
Abstract
Bird species depending on seeds from masting trees face large temporal and spatial variation in food availability and perform nomadic or irruptive movements to locate food. In northern Europe, the Common Crossbill (Loxia curvirostra) is specialized on seeds from Norway Spruce (Picea abies) which usually has peak years followed by crop failure. Common Crossbills breed in mast years and emigrate when spruce seeds are no longer available. On the other hand, the Parrot Crossbill (L. pytyopsittacus) is specialized on seeds from Scots Pine (Pinus sylvestris) in which seed production is more stable from year to year. The Parrot Crossbill is therefore thought to be more sedentary. However, both species may switch food if there is low abundance of their 'own' seed type, but little is known about the relative importance of crop size of the two tree species on breeding population size and migration of the two crossbill species. Here, we analyse time series (up to 36 years) of crossbills and crop size of spruce and pine in Norway and Sweden to investigate this. We found that breeding population sizes of both species were positively related to spruce crop size. Surprisingly, pine crop size had no positive influence, not even on the Parrot Crossbill. Emigration of both crossbill species (based on autumn migration counts in southern Sweden) increased when there were small seed crops of spruce, but pine crop size had no influence, not even on the Parrot Crossbill. There was no influence of population size on emigration. These results suggest that alternative food resources had an asymmetric effect, only influencing the Parrot Crossbill. This asymmetry explains why the two crossbill species often have synchronous eruptions despite being specialized on different tree species with different seed crop size dynamics.
Zusammenfassung
Schwankungen in der Populationsgröße und im Wanderverhalten von zwei Kreuzschnabelarten in Abhängigkeit von der Samenmenge von Fichten und Kiefern: asymmetrische Bedeutung alternativer Nahrungsressourcen
Vogelarten, die auf Samen von Mastbäumen angewiesen sind, werden mit großen zeitlichen und räumlichen Schwankungen in der Nahrungsverfügbarkeit konfrontiert und führen somit nomadische oder unregelmäßige Wanderbewegungen zur Nahrungssuche durch. In Nordeuropa hat sich der Fichtenkreuzschnabel (Loxia curvirostra) auf die Samen der Fichte (Picea abies) spezialisiert, die in der Regel Mastjahre hat, auf die eine geringere Samenproduktion folgt. Der Fichtenkreuzschnabel brütet in den Mastjahren und wandert ab, wenn keine Fichtensamen mehr zur Verfügung stehen. Der Kiefernkreuzschnabel (L. pytyopsittacus) hingegen ist auf die Samen der Waldkiefer (Pinus sylvestris) spezialisiert, deren Samenproduktion über die Jahre stabiler ist. Daher wird der Kiefernkreuzschnabel für sesshafter gehalten. Beide Arten können jedoch auf andere Nahrungsquellen ausweichen, wenn ihre „eigene“ Samenart nicht in ausreichender Menge vorhanden ist. Über die relative Bedeutung der Samenmenge der beiden Baumarten für die Populationsgröße und das Wanderverhalten der beiden Kreuzschnabelarten ist jedoch wenig bekannt. Um dies zu untersuchen, analysieren wir hier Zeitreihen (bis zu 36 Jahren) von Kreuzschnabelvorkommen und Samenmengen der Fichte und Kiefer in Norwegen und Schweden. Wir fanden heraus, dass die Populationsgrößen beider Arten positiv mit der Menge an Fichtensamen zusammenhängen. Überraschenderweise hatte die Menge der Kiefernsamen jedoch keinen positiven Einfluss, nicht einmal auf den Kiefernkreuzschnabel. Die Abwanderung beider Kreuzschnabelarten (basierend auf Zählungen während des Herbstzuges in Südschweden) nahm zu, wenn es geringere Mengen an Fichtensamen gab; aber die Kiefernsamenmenge hatte keinen Einfluss, auch nicht auf den Kiefernkreuzschnabel. Es wurde jedoch kein Effekt der Populationsgröße auf die Abwanderung beobachtet. Diese Ergebnisse lassen vermuten, dass alternative Nahrungsressourcen einen asymmetrischen Effekt hatten, der sich nur auf die Kiefernkreuzschnäbel auswirkte. Diese Asymmetrie erklärt, warum die beiden Kreuzschnabelarten häufig synchron auftreten, obwohl sie auf verschiedene Baumarten mit unterschiedlicher Dynamik der Samenproduktion spezialisiert sind.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
A number of bird species in boreal forests depend on seeds from one or more tree species. However, seed production of some tree species varies enormously from year to year where peak seed crop years are often followed by years with very poor crops (called masting; Silvertown 1980; Kelly 1994; Koenig and Knops 2000). In addition, masting is usually spatially synchronous over distances of hundreds or thousands of kilometers (Koenig and Knops 2000; LaMontagne et al. 2020). Thus, these seed-eating birds face large temporal and spatial variations in food abundance, and may deal with this by performing nomadic or irruptive movements (Bock and Lepthien 1976; Koenig and Knops 2001; Newton 2006a, 2008). Individual birds may therefore breed or winter in locations that are up to thousands of kilometers apart in different years (Newton 2006a, 2008, 2012). The movements may be triggered by lack of food where they come from (push factor; Svärdson 1957; Bock and Lepthien 1976), but likely also drawn towards more abundant food in the areas they move to (pull factor; Strong et al. 2015; Dale 2023). Emigration may also be due to the combination of food shortage and high population density due to high reproduction in the peak seed crop year (Lack 1954; Koenig and Knops 2001).
