
Abstract
Intense anthropogenic impacts in tidal rivers can cause habitat loss and ecosystem degradation. In addition, changes in water temperature associated with climate change are significantly impacting the distribution area of fish fauna within tidal rivers. In the present study, we used long-term fish fauna data to determine the relationship between climate change-induced increases in water temperature and changes in the distribution of fish species in tidal rivers in the Japanese archipelago. The distribution ranges of many subtropical and tropical fish species were found to move northward in areas affected by warm currents, suggesting further possible distributional dispersal in future. This study is the first to examine the nationwide distributional changes and future projections of fish fauna in tidal rivers. The results suggest that many subtropical and tropical fishes are expanding their distribution areas in tidal rivers and in coastal and estuarine areas.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Estuaries are ecosystems located at the boundary between land and sea, and thus, demonstrate characteristics of both freshwater and seawater. These areas are subject to constantly changing periodic tides, waves, and river flows (Dyer 1997; Schröder-Adams et al. 2014; Barletta et al. 2017). Tidal rivers are important habitats not only for organisms that are endemic to brackish waters but also for marine and freshwater organisms (Sousa et al. 2007; Sheaves and Johnston 2008; Whitfield et al. 2012). In addition, tidal rivers are extremely important habitats for juvenile fishes due to high food availability and low predation pressure (Haedrich 1983; da Silva et al. 2018). Further, estuarine and coastal ecosystems, including tidal rivers, provide various ecosystem services, such as coastal ecosystem conservation, water purification, carbon sequestration, fishery resources, raw materials, and ecotourism, all of which contribute to human well-being (Costanza et al. 1997; Sloterdijk et al. 2017). However, estuarine and coastal areas are more susceptible to anthropogenic impacts due to their several ecosystem services (Dafforn et al. 2012; Barletta and Lima 2019), consequently, resulting in environmental issues, such as water quality deterioration, habitat loss, and resource degradation (Zhao et al. 2018; Brophy et al. 2019; Elliott et al. 2019).
Biodiversity loss due to anthropogenic impacts may be further accelerated by climate change; therefore, the effects of climate change on biodiversity have been extensively studied (Nunez et al. 2019; Weiskopf et al. 2020). Recent studies on ecosystem responses to climate change, conducted in different terrestrial and aquatic ecosystems have reported biota changes, life history shifts, and biogeographical shifts (Beaugrand 2009; Pasquaud et al. 2012; Piñeiro-Corbeira et al. 2018).
Similar studies have been conducted on the effects of climate change on habitats and biota in estuarine and coastal areas. For example, climate change is expected to increase the proportion of estuarine nutrients through increased river flows, increased coastal storm activity, and changes in phytoplankton productivity, with the effects being more pronounced in the subarctic zone, where permafrost has been decreasing (Statham 2012). Across the UK, with increasing temperatures, many coastal reef species are moving northward, particularly the invasive oyster Crassostrea gigas, which shows an expanding distribution (Mieszkowska et al. 2014). The 22-year increase in fish species richness in the North Sea, which is associated with warmer water temperatures, confirmed that species richness might increase at temperate latitudes due to the interactions between large-scale biogeographic patterns and climate change (Hiddink and Ter Hofstede 2008). Although the response of seagrass beds (main ecosystem type in coastal areas) to climate change remains unclear, seagrass bed loss rate has been severe, accelerating from approximately 1% yr−1 before 1940 to 7% yr−1 in the 1990s (Waycott et al. 2009). Seagrass beds in Europe have declined dramatically by 50–80% over the past century (Short and Wyllie-Echeverria 1996), and increasing CO2 concentrations and water temperatures due to climate change are expected to lead to further severe conditions (Koch et al. 2013; Robins et al. 2016).
Owing to the strong influence of freshwater inflows, changes in the habitats and fish fauna due to climate change in tidal rivers and coastal and estuarine areas differ. Climate change-induced sea level rise and reduced river discharge will increase salinity and significantly affect tidal rivers and freshwater intertidal ecosystems (Prandle and Lane 2015; Little et al. 2017). Moreover, the climate change-induced changes fish fauna of estuary will significantly impact human communities that are dependent on these resources. Hence, given that changes in coastal fishery resources also affect the establishment of marine protected areas or economic activities, understanding the effects of climate change on estuaries is important to adequately inform ecosystem conservation and management and to promote adaptation (Roessig et al. 2004; Gillanders et al. 2022). Although tidal rivers have been studied to evaluate the overall effects of climate change (Gillanders et al. 2011; Robins et al. 2016), studies on the effects of climate change based on long-term monitoring on a nationwide scale have not yet been conducted. Understanding the influence of climate change on the biota of tidal rivers, which are exposed to diverse anthropogenic pressures, is gaining attention.
The effects of climate change on the fish fauna of estuaries have been reported for the Brazilian coast (Araújo et al. 2018), San Francisco Estuary, California, USA (Feyrer et al. 2015), Gulf of Mexico (Fodrie et al. 2010; Gericke et al. 2013), English and Bristol Channel (Genner et al. 2004; Henderson 2007), Long Island Sound, USA (Howell and Auster 2012), South Africa (James et al. 2008, 2013, 2016), Australia (Koehn et al. 2011), and the Atlantic European seaboard, from Portugal to Scotland (Nicolas et al. 2011). However, aside from regional reports, no similar studies have investigated the Japanese archipelago, which serves as the habitat for tropical, subtropical, arctic, and subarctic fish species in addition to temperate zone species. There are also very few reports of predicted changes in distribution areas in response to changes in water temperature (Henderson 2007).
