
Abstract
One of the most critical functions of nests in parids is thermal insulation, particularly during egg incubation by females and during the first week of the hatchlings’ life when they are poikilothermic. Several avian studies reported associations between aspects of breeding success and nest parameters. In this study, we present results regarding long-term (eleven breeding seasons) impact of nest parameters on breeding characteristics in Great Tits (Parus major) and Eurasian Blue Tits (Cyanistes caeruleus) in a deciduous forest and an urban parkland study area. Our findings indicate that in Great Tits and Eurasian Blue Tits the number of fledglings (and in Eurasian Blue Tits also clutch size tended to be) was positively associated with lining mass and was influenced by the study area, with higher values observed in the forest. Additionally, there was a relationship between lining mass and higher hatching success in both tit species. We also demonstrated that both clutch size and number of fledglings were higher in Great Tits in the forest and in Eurasian Blue Tits the number of fledglings was higher in the forest. The study site also impacted hatching success, with Great Tits showing higher rates in the forest, and fledging success for Eurasian Blue Tits being higher in the forest. The number of fledglings differed between years in Great Tits and hatching success and fledging success differed between years in both Great and Eurasian Blue Tit species. We conclude that avian nests play an important role during reproduction, and the materials used to construct nests contribute to various functional properties. However, further studies are needed to establish clear relationships between nest parameters and breeding performance.
Zusammenfassung
Nestbauweise und Bruterfolg bei zwei Meisenarten: eine Langzeitstudie
Eine der wichtigsten Funktionen von Meisennestern ist ihre Wärmeisolierung, insbesondere während der Bebrütung der Eier durch die Weibchen und während der ersten Lebenswoche der Jungtiere, wenn diese noch wechselwarm sind. In mehreren Untersuchungen konnte ein Zusammenhang zwischen Merkmalen für den Bruterfolg und Eigenschaften der Nester festgestellt werden. In dieser Studie werden die Ergebnisse langfristiger (elf Brutsaisons) Auswirkungen von Nestparametern auf die Bruteigenheiten von Kohlmeisen (Parus major) und Blaumeisen (Cyanistes caeruleus) sowohl in einem Laubwald als auch in einer städtischen Parklandschaft untersucht. Unsere Ergebnisse deuten darauf hin, dass bei Kohlmeisen und Blaumeisen die Anzahl der Jungvögel (und bei Blaumeisen tendenziell auch die Gelegegröße) positiv mit der Ausfütterung des Nests und dem Untersuchungsgebiet zusammenhängt, wobei im Wald höhere Werte für einen solchen Zusammenhang festgestellt wurden. Außerdem gab es bei beiden Meisenarten einen Zusammenhang zwischen der Ausfütterung des Nests und einem größeren Schlüpferfolg. Wir konnten auch nachweisen, dass sowohl die Gelegegröße als auch die Zahl der Jungvögel von Kohlmeisen im Wald gößer war, bei den Blaumeisen nur die Anzahl der Jungvögel. Es gab ebenfalls einen Zusammenhang zwischen dem Untersuchungsgebiet selbst und dem Bruterfolg, wobei Kohlmeisen im Wald höhere Schlüpfraten aufwiesen, während bei den Blaumeisen mehr Jungvögel flügge wurden. Die Anzahl der flügge werdenden Jungvögel war bei den Kohlmeisen von Jahr zu Jahr unterschiedlich wie auch der Schlüpferfolg und die Anzahl der flügge werdenden Jungen bei Kohl- und Blaumeisen. Wir schließen daraus, dass die Nester selbst bei der Fortpflanzung eine wichtige Rolle spielen und die beim Nestbau verwendeten Materialien zu verschiedenen funktionalen Eigenschaften eines Nests beitragen. Weitere Untersuchungen sind jedoch notwendig, um eindeutige Zusammenhänge zwischen solchen Nestparametern und dem Bruterfolg nachzuweisen.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
For many animals, the rearing microenvironment is a crucial factor influencing early offspring development and behaviour. Many bird offspring develop in nests and nest architecture is an extended phenotype that results from the interaction between the evolutionarily adaptive behaviour of birds and the environment (Dawkins 1982; Hansell 2000). In altricial birds, a nest may be considered as a product of niche construction provided by the parents and aimed to optimize living conditions of the offspring (Mainwaring 2017; Maziarz et al. 2017; Vanadzina et al. 2023). Nests reduce both heat loss and overheating and, therefore, may create a suitable microclimate, as well as they can render protection from predators and help to maintain optimal position of eggs during incubation (Heenan 2013). Nests may also protect nestlings from ectoparasites and pathogens or facilitate nestling care and, therefore, nest may improve nestling survival and increase fitness (Mainwaring et al. 2014; Lambrechts et al. 2017; Glądalski et al. 2020a, b; Nuhlíčková et al. 2021).
