
Abstract
During polar nights of 2012 and 2017, bioluminescence (BL) potential surveys demonstrated high emissions at depths around and below 100 m at offshore stations to the north of a Svalbard fjord (Rijpfjorden). We demonstrated that the highest bioluminescent emissions for offshore stations are located at depths below depths of modelled/ambient light intensities corresponding to the reported irradiance thresholds for the behavioral light sensitivity of krill and copepods, and suggest that behavioral light sensitivity is one of the reasons for high values of BL potential observed below 100 m at offshore stations. In order to understand sources of bioluminescent taxa responsible for the observed high values of BL potential in offshore waters, we have investigated the origin and pathways of water masses circulating to the north, offshore of the fjord Rijpfjorden by using a hydrodynamic model. For both 2012 and 2017, the model water masses mostly originate from the west, where the Atlantic water is flowing northward, then along the shelf and shelf slope of northern Svalbard, and to the offshore of the fjord. This indicates that the advection of zooplankton by North Atlantic Water is one possible source of bioluminescent organisms offshore of northern Svalbard. In 2012, water masses also originated from the inflow through Hinlopen trench and strait, while, in 2017, the offshore water was advected and upwelled into the fjord on time scales less than 10 days, and after that there was a recirculation back from the fjord to offshore on time scales larger than 10 days. This recirculation from the fjord might be another source of bioluminescent organisms in the offshore waters.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
1 Introduction
The polar night period presents challenging optical conditions for Arctic pelagic organisms. With the sun remaining below the horizon from one day at the Arctic Circle to 6 months at the North Pole, prolonged darkness limits light-mediated predator prey interactions in the plankton. For macrozooplankton and micronekton, the twilight optical conditions present during polar night, while dim, do appear sufficient to drive physiological processes such as entrainment of biological rhythms (Cohen et al. 2021) and behavioral processes such as vertical migrations on diel (Ludvigsen et al. 2018) and lunar (Last et al. 2016) timescales. Along with diffuse atmospheric light from the sun and moon, bioluminescence represents an in-water light source that can facilitate ecological interactions in an otherwise dim-light environment. Indeed, bioluminescence is a common feature of the pelagic during polar night (Berge et al. 2012; Johnsen et al. 2014; Cronin et al. 2016; Cohen et al. 2020). For bioluminescent flashes to be effective in predator–prey interactions, they must be observable over background light (Nilsson et al. 2014). Yet, we know little about the distribution of bioluminescence as related to environmental light during Arctic Polar night. Because most bioluminescent organisms in the marine environment generate light in response to mechanical stimulation, bioluminescence is commonly measured as bathyphotometer bioluminescence potential (BL potential), defined as mechanically stimulated light measured inside of a chambered pump-through bathyphotometer (Herren et al. 2005). The bathyphotometer pumps water into its detection chamber and mechanically stimulates marine organisms to produce light inside the detection chamber.
As demonstrated in Shulman et al. (2020, 2022), two surveys during polar nights of 2012 and 2017 (12–16 January 2012 and 10–14 January 2017) recorded higher bioluminescence (BL) potential emissions in the top 50 m in the northern Svalbard fjord (Rijpfjorden) in comparison to that measured at offshore stations. Around 56% of BL potential non-zero emissions were larger or equal to 108 photon/s in the fjord area in 2012 and 39% in 2017, while no more than 22% in 2012 and 20% in 2017 were observed at offshore stations located on the shelf-break, shelf-slope areas and in the Arctic basin (Shulman et al. 2020, 2022). The value of 108 photons/s is about 100 times larger than background BL potential from filtered seawater in the Arctic (Johnsen et al. 2014; Cronin et al. 2016). At the same time, observations of BL potential also indicated high emissions at depths below 100 m at offshore stations for both years. Our previous studies demonstrated that these high offshore values of BL potential below 100 m are upwelled and advected to the top 50 m of the fjord (Shulman et al. 2020, 2022). We demonstrated that upwelling and advection of these deep high BL potential values (and therefore, upwelling and advection of corresponding bioluminescent taxa) from offshore areas are dominant factors in the observed BL potential dynamics in the top 50 m of the fjord (Shulman et al. 2020, 2022). In the present paper, we address the following questions expanding upon our previous studies: (1) why were bioluminescent taxa observed predominantly below 100 m in offshore waters of the fjord? and (2) what are origins and pathways of water masses circulating to the offshore? This will help to understand the origin and sources of the observed bioluminescent taxa in offshore waters of the fjord.