In order to elucidate the effects of climate change on tidal rivers, it is necessary to use a wide range of long-term data to identify areas undergoing significant changes, make accurate future projections, and establish priority areas for ecosystem conservation and resource management. For this purpose, it is important to identify species that are clearly affected by changes in water temperature and to model their distributions based on long-term changes. In this study, the effects of climate change on the distribution of fish fauna in tidal rivers were determined using fish fauna data (114 families and 337 species) collected from 1992 to 2019 for 110 tidal rivers located in the Japanese archipelago. Additionally, future occurrences and distributions of the fish fauna were predicted. To the best of our knowledge, this is the first study to examine changes in the distribution areas and predict the occurrence of fish species within tidal rivers of the Japanese archipelago over multiple climatic zones as well as the occurrence of arctic, subarctic, temperate, subtropical, and tropical fish species.
Methods
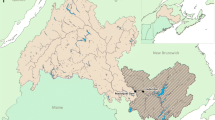
Four major ocean currents in the seas around the Japanese archipelago play an important role in the formation of fish fauna in coastal, coastal estuarine, and tidal rivers. They are the Kuroshio Current, which enters the Pacific Ocean from the East China Sea and flows northward along the Japanese archipelago coast; the Tsushima Current, which comprises water from the Kuroshio Current and coastal waters from the East China Sea that flow into the Sea of Japan through the Tsushima Strait; the Kuril Current, which flows from the North Pacific Ocean and the Sea of Okhotsk; and the Liman Current, a cold current that flows southward from near the Mamiya Strait along the Eurasian continent to the Sea of Japan (Kume et al. 2021) (Fig. 1). The Japanese archipelago also has a delicately complex coastal topography, with a long coastline of approximately 35,000 km. The topographic complexity and the diverse ocean currents are majorly responsible for the diverse estuarine biota (Itsukushima 2019).

Location of the study sites. The coasts of the Japanese archipelago selected in this study are surrounded by four major currents (Kuroshio, Tsushima, Kurile, and Liman currents). Fish fauna data were obtained from surveys on 110 tidal rivers spanning from 1992 to 2019
To investigate the effects of climate change on the fish fauna of tidal rivers, fish fauna data from 110 tidal rivers were analyzed in the Japanese archipelago (coordinates: 31.3576 and 44.922 Latitude; 129.993 and 144.366 Longitude) through regular surveys of the fish fauna in selected tidal rivers. Fish fauna data were acquired from the results of the National Census on River Environments (1992–2019) conducted by the Ministry of Land, Infrastructure, Transport, and Tourism. Surveys were conducted at approximately five-year intervals for each river, and the corresponding results were divided into three time periods, 1990s (1992–2000), 2000s (2001–2010), and 2010s (2011–2019), to examine changes in the species occurrence. Further, 1–3 surveys were conducted for each tidal river in the targeted age groups. Only species identified to the species level were included in the analysis.
Water temperature data from tidal rivers and coastal areas were compiled to investigate the relationship between fish fauna and water temperature. Data from the most downstream water temperature survey station in each watershed were used for the tidal rivers, and data from the station closest to each tidal river were used for the coastal area. The maximum and minimum observation periods were 58 and 20 years for the tidal rivers and 39 and 25 years for the coastal area, respectively. The annual mean temperature was calculated from the monthly mean temperature using arithmetic mean. The data source for calculating the water temperature was obtained from Ministry of Environment (https://www.nies.go.jp/igreen/) and Japan Meteorological Agency (https://www.data.jma.go.jp/gmd/kaiyou/data/db/kaikyo/dbindex_jpn.html). Subsequently, Mann–Kendall rank statistics were conducted to identify the trends in water temperature changes. Data with missing periods were interpolated using linear interpolation. The interpolated missing periods comprised only 1.8% of the total observation period. In areas, where the Mann–Kendall rank statistics indicated significant changes in water temperature (p < 0.05), the slope of the linear regression equation between the observation year and water temperature was calculated and used to assess the annual amount of water temperature change.
The tidal reach length of the 110 targeted rivers was 9.32 ± 11.25 km (average ± standard deviation), the distance from the mouth of the river to the point where water temperature was measured was 2.74 ± 2.79 km, and the distance from the mouth of the river to the midpoint of the fish survey area was 1.63 ± 1.86 km.
Furthermore, a habitat probability model was constructed using logistic regression analysis for fish species inhabiting tidal rivers with significant changes in the water temperature identified through trend analysis. Water temperature of the tidal rivers was used as the explanatory variable and the presence/ absence of fish habitat was considered as the objective variable. The objective variable comprised the presence/absence data for the 1990s, 2000s, and 2010s for each tidal river. The goodness of fit of the model was determined by calculating the AUC value by drawing a receiver operating characteristic curve using the objective variable of the logistic regression and the calculated predicted probability. Further, the occurrence probability of the target fish species in 2070 and 2120 was calculated using logistic regression equations for species considering a p-value of less than 0.05 for the tidal river temperature in the logistic regression analysis and an AUC value of 0.7 or greater. Lastly, water temperatures in the tidal rivers in 2070 and 2120 were calculated based on linear regression equations.
Results
In this study, 580 surveys of fish fauna in the 110 tidal rivers in Japan were conducted between 1992 and 2019. Consequently, 337 estuarine and marine fish species (only those identified to the species level were considered, except for 32 genera that were only identified to the genus level) were identified. The top six families with the highest number of species were Gobiidae (64 species), Tetraodontidae (15 species), Carangidae (13 species), Syngnathidae (11 species), Pleuronectidae (10 species), and Cottidae (10 species). These families comprised 123 species, accounting for 36.5% of the total number of species.
