
Abstract
Currently, there is a trend of increasing incidence in pulmonary non-tuberculous mycobacterial infections (PNTM) together with a decrease in tuberculosis (TB) incidence, particularly in developed countries. The prevalence of PNTM in underdeveloped and developing countries remains unclear as there is still a lack of detection methods that could clearly diagnose PNTM applicable in these low-resource settings. Since non-tuberculous mycobacteria (NTM) are environmental pathogens, the vicinity favouring host-pathogen interactions is known as important predisposing factor for PNTM. The ongoing changes in world population, as well as socio-political and economic factors, are linked to the rise in the incidence of PNTM. Development is an important factor for the improvement of population well-being, but it has also been linked, in general, to detrimental environmental consequences, including the rise of emergent (usually neglected) infectious diseases, such as PNTM. The rise of neglected PNTM infections requires the expansion of the current efforts on the development of diagnostics, therapies and vaccines for mycobacterial diseases, which at present, are mainly focused on TB. This review discuss the current situation of PNTM and its predisposing factors, as well as the efforts and challenges for their control.
Similar content being viewed by others

Introduction
Multiple bacterial agents are associated with pulmonary infections [1]; among them, non-tuberculous mycobacteria (NTM) have a rising incidence [2]. NTM are also known as mycobacteria other than tuberculosis, atypical mycobacteria or environmental mycobacteria [3]. NTM are labelled as environmental mycobacteria because they are widely distributed in the environment, such as in soil, marshland, streams, rivers, estuaries, dust, domestic and wild animals and food [4]. NTM are opportunistic pathogens which rarely cause disease in human unless host defence is impaired [5]. They are associated with disseminated and local infections in lungs, pleura, skin, eye, central nervous system, soft tissue, genitourinary system and lymph nodes, among others [6,7,8,9,10,11,12,13,14,15,16] (Table 1).
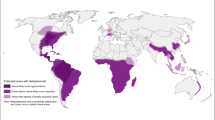
Pulmonary infection is the most common disease caused by NTM (PNTM) and it has substantially increased worldwide [28]. Based on the data published by Hoefsloot et al., (2013) from 30 countries across six continents, M. avium complex (MAC) (consisting of M. avium and M. intracellulare) is the most prevalent NTM found in respiratory samples, followed by M. gordonae and M. xenopi [29]. Among the six continents, the relative contribution of MAC per continent was highest in Australia (71.1%), followed by Asia (53.8%), North America (52.0%), South Africa (50.5%), Europe (36.9%) and South America (31.3%) [29]. In Europe, M. gordonae is most prevalent in Germany, while M. xenopi is prevalent in Hungary [29]. Rapidly growing Mycobacterium (RGM) such as M. fortuitum and M. abscessus are the major species associated with pulmonary disease in Asia, particularly in Taiwan, South Korea, Saudi Arabia, India, Singapore and Malaysia [29,30,31,32,33] (Table 1).
PNTM is a recognized disease in the developed world as the incidence rate of PNTM is higher than TB in countries such as Japan [34], USA [35] and Australia [36]. Indeed, the inverse trend of the incidence rates of NTM and TB was observed in 75% of 16 geographic areas across four continents [37]. The average annual prevalence of PNTM in the USA ranges from 1.4 to 13.9/100,000 and can reach up to 44/100,000 in Hawaii, with an estimated increment of 2.5–8% annually [38]. In the USA, from year 1999 to 2014, even though the total deaths due to TB are higher compared to PNTM, the number of TB deaths has decreased periodically, while the number of PNTM deaths has increased [39].