Full moon or near full moon conditions were present during both BL potential sampling periods under consideration. Marine organisms are sensitive to light, and numerous recent studies have demonstrated that the position of zooplankton is controlled by light, including moonlight (Cohen et al. 2020, 2021; Webster et al. 2015; Berge et al. 2012, 2020; Hobbs et al. 2021; Last et al. 2016; Batnes et al. 2015). Therefore, the vertical migration of the bioluminescent organisms (due to behavioral sensitivity to the light conditions) might be one of the reasons why high values of BL potential were observed below 100 m during both polar nights under consideration. In the present paper, we address question (1) above by relating bioluminescent taxa behavioral light sensitivity to BL potential distributions with depth by using observed BL potential during both polar nights’ surveys.
There is increasing evidence showing the influence of Atlantic water (Atlantification) in the Arctic generally, and in Svalbard fjords in particular (Polyakov et al. 2020; Csapo et al. 2021; Lind et al. 2018). Indeed, Falk-Petersen et al. (2015) suggested extensive upwelling of Atlantic Water north of Svalbard in January 2012. Based on modeling results of Lind and Ingvaldsen (2012), Falk-Petersen et al. (2015) suggest that an Ekman transport toward the north along the northern coast of Svalbard increases the cross-shelf exchange and the penetration of Atlantic Water onto the shelf. Basedow et al. (2018) observed a strong influx of zooplankton with North Atlantic Water during January of 2014. This is also supported by recent studies of ecosystem dynamics around the Svalbard area (Wassmann et al. 2019; Vernet et al. 2019). Advection of zooplankton by the North Atlantic Water is one possible source of bioluminescent organisms offshore of northern Svalbard during the polar night. In the present paper, we address question (2) above by testing this using a hydrodynamic model to investigate the origin and pathways (hydrographic history) of water masses circulating to the north of the Svalbard fjord Rijpfjorden.
2 Methods
2.1 Bioluminescence potential observations during polar nights of 2012 and 2017
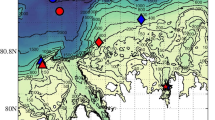
The area of study and locations of observational stations sampled during the polar nights of January 10–14, 2012, and January 10–14, 2017, are shown in Fig. 1. Stations A2012 and A2017 are located at the mouth of the fjord Rijpfjorden; stations B2012 and B2017 are to the north of the fjord and in the shelf-break, shelf-slope area; station C2012 is also located in the shelf-break, shelf-slope area, but is to the west-northwest of the fjord, and finally station D2017 is the further-most offshore and in the Arctic basin.

Study area around Svalbard, NO, and locations of observational stations. Stations sampled in 2012 are as follows: A2012, blue star; B2012, blue diamond; C2012, blue triangle. Stations sampled in 2017 are as follows: A2017, red star; B2017, red diamond; D2017, red circle
Surveys of BL potential were conducted using a bathyphotometer called the Underwater Bioluminescence Assessment Tool (UBAT). For both years, the UBAT was mounted on a profiling cage. In 2012, downcasts surveys down to 500 m depth were used in the present study. In 2017, each profile consisted of 4 min stops at 20-m depth intervals down to 120-m depth. The UBAT outputs light data at 60 Hz (60 outputs per second), as well as averaged data over 1 s. For more information about the BL observations during polar nights of 2012 and 2017, see Shulman et al. (2020) and (2022).
2.2 Ranges of behavioral light sensitivity for copepods and krill, and surface light data during polar night
Batnes et al. (2015) reported ranges of the behavioral light sensitivity for copepods (Calanus spp.) between 5 × 10−8 and 1.7 × 10−7 μmol photons m−2 s−1, while Myslinski et al. (2005) reported ranges of the behavioral light sensitivity for krill (Meganyctiphanes norvegica) between 108 and 107 photons cm−2 s−1 (which is between 1.66 × 10−7 and 1.66 × 10−6 μmol photons m−2 s−1). Note, that the value of 10−6 μmol photons m−2 s−1 is reported as light sensitivity for another bioluminescent krill Thysanoessa inermis in Cohen et al. (2021). As we stated in the introduction, full moon or near full moon conditions occurred during both sampling periods under consideration. Surface light data from the ArcLight observatory in Ny-Ålesund, Svalbard (79° N) provides a multi-year record of light throughout the annual cycle (Johnsen et al. 2021). Cohen et al. (2020) analyzed hourly ArcLight data for the Polar night period, separating measurements throughout the 24-h day when the lunar disk was > 50% full from measurements when it was < 50% full. This analysis suggested that the brightest light conditions, occurring when the lunar disk was > 50% full (including the full moons), reached intensities between 10−4 and 10−3 μmol photons m−2 s−1 when midday solar elevation was approximately 10° below the horizon (comparable to Rijpfjorden at 81°N during our bioluminescence sampling periods). We therefore used these values as incident downwelling light at the sea surface for models of underwater light.