Further, 220, 247, and 235 species were observed for the 1990s, 2000s, and 2010s, respectively. Tropical and subtropical species, such as the Hippichthys penicillus, Acanthopagrus latus, and Lutjanus argentimaculatus, mainly expanded their distribution area. In the 1990s, the distribution areas, such as the southern coasts of Kyushu and Shikoku, were strongly influenced by the Kuroshio Current due to its high water temperature; however, the distribution areas expanded over time (Fig. 2a and b). Thirty-three new fish species were identified during the 2010s survey, with a major proportion of tropical and subtropical species (23), followed by temperate species (7), and cold/subarctic species (3). In addition, the number of newly confirmed fish species in each tidal river was examined. Moreover, in 25 of the targeted tidal rivers, several fish species were confirmed for the first time, with many of the newly confirmed species found in tidal rivers flowing into the coastal areas of the Kuroshio Current, especially in Kyushu and Shikoku. The largest number of the newly confirmed fish species (seven species) was found at the mouth of the Watari River, southwest of Shikoku (Fig. 3).

(a) Temporal changes in the distribution of Hippichthys penicillus (Cantor, 1849). The status of confirmed inhabitation is shown by age. Among the 110 targeted tidal rivers, inhabitation was confirmed in 40 tidal rivers through the survey. In the 1990s, this species was distributed only in some parts along the Pacific Ocean in southwestern Japan; however, its distribution area expanded, and in the 2000s, it was confirmed to be present in the Seto Inland Sea and the Sea of Japan side. (b) Temporal changes in the distribution of Acanthopagrus latus (Houttuyn, 1782). The status of confirmed inhabitation is shown by age. Among the 110 targeted tidal rivers, inhabitation was confirmed in 50 tidal rivers through the survey. In the 1990s, the distribution of this species was limited to tidal rivers on the Shikoku side of the Seto Inland Sea; however, in the 2000s, its presence was confirmed in almost all areas of the Seto Inland Sea and Kyushu. In the 2010s, the species distribution also expanded into the Sea of Japan.

Number of newly confirmed species in 2010s. The seven new species were most abundant in the Watari River on the southern coast of Shikoku, which is strongly influenced by warm currents. Subtropical and tropical species, including Bathygobius fuscus, were newly identified in the Watari River. Similarly, many newly distributed species were found in tidal rivers influenced by warm currents. Conversely, three new species were also observed in tidal rivers along the Sea of Japan. The newly identified species (Pseudopleuronectes obscurus) in Hokkaido is a cold/subarctic species
The Mann–Kendall rank statistics showed significant increases in the annual mean water temperatures of tidal rivers. Further, the annual mean water temperature increased significantly in 82 rivers of the coastal zone. A significant upward trend was observed in 59 areas with tidal rivers and coastal areas, and in 19 tidal rivers, and 24 coastal areas. Conversely, the water temperature did not significantly increase in only eight of the tidal rivers and coastal areas. Areas with significant increases in both tidal rivers and coastal areas were particularly abundant in southwestern Japan and along the coast of the Sea of Japan. However, tidal rivers in non-coastal areas of warm currents, such as Hokkaido and Tohoku, did not show an increasing trend in the annual mean water temperature (Fig. 4). Furthermore, the average value of the maximum annual mean increase in water temperature (slope of the regression line) was 0.039, and 0.072 for the largest river (Yamato River).

Increasing trends in tidal rivers and coastal water temperatures in the target sites. The results of the Mann–Kendall rank statistics with a significance level of p < 0.05 indicate a trend of increasing water temperature. Although an increasing trend was observed nationwide, the temperatures of both tidal rivers and coastal water were increasing in many tidal rivers, especially in southwestern Japan and along the coast of the Sea of Japan, while the tidal river temperatures in certain areas of Hokkaido and Tohoku did not increase
Logistic regression analysis was used to examine the relationship between water temperature of tidal rivers and the occurrence of fish species. The corresponding results showed that 72 species showed a significant relationship (p < 0.05) with the annual mean water temperature in tidal rivers. Among these species, the probability of occurrence of 58 species increased with increasing water temperature, whereas that of 14 species decreased. Further, out of the 72 species, the probability of occurrences of the 46 species that were found to have a strong relationship (p < 0.01) with water temperature, and an area under curve (AUC) of 0.7 or higher, were estimated for 2070 and 2120 using logistic regression equations. H. penicillus, which has been expanding its distribution range over the past 30 years, may further greatly expand its distribution range to 17 new tidal rivers by 2070 (50% or greater probability of inhabitation) and to 11 tidal rivers by 2120 (50% or greater probability of inhabitation) (Fig. 5a). The probability of occurrence for A. latus, which has already expanded its distribution area in southwestern Japan, was more than 50% in the tidal rivers flowing into Tokyo Bay and in the western part of the Sea of Japan by 2070, and in some tidal rivers in northeastern Japan by 2120 (Fig. 5b). In addition, tropical and subtropical species, such as Sphyraena barracudaa and Elops hawaiensis, which currently occur only in few tidal rivers, showed a greater than 50% probability of occurrence by 2120 in many tidal rivers. However, the probability of occurrence of cold-water fish species declined significantly, and Oncorhynchus keta, which was recorded in 34 tidal rivers through previous surveys, will have less than 50% probability of occurrence in 12 of the 34 tidal rivers by 2070.