The rise of PNTM may be partially associated to the advancement of detection methods of mycobacteria, but its annual augmentation is multifactorial, contributed by the pathogen, host, host-pathogen interactions and the still insufficient management of the disease [40]. The current world dynamic landscape, characterized by a growing population, development associated changes with environmental impact, increase of life expectancy and an increasing pool of immunosuppressed individuals associated with chronic communicable and non-communicable diseases and their interactions, among other factors, configure a context where the increase of PNTM is expected in the foreseeable future [38, 41, 42]. To bridge the gaps in prevention, diagnosis and treatment of PNTM, the National Institute of Allergy and Infectious Diseases (NIAID) and the NTM Research Consortium (NTMRC) established by North American clinicians have organized workshops to gather the experts in discussions for better understanding of the pathogen diversity, host-pathogen interactions and the development of efficient control strategies [43, 44].
This review is divided into two main sections: (1) The disease, which includes the most important factors related to the host, the pathogens and their interactions and (2) the control of the disease, comprising (a) prophylaxis—measures to prevent the disease and (b) management of disease—measures focused in the diagnosis and therapy.
The disease (PNTM)
In this section will be discussed the most important factors involved in PNTM: (1) the host, (2) the pathogens and (3) the host-pathogen interactions (Table 2).
Host
The risk of PNTM development increases due to multiple host-related-factors, such as structural lung defects [327,328,329], genetic factors [74] and immunodeficiencies [162, 330], among others; all of them are discussed in more detail in the following subsections (Table 2).
Structural lung defects
PNTM are frequently associated with diverse structural lung defects, mainly with chronic obstructive pulmonary disease (COPD) and bronchiectasis, which are linked to different pathologies (Table 2).
Chronic obstructive pulmonary disease
Chronic obstructive pulmonary disease (COPD) is a disorder characterized by airflow limitation and persistent respiratory symptoms, which have been mainly associated with chronic bronchitis, emphysema and chronic obstructive asthma [45, 46]. The association of PNTM and COPD has been reported. It is considered that COPD predispose to PNTM and the infection with NTM can worsen the evolution of COPD and increase the mortality [47,48,49,50,51, 329]. The use of inhaled corticosteroids in COPD and other chronic pulmonary diseases is considered a risk factor for the development of PNTM [52] (Table 2).
Bronchiectasis
Bronchiectasis is a syndrome characterized by chronic cough and viscous sputum production, bronchial dilatation and thickening of the bronchial wall, which can be idiopathic or associated with diverse aetiologies and comorbidities such as TB infection, cystic fibrosis (CF), allergic bronchopulmonary aspergillosis (ABPA) and impaired mucociliary clearance, among others [53,54,55,56]. Independently of the aetiology, the presence of bronchiectasis predisposes to PNTM [53,54,55,56] (Table 2).
-
(a)
Post-TB infection
Retrospective data on patients admitted for bronchiectasis in a large single centre in China for a period of 17 years showed that pulmonary TB was the major predisposing factor (30%), mainly in patients between 30 and 39 and 60–69 years old [57]. Previous history of TB infection is one of the strongest risk factors associated to PNTM [58]. It is estimated that 10 million new cases of TB occurred in 2017 and about one-quarter of the world population has latent TB [59]. In Korea, both TB and PNTM burden have increased, which leads to the speculation that previous history of TB cause structural lung damage and increase the vulnerability to PNTM [60] (Table 2).
-
(b)
Cystic fibrosis
Cystic fibrosis (CF) is an autosomal recessive disease, caused by mutations of the cystic fibrosis transmembrane conductance regulator (CFTR) gene predominantly found in Caucasian populations, is characterized by CFTR dysfunction [61]. The CF patients have decreased secretion of chloride and bicarbonate across the CFTR channel and increased absorption of sodium through the epithelial sodium channel, resulting in increased mucus viscosity, compromised mucociliary clearance and airway obstruction, among other defects [62], predisposing to PNTM (mainly MAC and M. abscessus) [63, 64].
As CF patients have high prevalence (20%) of NTM infections, the CF Foundation has recommended an annual screening of NTM in these patients [65] and the need to take precautionary measures to limit the transmission of NTM in CF clinics [66] (Table 2).