Nest parameters vary largely between and even within taxa according to their particular life-history (Alabrudzińska et al. 2003; Heenan 2013; Sonnenberg et al. 2020; Glądalski et al. 2024). In our study populations, a tit nest usually consists of two layers. The moss part (usually larger) is considered rather as a sanitary layer (mainly important after hatching), and it also provides a structural support for offspring and parents (Stenning 2018; Rydgren et al. 2023; but also note that some moss species were shown to have antimicrobial and antifungal properties, see Wolski et al. 2021). The lining part (usually thinner) of the nest was shown to be associated mainly with its insulating capacity (Soler et al. 1998; Heenan and Seymour 2011).
Nest size and structure have been shown to be positively related to reproductive parameters, such as clutch size, fledging and hatching success or recruitment success, as well as quality and condition of the nest-building female (Alabrudzińska et al. 2003; Álvarez and Barba 2008, 2011; Mainwaring and Hartley 2008; Álvarez et al. 2013; Glądalski et al. 2016a, b). On the other hand, several field studies showed that associations between breeding parameters and nest parameters were non-significant, rather weak or differed between years within study areas (Britt and Deeming 2011; Lambrechts et al. 2016; see Harnist et al. 2020). It was suggested that it is not nest size per se (volume or mass), but the mass of the lining material in and surrounding the nest cup that might directly impact on breeding success in all positive-effect studies. Järvinen and Brommer (2020) conclude their eight-season study on the nests of Eurasian Blue Tits (Cyanistes caeruleus) that a higher proportion of feathers in the nest lining is positively associated with the probability of offspring to recruit as a breeding adult later in life and, therefore, the choice of nest material may be a maternal behaviour with potential lifelong effects on her offspring. On the other hand, Sonnenberg et al. (2020) conclude their two-season study on the Mountain Chickadee (Poecile gambeli) that in some tit species, the large variation in nest size, both across and within sites, may be rather a result of consistent individual preferences, which may not have clear fitness consequences.
Studies examining the impact of nest parameters on breeding performance and success are not numerous (especially when compared to other stages of bird reproduction) and the number of nest-studies is slowly increasing in recent years (Mainwaring et al. 2014; Harnist et al. 2020). Especially, long-term studies are still rare and the vast majority of reports refer usually to one to two seasons of research and inferring from such a small number of seasons in some cases can give incorrect or even opposite results. The aim of this paper is to study the long-term relation between nest structure and reproductive performance, including hatching success and fledging success of Great Tits (Parus major) and Eurasian Blue Tits in two floristically and structurally contrasting study areas: a rich deciduous forest and an urban parkland during eleven breeding seasons (2012–2022). Our prediction is that in both tit species clutch size and the number of fledglings will be positively associated with lining mass (and that both breeding parameters will be affected by the study area). We also predict that breeding success will be positively affected by nest characteristics (nest height/mass or/and lining mass).
Materials and methods
This study was carried out as part of long-term research project concerning the breeding biology of secondary hole nesting birds occupying nest boxes in the city of Łódź (central Poland, Glądalski et al. 2018). Nests examined for this study were obtained between 2012 and 2022 (eleven breeding seasons) in two structurally and floristically contrasting study areas: a rich deciduous forest and an urban parkland (located c.10 km apart). Both study areas were supplied with standardized wooden nestboxes with a removable front wall (Lambrechts et al. 2010). A 30 mm diameter entrance is located 20 cm from the bottom of the nestbox and internal dimensions are: 11.5 cm (depth) × 11 cm (width) × 30 cm (height).