(a) Predicted occurrence of Hippichthys penicillus (Cantor, 1849) in 2070 and 2120. Tidal rivers with a significant increase in the temperature of tidal rivers were plotted with the probability of occurrence, exceeding the 50% threshold annually. The species is currently distributed in southwestern Japan, excluding parts of the Sea of Japan; however, it will expand its distribution to the Sea of Japan by 2070 and may appear in the Tohoku region by 2120. (b) Predicted occurrence of Acanthopagrus latus (Houttuyn, 1782) in 2070 and 2120. Tidal rivers with a significant increase in the temperature of the tidal rivers were plotted with the probability of occurrence exceeding the 50% threshold annually. The species is widely distributed in southwestern Japan; however, by 2070 its distribution may expand to almost all areas of southwestern Japan as well as the Sea of Japan and Tokyo Bay.
Discussion
Changes in the fish fauna of estuaries and coasts due to climate change-induced increases in sea surface temperatures have been reported in many parts of the world. For instance, in southeastern Brazil, located in the transition zone between the tropics and subtropics, 40 years of monitoring has revealed that populations of tropical fish species are increasing, while the number of subtropical species is declining, or they are disappearing (Araújo et al. 2018). Meanwhile, within the northern Gulf of Mexico, an increase in the number of tropical and subtropical fish species has also been observed, suggesting a relationship between temperature and sea level rise (Fodrie et al. 2010; Gericke et al. 2013). Similarly in South Africa, which is located in the temperate zone, tropical fish species have been established since ~ 2000 (James et al. 2008; 2016). Additionally, studies in colder zones have reported a decrease in the number of cold-adapted species along with an increase in that of subtropical fish species (Genner et al. 2004; Howell and Auster 2012). In fact, within Bridgewater Bay, UK, a 2 °C increase in coastal seawater temperatures is projected to increase the total number of fish species by 10% due to an increase in warm water tourists (Henderson 2007).
The current study involved 337 species that occur in the 110 tidal rivers in the Japanese archipelago. Habitat classification, based on existing literature and information related to distribution areas (Kimura et al. 2013; Kawano et al. 2014), revealed that 145 (43.0%) of these species were tropical and subtropical, 160 (47.5%) were temperate, 31 (9.2%) were arctic and subarctic, and 1 (0.3%) was a deep-sea species. Among the species analyzed in this study, tropical/subtropical species, such as Scatophagus argus, Redigobius bikolanus, and H. penicillus have been reported to follow a northward expansion along the Pacific coast (Kimura et al. 1997; Kudo 2011; Yamakawa et al. 2018, 2020). However, spatiotemporal constraints, including local regional studies, have limited the assessment of distribution expansion in multiple regions. In this study, changes in the distribution area of estuarine and marine fishes on a nationwide scale were determined using long-term data from tidal rivers in the Japanese archipelago; additionally, by constructing a habitat model using water temperatures in tidal rivers, the possibility of future distribution was identified.
Although the Kuroshio Current is a source of tropical and subtropical species that invade coastal waters in areas influenced by warm currents around the Japanese archipelago (Tashiro et al. 2017), few reports exist on the changes in fish fauna distribution in tidal rivers associated with increasing water temperatures, except in some areas. In addition, the Sea of Japan is originally known to have a poorer fish fauna distribution than the surrounding marine areas possibly because of the shallow depth of the Tsushima Strait, which prevents the entry of mesopelagic fish species that inhabit the Pacific Ocean (Kawano et al. 2014). However, in the southernmost part of the Japan Sea, tropical and subtropical species have increased rapidly since the late 1990s (Kobayashi et al. 2006). However, the distribution of these tropical and subtropical species has not rapidly increased in tidal rivers in various rivers along the Sea of Japan (Fig. 3). Further, various rivers along the Sea of Japan are experiencing an increase in the water temperature in tidal rivers and coastal areas (Fig. 4), which may promote northward migration of tropical and subtropical species that have already invaded the area. In addition, as changes in the distribution areas of estuarine and marine fish have only been observed in the coastal areas, collecting information on the coastal-estuarine-tidal rivers areas is essential that can assist in determining whether the species follow abortive migration or colonization.
Changes in the fish distribution patterns in many ocean regions (Last et al. 2011) and species temporal turnover (Albouy et al. 2012) due to climate change-induced temperature changes have been reported. However, although a northward expansion of tropical and subtropical species was reported in this study, disturbance of native tidal river ecosystems by the invasion of these species, such as fish fauna replacement, has not been observed possibly because areas affected by warm currents are originally inhabited by tropical, subtropical, and temperate species, and the global warming effects are less pronounced. Additionally, the Pacific coastal areas, showing northward migration of tropical and subtropical species, are less susceptible to the introduction of tropical and subtropical species due to the abundance of biota and robust native ecosystems.
Conversely, the temperatures in areas affected by cold currents are not yet tolerable for the reproduction of tropical and subtropical species, and thus, the effects of climate change are expected to be limited. In addition, most targeted tidal rivers have been subjected to anthropogenic impacts, such as land reclamation, land use change, revetment, and dam construction (Yoshimura et al. 2005), and their influences are larger than the influence of water temperature increase due to climate change. However, as climate change worsens its effects on ecosystems that are already affected by human activities (Schindler 2001; Graham and Harrod 2009), monitoring the changes in these ecosystems is necessary. In some tidal rivers, changes in water temperature due to climate change cannot be neglected; for example, in the Tsurumi River, where 85% of the watershed is urbanized, the annual average water temperature change is remarkably high (0.06 °C/year); consequently, the fish fauna may change dramatically in a short period. Therefore, ecosystem conservation and resource management should be implemented using continuous monitoring.