Crossbills (Loxia spp.) are extreme examples of food specialization where seeds from conifer trees form the diet most of the year. Adaptations in particular regarding bill morphology has led to resource specialization between species (Benkman 1987a) and even between populations within a species (Benkman 1993; Newton 2008), thereby illustrating the importance of feeding specializations for adaptive radiation (Benkman 2003). In Europe, two crossbill species are specialized on the seeds of different tree species (Newton 1972; Cramp and Perrins 1994): the Common Crossbill (L. curvirostra) feeds mainly on seeds of Norway Spruce (Picea abies) whereas the Parrot Crossbill (L. pytyopsittacus) has a larger bill and is better able to handle the tougher cones of Scots Pine (Pinus sylvestris). In northern Europe, these two trees are the main conifer species present. The spruce and the pine differs in that the former displays typical masting whereas seed production in the pine is less variable between years (Lindén et al. 2011; Nussbaumer et al. 2016). Thus, Common Crossbills show extraordinary movement patterns and large spatial and temporal variation in population size which follow fluctuations in spruce seed crop size (Reinikainen 1937; Svärdson 1957; Newton 1972; Förschler et al. 2006). Successive breeding sites are often located thousands of kilometers apart (Newton 1972, 2006b; Marquiss et al. 2012). On the other hand, the Parrot Crossbill is considered to be more sedentary with shorter and less frequent movements (Alerstam 1982; Cramp and Perrins 1994), although there have been no quantitative studies of this species.
However, although the two European crossbill species are specialized to handle the cones of different tree species, both species may switch to the other tree species when their 'own' tree has poor seed production (Reinikainen 1937; Alerstam 1982; Cramp and Perrins 1994). An alternative idea is that use of other tree species occurs when they provide abundant food (Benkman 1987b), irrespective of the crop size of their 'own' tree species. In any case, it is therefore possible that breeding and migration may be influenced by seed crop size of both tree species. Eruptions of crossbill species are often synchronous (Newton 1972; Alerstam 1982; Cramp and Perrins 1994), and Alerstam (1982) suggested that this is because Parrot Crossbills are able to survive on spruce seeds and emigrate only when also the spruce crop size is poor, thereby coinciding with Common Crossbill emigration. Massive emigration may therefore occur in particular when both tree species have poor crops, whereas high breeding density may occur when both seed crops are good. However, these ideas have not been tested quantitatively.
Here, we analyse how crossbill breeding densities in Norway and Sweden and migration away from these countries are influenced by yearly variation in seed crop size of both spruce and pine. We predict that breeding population size is positively related to seed crop size, and that breeding population size is highest when both spruce and pine have good crops. Similarly, we predict that emigration is related to seed crop failure, and that emigration of both species is highest when both spruce and pine have poor crops. Furthermore, we predict that emigration is also related to breeding population size so that the largest eruptions occur when there is seed crop failure in combination with high breeding population size. These predictions are tested with time series of crossbill breeding numbers and migration, and crop size of spruce and pine, from Norway and Sweden. We acknowledge that distinguishing the two crossbill species in the field may be challenging, and we therefore provide detailed accounts of the data sources (see Methods) and a thorough evaluation of the reliability of the data (see Discussion), and conclude that the results are likely to reflect real patterns.
Methods and materials
Study area and study species
In this study, time series of crossbill numbers and seed crop sizes from Norway and Sweden are used (Table 1, see below for details). The Parrot Crossbill is distributed in the boreal zone from northwestern Europe and east to the Ural mountains, whereas the Common Crossbill has a wider distribution, both further south and east, and also including North America (Cramp and Perrins 1994). Both crossbill species breed over most parts of both Norway and Sweden. Population size of the Common Crossbill is on average approximately six times larger than that of the Parrot Crossbill in both Norway and Sweden, but both species fluctuate considerably from year to year (approximate population estimates of Common Crossbill; Norway: 15,000–150,000 pairs, Sweden: 58,000–580,000 pairs; Parrot Crossbill; Norway: 2500–25,000 pairs, Sweden: 10,000–100,000 pairs; Shimmings and Øien 2015; Ottosson et al. 2012). The ranges of the estimates may reflect both actual population fluctuations and uncertainties of estimates. Although the Common Crossbill is known to have several ecotypes that differ in bill size and calls and that are associated with different conifer species both in North America (Benkman 1993) and parts of Europe, especially in the Mediterranean (Newton 2008), no differentiation into ecotypes is known from Norway or Sweden.
Spruce and pine are also widespread across both countries except that spruce has few natural populations in western Norway, but is widely planted there. Most of the forests in Norway and Sweden are in the boreal vegetation zone where spruce and pine are dominant tree species. The spruce flowers in early summer and crossbills can extract seeds from July and through autumn and winter until the cones open and seeds are shed during late winter the next year (Newton 1972; Alerstam 1982), although seed shedding may sometimes occur in part during autumn. The main breeding season of Common Crossbills is during February–April (Haftorn 1971; Newton 1972; Alerstam 1982; Cramp and Perrins 1994). Because few or no spruce flowers are produced after a masting year, crossbills will not have access to a new good spruce crop in the same area until at least one year (but usually several years) after the previous crop was shed. Thus, migration of Common Crossbills from one breeding area to the next usually starts in May and may continue until autumn (Haftorn 1971; Newton 1972; Alerstam 1982; Cramp and Perrins 1994).
On the other hand, the Scots Pine has a reproductive cycle of two years (Owens and Blake 1985). After flowering and pollination during spring–early summer in the first growing season, fertilization and seed development takes place during summer in the second growing season. The seeds are available to crossbills thereafter, generally from September–October until they are shed in spring and early summer in the year after they ripened (Newton 1972; Alerstam 1982). The main breeding season of Parrot Crossbills is during February–June (Haftorn 1971; Newton 1972; Alerstam 1982; Cramp and Perrins 1994) which is later than for the Common Crossbill because pine seeds are available for longer during spring and early summer. If food shortage or other factors cause migration after breeding, this takes place from late summer and through autumn (Haftorn 1971; Newton 1972; Alerstam 1982; Cramp and Perrins 1994), again later than for Common Crossbills.