-
(c)
Allergic bronchopulmonary aspergillosis
Allergic bronchopulmonary aspergillosis (ABPA) is a hypersensitivity reaction to the presence of Aspergillus fumigatus associated with asthma and CF [67,68,69]. Patients with ABPA are at risk of developing PNTM [70, 71] and the infection with MAC and M. kansasii is associated with a higher risk of developing chronic pulmonary aspergillosis (CPA) [72], with poor prognosis related to the use of systemic corticosteroid treatment [72]. The probability to develop bronchiectasis decreases if patients with ABPA receive proper treatment [73] (Table 2).
-
(d)
Impaired mucociliary clearance
The mucociliary clearance impairment, with low ciliary beat frequency, low nitric oxide production and impaired toll-like-receptors function, as seen in CF and primary ciliary dyskinesia patients is a critical determinant in PNTM infection [75]. Defect in several genes has been associated with primary ciliary dyskinesia, e.g. DNAH5, DNAI1, DNAI2, DNAL1, CCDC114, TXNDC3, DNAAF1, DNAAF2, DNAAF3, CCDC103, HEATR2, LRRC6, CCDC39 and CCDC40 [74].
A study by Matsuyama et al. (2018) showed that in respiratory cells infected with NTM, immune signalling leads to downregulation of ciliary genes, upregulation of the inflammatory cytokine IL-32 and cholesterol biosynthesis [76]. A recent study showed that mutations at MST1R gene were associated with decreased airway ciliary function and interferon-γ (IFN-γ) production [77] (Table 2).
Genetic defects
Multiple genetic alterations have been associated with the susceptibility to PTMN. For example, in the case of structural lung defects, specific gene defects [mentioned in “Cystic fibrosis” and “Impaired mucociliary clearance” subsections] have been described [74]. Specific gene defects associated with immunodeficiencies will be described in the next subsection. Defects on CHP2 gene have been associated to PNTM due to MAC [78]. Genetic defects in genes related with immune response, CFTR, cilia, and connective tissue have been found with increased frequency in PNTM patients compared with their unaffected family members and control subjects. Many of the patients had simultaneous defects in various genes, comprising of different categories [79]. Evidence of genetic linkage on chromosome 6q12-q16 with PNTM and the identification of TTK as a candidate gene for PNTM have been found in PNTM patients [80]. Also, haplotypic association with PNTM has been reported [81]. Some studies showed association of different HLA antigens and susceptibility to PNTM. In this regard, HLA-A33, HLA-DR6 and the haplotype A33-B44-DR6 were found with higher frequency in PNTM, and the presence of HLA-A26 has been associated with a bad clinical evolution [82, 83]. The studies related with the genetic defects associated with PNTM support the notion that PNTM is a complex, multifactorial disease with the simultaneous presence of several concomitant genetic alterations [79] (Table 2).
Immunodeficiencies
Immunodeficiencies, primary and secondary, are associated with the increase of susceptibility to infectious diseases [84]. It has been documented the increase of NTM infections in immunosuppressed individuals [85, 86]. In the following subsections, examples of association of primary and secondary immunodeficiencies and PNTM will be discussed (Table 2).
Primary immunodeficiency
Primary defects of the immune system also known as inborn errors of immunity comprise more than 350 hereditary entities associated to single gene mutations, and their classification has been recently updated by the Primary Immunodeficiency Diseases Committee, under the International Union of Immunological Societies [84].
In the category “Combined immunodeficiencies with associated or syndromic features”, patients affected with “Anhidrotic ectodermal dysplasia with immunodeficiency (EDA-ID)”, in its both variants, EDA-ID due to NEMO or EDA-ID due to IKBA, produced by genetic defects in IKBKG or IKBA NFKBIA, respectively, have increased susceptibility to NTM [87,88,89].