Of the 337 species covered in this study, the Gobiidae, a major family in the tidal rivers of the Japanese archipelago, accounted for the largest number of species. Among the 64 species of Gobiidae confirmed in this study, 61 are considered tropical, tropical and subtropical, or temperate species. Distribution areas of Redigobius bikolanus and Rhinogobius similis have been confirmed to have expanded since the 1990s. Regional reports also describe a northward trend for Glossogobius biocellatus and Callogobius tanegasimae (Kobayashi et al. 2006; Last et al. 2011). These goby species are likely to be found in more rivers in the future if the trend of increasing water temperatures continues. In addition, some species of Gobiidae have been reported to expand their distribution range worldwide due to wide temperature tolerances and competition (Cross and Rawding 2009; Chargulaf 2010). For example, Neogobius melanostomus has expanded its brackish habitat over the past decade, primarily in North America and Europe, and there is concern that climate change will further expand its distribution range (Kornis et al. 2012; Kotta et al. 2016). No non-native gobies were identified in this study, and the species tending to move northward are native species, however, it is quite possible that non-native species could be introduced to the tidal rivers of the Japanese archipelago through ballast waters or by other means. Therefore, future research should also include monitoring non-native fish species because species with a high water temperature tolerance and high competitiveness may rapidly expand their distribution areas and affect native ecosystems.
Observations from past decades have shown that many of the world's coastal and estuarine ecosystems are continuously changing, and that the rate of change is faster than expected (Cloern et al. 2016). In addition to changes in water temperature, the effects of climate change are diverse, including changes in freshwater inflows from land, sea level rise, the magnitude and frequency of storm surges, and phytoplankton productivity (Statham 2012; Robins et al. 2016). Indeed, factors other than temperature change can significantly impact species range shifts and, if not considered, the effects of climate change on species populations and ecosystem services will be grossly underestimated (Lauchlan and Nagelkerken 2019). More specifically, increased freshwater runoff in watersheds due to climate change has been shown to significantly impact estuarian fish fauna, with demersal and pelagic fish community structures detected in the oligohaline, mesohaline, polyhaline, and euhaline guilds (Feyrer et al. 2015). It has also been suggested that estuarine habitats will become significantly degraded by rises in sea level (James et al. 2013). However, it is also necessary to consider anthropogenic stressors and climate change scenarios; that is, climate change effects on the estuary ecosystem are compounded by other anthropogenic impacts, such as land use, water resource use, and dam construction (Toft et al. 2018). Hence, it is essential to predict the response of estuarian fish fauna to combined impacts, to effectively anticipate changes, and support management actions that consider diverse actors, including fisheries, environmental, socioeconomic and political layers (Koehn et al. 2011).
Conclusion
This study determined the relationship between rising water temperatures on a nationwide scale and changes in the distribution of fish in tidal rivers using long-term (1992–2019) fish fauna data. The distribution of 337 brackish and saltwater fish species in 110 rivers in the Japanese archipelago over the past 30 years revealed that tropical and subtropical species such as the H. penicillus, A. latus, and L. argentimaculatus, have expanded their distribution areas. In the 2010s, 33 new fish species were identified, 23 of which were tropical and subtropical species, indicating an expansion of the distribution range of tropical and subtropical species. The Mann–Kendall rank statics showed that the annual mean temperature of tidal rivers was found in 76 rivers, with a marked upward trend in tidal rivers affected by warm currents and tidal rivers with significant urbanization. Furthermore, future occurrence forecasts for species that are strongly associated with tidal rivers water temperatures indicate that the probability of occurrence of cold-water fish species will decrease significantly, while the further northward expansion of H. penicillus and A. latus, which have been expanding their distribution areas over the past 30 years, is predicted. The distribution ranges of many subtropical and tropical fish species were found to move northward in areas affected by warm currents, suggesting further possible distributional dispersal in future. Hence, collectively, this study examines the nationwide distributional changes and future projections of fish fauna in tidal rivers. The results suggest that many subtropical and tropical fishes are expanding their distribution areas in tidal rivers and in coastal and estuarine areas.
Data and materials availability
Data are available upon request. Data have been generated at the Tokyo Institute of Technology and are available from the corresponding author RI on request.