2.3 Estimation of origin and pathways (hydrographic history) of water masses circulating to the offshore of the Svalbard fjord (Rijpfjorden)
In order to investigate the origin and pathways (hydrographic history) of the modeled water masses circulating to the offshore of the Svalbard fjord (Rijpfjorden), we conducted numerical experiments with the adjoint to the tracer model (see Appendix). The adjoint tracer distribution provides information on the model tracer history and identifies the origin and pathways of the model tracer-tagged water masses in the past, which circulated into the area of interest (Appendix, Shulman et al. 2010, 2011). In other words, the adjoint quantifies where water masses were, for example, three days before circulating into a specific geographical area of interest. Therefore, adjoint distributions show the hydrographic history of water masses advected and mixed into the area of interest at evaluation time T.
3 Results
3.1 Impact of behavioral light sensitivity on BL potential distributions with depth
Figure 2 shows the observed vertical profiles of BL potential at stations B2012 and C2012 located offshore in the shelf-break, shelf-slope areas (Fig. 1). Ranges of irradiance at surface during the polar night from moonlight (between 10−4 and 10−3 μmol photons m−2 s−1) and ranges of the behavioral light sensitivity for krill (Myslinski et al., 2005) and copepods (Batnes et al., 2015) are also shown in Fig. 2 (note that minimal range of the behavioral light sensitivity for krill (1.66 10−7 μmol photons m−2 s−1) is almost equal to the value of the maximal range of the behavioral light sensitivity for copepods (1.7 10−7 μmol photons m−2 s−1). The ranges of the irradiance from the moon at depths are shown for two values of the attenuation coefficient \({k}_{PAR}(z)\) (for the attenuation of scalar irradiance in the visible spectral range): \({k}_{PAR}(z)\)= 0.0708 (Fig. 2a and b) and \({k}_{PAR}(z)\)=0.18 (Fig. 2c and d). The attenuation coefficient \({k}_{PAR}(z)\)= 0.0708 was reported in Batnes et al. (2015), and the value is close to the attenuation coefficient for clear water (Jerlov, 1976). The second attenuation coefficient \({k}_{PAR}(z)\) = 0.18 was reported in Cronin et al. (2016) as the lowest value of the attenuation coefficient in the Svalbard Kongsfjorden fjord located on the western site of the Svalbard at around 79 N. In accord with Fig. 2a and b, the ranges of the behavioral light sensitivity for copepods are crossing the ranges of the irradiance from the moon at depths around 100–140 m. The ranges of behavioral light sensitivity for krill are crossing the ranges of the attenuated atmospheric light from the moon at depths around 90–120 m. There are observed maxima values of BL potential near 120 m at station B2012 below depths of modelled/ambient light intensities corresponding to the reported thresholds for the behavioral light sensitivity of krill and copepods. Similar relations found for station C2012 in Fig. 2a and b, where high values of BL potential are also observed below 100 m (around 110 m). Using \({k}_{PAR}(z)\) = 0.18 for the moonlight attenuation, Fig. 2c and d and Fig. 3a and b demonstrates clearly that the highest bioluminescent emissions for all four offshore stations are located below depths of modelled/ambient light intensities corresponding to the reported thresholds for the behavioral light sensitivity of krill and copepods. In accord with Fig. 2 and Fig. 3a and b, the depths of the observed BL potential maxima, at offshore stations, are depths to which copepods and krill will migrate under moonlight conditions during polar night. Therefore, our results demonstrate that the observed presence of bioluminescent taxa below 100 m offshore is consistent with the behavioral light sensitivity of bioluminescent taxa (in the case of krill and copepods), and these organisms positioning themselves in relative darkness may be one of the reasons for high values of BL potential observed below 100 m at offshore stations.