Swedish crossbill breeding numbers
Data on crossbill breeding numbers for Sweden were taken from the Swedish Bird Survey (www.fageltaxering.lu.se; Green et al. 2021). Results were based on 716 fixed routes spread across Sweden and surveyed once per year during the breeding season of birds in general (May–July). Data were available for the period 1998–2021 (n = 24 years). Breeding numbers were expressed as an index where 1998 was set to a value of 1.0. Because breeding of crossbills is dependent on food availability (Newton 1972; Cramp and Perrins 1994), presence during the main breeding season does not necessarily imply actual breeding, and numbers reported could include non-breeders in particular during years with low numbers of crossbills and low food availability.
Reports included separate data for Common and Parrot Crossbill (average numbers seen per year was 1228 and 272, respectively), but also included 'unidentified crossbills' (average 1485). Unidentified crossbills were correlated to the separate indices for both species, but was best explained by Common Crossbill indices (GLM, centred and scales predictors; Common Crossbill: estimate = 3.45, SE = 0.63, p < 0.001, Parrot Crossbill: estimate = 1.45, SE = 0.63, p = 0.022). Thus, unidentified crossbills were likely to a large degree Common Crossbills. In any case, we used the species-specific indices for analyses. Breeding numbers of both species increased over the study period, although significantly only for the Parrot Crossbill (Pearson correlations; Common Crossbill: r = 0.34, p = 0.12, Parrot Crossbill: r = 0.66, p = 0.002).
The Swedish Bird Survey also includes winter counts (Green et al. 2021). In this case, counts are conducted in areas selected by the observers, so unlike the breeding bird survey described above, the counts are not spatially representative for the whole of Sweden, and have a strong bias to the southern half of Sweden. Counts were conducted during October–March, thus to some degree overlapping with the breeding season of crossbills. The winter counts were considered to represent to a large degree pre-breeding numbers, and because crossbills do not perform regular migration during most of this period the winter counts may therefore be an alternative indicator of breeding numbers. Data were available for the winters 1998/99–2020/21 (n = 23 years). The winter count 1998/99 was taken as an indication of breeding numbers for 1999 and so on. Numbers were expressed as an index where 1998/99 was set to a value of 1.0. On average, winter counts totalled 309 Common Crossbills, 36 Parrot Crossbills, and 570 unidentified crossbills per year. As for the breeding numbers above, the species-specific indices were used for analyses. Winter counts increased during the study period for both species, although significantly only for the Parrot Crossbill (Common Crossbill: r = 0.17, p = 0.44, Parrot Crossbill: r = 0.44, p = 0.035).
Population fluctuations in a more restricted area were also analysed to avoid large-scale asynchrony in peak and low years. For this purpose the number of records submitted to the Swedish Species Observation System (www.artportalen.se) were extracted. Most reports of birds are submitted by members of BirdLife Sweden and other birders. The unit used for analyses was a 'record', i.e. one or more individuals observed in one place at one time (termed 'fynd'). Records from parts of southern Sweden were used (the regions Värmland, Västmanland, Uppland, Södermanland, Närke, Dalsland, Bohuslän, Halland, Västergötland, Östergötland and Småland). The choice of regions was based on regions showing similar fluctuations in spruce crop size (see below). For the Common Crossbill, all records during February–April (main breeding season) in the period 2001–2021 (n = 21 years) were used. There were relatively few records in the years before 2001. Observations from May and later were excluded to avoid the post-breeding influx of birds from distant populations which typically starts during May–June. For the Parrot Crossbill, post-breeding movements start later and records during February–June which reflect the breeding season were therefore used. Number of records of both species increased over the study period, most likely due to increased reporting frequency by birders over time (Common Crossbill: r = 0.41, p = 0.07, Parrot Crossbill: r = 0.71, p < 0.001).
Norwegian crossbill breeding numbers
For Norway, crossbill breeding numbers were taken from the Norwegian Breeding Bird Survey (Norsk hekkefuglovervåking) (www.hekkefuglovervakingen.nina.no; Kålås et al. 2019). Results from 492 fixed routes spread across Norway, of which at least 80% were surveyed each year during the general bird breeding season (May–July), in the period 2008–2021 were used (n = 14 years). Breeding numbers were expressed as an index where 2008 was set to a value of 1.0. There was enough data to calculate yearly indices for the Common Crossbill, but not for the Parrot Crossbill. The average abundance of Common Crossbills recorded per year has not been specified, but is likely in the order of 200 pairs per year. Breeding numbers of the Common Crossbill in Norway did not show a long-term trend (r = 0.13, p = 0.67).
Because data on Parrot Crossbills were not available from the Norwegian Breeding Bird Survey, crossbill numbers based on records submitted to the National Biodiversity Information Centre in Norway (www.artsobservasjoner.no) were extracted for both species. Most reports of birds are submitted by members of BirdLife Norway and other birders. The unit used for analyses was a 'record', i.e. one or more individuals observed in one place at one time (termed 'funn'), similar to the Swedish Species Observation System because both Norway and Sweden use the same data base platform. Records from all regions of Norway were used, and for the same months of the year as described above for Sweden. Data were extracted for the period 2008–2021 (n = 14 years) because there were relatively few records in the years before 2008. Number of records of in particular Parrot Crossbill increased over the study period, again most likely due to increased reporting frequency by birders over time (Common Crossbill: r = 0.20, p = 0.50, Parrot Crossbill: r = 0.55, p = 0.042).