Two diseases included in the category “Congenital defects of phagocyte number or function”, X-linked chronic granulomatous disease and GATA2 deficiency (MonoMac syndrome), with defects in CYBB and GATA2 genes, respectively, has been reported with increased susceptibility to NTM [84, 90,91,92,93].
The diseases from the group known as “Mendelian Susceptibility to Mycobacterial Disease (MSMD)”, included in the category “Defects in intrinsic and innate immunity”, mainly associated with gene defects related with the function of the IL-12/IFN-γ pathway (IL12RB1, IL12B, IFNGR1, IFNGR2, STAT1, CYBB, IRF8, TYK2, ISG15, RORC and JAK1), are characterized for increased susceptibility to NTM [84, 88, 94,95,96,97].
Patients affected by “Adult-onset immunodeficiency with susceptibility to mycobacteria”, belong to the category “Phenocopies of inborn errors of immunity”, produce auto-antibodies against IFN-γ and have increased susceptibility to NTM [84, 98,99,100,101,102,103,104,105,106,107,108].
Also, increased susceptibility to PNTM has been associated to C4 complement deficiency [109].
Secondary immunodeficiency
Many different pathological conditions compromise the function of the immune system predisposing to infections, including PNTM. In this subsection will be discussed a group of heterogeneous situations [human immunodeficiency virus (HIV) infection, autoimmune diseases, cancer, Immunosuppressive drugs, surgery, age, malnutrition, vitamin and trace elements deficiencies, addictions and Lady Windermere syndrome] where the presence of malfunction of the immune system increase the susceptibility to PNTM (Table 2).
-
(a)
Human immunodeficiency virus
One of the most demonstrative examples of the impact of immunosuppression on the susceptibility to PNTM is the devastating effect of MAC infection in acquired immunodeficiency syndrome (AIDS) patients [110,111,112,113,114,115].
Severely immunocompromised patients, such as those with HIV infection, with low CD4+ lymphocyte counts are at high risk of PNTM infection even with the introduction of anti-retroviral therapy (ART) [110, 116]. The infection might not be limited to PNTM, but it may progress into disseminated disease (dNTM) caused by MAC [110, 117]. The mycobacterial infections have caused high morbidity and mortality among HIV-positive individuals [110, 117]. In 1996, the introduction of ART in 1996, together with antibiotic prophylaxis, such as azithromycin or clarithromycin, has successfully reduced the development of disseminated MAC among HIV patients [118].
Even at this ART era, the management of dNTM in HIV patients is difficult as in one study, and 79% of the patients had the immune-reconstitution syndrome, which introduce important therapeutic dilemmas [119].
-
(b)
Autoimmune diseases
In patients with systemic autoimmune rheumatic diseases, PNTM could develop and exacerbate [120].
-
(c)
Cancer
Cancer patients are prone to develop PNTM, which could be favoured by the intrinsic immune suppressive effect of cancer and by inherent additional factors such as malnutrition, immunosuppressive treatment, radiation, stress and surgery [121, 122]. Among cancer patients, PNTM infection is most common in lung cancer and MAC is the most common causative agent [123].
-
(d)
Immunosuppressive drugs
Immunosuppressive drugs used to treat cancer [124], autoimmune diseases [125], and following transplantation [126], increase the risk of opportunistic infections [127].
The use of immunosuppressive drugs and TNF-α blockers to treat rheumatoid arthritis increases the risk of NTM infections, mostly by M. avium, followed by RGM, particularly in the lungs [128, 129].
In the case of patients under immunosuppressive treatment, other factors such as the underlying pathology and surgery, among other concomitant factors, potentiate the immune dysfunction.
-
(e)
Surgery
Any major surgical procedure means a great trauma for the patient with many immunosuppressive associated factors during the pre-operative (pre-operative anxiety), intra-operative (tissue damage, disruption of natural protective barriers, blood loss, transfusion, hypothermia, pain, analgesia, anaesthesia and stress), and post-operative (activation of the hypothalamic-pituitary-adrenal axis, endogenous opioids, prostaglandins, cytokines and their agonists) periods [130,131,132,133]. The specific role of surgery per se, in the predisposition of PNTM, is difficult to evaluate, but is a factor that need to be considered in general, particularly in individuals belonging to risk groups of PNTM.