Observed vertical profiles of BL potential (1s averaged) at stations B2012 and C2012 with overlaid ranges of irradiance in the polar night from the moon (dotted lines, \({k}_{PAR}(z)\)=0.0708 for a and b; \({k}_{PAR}(z)\)=0.18 for c and d), and ranges of the behavioral light sensitivity for krill (solid vertical) and copepods (dashed vertical) lines

Observed vertical profiles of BL potential (1s averaged) at stations B2017, D2017, A2012, and A2017 with overlaid ranges of irradiance in the polar night from the moon (dotted lines, \({k}_{PAR}(z)\)=0.18), and ranges of the behavioral light sensitivity for krill (solid vertical) and copepods (dashed vertical) lines
For stations A2012 and A2017 in the fjord (Fig. 3c and d), there is no clear relation between the depths of observed high BL potential values and behavioral light sensitivity as at offshore stations for both years. At station A2012, high values of BL potential are at depths shallower than depths of crossings of moonlight attenuation with the thresholds of behavioral light sensitivities. Therefore, other processes dominate the BL potential dynamics in the fjord during polar night of 2012, one of which is advection and upwelling of the deep offshore bioluminescent taxa as demonstrated in Shulman et al. (2020). At station A2017, there is a strong maximum of BL potential located below the crossings of moonlight attenuation with the thresholds of behavioral light sensitivities. However, high values of BL potential are also present in shallow waters in the fjord during polar night of 2017.
3.2 Origin and pathways (hydrographic history) of water masses circulating to the offshore of the Svalbard fjord (Rijpfjorden)
In the present study, we are interested in the origin and pathways of water masses circulating into a subsurface around 100 m, in the area of the offshore station B2012, located to the north of the fjord. For this reason, our area of interest is the box of 10 × 10 grid cells of the circulation model (described in Shulman et al. 2020, 2022) around offshore station B2012 (Fig. 4a, red box) with depth between 70 and 150 m. For the polar nights of 2012 and 2017, we selected the evaluation time as January 14th 12Z (= 12:00 UTC) for both years.

Adjoint distribution maps for the polar night of 2012. a Initial distribution of the adjoint. b Distribution of the adjoint after 4 days of integration of the adjoint equation (vertically integrated adjoint for each model grid normalized by the volume of the area of interest). c As in b but after 10 days of integration of the adjoint equation. d As in b but after 20 days. e As in b but after 30 days of integration of the adjoint equation. Those adjoint maps show areas where the model tracer-tagged water masses were 30, 20, 10 and 4 days before circulating into the area of interest (red box around station B2012)
Figure 4a shows the distribution of the adjoint at evaluation time January 14th 12Z of 2012, when the adjoint is set to 1 in the area of interest (the red box around station B2012), and equals 0 everywhere outside of the area of interest. From these initial values at t = T, the adjoint equation to the tracer model is integrated backward in time (Appendix, Eq. (4), see also Shulman et al., 2010, 2011). Vertically integrated adjoint distributions (normalized by the volume of the area of interest) after 4, 10, 20, and 30 days of the backward adjoint integration in time are shown in Fig. 4b–e. Similarly, Fig. 5 shows the distribution of the adjoint at the time evaluation January 14th 12Z of 2017 for the 2017, and after 4, 10, 20, and 30 days of the backward adjoint integration in time. As stated previously, those adjoint maps show areas where the model tracer-tagged water masses were 30, 20, 10, and 4 days before circulating into the area of interest (red box around station B2012) for 2012 (Fig. 4) and 2017 (Fig. 5). For both years, those model water masses mostly originated from the west of station B2012, where the Atlantic water is flowing northward, then along the shelf and shelf slope of the northern Svalbard, and to the area around station B2012. This is in accord with numerous observational and modeling studies conducted previously in the area (Beszczynska-Moller et al. 2012; Falk-Petersen et al. 2015; Basedow et al. 2018; Wassmann et al. 2019; Vernet et al. 2019). At the same time, the adjoint maps show striking differences between the two years. In accord with adjoint maps of 30 days (Fig. 4e) and 20 days (Fig. 4d) for 2012, water masses also originated from the inflow through Hinlopen trench and strait before circulating to the area around station B2012. This is in agreement with Menze et al. (2019), where it is demonstrated that the Atlantic water also enters the shelf of the northern Svalbard through the Hinlopen trench on the western side, transporting heat and nutrients that could boost local productivity. Therefore, in 2012, the model shows that water masses originated from the