Crossbill migration data
Data on emigration of crossbills from northern Europe was based om migration counts from Falsterbo bird station at the southwestern tip of Skåne, southernmost Sweden (www.falsterbofagelstation.se). Birds migrating south during autumn head for Denmark and Germany. Daily counts during the period 1 August–20 November were conducted each year (except that they were from 11 August until 2000), and data on both crossbill species were available for the period 1986–2021 (n = 36 years; www.falsterbofagelstation.se/strack/art-alla-ar/). During most of these years all crossbills were identified to species, and 'unidentified crossbills' were only recorded in some years during 1986–1999. In nine years with at least 10 unidentified crossbill individuals recorded, they constituted on average 22% of all crossbills recorded (median 19%). In one additional exceptional year 19 unidentified crossbills were recorded when no crossbills were identified. Furthermore, the mean number of Common Crossbills across all 36 years was 3974, for Parrot Crossbills 804, and for unidentified crossbills 164. We conclude that unidentified crossbills constituted a small proportion and were therefore excluded, and only numbers for identified crossbills were used in analyses. Migration counts of neither species showed long-term changes (Common Crossbill: r = 0.18, p = 0.30, Parrot Crossbill: r = 0.19, p = 0.26).
Spruce and pine seed crop size
In Sweden, seed crop size estimation was conducted by Skogforsk (Forestry Research Institute of Sweden; www.skogforsk.se) by counting the number of cones on sample trees at a large number of sites across the whole country (Riksskogstaxeringen). Although the number of cones is comparable between spruce and pine, it should be noted that medium-sized spruce cones usually contain 100–200 filled seeds whereas medium-sized pine cones usually contain only 25–30 filled seeds (Fystro 1962). Each year cones on ca. 2000 trees (range ca. 1000–3000) of spruce and similar numbers of pine trees were recorded. Average values across individual trees were reported for each of 31 regions and for each year. Data were available from 1983, but to match crossbill data, the period 1986–2021 was used (n = 36 years). Counts were conducted in autumn, and for pine second year cones (mature cones) were counted. Counts were conducted in two different ways: in interval groups (up to and including 2014) and as exact numbers (from 2011). Thus, there were four years where both methods were used, and in these years counts were strongly correlated (spruce: r = 0.99, n = 111, p < 0.001; pine: r = 0.99, n = 114, p < 0.001). However, interval values were consistently higher than exact counts [mean difference: 7.1 for spruce (SE = 0.25) and 16.7 for pine (SE = 0.69)], and this was partly because the first interval (0–16 cones) was coded as 8 so that even years with no cones had a value of 8. Thus, exact values for 2015–2021 were adjusted by adding the mean difference. This was preferred over subtracting from interval values because this would have produced a few negative values. For remaining years (1986–2014), interval values were used.
For national indices for Sweden, values were averaged across regions. However, spruce data from three regions (northern Dalarne, Malmöhus, Gotland) were excluded because of low number of sampled trees (< 10) in many years. Seven years from the remaining 28 regions were also excluded for the same reason. For pine, two regions (Malmöhus, Blekinge) were excluded together with 24 years from the remaining 29 regions. Thus, analyses were based on 1,001 data points for spruce and 1,020 for pine. Yearly crop size indices for Sweden did not show temporal trends (spruce: r = 0.13, p = 0.46, pine: r = 0.19, p = 0.27).
Spruce crop sizes across Sweden often fluctuated in synchrony, but in some years in particular northern parts of Sweden and the southernmost nemoral areas differed from south-central parts (data not shown). To provide data for a more restricted area where spruce crop size fluctuated relatively synchronously, a subset of the data based on regions in southern Sweden were used (the regions Värmland, Västmanland, Uppland, Södermanland, Närke, Dalsland, Bohuslän, Halland, Västergötland, Östergötland and Småland). Although there were less clear regional patterns in crop size of pine, an index based on the same regions was also used for pine. Yearly crop size indices for southern Sweden did not show temporal trends (spruce: r = 0.21, p = 0.22, pine: r = 0.10, p = 0.57).
Data on seed crop sizes for spruce and pine in Norway were based on the yearly amount of cones collected by Stiftelsen Det norske Skogfrøverk (www.skogfroverket.no). Cones are collected to provide seeds for tree cultivation, and the amount collected depends on cone availability and therefore provides an index of seed crop size. Collection was conducted in autumn and winter, and for pine second year cones (mature cones) were collected. Amount collected was measured in kg, and numbers were based on total amount collected throughout Norway. Data were available from 1961, but to match crossbill data, the period 1986–2021 was used (n = 36 years). Amounts collected of both spruce and pine cones decreased over the study period, although significantly only for pine (spruce: r = -0.23, p = 0.19, pine: r = -0.53, p < 0.001). The decreasing trends are due to both decreasing demand for saplings and more efficient production of saplings (i.e. more saplings produced from the same amount of seeds).
Statistical analyses
Fluctuations in crossbill numbers and seed crop sizes are shown with yearly mean and median values, and range of values. In addition, to compare variability between the two crossbill species and the two tree species, coefficients of variation (CV) were calculated (standard deviation/mean).
Correlation analyses were used to explore the temporal synchrony of crossbill numbers and crop sizes. Analyses included year to account for long-term trends in crossbill numbers and crop sizes, and partial correlation coefficients when controlling for the effect of year are reported. Analyses used log10-transformed numbers (log x + 1) for number of crossbills on migration and crop sizes in Norway.
