–82
Zhang Y, Chen HYH, Reich PB (2012) Forest productivity increases with evenness, species richness and trait variation: a global meta-analysis. J Ecol 100(3):742–749
Acknowledgements
The study was supported by General Directorate of State Forests (BLP-278 and BLP359). We are grateful to Pisz Forest District Headquarters for their help in the study. Special thanks are directed to Andrzej Pawelczyk, forester from Wilczy Dół Forest Sub-district.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by Christian Ammer.
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.
About this article

Cite this article
Borkowski, J., Dobrowolska, D., Dąbrowski, W. et al. Young conifer stands form a deer browsing refuge for an oak admixture: silvicultural implications for forest regeneration under herbivore pressure. Eur J Forest Res 136, 787–800 (2017). https://doi.org/10.1007/s10342-017-1070-3
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10342-017-1070-3