west all the way to the Fram strait and also through the Hinlopen trench and strait inflow. In 2017, there is no such pronounced inflow through the Hinlopen strait; however, the adjoint maps for 20 days (Fig. 5d) and 30 days (Fig. 5e) show circulation of water masses from the east of the B2012 station and from the fjord Rijpfjorden (location stations A2012 and A2017, Fig. 1). Therefore, in accord with the adjoint maps of Fig. 5, the fjord water masses are reaching offshore station B2012 on time scales around 20 days. This is supported by cross-section plots (Fig. 6) of those adjoint maps: the adjoint values are plotted along a section crossing the mouth of the fjord (see Fig. 5) at longitude 22.08° E. Figure 6 shows where water masses were along latitude 22.08E before they were circulating to the offshore station B2012. It shows that water masses from the fjord (the A2017 station latitude is marked by the red star in Fig. 6) begin to reach the area around offshore station B2012 after 10 days (starting with deeper water masses) and, mostly, the water masses from the fjord reach the offshore after 20 days. At the same time, Shulman et al. (2020) and (2022) demonstrated that water masses from offshore below 100 m (from the area around station B2012) are advected and upwelled into the top 50 m of the fjord (area around stations A2012 and A2017) on time scales less than 10 days for both years. Therefore, for 2017, the offshore water masses are advected and upwelled into the top 50 m of the fjord on time scales less than 10 days, and after that, there is a recirculation (starting with the deeper water) back from the fjord to the offshore on time scales larger than 10 days. Because of high BL potential emissions observed in the fjord (Fig. 3), this recirculation from the fjord might be another (along with Atlantic water inflow) source of bioluminescent organisms (and therefore, the observed high BL potential) in the offshore waters below 100 m in 2017.

Adjoint distribution maps for the Polar night of 2017. a Initial distribution of the adjoint. b Distribution of the adjoint after 4 days of integration of the adjoint equation (vertically integrated adjoint for each model grid normalized by the volume of the area of interest). c As in b but after 10 days of integration of the adjoint equation. d As in b but after 20 days. e As in b but after 30 days of integration of the adjoint equation. Those adjoint maps show areas where the model tracer-tagged water masses were 30, 20, 10, and 4 days before circulating into the area of interest (red box around station B2012). The location of section plotted in Fig. 6 is shown with maroon color

Adjoint maps plotted in Fig. 5 along the section crossing the mouth of the fjord at 22.08E of longitude (shown in maroon color in Fig. 5). Latitude of station A2017 is shown with the red star, while latitude of station B2012 with the blue rhomb. a Distribution of the adjoint after 4 days. b As in a but after 10 days. c As in a but after 20 days. d As in a but after 30 days of the integration of the adjoint equation
4 Conclusions and discussions
We have related the behavioral light sensitivity of copepods and krill to bioluminescence potential distributions with depth offshore of the northern Svalbard fjord Rijpfjorden during 2012 and 2017 polar nights. For attenuation of moonlight with the depth we used two values: 0.0708 (Batnes et al. 2015) and 0.18 (Cohen et al. 2020). The first value is close to the attenuation coefficient for the clear water type (Jerlov, 1976), while the second value of the attenuation coefficient was reported as the lowest value of \({k}_{PAR}(z)\) in the Kongsfjorden fjord during polar night. With \({k}_{PAR}(z)\) = 0.0708, the ranges of the behavioral light sensitivity for copepods are crossing the ranges of the irradiance from the moon at depths around 100–140 m. The ranges of behavioral light sensitivity for krill are crossing the ranges of the attenuated atmospheric light from the moon at depths around 90–120 m. This coincides with observed maxima values of BL potential at offshore stations which are also near 120 m and 110 m.
With \({k}_{PAR}(z)\)=0.18, our results clearly demonstrate that highest bioluminescent emissions for all offshore stations during 2 years are located below depths of modelled/ambient light intensities corresponding to the reported thresholds for the behavioral light sensitivity of krill and copepods. Therefore, our results demonstrate that the observed presence of bioluminescent taxa below 100 m offshore relates to behavioral light sensitivity of krill and copepods, and the behavioral light sensitivity is one of the reasons why high values of BL potential are observed below 100 m at offshore stations. The depths of the observed deep high values of BL potential are below the depths to which copepods and krill migrate deep due to their respective behavioral light sensitivities under moonlight conditions during polar night.