The main analyses of the influence of crop sizes on crossbill breeding numbers and migration were conducted with generalized linear models (GLM) with Gaussian error distribution. The response variable was breeding numbers or migration, and explanatory variables were spruce crop size and pine crop size. Year was included as a covariate to control for temporal trends in several variables. No interactions were included. Breeding numbers were analysed in relation to crop size in the previous year based on the observation that seed crops measured in autumn are available to crossbills until the next year when seeds are shed in late winter or in spring. Crossbill numbers on migration during autumn were analysed in relation to crop size in the same year. In an additional analysis, crossbill autumn migration was analysed in relation to crop sizes of both spruce and pine in the same year and breeding numbers in the same year, with year as a covariate. Analyses used log10-transformed numbers (log x + 1) for number of crossbills on migration and crop sizes in Norway. Results are presented with parameter estimates based on centered and scaled predictors to allow comparisons of the relative importance of predictors.
Relationships between explanatory variables (spruce and pine seed crop sizes) and response variables (breeding numbers and migration) were illustrated with univariate regression plots. To remove temporal trends in the data, plots were based on detrended data for variables with significant year effects (crossbill citizen data, Swedish monitoring data on Parrot Crossbill, Norwegian seed crop sizes). Detrended data were based on residuals from regressions of the focal variables on year. All statistical analyses were conducted in JMP Pro version 16.
Results
Fluctuations in crossbill numbers and seed crop size
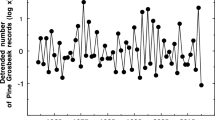
There was substantial fluctuations from year to year for both crossbill numbers and seed crop size (Fig. 1). In general, coefficients of variation were larger for time series of Common Crossbills than for Parrot Crossbills, and for Norway Spruce than for Scots Pine (Online Resource 1, Table S1), although there was no difference for the breeding monitoring data of crossbills in Sweden.



Time series of fluctuations in breeding population size and migration of two crossbill species (Common Crossbill Loxia curvirostra and Parrot Crossbill L. pytyopsittacus), and seed crop size of Norway Spruce (Picea abies) and Scots Pine (Pinus sylvestris) in Norway and Sweden. Year 85 = 1985
Synchrony of crossbill numbers and seed crop size
In general, there were significant correlations both within and between crossbill species (Online Resource 1; Fig. S1–S3, Table S2). Nine out of 16 analyses showed synchrony in within-species population fluctuations of crossbills between Norway and Sweden, or between independent data sets within the same country. All five analyses of between-species population fluctuations of crossbills within the same regions showed that they fluctuated in synchrony. Furthermore, emigration from Sweden was synchronous for the two species.
Similarly, crop size fluctuations within tree species were synchronous between Norway and Sweden (Online Resource 1; Fig. S4, Table S2). However, crop sizes of spruce and pine were only weakly related at national scales, but were synchronous within southern Sweden (Online Resource 1; Fig. S4, Table S2).
Determinants of crossbill breeding population size
Breeding population size of Common Crossbills was largest when there had been a good spruce seed crop in the year before, whereas the size of the pine crop was not positively related to population size (Fig. 2, Table 2). Surprisingly, Parrot Crossbill breeding population size was not related to the size of the pine crop in the previous year, whereas spruce crop size had positive relationships with breeding population size, and significantly so in two out of four analyses (Fig. 3, Table 2).

Breeding population size of Common Crossbill (Loxia curvirostra) in relation to seed crop size of Norway Spruce (Picea abies) and Scots Pine (Pinus sylvestris) in Norway and Sweden. Seed crop size was measured in the previous year, but was assumed to be available to crossbills into the next year. Detrended variables are shown with *. Regression lines and correlation coefficients are reported in each panel irrespective of significance level

Breeding population size of Parrot Crossbill (Loxia pytyopsittacus) in relation to seed crop size of Norway Spruce (Picea abies) and Scots Pine (Pinus sylvestris) in Norway and Sweden. Seed crop size was measured in the previous year, but was assumed to be available to crossbills into the next year. Detrended variables are shown with *. Regression lines and correlation coefficients are reported in each panel irrespective of significance level
Determinants of crossbill migration
Analyses of crossbill migration in relation to crop sizes of spruce and pine showed that numbers were highest when there was a poor spruce seed crop in the same year, whereas numbers were not related to pine crop size (Figs. 4–5, Table 2). Analyses which included breeding population size as an additional explanatory variable did not reveal consistent relationships between migration and population size earlier in the same year (Table 3).

Autumn migration in southern Sweden of Common Crossbill (Loxia curvirostra) in relation to seed crop size in the same year of Norway Spruce (Picea abies) and Scots Pine (Pinus sylvestris) in Norway and Sweden. Detrended variables are shown with *. Regression lines and correlation coefficients are reported in each panel irrespective of significance level

Autumn migration in southern Sweden of Parrot Crossbill (Loxia pytyopsittacus) in relation to seed crop size in the same year of Norway Spruce (Picea abies) and Scots Pine (Pinus sylvestris) in Norway and Sweden. Detrended variables are shown with *. Regression lines and correlation coefficients are reported in each panel irrespective of significance level
Discussion
Identification of crossbills
Field identification of Common and Parrot Crossbill is based mainly on vocalizations and bill morphology. Although both calls and bill sizes differ on average (Cramp and Perrins 1994) this may be difficult to judge by observers with limited experience with the species. Thus, it is important to evaluate whether the data used in analyses could contain biases due to misidentification. The field work of the Swedish Bird Survey and the Norwegian Breeding Bird Survey is conducted by a large number of different persons who may differ in their experience with crossbills. Thus, the Swedish Bird Survey operates with the category 'Unidentified crossbills' which constituted 50% of all crossbills recorded in the breeding surveys whereas Parrot Crossbills were only 9% (see Methods). Thus, even moderate variation in the proportion of unidentified crossbills that were actually Parrot Crossbills could possibly change the breeding index of the rarer Parrot Crossbill substantially, and likely more than unidentified crossbills would change the Common Crossbill index. Whether these unidentified crossbills, had they been identified, would have made the fluctuations of Parrot Crossbills more or less similar to the fluctuations of Common Crossbills is difficult to say.