References
Albouy C, Guilhaumon F, Araújo MB, Mouillot D, Leprieur F (2012) Combining projected changes in species richness and composition reveals climate change impacts on coastal Mediterranean fish assemblages. Glob Change Biol 18:2995–3003. https://doi.org/10.1111/j.1365-2486.2012.02772.x
Araújo FG, Teixeira TP, Guedes AP, Azevedo MC, Pessanha AL (2018) Shifts in the abundance and distribution of shallow water fish fauna on the southeastern Brazilian coast: a response to climate change. Hydrobiologia 814:205–218. https://doi.org/10.1007/s10750-018-3537-8
Barletta M, Lima AR, Costa MF, Dantas DV (2017) Estuarine ecoclines and the associated fauna: Ecological information as the basis for ecosystem conservation. In: Finkl C, Makowski C (eds) Coastal wetlands: Alteration and remediation. Coastal research library 21, Springer Cham. https://doi.org/10.1007/978-3-319-56179-0_16
Barletta M, Lima AR (2019) Systematic review of fish ecology and anthropogenic impacts in South American estuaries: Setting priorities for ecosystem conservation. Front Mar Sci 6:237. https://doi.org/10.3389/fmars.2019.00237
Beaugrand G (2009) Decadal changes in climate and ecosystems in the North Atlantic ocean and adjacent seas. e 56:656–673. https://doi.org/10.1016/J.DSR2.2008.12.022
Brophy L, Greene CM, Hare VC, Holycross B, Lanier A et al (2019) Insights into estuary habitat loss in the western United States using a new method for mapping maximum extent of tidal wetlands. PLoS ONE 14:e0218558. https://doi.org/10.1371/journal.pone.0218558
Chargulaf CA (2010) The ecology of soft sediment tidepool dwelling gobies (Pisces: Gobiidae) in Moreton Bay. The University of Queensland, Australia
Cloern JE, Abreu PC, Carstensen J, Chauvaud L, Elmgren R et al (2016) Human activities and climate variability drive fast-paced change across the world’s estuarine-coastal ecosystems. Glob Change Biol 22:513–529. https://doi.org/10.1111/gcb.13059
Costanza R, D’arge RC, Groot RD, Farber SB, Grasso M et al (1997) The value of the world’s ecosystem services and natural capital. Nature 387:253–260. https://doi.org/10.1038/387253a0
Cross EE, Rawding R (2009) Acute thermal tolerance in the round goby, Apollonia melanostoma (Neogobius melanostomus). J Therm Biol 34:85–92. https://doi.org/10.1016/J.JTHERBIO.2008.10.006
da Silva RS, Baeta A, Pessanha AL (2018) Are vegetated areas more attractive for juvenile fish in estuaries? A comparison in a tropical estuary. Environ Biol Fishes 101:1427–1442. https://doi.org/10.1007/s10641-018-0790-7
Dafforn KA, Simpson SL, Kelaher BP, Clark GF, Komyakova V et al (2012) The challenge of choosing environmental indicators of anthropogenic impacts in estuaries. Environ Pollut 163:207–217. https://doi.org/10.1016/j.envpol.2011.12.029
Dyer KR (1997) Estuaries, a physical introduction, 2nd edn. John Wiley & Sons, Chichester
Elliott M, Day JW, Ramachandran R, Wolanski EA (2019) Synthesis: What is the future for coasts, estuaries, deltas and other transitional habitats in 2050 and beyond? Coasts and Estuaries:1–28. https://doi.org/10.1016/B978-0-12-814003-1.00001-0
Feyrer F, Cloern JE, Brown LR, Fish M, Hieb KA et al (2015) Estuarine fish communities respond to climate variability over both river and ocean basins. Glob Change Biol 21:3608–3619. https://doi.org/10.1111/gcb.12969
Fodrie FJ, Heck KL, Powers S, Graham WM, Robinson KL (2010) Climate-related, decadal-scale assemblage changes of seagrass-associated fishes in the northern Gulf of Mexico. Glob Change Biol 16:48–59. https://doi.org/10.1111/j.1365-2486.2009.01889.x
Genner MJ, Sims DW, Wearmouth VJ, Southall EJ, Southward AJ et al (2004) Regional climatic warming drives long–term community changes of British marine fish. P Roy Soc Lond B Bio 271:655–661. https://doi.org/10.1098/rspb.2003.2651
Gericke RL, Heck KL, Fodrie FJ (2013) Interactions between Northern-Shifting Tropical Species and Native Species in the Northern Gulf of Mexico. Estuar Coast 37:952–961. https://doi.org/10.1007/s12237-013-9733-x
Gillanders BM, Elsdon TS, Halliday IA, Jenkins GP, Robins JB et al (2011) Potential effects of climate change on Australian estuaries and fish utilising estuaries: a review. Mar Freshw Res 62:1115–1131. https://doi.org/10.1071/MF11047
Gillanders BM, McMillan MN, Reis-Santos P, Baumgartner LJ, Brown LR et al (2022) Climate change and fishes in estuaries. In: Whitfield AK, Able KW, Stephen JM, Elliott M (eds) Fish and fisheries in estuaries: A global perspective. Wiley, New York, pp 380–457. https://doi.org/10.1002/9781119705345.ch7
Graham CT, Harrod C (2009) Implications of climate change for the fishes of the British Isles. J Fish Biol 74:1143–1205. https://doi.org/10.1111/j.1095-8649.2009.02180.x
Haedrich RL (1983) Estuarine fishes. In: Ketchum B (ed) Ecosystems of the world: estuaries and enclosed seas. Elsevier, Amsterdam, pp 183–207
Henderson PA (2007) Discrete and continuous change in the fish community of the Bristol Channel in response to climate change. J Mar Biol Assoc UK 87:589–598. https://doi.org/10.1017/S0025315407052447