We have investigated the origin and pathways of the model water masses circulating to the offshore of the Svalbard fjord (Rijpfjorden). For both polar nights, those model water masses mostly originated from the west, where Atlantic water is flowing northward, then along the shelf and shelf slope of the northern Svalbard, and to the offshore of the fjord. Basedow et al. (2018) found that the transport of North Atlantic Water into the Arctic is larger during the polar night (January) than in May and August, and that there is a strong influx of zooplankton with North Atlantic Water during the polar night. Therefore, the inflow of zooplankton with the Atlantic water might be one of the sources of high BL potential offshore. In accord with our results for 2012, water masses also originated from the inflow through Hinlopen trench and strait before circulating to the offshore area of the fjord. In 2017, there is no such pronounced inflow through the Hinlopen strait; however, modeling results show circulation of water masses from the fjord into the offshore area. Therefore, in 2017, the offshore water is advected and upwelled into the top 50 m of the fjord on time scales less than 10 days (in accord with Shulman et al. (2020) and (2022)), and after that, there is a recirculation back (starting with the deeper water) from the fjord to offshore on time scales larger than 10 days. Because of high BL potential emissions observed in the fjord, this recirculation from the fjord might be another (along with Atlantic water inflow) source of bioluminescent organisms (and therefore, the observed high BL potential) in the offshore waters below 100 m. Our search of existing literature did not produce any observational or modeling studies related to the exchange between the fjord Rijpfjorden and offshore; therefore, the above conclusions about the fjord and offshore exchange based on model predictions for only one polar night of 2017 should be taken with caution and need further verification.
Data availability
The datasets generated during the current study are available from the corresponding author on reasonable request.
References
Basedow SL, Sundfjord A, von Appen W-J, Halvorsen E, Kwasniewski S, Reigstad M (2018) Seasonal variation in transport of zooplankton into the arctic basin through the atlantic gateway, fram strait. Front Mar Sci 5:194. https://doi.org/10.3389/fmars.2018.00194
Batnes AS, Miljeteig C, Berge J, Greenacre M, Johnsen G (2015) Quantifying the light sensitivity of Calanus spp. during the polar night: potential for orchestrated migrations conducted by ambient light from the sun, moon, or aurora borealis? Polar Biol 38:51–65. https://doi.org/10.1007/s00300-013-1415-4
Berge J, Batnes AS, Johnsen G, Blackwell SM, Moline MA (2012) Bioluminescence in the high Arctic during the polar night. Mar Biol 159:231–237. https://doi.org/10.1007/s00227-011-1798-0
Berge J, Daase M, Hobbs L, Falk-Petersen S, Darnis G, Soreide J (2020) Zooplankton in the Polar night. In: Berge J, Johnsen G, Cohen JH (eds) Polar night marine ecology: life and light in the dead of night. Springer Nature, Switzerland, pp 116–130
Beszczynska-Moller A, Fahrbach E, Schauer U, Hansen E (2012) Variability in Atlantic water temperature and transport at the entrance to the Arctic Ocean, 1997–2010. ICES J Mar Sci 69(5):852–863. https://doi.org/10.1093/icesjms/fss056
Cohen JH, Berge J, Moline MA, Johnsen G, Zolich AP (2020) Light in the polar night. In: Berge J, Johnsen G, Cohen JH (eds) Polar night marine ecology: life and light in the dead of night. Springer Nature, Switzerland, pp 37–66
Cohen JH, Last KS, Charpentier CL, Cottier F, Daase M, Hobbs L et al (2021) Photophysiological cycles in Arctic krill are entrained by weak midday twilight during the polar night. PLoS Biol 19(10):e3001413. https://doi.org/10.1371/journal.pbio.3001413
Cronin HA, Cohen JH, Berge J, Johnsen G, Moline MA (2016) Bioluminescence as an ecological factor during high Arctic polar night. Sci Rep 6:36374. https://doi.org/10.1038/srep36374