During all the breeding seasons, the nestboxes were checked at least once a week (or more often if needed) to monitor breeding performance of tits. Parameters measured: clutch size, the number of hatchlings, and the number of fledglings. Hatching success refers to the proportion of eggs resulting in hatchlings and fledging success refers to the proportion of hatchlings resulting in fledglings. Only first clutches were studied.
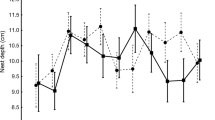
The height (depth) of the nest (as a distance from the nest base to the top rim of the nest cup), following Glądalski et al. (2016a, b), was measured at the end of tit laying stage with a ruler to the nearest 5 mm. Nest height of Eurasian Blue Tits were measured in 183 nests (114 from the parkland site and 69 from the forest site). Nest height of Great Tits was measured in 322 nests (175 from the parkland and 147 from the forest).
After the end of tit breeding season, all nests were collected from the nestboxes and placed in a freezer (− 80 °C) for 24 h in order to sterilize nests by deep freezing. After this procedure, the nests were dried for 24 h in a lab dryer (60 °C) until reaching constant dry mass and then deconstructed (by M.G.) into two basic structural parts: (1) the cup lining and (2) the moss (and each of the part was deconstructed to the basic structural elements, which took up to an hour per nest). The cup lining and the moss layer were weighed for every nest to the nearest 0.1 g (Glądalski et al. 2016a, b, 2021; Table 1). All dead nestlings, feaces, food remnants and all detectable organic powder fragments (skin like dust that is a byproduct of feather growth) were removed. The height of the nest was positively correlated with the nest mass in both Great and Eurasian Blue Tit species, therefore, we used only nest height parameter in the GLM (for Great Tits: r = 0.62, n = 322, p < 0.001, and for Eurasian Blue Tits: r = 0.66, n = 183, p < 0.001).
The effects of nest characteristics on clutch size and brood size at fledging, treated as Poisson variables, were tested using generalized linear models (GLM) with the log-link function (Crawley 2002; Heck et al. 2012). The effects of nest parameters on hatching and fledging successes (hatching success refers to the proportion of eggs resulting in hatchlings, and fledging success refers to the proportion of hatchlings resulting in fledglings, with abandoned broods excluded from the analysis) were examined using GLMs with binomial error distribution and the logit link function (Crawley 2002; Heck et al. 2012). The GLM modeling was performed using IBM SPSS v. 22 software (IBM SPSS Statistics 222013).
Results
The number of fledglings (Table 2) of the Great Tit was positively associated with lining mass and differed between years (Table 3). Hatching success in Great Tits was positively associated with lining mass. In Eurasian Blue Tits, clutch size and the number of fledglings were positively associated lining mass (Table 3). Hatching success in Eurasian Blue Tits was also positively associated with lining mass (Table 4). Clutch size differed between urban parkland and a forest study area in Great Tits (being higher in the forest) and the number of fledglings differed between urban parkland and a forest study area in both Great and Eurasian Blue Tits being higher in the forest. In the Great Tit, hatching success and fledging success were higher in the forest and in the Eurasian Blue Tit fledging success was also higher in the forest. Hatching success and fledging success differed between years in both Great and Eurasian Blue Tit species (Table 4).
Discussion
We showed that in Great and Eurasian Blue Tits the number of fledglings were positively associated with lining mass and were affected by the study area (being higher in the forest). In Eurasian Blue Tits, clutch size tended to be associated with lining mass. There was also a relationship between lining mass and higher hatching success in both tit species.