Transplantation
Solid organ transplant (SOT) is a special surgical case, because the immunosuppressive effect of surgery and the lifelong immunosuppressive therapy are combined.
The risk of PNTM is high among SOT recipients, especially in lung transplant patients, which have the highest risk of lung infection compared to other organ transplant recipients [134, 135]. The risk of PNTM in haematopoietic cell transplants has also been reported [136,137,138].
-
(f)
Age
The current world population is estimated to be 7.6 billion [139]. One of the most important characteristics of the current demographic evolution worldwide is the increase of individuals over 60 years of age [140]. The increase in life expectancy is accompanied by multiple challenges for health systems due to the inherent ageing-associated physiological/structural changes and the concomitant increase in associated diseases.
Immunosenescence is one of the processes associated with ageing which is characterized by an increase in predisposition to infection, cancer and autoimmunity and a decrease in response to vaccination [141, 142].
Age is an important risk factor which predisposes to PNTM. Adults older than 60 years old are more prone to PNTM, and their prognosis is commonly unfavourable, which is often complicated by the presence of structural lung defects such as bronchiectasis and COPD [40, 143,144,145,146,147,148,149,150].
This situation may burden health care costs as it is estimated that nearly 1.5 billion people are expected to be over 65 years old by 2050 [151].
-
(g)
Malnutrition
The immune response is affected by the effects related with the undernutrition, low body mass index (BMI), obesity and dietary deficits. Despite the immunological imbalances present in obesity, characterized by inflammation, increase susceptibility to infections and compromise response to vaccination, a clear connection between obesity and PNTM have not been reported, so the discussion will be focussed on undernutrition and dietary deficits [152,153,154,155,156,157].
In children living in low resource areas such as Mozambique, the prevalence of NTM in pulmonary samples of presumptive TB cases can reach 26.3%, and the most common clinical feature observed was malnutrition [158].
Low BMI was associated with PNTM and was a predictor of dissemination [159]. PNTM has also been associated with low visceral fat and low nutrient intake [160]. Low BMI and cholesterol levels were predictors of bad prognosis in another study [161].
-
(h)
Vitamin and trace elements deficiencies
Vitamin and trace elements are of paramount importance for the normal function of the immune system. Primary and secondary malnutrition have a high impact in the availability of micronutrients with increase susceptibility to infectious diseases. Primary malnutrition is frequently associated to poverty and extreme poverty, wars, natural disasters, migrations, malnutrition following beauty patterns, etc. Secondary malnutrition is associated with diseases such as cancer and parasitism, among others [163, 164].
Vitamin D
Deficiency of vitamin D has been observed in adolescent idiopathic scoliosis (AIS) as it affects the regulation of bone mineral density, postural control and fibrosis [165]. Pectus excavatum, a skeletal feature of rickets, is related to hypovitaminosis D, as this vitamin is essential for maintenance of healthy bones [166]. The development of lungs is also affected by the vitamin D level and deficiency of this vitamin may lead to deficits in lung function and volume [167]. In bronchiectasis patients, 50% have vitamin D deficiency and are frequently colonized with bacteria [168]. Generally, the level of vitamin D decreases with increasing age and decreased exposure to sunlight [169]. All the factors previously mentioned have been linked with PNTM; however, studies related with the link between vitamin D and PNTM are relatively scarce. Association of severe vitamin D deficiency and PNTM has been reported [162]. However, in another study, Fujita et al. (2018) showed association of vitamin D with bone mineral density and antimicrobial peptide levels (hCAP18/LL-37), without a direct link between the serum vitamin D level and PNTM [170].
In PNTM patients due to M. malmoense, association of susceptibility with vitamin D receptor gene polymorphisms was found [171].