Similarly, the indices of crossbill numbers based on citizen data likely even contains some proportion of misidentified species records, but it is impossible to know how large this proportion is. On the other hand, migration counts in southern Sweden were likely less affected by identification problems because counts were done by a few persons with many years of extensive experience, although these observers have not been tested for their reliability and consistency in identification. Despite these potential problems (unidentified and misidentified crossbills), there were positive correlations (four out of six were significant) between different and independent Parrot Crossbill breeding population size indices in Norway and Sweden. Theoretically, one could object that these correlations could have been mediated by the fluctuations in Common Crossbill numbers if some Common Crossbills were misidentified and recorded as Parrot Crossbills. However, different Parrot Crossbill indices remained positively correlated (five out of six were significant) even when controlling for Common Crossbill indices (Online Resource 1, Table S3). Thus, the Parrot Crossbill indices used likely reflected true population size. Similarly, a majority of different and independent Common Crossbill indices were positively related, and in combination with a larger annual variation in this species, the data likely reflected true population fluctuations.
Importance of alternative food resources
The results of the present study showed that the two crossbill species were differentially affected by the availability of alternative food sources. In line with a previous suggestion (Alerstam 1982), emigration of Parrot Crossbills occurred synchronously with emigration of Common Crossbills likely because the Parrot Crossbill used spruce seeds extensively as an alternative food source. Thus, both species emigrated when there was a poor spruce seed crop. On the other hand, migration of Common Crossbills was not related to pine crop size, and, surprisingly, neither was the Parrot Crossbill. Similarly, breeding population sizes of both species were positively related to spruce crop size in the previous year (seeds are available into the breeding season of the focal year), whereas not even breeding of the Parrot Crossbill was related to pine crop size. The patterns found for the Common Crossbill match those found in several previous studies (see references in Introduction, including that Reinikainen (1937) also did not find an effect of pine crop on the Common Crossbill), while there have been no previous quantitative studies of the Parrot Crossbill. The patterns found in the present study appeared to be robust because several independent data sets, both for crossbills and for crop sizes, showed similar patterns.
The asymmetric importance of alternative food resources may be explained by the difference in cone toughness between spruce and pine, similar to how cone structure affects foraging efficiency of North American ecotypes of Common Crossbills (Benkman 1987a, 1993, 2003). The Parrot Crossbill with its large bill can open spruce cones relatively easy, whereas Common Crossbills probably have more difficulties opening the tough pine cones. Thus, asymmetry in use of alternative food resources may be related to the costs and benefits of handling different food types. This asymmetry also provides an additional explanation for why Parrot Crossbills are less nomadic than Common Crossbills. Not only do Parrot Crossbills have a less fluctuating staple food resource, but they also apparently use alternative food resources to a larger extent than the Common Crossbill.
Parrot Crossbills and pine and spruce crop sizes
The lack of relationships between Parrot Crossbill numbers and pine crop size is surprising. Perhaps studies with better spatial resoultion would show that breeding Parrot Crossbills concentrate in areas with locally good pine seed crops. Personal observations (SD) from Oslo and Akershus counties in southeastern Norway in 2023 (not included in the present study) indicated that a good pine crop size coincided with an exceptionally large breeding density of Parrot Crossbills. However, partly because the Parrot Crossbill is rarer than the Common Crossbill, and partly because citizen data are generally scarce from the boreal forests in which they breed, local scale analyses are currently not possible due to little data.
However, the pattern that Parrot Crossbills were influenced more by their alternative food source (spruce) than their 'own' food source (pine) challenges the key conifer hypothesis that each crossbill species and ecotype is closely adapted to a single food resource (Benkman 1993, 2003). Actually, crossbill diet selection may sometimes be opportunistic and depend on availability of food sources (Benkman 1987b). For example, the Two-barred Crossbill (Loxia leucoptera) may shift seasonally between two spruce species and Tamarack (Larix laricina) in North America (Benkman 1987b), and irruptive populations in Eurasia may breed based on Norway Spruce (Dale 2021), even though Siberian Larch (Larix sibirica) is the main food in the native range (Cramp and Perrins 1994). Furthermore, Common Crossbill ecotypes in North America do not always stick to their key conifer species (Kelsey 2008; Centanni 2023). This suggests that resource specialization in crossbills is temporally variable, but with highest specialization when alternative resources are scarce.
Spruce crop dynamics
The results of the present study indicated that availability of spruce seeds was important for both species of crossbill. These results were partly based on spruce crop size indices at national levels. However, flowering and seed production of masting trees may be asynchronous at large spatial scales (Koenig and Knops 2000; LaMontagne et al. 2020). In Finland, Gallego Zamorano et al. (2018) found that spruce seed production was strongly correlated over distances of at least 500 km. However, the national indices used here covered distances of up to ca. 2000 km, and within these areas there may be latitudinal trends as well as elevational trends reflecting climatic influences on crop size (Pukkala et al. 2010; Gallego Zamorano et al. 2018). Thus, more local scale analyses might reveal correlations between crossbills and crop sizes more easily. To this end, we also used indices of crop size from southern Sweden, and analyses using these data sets corroborated the main findings.