Nest characteristics (like size, mass or structure) were shown before to be positively related to reproductive parameters, such as clutch size, hatching and fledging success or recruitment success, as well as quality and condition of the nest-building female (Heenan 2013; Deeming and Pike 2015; Mainwaring 2017; Stenning 2018; Deeming 2023). It was also suggested that the most important function of the nest is thermal insulation, especially during egg incubation by the female and during the first week of the hatchlings’ life when they are ectothermic (Perrins 1979). Nest material buffers ambient temperature fluctuations and helps to maintain egg temperature and humidity (Mertens 1977; Deeming 2011). Several authors reported that tits are characterised by a selective behaviour in their choice of building elements collected for nest construction (Wesołowski and Wierzcholska 2018; Järvinen and Brommer 2020; Glądalski et al. 2021; Fäth et al. 2023) and recently Deeming et al. (2020) showed that, by selecting particular materials whilst constructing a nest, bird can manipulate the thermal characteristics of the nest. Also Deeming and Pike (2015) showed on tits that the insulatory properties of nests during incubation can correlate with offspring fitness. Therefore, in many recent studies, authors were linking a lining (nest) mass with breeding success as a natural consequence (Heenan 2013; Mainwaring 2017; Stenning 2018). But after almost 2 decades of studies increasingly focusing on nests, some evidence is available that simple links may exist between lining mass and breeding success, yet not so strong as presumed before. For example, some of the authors did not find any effects (or effects were rather weak) of nest characteristics on breeding performance (Tomás et al. 2006; Lambrechts et al. 2016; for experimental study, see Lambrechts et al. 2012). Some authors suggested that there might be effects (between nest characteristics and breeding performance) in some years but not in other years (e.g., Britt and Deeming 2011). In addition, Harnist et al. (2020) showed on Great Tits that nest mass increases over the course of the nesting cycle (and basically the vast majority of the nest studies in tit literature is based on the post-season collection of the nests, similar findings were shown by Britt and Deeming 2011). Between the incubation and post-fledging stages the authors recorded an increase in fresh mass of the nest by nearly 70% (some of other studies reported about 9%, see Cruz et al. 2016). The change in mass between both stages is probably caused by the accumulation of organic powder, food remnants and feaces (Harnist et al. 2020). Authors also suggest that a better method of quantifying the nest size is measuring of the nest height/height, but it is not perfect because nest height may differ between different sizes of nestboxes and between various populations, it may also differ due to predator attacks and many other factors (Kaliński et al. 2014). Recently, Marcel Lambrechts after reading Harnist et al. (2020) suggested that it is probable that skin produced by nestlings accumulate inside the lining part and it can make the lining part of the nest with a higher fledging success or a larger number of nestlings heavier (Lambrechts, public information, 2021). Therefore, it seems reasonable that if hatching success in many studies (including ours) refers to the proportion of eggs resulting in hatchlings—more hatchings may deposit more organic powder in the lining layer of the nest and cause higher factor of hatching success (in other words and more general: organic powder may be correlated with hatching success/number of fledglings/clutch size and not with the mass of the lining). This hypothesis should be tested in the near future using lining and moss from the fresh nests. In the present study all the dead nestlings, feaces, food remnants and all distinguishable organic powder fragments were removed from the nests but it is important to emphasize that indeed some of the powder was tightly bound with the fragments of lining, which were simply impossible to remove (Glądalski, personal observation)—that could at least partly support Harnist et al. (2020) suggestion. In other words, although our results indicated a positive correlation between lining mass in the nest and hatching success, it is nonetheless possible that a greater proportion of nestlings produce a greater quantity of organic debris which consequentially increased lining mass.
Harnist et al. (2020) suggest also that the nest height/depth is a reliable method of quantifying the characteristics of the nest. Recently Lambrechts and Caro (2022) showed experimentally that egg cooling is associated with nest size in Great Tits—eggs cooled more quickly in thinner nests and when ambient temperature was lower. In southern France (Mediterranean region), the temperatures during breeding period are relatively high and stable and this effect (of faster egg cooling) may not occur or be just negligible, but in colder parts of Europe, like Poland, it could be important. During the breeding season in Poland, we have witnessed large drops in the mean ambient temperature (while tits were incubating), and during some breeding seasons even, to about 2 °C to 3 °C (Glądalski et al. 2020b). Therefore, a larger nest may be probably beneficial for the female (less heat loss) but it also may be beneficial for the nestlings because incubation temperature affects growth and energy metabolism in tit nestlings (Nord and Nilsson 2011). In Glądalski et al. (2016a), we showed that nest height (depth) positively affected fledging success in Great Tits, but with a larger set of data the effect disappeared in the present paper (and there is no effect for both tit species). The reason may be that in a large set of different seasons with various weather characteristics, the positive effect of nest size may be lost, but the reason may also be that the amount of moss (which dominates the nest mass/height) is mainly related to individual variability and does not affect fitness as suggested by Sonnenberg et al. (2020).