Vitamins A and E
A study by Oh et al. (2019) on serum vitamin levels in PNTM patients showed that they had significant lower vitamin A and vitamin E levels than healthy individuals, without significant changes in vitamin D levels [172]. It has been reported that in TB patients, vitamin A promotes autophagy to reduce bacterial burden in macrophages [173] and vitamin E-selenium increases antioxidant effects to reduce oxidative stress [174], suggesting that low level of these vitamins may inhibit the ability of host immune system to fight mycobacterial infections.
Trace elements deficiencies
Decreased levels of selenium and zinc were found in PNTM [175].
-
(i)
Addictions
Addictions such as smoking, alcoholism and drug abuse have a high impact in the normal function of the immune system predisposing to infectious diseases [176]. The link between addictions and PNTM has been reported [177,178,179,180,181].
Smoking
It was estimated that over 1/7 of world population (1.1 billion) were smokers in 2016 [182]. Looking at regional smoking prevalence, most Africans and Americans smoke at adolescence [182]. Many people still underestimate the relative risk of smoking [183], and young people perceive smoking as a normal and acceptable behaviour, supported by misconceptions as having stress-relieving effects and social bonding (peer-influence) [184]. Even patients who are diagnosed with COPD do not quit smoking as smoking cessation is difficult due to their lifelong smoking habit [185].
The damages caused by smoking vary significantly depending on the personal smoking practices (frequency of smoking, fraction of smoking, starting age, etc.) and the characteristics of the smoked product (cigarette, tobacco, pipe, chemical concentration, size of the compounds, charged particles formed, etc.) [186].
Cigarette smoke is a mixture of chemical compounds that are free in the gas phase and attached to aerosol particles. It has been estimated that cigarette smoke has 7357 chemical compounds [187], some of them are very toxic and others with great potential to be respiratory irritants, e.g. 1,3-butadiene, acrolein, acetaldehyde, cyanide, arsenic, cresols, N-nitrosamines and polycyclic aromatic hydrocarbons (PAHs), among others [188].
The antecedent of smoking in patients with PNTM has been reported [32, 86, 189, 190]. Smoking impairs the immune defences at the respiratory level by different mechanisms such as inhibition of bacterial killing due to bacterial phagocytosis defects, modulating CFTR dependent lipid-rafts and autophagy impairment, among others [191,192,193,194,195,196]. The predisposing role of smoking in PNTM could be mediated by direct effect on the immune response and indirectly, by its association with pathologies that increase the risk of PNTM such as COPD [196, 197].
Alcohol abuse
According to WHO, alcohol consumption was related to about 3 million deaths and 132.6 million disabilities in 2016 [198]. Even though several countries have adopted alcohol control policies as suggested by WHO since 1999, including restriction of alcohol marketing, the weak implementation of prevention programmes has not achieved significant impact on human health [199].
Alcohol is another causative agent for airway inflammation and injury as observed in COPD, and prolonged alcohol consumption leads to increased risk of mortality in these patients [200]. The multiple deleterious effects of alcohol in the function of the different arms of the immune system have been documented [179, 180, 201], including disruption of the immune defence mechanisms at the airways [202].
Alcohol abuse has been associated with the risk of PNTM in HIV infected and non-infected individuals [190, 193].
Drug abuse
Drug abuse induces important deficits in the immune system, which impacts the susceptibility to infectious diseases [176, 181].
A study on HIV patients with NTM diseases showed that their risk of infections was associated with both alcohol and drug abuse. Depending on the drugs used, the results showed that more SGM were isolated from intravenous and inhalation cocaine users, while RGM were isolated from inhalation crack users [203].
-
(j)
Lady Windermere syndrome
Generally, tall, lean, old Caucasian ladies with MAC lung disease, with an isolated lingular or middle lobe bronchiectasis pattern, are known as LWS. The syndrome is associated with a voluntary suppression of the normal cough reflex due to politeness, which impairs the clearness of airway secretions [330]. Patients affected with this syndrome also tend to have scoliosis, pectus excavatum or mitral valve prolapse even without any apparent immune defects [204, 205].