Csapo HK, Grabowski M, Węsławski JM (2021) Coming home – boreal ecosystem claims Atlantic sector of the Arctic. Sci Total Environ 771:144817. https://doi.org/10.1016/j.scitotenv.2020.144817
Falk-Petersen S, Pavlov V, Berge J, Cottier F, Kovacs KM, Lydersen C (2015) At the rainbow’s end: high productivity fueled by winter upwelling along an Arctic shelf. Polar Biol 38:5–11. https://doi.org/10.1007/s00300-014-1482-1
Herren CM, Haddock SHD, Johnson C, Orrico CM, Moline MA, Case JF (2005) A multi-platform bathyphotometer for fine-scale, coastal bioluminescence research, Limnol Oceanogr. Methods 3:247–262. https://doi.org/10.4319/lom.2005.3.247
Hobbs L, Banas NS, Cohen JH, Cottier FR, Berge J, Varpe O (2021) A marine zooplankton community vertically structured by light across diel to interannual timescales. Biol Let 20200810. https://doi.org/10.1098/rsbl.2020.0810
Jerlov NG (1976) Marine optics. Elsevier Scientific Publishing Company, Amsterdam
Johnsen G, Candeloro M, Berge J, Moline M (2014) Glowing in the dark: discriminating patterns of bioluminescence from different taxa during the Arctic polar night. Polar Biol 37(5):707–713. https://doi.org/10.1007/s00300-014-1471-4I
Johnsen G, Zolich A, Grant S, Bjørgum R, Cohen JH, McKee D, Kopec TP, Vogedes D, Berge J (2021) All-sky camera system providing high temporal resolution annual time series of irradiance in the Arctic. Appl Opt 60:6456–6468. https://doi.org/10.1364/AO.424871
Last KS, Hobbs L, Berge J, Brierley AS, Cottier F (2016) Moonlight drives ocean-scale mass vertical migration of zooplankton during the Arctic winter. Curr Biol 26:244–251. https://doi.org/10.1016/j.cub.2015.11.038
Lind S, Ingvaldsen RB (2012) Variability and impacts of Atlantic Water entering the Barents Sea from the north. Deep Sea Res Part I Oceanogr Res Pap 62:70–88. https://doi.org/10.1016/j.dsr.2011.12.007
Lind S, Ingvaldsen RB, Furevik T (2018) Arctic warming hotspot in the northern Barents Sea linked to declining sea-ice import. Nat Clim Chang 8:634–639. https://doi.org/10.1038/s41558-018-0205-y
Ludvigsen M, Berge J, Geoffroy M, Cohen JH, De La Torre PR, Nornes SM, Singh H, Sørensen AJ, Daase M, Johnsen G (2018) Use of an autonomous surface vehicle reveals small-scale diel vertical migrations of zooplankton and susceptibility to light pollution under low solar irradiance. Sci Adv 4:eaap9887
Menze S, Ingvaldsen RB, Haugan P, Beszczynska-Moeller A, Fer I, Sundfjord A, Falk-Petersen S (2019) Atlantic water pathways along the north-western Svalbard shelf mapped using vessel-mounted current profilers. J Geophys Res Oceans 124. https://doi.org/10.1029/2018JC014299
Myslinski TJ, Frank TM, Widder EA (2005) Correlation between photosensitivity and downwelling irradiance in mesopelagic crustaceans. Mar Biol 147:619–629
Nilsson D-E, Warrant E, Johnsen S (2014) Computational visual ecology in the pelagic realm. Philos Trans R Soc B 369:20130038. https://doi.org/10.1098/rstb.2013.0038
Polyakov IV, Alkire MB, Bluhm BA, Brown KA, Carmack EC, Chierici M, Danielson SL, Ellingsen I, Ershova EA, Gårdfeldt K, Ingvaldsen RB, Pnyushkov AV, Slagstad D, Wassmann P (2020) Borealization of the Arctic Ocean in response to anomalous advection from sub-Arctic seas. Front Mar Sci 7:491. https://doi.org/10.3389/fmars.2020.00491
Shulman I, Anderson S, Rowley C, deRada S, Doyle J, Ramp S (2010) Comparisons of upwelling and relaxation events in the Monterey Bay area. J Geophys Res 115:C06016. https://doi.org/10.1029/2009JC005483
Shulman I, Moline MA, Penta B, Anderson S, Oliver M, Haddock SHD (2011) Observed and modeled bio-optical, bioluminescent, and physical properties during a coastal upwelling event in Monterey Bay, California. J Geophys Res 116:C01018. https://doi.org/10.1029/2010JC006525
Shulman I, Moline MA, Cohen JH, Anderson S, Metzger EJ, Rowley C (2020) Bioluminescence potential modeling during polar night in the Arctic: impact of advection versus local sources. Ocean Dyn 70(9):1211–1223. https://doi.org/10.1007/s10236-020-01392-2