We also showed that both clutch size and number of fledglings were higher in both tit species in the forest and those results are in line with our earlier results on both tit species (for Eurasian Blue Tits Glądalski et al. 2015, for Great Tits Wawrzyniak et al. 2015). The site factor also affected hatching success (for Great Tits higher in the forest) and fledging success (for Eurasian Blue Tits higher in the forest) (for Eurasian Blue Tits Glądalski et al. 2017, for Great Tits Wawrzyniak et al. 2020). The main reason for the differences is probably food availability. The total number of native oaks (Pedunculate Oak Quercus robur and Sessile Oak Quercus petraea) per nestbox plot was 19 times greater in the forest than in the parkland (for the forest—65.0 trees / nestbox plot and for the parkland—3.4 trees/nestbox plot, Glądalski et al. 2017). In addition, both study areas differ in the abundance of caterpillars (considered as the most suitable kind of food for nestlings), as the differences in the amount of caterpillar frassfall suggests, with the parkland being characterized by ca. two to three times lower abundance of caterpillars than the forest area (Nadolski et al. 2021). Reduced hatching success in the urban parkland may be related to feeding and incubation rhythm disturbances. Exposure to human visitors in some birds may disturb the regularity of incubating and feeding nestlings by parents and may affect breeding success (Ruiz et al. 2002). The parkland site is one of the largest recreation and entertainment areas in Łódź and the number of visitors is large compared to the forest (Glądalski et al. 2016b). We previously showed that non-lethal disturbance events from humans could influence breeding success negatively in the parkland study site, especially during sunny and warm days, when the numbers of visitors are high (Glądalski et al. 2016b).
We also showed that hatching success and fledging success differed between years in both Great and Eurasian Blue Tits. As we showed in Glądalski et al. (2017), one of the most important components of the year factor in are food availability and weather conditions (precipitations, temperature, extreme weather events). The abundance of caterpillars (considered as the most suitable kind of food for nestlings) differs not only between habitats but also differs largely between years as shown for our study areas in a long-term study by Nadolski et al. (2021). Unfavorable weather conditions (like low temperatures of heavy rain in longer periods of time) may cause tits (and other birds) breed later and in lower numbers, lay smaller clutches, increase incubation bout length and have much lower breeding success (Bordjan and Tome 2014; Glądalski et al. 2017; Stenning 2018).
Breeding success is influenced by a large cocktail of factors and nest parameters are only small part of that mosaic of factors. There is no doubt that avian nests are important to reproduction and materials used to construct nests contribute to many and miscellaneous functional properties like insulation, control of humidity or weather-proofing, but new nest-studies, especially experimental are to be welcome.
Data availability
Data are available from the author on request.