A study by Chan et al. (2010), showed that NTM infections are more frequent in females even without any overt immune defects [206]. However, despite of the first perception that this syndrome was not related with immune compromise, defects in MST1R gene, associated with compromised airway ciliary function and reduction on IFN-γ production in response to NTM, have been found in patients with LWS as well as alterations in immune related genes [77].
“Lord” Windermere syndrome
Although the alterations associated with LWS have been associated with female gender, similar characteristics to the LWS have been described in men [207, 208].
Pathogens (NTM)
There are more than 180 NTM species that are classified into 4 groups based on growth rate and pigment production, known as Runyon classification. The first 3 groups are slow growers (SGM) which require more than 14 days to growth: (I) photochromogens, which develop pigments in or after being exposed to light, (II) scotochromogens, which become pigmented in light/dark conditions and (III) non-photochromogens, which do not form pigment. The group (IV) RGM require only 2 to 5 days to growth [331]. Some phenotypic [17, 18, 27] and genomic [19] characteristics of the most important NTM associated to PNTM are listed in Table 1.
Recently, based on the results of phylogenomic analysis and identified molecular signatures [Conserved Signature Indels (CSIs) and Conserved Signature Proteins (CSPs)], Gupta et al. (2018) described the division of mycobacterial species in one emended genus: “Mycobacterium”; and four novel genera: Mycolicibacterium, Mycolicibacter, Mycolicibacillus and Mycobacteroides, which have five clades: “Tuberculosis-Simiae”, “Fortuitum-Vaccae”, “Terrae”, “Triviale”, “Abscessus-Chelonae”, respectively. NTM belong to these five clades [17] (Table 1).
“Tuberculosis-Simiae” clade
This is the only clade that includes NTM and MTBC species. Some NTM, from this clade, has been reported as pathogenic Mycobacterium species including MAC, M. gordonae, M. kansasii, M. xenopi, M. simiae, M. szulgai and M. interjectum causing PNTM [17].
“Fortuitum-Vaccae” clade
This clade includes M. fortuitum and has been described as causing PNTM [17].
“Terrae” and “Triviale” clades
Some members of “Terrae” and “Triviale” clades have been described as opportunistic pathogens [17].
“Abscessus-Chelonae” clade
This clade has six members and has been described as pathogenic to humans. PNTM has been described caused by M. abscessus and M. chelonae [17].
Development in molecular methods, especially DNA sequencing and polymerase chain reaction (PCR) tests, has enabled NTM species-species differentiation and strain typing [332]. Most NTM are considered as non-pathogenic microorganisms which are harmless and unlikely to cause disease but become opportunistic pathogens especially in immunocompromised hosts [333]. On the other hand, those within the Mycobacterium tuberculosis complex (MTBC) are regarded as pathogenic microorganisms which cause a specific disease, TB [334].
Virulence factors
Genomic profiling of 41 NTM species showed that MTBC and NTM do share similar virulence factors, e.g. (1) cell surface hypervariable PE/PPE proteins that are important in the evasion of the host immune response, except for PE5, which is only found in NTM; (2) ESX or type VII secretion system (T7SS) for PE/PPE proteins transportation, where ESX-3 is conserved in both groups and ESX-1 which was thought to be specific for Mtb, is also expressed in M. gordonae, M. riyadhense and M. szulgai; (3) Mce proteins that are involved in host-cell invasion, namely, Mce1, Mce2, Mce3 and Mce4 which are present in Mtb, are also found in many NTM, with Mce5, Mce6, Mce7, Mec8 and Mce9 only present in NTM; (4) Sec export systems, SecA1 and SecA2, that are important to export Mtb lipoproteins are conserved in all species; and (5) Tat export system for Mtb virulence is also conserved in all species [209]. The genes involved in biosynthesis of mycolic acids in NTM are similar to those for Mtb, although genes for dimycocerate esters (DIM) biosynthesis are only found in SGM, particularly in the pathogenic mycobacteria, i.e. MTBC, M. leprae, M. kansasii, M. marinum, M. ulcerans and M. haemophilum [209].