Hiddink JG, ter Hofstede R (2008) Climate induced increases in species richness of marine fishes. Glob Change Biol 14:453–460. https://doi.org/10.1111/j.1365-2486.2007.01518.x
Howell PT, Auster PJ (2012) Phase Shift in an Estuarine Finfish Community Associated with Warming Temperatures. Mar Coastal Fisheries: Dyn Manage Ecosys Sci 4:481–495. https://doi.org/10.1080/19425120.2012.685144
Itsukushima R (2019) Study of aquatic ecological regions using fish fauna and geographic archipelago factors. Ecol Indic 96:69–80. https://doi.org/10.1016/J.ECOLIND.2018.08.057
James NC, Whitfield AK, Cowley PD (2008) Preliminary indications of climate-induced change in a warm-temperate South African estuarine fish community. J Fish Biol 72:1855–1863. https://doi.org/10.1111/J.1095-8649.2008.01850.X
James NC, Niekerk LV, Whitfield AK, Potts WM, Götz A et al (2013) Effects of climate change on South African estuaries and associated fish species. Clim Res 57:233–248. https://doi.org/10.3354/CR01178
James NC, Whitfield AK, Harrison TD (2016) Grey mullet (Mugilidae) as possible indicators of global warming in South African estuaries and coastal waters. Mar Environ Res 122:188–195. https://doi.org/10.1016/j.marenvres.2016.11.002
Kawano M, Miyake H, Hoshino N, Ito K, Yamanaka T et al (2014) List of fishes in the Japan sea. Bull Yamaguchi Prefectural Fish 11:1–30
Kimura K, Hagiwara K, Nakane M (1997) Distributional notes on five species of freshwater fishes in Kanagawa prefecture. Natural History Report of Kanagawa 18:79–82
Kimura T, Abe K, Matsumoto T (2013) Benthic environment and fish assemblages at the water in front of Yokonami Rinkai Experimental Station in Kochi, western Japan. Kuroshio Science 6:194–206
Kobayashi T, Hori S, Hiroyuki D, Kawano M (2006) Noteworthy phenomena on the marine organisms in the coastal region of Yamaguchi Prefecture, southwestern Japan Sea. Bull Yamaguchi Prefectural Fish 4:19–56
Koch MS, Bowes G, Ross C, Zhang XH (2013) Climate change and ocean acidification effects on seagrasses and marine macroalgae. Glob Change Biol 19:103–132. https://doi.org/10.1111/j.1365-2486.2012.02791.x
Koehn JD, Hobday AJ, Pratchett MS, Gillanders BM (2011) Climate change and Australian marine and freshwater environments, fishes and fisheries: synthesis and options for adaptation. Mar Freshwater Res 62:1148–1164. https://doi.org/10.1071/MF11139
Kornis MS, Mercado-Silva N, Vander Zanden MJ (2012) Twenty years of invasion: a review of round goby Neogobius melanostomus biology, spread and ecological implications. J Fish Biol 80:235–285. https://doi.org/10.1111/j.1095-8649.2011.03157.x
Kotta J, Nurkse K, Puntila R, Ojaveer H (2016) Shipping and natural environmental conditions determine the distribution of the invasive non-indigenous round goby Neogobius melanostomus in a regional sea. Estuar Coast Shelf Sci 169:15–24. https://doi.org/10.1016/J.ECSS.2015.11.029
Kudo T (2011) New records of fishes from the coasts of the Yokohama, Kawasaki and Nakanose areas, Tokyo Bay – V. Natural History Report of Kanagawa 32:127–133. https://doi.org/10.32225/nkpmnh.2011.32_127
Kume M, Lavergne E, Ahn H, Terashima Y, Kadowaki K et al (2021) Factors structuring estuarine and coastal fish communities across Japan using environmental DNA metabarcoding. Ecol Indic 121:107216. https://doi.org/10.1016/j.ecolind.2020.107216
Last PR, White WT, Gledhill DC, Hobday AJ, Brown R et al (2011) Long-term shifts in abundance and distribution of a temperate fish fauna: A response to climate change and fishing practices. Glob Ecol Biogeogr 20:58–72. https://doi.org/10.1111/j.1466-8238.2010.00575.x
Lauchlan SS, Nagelkerken I (2019) Species range shifts along multistressor mosaics in estuarine environments under future climate. Fish Fish 21:32–46. https://doi.org/10.1111/faf.12412
Little S, Wood PJ, Elliott M (2017) Quantifying salinity-induced changes on estuarine benthic fauna: The potential implications of climate change. Estuarine Coast Shelf Sci 198:610–625. https://doi.org/10.1016/J.ECSS.2016.07.020
Mieszkowska N, Firth LB, Bentley MG (2014) Impacts of climate change on intertidal habitats. MCCIP Sci Rev:180–192. https://doi.org/10.14465/2013.arc19.180-192
Nicolas D, Chaalali A, Drouineau H, Lobry J, Uriarte A et al (2011) Impact of global warming on European tidal estuaries: Some evidence of northward migration of estuarine fish species. Reg Environ Change 11:639–649. https://doi.org/10.1007/S10113-010-0196-3
Nunez S, Arets E, Alkemade R, Verwer C, Leemans R (2019) Assessing the impacts of climate change on biodiversity: Is below 2 °C enough? Clim Change 154:351–365. https://doi.org/10.1007/s10584-019-02420-x
Pasquaud S, Béguer M, Larsen MH, Chaalali A, Cabral HN et al (2012) Increase of marine juvenile fish abundances in the middle Gironde estuary related to warmer and more saline waters, due to global changes. Estuarine Coast Shelf Sci 104–105:46–53. https://doi.org/10.1016/J.ECSS.2012.03.021
Piñeiro-Corbeira C, Barreiro R, Cremades J, Arenas F (2018) Seaweed assemblages under a climate change scenario: Functional responses to temperature of eight intertidal seaweeds match recent abundance shifts. Sci Rep 8:12978. https://doi.org/10.1038/s41598-018-31357-x