Shulman I, Cohen JH, Moline MA et al (2022) Modeling studies of the bioluminescence potential dynamics in a high Arctic fjord during polar night. Ocean Dyn 72:37–48. https://doi.org/10.1007/s10236-021-01491-8
Vernet M, Ellingsen IH, Seuthe L, Slagstad D, Cape MR, Matrai PA (2019) Influence of phytoplankton advection on the productivity along the Atlantic water inflow to the Arctic Ocean. Front Mar Sci 6:583. https://doi.org/10.3389/fmars.2019.00583
Wassmann P, Slagstad D, Ellingsen I (2019) Advection of mesozooplankton into the northern Svalbard shelf region. Front Mar Sci 6:458. https://doi.org/10.3389/fmars.2019.00458
Webster CN, Varpe O, Falk-Petersen S et al (2015) Moonlit swimming: vertical distributions of macrozooplankton and nekton during the polar night. Polar Biol 38:75–85. https://doi.org/10.1007/s00300-013-1422-5
Acknowledgements
We thank Dr. Batnes of NTNU for providing data used in Batnes et al. (2015) and for helpful discussions. We thank anonymous reviewers for provided comments and recommendations.
Funding
This research was funded through the US Naval Research Laboratory under program element 61153 N. Support for JHC and MAM for observational work came from the Norwegian Research Council (NFR) projects ArcticABC and DeepImpact (NFR grants 244319 and 300333). Computer time for the numerical simulations was provided through a grant from the Department of Defense High Performance Computing Initiative. This manuscript is US NRL contribution NRL/7330/JA—2022/5.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare no competing interests.
Additional information
Responsible Editor: Alejandro Orfila
Appendix. The tracer model and its adjoint
Appendix. The tracer model and its adjoint
Consider a passive tracer model for concentration C:
where U is thethree-dimensional (x,y,z) velocity, k represents the horizontal and vertical diffusivities (velocity and diffusivities are taken from the physical circulation model, described in Shulman et al. (2020, 2022)). Initial condition for (1) at \(t={t}_{0}\) has the form:
where D is the modeling domain, and concentration \({C}_{0}(x,y,z)\) is the initial conditions of the tracer in the modeling domain D.
Let us consider the following objective function J at time \(T>{t}_{0}\):
where V is an area of interest (particular sub-domain of the modeling domain), τ is the location in the model domain with coordinates (x,y,z), and dτ is a volume element. Therefore, function J is the normalized content of tracer C in the domain V at time T.
Consider the following adjoint variable λ satisfying the adjoint to (1) equation:
with initial conditions at t = T:
and boundary conditions:
Boundary Sd includes all boundaries of D: lateral boundaries, surface and bottom.
The gradient of the function J (3) at time T with respect to the initial concentration C0 at time t0, can be estimated:
where \(\lambda (\tau ,{t}_{0})\) is result of integration adjoint models (4)–(6) backward in time, from initial conditions (5) at \(t=T\) to time t = t0. Let us introduce some finite perturbation \(\Delta {C}_{0}\), to the initial concentration C0 at time t0, according to (7) we would have:
According to (8), the adjoint tracer distribution \(\lambda (\tau ,{t}_{0})\) represents a fraction of tracer \(\Delta {C}_{0}\), which makes its way to the volume V from time t0 to time T. Due to the linearity of the passive tracer and its adjoint problems, the adjoint tracer distribution \(\lambda (\tau ,{t}_{0})\) will represent the fraction of the tracer-tagged water that makes its way from location \(\tau\) at time \({t}_{0}\) to the volume V at time T. Therefore, the adjoint tracer distribution provides information on the model tracer history and identifies origin and pathways of the model tracer-tagged water masses in the past.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Shulman, I., Cohen, J.H., Moline, M.A. et al. Bioluminescence potential during polar night: impact of behavioral light sensitivity and water mass pathways. Ocean Dynamics 72, 775–784 (2022). https://doi.org/10.1007/s10236-022-01533-9
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10236-022-01533-9