Large mast years of spruce are irregular and in our data sets occurred at intervals of two or more years (Fig. 1), sometimes up to 6–8 years between really good crop years. Selås (1997) reported that mast years of spruce (bumper crops) occurred at 7–13 year intervals in southern Norway during 1930–1985, whereas Nussbaumer et al. (2016) reported 1–9 year intervals in Norway during 1987–2012. In Finland, large seed crops of spruce occurred at intervals of 2–6 years (Lindén et al. 2011). It is well known that these crop fluctuations have large impacts on population dynamics and movements of Common Crossbills (Reinikainen 1937; Svärdson 1957; Newton 1972, 2006b; Förschler et al. 2006; Marquiss et al. 2012). The present results indicate that also Parrot Crossbills are strongly influenced by spruce seed crop dynamics, and although Parrot Crossbills are less nomadic than Common Crossbills and have smaller population fluctuations, this helps explain irruptive movements of the Parrot Crossbill (e.g. Catley and Hursthouse 1985). Several other studies have also found that migratory behaviour of crossbills may be related to the stability of food resources (Arizaga et al. 2014; Alonso et al. 2017).
Although climate has an indirect influence on crossbills through timing of crop years (Pukkala et al. 2010; Gallego Zamorano et al. 2018), there have been no previous suggestions that weather conditions have a direct influence on breeding or migration (Newton 1972; Cramp and Perrins 1994). On the contrary, crossbills are known for being able to breed in sub-zero temperatures during winter (Haftorn 1971; Cramp and Perrins 1994), and this is likely because access to food (cones in treetops) is not affected by temperature and usually not even by snowfalls. Thus, population dynamics and migration are probably mostly driven by fluctuations in food availability.
Population consequences of differences in resource use
It may appear paradoxical that the Parrot Crossbill is the rarer of the two crossbill species, considering that the results suggest that the Parrot Crossbill is better able to utilize alternative food resources. However, although the Common Crossbill is more abundant on average, it may be completely absent over large areas in years when the spruce has very small seed crops. On the other hand, in mast years we suggest that the Common Crossbill may reach larger abundance because it may be more efficient than the Parrot Crossbill when foraging on spruce seeds. Perhaps even more important, spruce mast years may provide a superabundant food supply for which a numerical response by crossbills is better achieved through immigration (immediate response of the migratory Common Crossbill) than through reproduction (slow response of the more sedentary Parrot Crossbill). Thus, the migratory response of the Common Crossbill to widely fluctuating food supply may give larger average population size than the less nomadic lifestyle of the Parrot Crossbill as a response to a more stable food supply. Thus, the specialization of two closely related species to different main food types have a wide range of consequences, including use of alternative food resources, migration and population size.
Conclusions
Based on the best data available from northern Europe, this study has explored the relative importance of spruce and pine seed crop sizes on breeding and migration of two crossbill species. Likely due to the interspecific differences in bill size and shape, Parrot Crossbill numbers were related more strongly to alternative food resources than Common Crossbill numbers. The patterns found in the present study should be followed up by more detailed and small-scale studies of local populations of crossbills through different phases of crop fluctuations, in particular to gain more knowledge of how pine crop size affects the Parrot Crossbill.
Data availability
Data are available from the corresponding author on request.
References
Alerstam T (1982) Fågelflyttning. Bokförlaget Signum, Lund, Sweden. (In Swedish)
Alonso D, Arizaga J, Meier CM, Liechti F (2017) Light-level geolocators confirm resident status of a Southern European Common Crossbill population. J Ornithol 158:75–81. https://doi.org/10.1007/s10336-016-1388-5
Arizaga J, Alonso D, Hobson KA (2014) Disentangling the origin of crossbills using morphology and isotopic (δ2H) characters. Are southern European crossbills restricted to population-specific key resources? J Ornithol 155:1027–1035. https://doi.org/10.1007/s10336-014-1089-x
Benkman CW (1987a) Crossbill foraging behavior, bill structure, and patterns of food profitability. Wilson Bull 99:351–368
Benkman CW (1987b) Food profitability and the foraging ecology of crossbills. Ecol Monogr 57:251–267. https://doi.org/10.2307/2937083
Benkman CW (1993) Adaptation to single resources and the evolution of crossbill (Loxia) diversity. Ecol Monogr 63:305–325. https://doi.org/10.2307/2937103
Benkman CW (2003) Divergent selection drives the adaptive radiation of crossbills. Evolution 57:1176–1181. https://doi.org/10.1111/j.0014-3820.2003.tb00326.x
Bock CE, Lepthien LW (1976) Synchronous eruptions of boreal seed-eating birds. Am Nat 110:559–571. https://doi.org/10.1086/283091
Catley GP, Hursthouse D (1985) Parrot Crossbills in Britain. Brit Birds 78:482–505
Centanni C (2023) Distribution and ecology of red crossbill call types in western Oregon. Oregon State University, Thesis
Cramp S, Perrins CM (1994) The Birds of the Western Palearctic, vol VIII. Oxford University Press, Oxford, Crows to finches
Dale S (2021) Ecology of a Two-barred Crossbill (Loxia leucoptera) irruption to Norway in 2019–20: altitudinal migration and interspecific habitat differences. Ornis Norv 44:19–30
Dale S (2023) Irruptions of pine grosbeaks pulled by rowanberry peaks in southern areas. J Ornithol 164:353–366. https://doi.org/10.1007/s10336-022-02032-w