Prandle D, Lane A (2015) Sensitivity of estuaries to sea level rise: Vulnerability indices. Estuarine Coast Shelf Sci 160:60–68. https://doi.org/10.1016/J.ECSS.2015.04.001
Robins PE, Skov MW, Lewis MJ, Giménez L, Davies AG et al (2016) Impact of climate change on UK estuaries: A review of past trends and potential projections. Estuarine Coast Shelf Sci 169:119–135. https://doi.org/10.1016/J.ECSS.2015.12.016
Roessig JM, Woodley CM, Cech JJ, Hansen LJ (2004) Effects of global climate change on marine and estuarine fishes and fisheries. Rev Fish Biol Fisher 14:251–275. https://doi.org/10.1007/s11160-004-6749-0
Schindler DW (2001) The cumulative effects of climate warming and other human stresses on Canadian freshwaters in the new millennium. Can J Fish Aquat Sci 58:18–29. https://doi.org/10.1007/978-1-4615-1493-0_11
Schröder-Adams CJ, Boyd RL, Tran T (2014) Estuarine foraminiferal biofacies pattern compared to the brackish ichnofacies model: Port Stephens, southeast Australia. Estuar Coast Shelf S 139:78–87. https://doi.org/10.1016/J.ECSS.2013.12.006
Sheaves M, Johnston R (2008) Influence of marine and freshwater connectivity on the dynamics of subtropical estuarine wetland fish metapopulations. Mar Ecol Prog Ser 357:225–243. https://doi.org/10.3354/MEPS07292
Short FT, Wyllie-Echeverria S (1996) Natural and human-induced disturbance of seagrasses. Environ Conserv 23:17–27. https://doi.org/10.1017/S0376892900038212
Sloterdijk H, Brehmer P, Sadio O, Müller H, Döring J et al (2017) Composition and structure of the larval fish community related to environmental parameters in a tropical estuary impacted by climate change. Estuarine Coast Shelf Sci 197:10–26. https://doi.org/10.1016/J.ECSS.2017.08.003
Sousa R, Antunes C, Guilhermino L (2007) Species composition and monthly variation of the molluscan fauna in the freshwater subtidal area of the River Minho estuary. Estuar Coast Shelf S 75:90–100. https://doi.org/10.1016/J.ECSS.2007.02.020
Statham PJ (2012) Nutrients in estuaries -an overview and the potential impacts of climate change. Sci Total Environ 434:213–227. https://doi.org/10.1016/j.scitotenv.2011.09.088
Tashiro F, Suzuki K, Ueno Y, Funakoshi Y, Ikeguchi S et al (2017) Biogeographic and taxonomic notes regarding the diversity of fish in the Sea of Japan. In: TAXA Research Center Aquarium Proceedings of the Japanese Society of Systematic Zoology, vol 42, pp 22–40. https://doi.org/10.19004/taxa.42.0_22
Toft JD, Munsch SH, Cordell JR, Siitari K, Hare VC et al (2018) Impact of multiple stressors on juvenile fish in estuaries of the northeast Pacific. Glob Change Biol e:2008–2020. https://doi.org/10.1111/gcb.14055
Waycott M, Duarte CM, Carruthers TJ, Orth RJ, Dennison WC et al (2009) Accelerating loss of seagrasses across the globe threatens coastal ecosystems. Proc Natl Acad Sci USA 106:12377–12381. https://doi.org/10.1073/pnas.0905620106
Weiskopf SR, Rubenstein MA, Crozier LG, Gaichas SK, Griffis RB et al (2020) Climate change effects on biodiversity, ecosystems, ecosystem services, and natural resource management in the United States. Sci Total Environ 733:137782. https://doi.org/10.1016/j.scitotenv.2020.137782
Whitfield AK, Elliott M, Basset A, Blaber SJM, West RJ (2012) Paradigms in estuarine ecology - A review of the Remane diagram with a suggested revised model for estuaries. Estuar Coast Shelf S 97:78–90. https://doi.org/10.1016/J.ECSS.2011.11.026
Yamakawa U, Mitsui S, Maruyama T, Kato S, Sakai S et al (2018) Notes on eighteen fish species recorded from the river and coastal areas of Sagami Bay and adjacent waters, Japan: Northward range extension of warm water fishes in recent years. Bull Kanagawa Prefect Mus (Natl Sci) 47:35–57. https://doi.org/10.32225/bkpmnh.2018.47_35
Yamakawa U, Mitsui S, Oda T, Morita Y, Aoki T et al (2020) Records of seven fish species showing a northward shift in distribution pattern in the Sagami Bay region, Japan. Natural History Report of Kanagawa 41:71–82. https://doi.org/10.32225/nkpmnh.2020.41_71
Yoshimura C, Omura T, Furumai H, Tockner K (2005) Present state of rivers and streams in Japan. River Res Appl 21:93–112. https://doi.org/10.1002/RRA.835
Zhao Y, Xu M, Liu Q, Wang Z, Zhao L et al (2018) Study of heavy metal pollution, ecological risk and source apportionment in the surface water and sediments of the Jiangsu coastal region, China: A case study of the Sheyang Estuary. Mar Pollut Bull 137:601–609. https://doi.org/10.1016/j.marpolbul.2018.10.044
Acknowledgements
This work was supported by JSPS KAKENHI (Grant Number: 19H02250).
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by Anne Bousquet-Melou
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Itsukushima, R. Effects of climate change-induced water temperature changes on the distribution of tidal river fish fauna in the Japanese archipelago. Reg Environ Change 23, 100 (2023). https://doi.org/10.1007/s10113-023-02098-z
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s10113-023-02098-z