References
Alabrudzińska J, Kaliński A, Słomczyński R, Wawrzyniak J, Zieliński P, Bańbura J (2003) Effects of nest characteristics on breeding success of Great Tits Parus major. Acta Ornithol 38:151–154
Álvarez E, Barba E (2008) Nest quality in relation to adult bird condition and its impact on reproduction in Great Tits Parus major. Acta Ornithol 43:3–9
Álvarez E, Barba E (2011) Nest characteristics and reproductive performance in Great Tits Parus major. Ardeola 58:125–136
Álvarez E, Belda EJ, Verdejo J, Barba E (2013) Variation in Great Tit nest mass and composition and its breeding consequences: a comparative study in four Mediterranean habitats. Avian Biol Res 6:39–46
Bordjan D, Tome D (2014) Rain may have more influence than temperature on nest abandonment in the Great Tit Parus major. Ardea 102:79–85
Britt J, Deeming DC (2011) First-egg date and air temperature affect nest construction in Blue Tits Cyanistes caeruleus, but not in Great Tits Parus major. Bird Study 58:78–89
Crawley MJ (2002) Statistical computing: An introduction to data analysis using S-Plus. Wiley, Chichester
Cruz A, Álvarez E, Barba E (2016) Nest insulating capacity during incubation and after fledging are related. Avian Biol Res 9:22–27
Dawkins R (1982) The extended phenotype: long reach of the gene. Oxford University Press, Oxford
Deeming DC (2011) Importance of nest type on the regulation of humidity in bird nests. Avian Biol Res 4:23–31
Deeming DC (2023) A review of the roles materials play in determining functional properties of bird nests. Acta Ornithol 58:1–28
Deeming DC, Pike TW (2015) Nest surface temperature predicts fledging success of blue tits Cyanistes caeruleus but not great tits Parus major. Acta Ornithol 50:247–251. https://doi.org/10.3161/00016454AO2015.50.2.012
Deeming DC, Dickinson AM, Broughton RE, Locke E, Gray LA, Bennett S, Gilchrist R, Muniz S, Goodman AM, Biddle LE (2020) Factors affecting thermal insulation of songbird nests as measured using temperature loggers. Physiol Biochem Zool 93:488–504
Fäth L, Obermaier E, Pfeifer R (2023) Picky blue tits—selective moss utilization for nest construction in Cyanistes caeruleus. Ornithol Anz 61:1–15
Glądalski M, Bańbura M, Kaliński A, Markowski M, Skwarska J, Wawrzyniak J, Zieliński P, Cyżewska I, Bańbura J (2015) Inter-annual and inter-habitat variation in breeding performance of Blue Tits (Cyanistes caeruleus) in central Poland. Ornis Fennica 92:34–42
Glądalski M, Bańbura M, Kaliński A, Markowski M, Skwarska J, Wawrzyniak J, Zieliński P, Cyżewska I, Bańbura J (2016a) Effects of nest characteristics on reproductive performance in blue tits Cyanistes caeruleus and Great Tits Parus major. Avian Biol Res 9:37–43. https://doi.org/10.3184/175815516X14447556559088
Glądalski M, Bańbura M, Kaliński A, Markowski M, Skwarska J, Wawrzyniak J, Zieliński P, Cyżewska I, Mańkowska D, Bańbura J (2016b) Effects of human-related disturbance on breeding success of urban and non-urban Blue Tits (Cyanistes caeruleus). Urban Ecosyst 19:1325–1334. https://doi.org/10.1007/s11252-016-0543-3
Glądalski M, Bańbura M, Kaliński A, Markowski M, Skwarska J, Wawrzyniak J, Zieliński P, Cyżewska I, Bańbura J (2017) Differences in the breeding success of Blue Tits Cyanistes caeruleus between a forest and an urban area: a long-term study. Acta Ornithol 52:59–68. https://doi.org/10.3161/00016454AO2017.52.1.006