Förschler MI, Förschler L, Dorka U (2006) Population fluctuations of siskins Carduelis spinus, common crossbills Loxia curvirostra, and citril finches Carduelis citrinella in relationship to flowering intensity of spruce Picea abies. Ornis Fenn 83:91–96
Fystro I (1962) Skogbrukets frøforsyning. Skogforlaget A/S, Oslo ((In Norwegian))
Gallego Zamorano J, Hokkanen T, Lehikoinen A (2018) Climate-driven synchrony in seed production of masting deciduous and conifer tree species. J Plant Ecol 11:180–188. https://doi.org/10.1093/jpe/rtw117
Green M. Haas F, Lindström Å, Nilsson L (2021) Monitoring population changes of birds in Sweden. Annual report for 2020. Department of Biology, Lund University
Haftorn S (1971) Norges fugler. Universitetsforlaget, Oslo ((In Norwegian))
Kålås JA, Husby M, Vang R (2019) Endringer for Våre Fuglebestander Vår Fuglefauna 42:19–23 ((In Norwegian))
Kelly D (1994) The evolutionary ecology of mast seeding. Trends Ecol Evol 9:465–470. https://doi.org/10.1016/0169-5347(94)90310-7
Kelsey TR (2008) Biogeography, foraging ecology, and population dynamics of red crossbills in North America. PhD thesis, University of California, Davis
Koenig WD, Knops JMH (2000) Patterns of annual seed production by Northern Hemisphere trees: a global perspective. Am Nat 155:59–69. https://doi.org/10.1086/303302
Koenig WD, Knops JMH (2001) Seed-crop size and eruptions of North American boreal seed-eating birds. J Anim Ecol 70:609–620. https://doi.org/10.1046/j.1365-2656.2001.00516.x
Lack D (1954) The Natural Regulation of Animal Numbers. Clarendon Press, Oxford
LaMontagne JM, Pearse IS, Greene DF, Koenig WD (2020) Mast seeding patterns are asynchronous at a continental scale. Nature Plants 6:460–465. https://doi.org/10.1038/s41477-020-0647-x
Lindén A, Lehikoinen A, Hokkanen T, Väisänen RA (2011) Modelling irruptions and population dynamics of the great spotted woodpecker – joint effects of density and cone crops. Oikos 120:1065–1075. https://doi.org/10.1111/j.1600-0706.2010.18970.x
Marquiss M, Newton I, Hobson KA, Kolbeinsson Y (2012) Origins of irruptive migrations by common crossbills Loxia curvirostra into northwestern Europe revealed by stable isotope analysis. Ibis 154:400–409. https://doi.org/10.1111/j.1474-919X.2012.01221.x
Newton I (1972) Finches. Collins, London
Newton I (2006a) Advances in the study of irruptive migration. Ardea 94:433–460
Newton I (2006b) Movement patterns of common crossbills Loxia curvirostra in Europe. Ibis 148:782–788. https://doi.org/10.1111/j.1474-919X.2006.00585.x
Newton I (2008) The Migration Ecology of Birds. Academic Press, London
Newton I (2012) Obligate and facultative migration in birds: ecological aspects. J Orn 153:S171–S180. https://doi.org/10.1007/s10336-011-0765-3
Nussbaumer A, Waldner P, Etzold S, Gessler A, Benham S, Thomsen IM, Jørgensen BB, Timmermann V, Verstraeten A, Sioen G, Rautio P, Ukonmaanaho L, Skudnik M, Aputhin V, Braun S, Wauer A (2016) Patterns of mast fruiting of common beech, sessile and common oak, Norway spruce and Scots pine in Central and Northern Europe. Forest Ecol Manag 363:237–251. https://doi.org/10.1016/j.foreco.2015.12.033
Ottosson U, Ottvall R, Elmberg J, Green M, Gustafsson R, Haas F, Holmqvist N, Lindström Å, Nilsson L, Svensson M, Svensson S, Tjernberg M (2012) Fåglarna i Sverige – antal och förekomst. Swedish Ornithological Society, Halmstad, Sweden ((In Swedish))
Owens JN, Blake MD (1985) Forest tree seed production. A review of the literature and recommendations for future research. Information Report PI-X-53, Petawawa National Forestry Institute, Canadian Forestry Service, Agriculture Canada
Pukkala T, Hokkanen T, Nikkanen T (2010) Prediction models for the annual seed crop of Norway Spruce and Scots Pine in Finland. Silva Fenn 44:629–642
Reinikainen A (1937) The irregular migrations of the crossbill, Loxia c. curvirostra, and their relation to the cone-crop of the conifers. Ornis Fenn 14:55–64
Selås V (1997) Cyclic population fluctuations of herbivores as an effect of cyclic seed cropping of plants: the mast depression hypothesis. Oikos 80:257–268
Shimmings P, Øien IJ (2015) Bestandsestimater for norske hekkefugler. Norsk Ornitologisk Forening, Norway ((In Norwegian))
Silvertown JW (1980) The evolutionary ecology of mast seeding in trees. Biol J Linn Soc 14:235–250. https://doi.org/10.1111/j.1095-8312.1980.tb00107.x
Strong C, Zuckerberg B, Betancourt JL, Koenig WD (2015) Climatic dipoles drive two principal modes of North American boreal bird irruption. Proc Natl Acad Sci USA 112:E2795–E2802. https://doi.org/10.1073/pnas.1418414112
Svärdson G (1957) The “invasion” type of bird migration. Brit Birds 50:314–343
Acknowledgements
We thank Ulfstand Wennström (Skogforsk, Forestry Research Institute of Sweden) for access to Swedish cone estimates, and two anonymous reviewers for comments on the manuscript.
Funding
Open access funding provided by Norwegian University of Life Sciences.
Author information
Authors and Affiliations
Contributions
SD conceived the study, SD and ØME acquired data, SD analysed data and drafted the manuscript, SD and ØME revised the manuscript and approved the final manuscript.
Corresponding author
Ethics declarations
Conflicts of interests:
The authors have no conflicts of interest to declare.
Ethical approval
Not applicable.
Additional information
Communicated by N. Chernetsov.
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Dale, S., Edvardsen, Ø.M. Fluctuations in population size and migration of two species of crossbills in relation to seed crop size of spruce and pine: asymmetric importance of alternative food resources. J Ornithol (2024). https://doi.org/10.1007/s10336-024-02173-0
Received:
Revised:
Accepted:
Published:
DOI: https://doi.org/10.1007/s10336-024-02173-0