Glądalski M, Bańbura M, Kaliński A, Markowski M, Skwarska J, Wawrzyniak J, Zieliński P, Bańbura J (2018) Hatching delays in great tits and blue tits in response to an extreme cold spell: a long-term study. Int J Biometeorol 62:1437–1445
Glądalski M, Bańbura M, Kaliński A, Markowski M, Skwarska J, Wawrzyniak J, Zieliński P, Bańbura J (2020a) Consequences of experimental addition of fresh, aromatic plants into nests of Blue Tits (Cyanistes caeruleus) on the physiological condition of nestlings. Behav Ecol Sociobiol 74:29. https://doi.org/10.1007/s00265-020-2812-7
Glądalski M, Bańbura M, Kaliński A, Markowski M, Skwarska J, Wawrzyniak J, Zieliński P, Bańbura J (2020b) Extreme temperature drop alters hatching delay, reproductive success and physiological condition in Great Tits. Int J Biometeorol 64:623–629. https://doi.org/10.1007/s00484-019-01851-6
Glądalski M, Kaliński A, Markowski M, Skwarska J, Wawrzyniak J, Bańbura J (2024) Nest size parameters of Great Tits and Blue Tits: a long-term study. J Ornithol 165:381–389. https://doi.org/10.1007/s10336-023-02106-3
Hansell M (2000) Bird nest and construction behaviour. Cambridge University Press, Cambridge
Harnist I, Dubiec A, Mazgajski TD (2020) Changes of nest mass in relations to nesting stages in the Great Tit Parus major. Bird Study 67:292–299
Heck RH, Thomas SL, Tabata LN (2012) Multilevel modelling of categorical outcomes using IBM SPSS. Routledge, New York
Heenan CB (2013) An overview of the factors influencing the morphology and thermal properties of avian nests. Avian Bioly Res 6:104–118
Heenan CB, Seymour RS (2011) Structural support, not insulation, is the primary driver for avian cup-shaped nest design. Proc R Soc B 278:2924–2929
IBM SPSS Statistics 22 (2013) SPSS for windows release 22.0. Armonk: IBM Corporation
Järvinen P, Brommer JE (2020) Lining the nest with more feathers increases offspring recruitment probability: selection on an extended phenotype. Ecol Evol 10:13327–13333. https://doi.org/10.1002/ece3.6931
Kaliński A, Wawrzyniak J, Bańbura M, Skwarska J, Zieliński P, Glądalski M, Bańbura J (2014) Does the threat of European Pine Marten (Martes martes) predation influence the height of nests built by Blue Tits (Cyanistes caeruleus) and Great Tits (Parus major)? Avian Biol Res 7:83–90
Lambrechts ML, Caro SP (2022) Egg cooling associated with nest size in a passerine bird. J Therm Biol 110:103383. https://doi.org/10.1016/j.jtherbio.2022.103383
Lambrechts M, Adriaensen F, Ardia DR, Artemyev AV, Atiénzar F, Bańbura J et al (2010) The design of artificial nestboxes for the study of secondary hole-nesting birds: a review of methodological inconsistencies and potential biases. Acta Ornithol 45:1–26
Lambrechts M, Midamegbe A, Galan M-J, Perret P, Grégoire A, Doutrelant C (2012) Nest size and breeding success in first and replacement clutches: an experimental study in Blue Tits Cyanistes caeruleus. J Ornithol 153:173–179
Lambrechts MM, Marrot P, Fargevieille A, Giovannini P, Lucas A, Demeyrier V, Midamegbe A, Perret P, Grégoire A, Charmantier A, Doutrelant C (2016) Nest size is not closely related to breeding success in Blue Tits: a long-term nest-box study in a Mediterranean oak habitat. Auk 133:198–204
Lambrechts MM, Blondel J, de Franceschi C, Doutrelant C (2017) Nest size is positively correlated with fledging success in Corsican Blue Tits (Cyanistes caeruleus) in an insular-oak-dominated habitat mosaic. J Ornithol 158:125–132. https://doi.org/10.1007/s10336-016-1377-8
Lambrechts M (2021) https://www.researchgate.net/profile/Marcel-Lambrechts/answers; public information.
Mainwaring MC (2017) Causes and consequences of intraspecific variation in nesting behaviors: insights from Blue Tits and Great Tits. Front Ecol Evol 5:39. https://doi.org/10.3389/fevo.2017.00039
Mainwaring MC, Hartley IR (2008) Seasonal adjustments in nest cup lining in Blue Tits Cyanistes caeruleus. Ardea 96:278–282. https://doi.org/10.5253/078.096.0213
Mainwaring MC, Hartley IR, Lambrechts MM, Deeming DC (2014) The design and function of birds’ nests. Ecol Evol 20:3909–3928
Manly BJF (1986) Multivariate statistical methods. Chapman and Hall, London
Maziarz M, Broughton RK, Wesołowski T (2017) Microclimate in tree cavities and nest-boxes: implications for hole-nesting birds. Forest Ecol Manag 389:306–313
Mertens JAL (1977) Thermal conditions for successful breeding in Great Tits (Parus major L.). Oecologia 28:1–29
Nadolski J, Marciniak B, Loga B, Michalski M, Bańbura J (2021) Long-term variation in the timing and height of annual peak abundance of caterpillars in tree canopies: some effects on a breeding songbird. Ecol Indic 121:107–120. https://doi.org/10.1016/j.ecolind.2020.107120
Nord A, Nilsson J-Å (2011) Incubation temperature affects growth and energy metabolism in Blue Tit nestlings. Amer Natur 178:639–651
Nuhlíčková S, Svetlík J, Eckenfellner M, Knauer F, Hoi H (2021) Interaction between nestling behaviour and nest-space use. Ethol Ecol Evol 33:496–514. https://doi.org/10.1080/03949370.2020.1858173
Perrins CM (1979) British tits. Collins, London
Ruiz G, Rosenmann M, Novoa FF, Sabat P (2002) Hematological parameters and stress index in Rufous-collared Sparrows dwelling in urban environments. Condor 104:162–166
Rydgren K, Indreeide B, Slagsvold T, Lampe HM (2023) Nest building in titmice Paridae: selectivity in bryophyte use. Ecol Evol 13:e9852. https://doi.org/10.1002/ece3.9852
Soler JJ, Møller AP, Soler M (1998) Nest building, sexual selection and parental investment. Evol Ecol 12:427–441
Sonnenberg BR, Branch CL, Benedict LM, Pitera AM, Pravosudov VV (2020) Nest construction, ambient temperature and reproductive success in a cavity-nesting bird. Anim Behav 165:43–58
Stenning M (2018) The blue tit. T & AD Poyser, London
TIBCO Statistica® 13.3.0 (2017) Data analysis software system, version 13.3.0, URL: http://www.tibco.com
Tomás G, Merino S, Moreno J, Sanz JJ, Morales J, García-Fraile S (2006) Nest weight and female health in the Blue Tit (Cyanistes caeruleus). Auk 123:1013–1021
Vanadzina K, Street SE, Healy SD, Laland KN, Sheard C (2023) Global drivers of variation in cup nest size in passerine birds. J Anim Ecol 92:338–351
Wawrzyniak J, Kaliński A, Glądalski M, Bańbura M, Markowski M, Skwarska J, Zieliński P, Cyżewska I, Bańbura J (2015) Long-term variation in laying date and clutch size of the Great Tit Parus major in central Poland: a comparison between urban parkland and deciduous forest. Ardeola 62:311–322
Wawrzyniak J, Glądalski M, Kaliński A, Bańbura M, Markowski M, Skwarska J, Zieliński P, Bańbura J (2020) Differences in the breeding performance of Great Tits Parus major between a forest and an urban area: a long term study on first clutches. Eur Zool J 87:294–309
Wesołowski T, Wierzcholska S (2018) Tits as bryologists: patterns of bryophyte use in nests of three species cohabiting a primeval forest. J Ornithol 159:733–745
Wolski GJ, Sadowska B, Fol M, Podsędek A, Kajszczak D, Kobylińska A (2021) Cytotoxicity, antimicrobial and antioxidant activities of mosses obtained from open habitats. PLoS ONE 16:e0257479. https://doi.org/10.1371/journal.pone.0257479
Acknowledgements
All procedures were approved by the Local Ethical Committee and the State Office for Environment Protection. We thank A. Jaksa, M. Winsche, D. Mańkowska, and J. Białek for their help and consent to conducting research in the areas under their administration. We thank J. Sinclair for linguistic consultation.
Funding
The study was founded by University of Lodz (No. 506/1145).
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare no conflict of interest.
Ethical statement
Bird catching and handling was conducted according to Polish laws. All procedures were approved by the Local Ethical Committee and the State Office for Environment Protection."
Additional information
Communicated by F. Bairlein.
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Glądalski, M., Bańbura, M., Kaliński, A. et al. Nest structure and breeding success in two tit species: a long-term study. J Ornithol (2024). https://doi.org/10.1007/s10336-024-02193-w
Received:
Revised:
Accepted:
Published:
DOI: https://doi.org/10.1007/s10336-024-02193-w