In terms of immunogenic proteins, previously detected in Mtb, with some role in virulence, a study on 4 NTM species showed that CFP-10 was detected in M. malmesburii sp. nov.; GroES was detected in M. komanii sp. nov., M. nonchromogenicum and M. fortuitum ATCC 6841; and DnaK and GroEL were detected in all the 4 NTM [210]. Also, CFP-10 and ESAT-6 of Mtb are detected among M. kansasii, M. szulgai, M. marinum and M. riyadhense [211]. The presence of shared antigens between Mtb and NTM has given new insights to consider NTM as potentially pathogenic and could be associated with false positive results with the current interferon gamma release assays (IGRAs) in use [212].
Cell wall components of mycobacteria, including NTM, are important elements related to virulence, immunogenicity, immunomodulation and drug sensitivity [213,214,215,216] which have a high impact on the development of diagnostics, therapies and vaccines.
Important cell wall components of NTM are glycopeptidolipids (GPLs) and lipooligosaccharides (LOSs) [213,214,215]. GPLs, which are absent in Mtb [213] can be species-specific (ssGPLs) or non-specific (nsGPLs) [214]. The addition of an oligosaccharide to nsGPLs confers further immunogenicity producing the ssGPLs, which determine 31 serotypes among MAC members [214]. GPLs are associated with immunopathological responses during infection [214]. The serotypes of MAC are associated with pathological characteristics as in the case of serotype 4 which is linked to disseminated infection in AIDS patients [214, 217]. LOSs are also present in M. canettii and some MTBC, but absent in Mtb, such as H37Rv [215, 218]. LOSs are strongly immunogenic and associated with immunomodulation and virulence [218,219,220,221,222].
Biofilm
It is well documented that NTM such as M. avium [24], M. marinum, M. kansasii [26], M. smegmatis [223], M. abscessus [20], M. fortuitum and M. chelonae [22] can form biofilms which are important for resisting physicochemical stress, antimicrobials and immune defence mechanisms, favouring their persistence [224].
Besides mycolic acids, the mycobacterial cell wall has extractable lipids such as GPLs [20, 22, 24, 223], LOSs [26], and trehalose dimycolate (TDM) [20] together with extracellular DNA (eDNA) [21, 23, 25] which contribute to the biofilm development. Biofilm development is also partly influenced by the presence of nutrients [225,226,227] (Table 1).
As mycobacteria lack of pili and fimbriae, GPLs play an essential role in attachment on surfaces for biofilm formation and sliding motility for biofilm spreading [228]. Also, the presence or absence of GPLs determine the morphology and virulence of M. avium and M. abscessus, i.e. less virulent, non-cord-forming smooth variants has increased GPLs production, and more virulent, cord-forming rough variants lack GPLs [20, 24, 229].
Transmission
Transmission of NTM is similar to TB via aerosolization, but the source of transmission is from the environment, not human or animal, suggesting that PNTM is not contagious [332]. The propensity to PNTM increase due to exposure to environmental factors related to water and soil [230]. Although transmission of NTM from human-to-human is not common, in 2010, a first case of M. kansasii isolated from a couple in East London, an area with high TB burden, was reported [231]. The isolates were genetically identical, and the authors were unable to prove a shared domestic exposure [231]. Whole genome sequencing (WGS) has revealed the possibilities of human-to-human transmission, but most probably in an indirect manner via fomite contamination or inhalation of NTM in airborne water droplets [232]. Contamination of the environment after natural disasters and human mobility might be associated with PNTM transmission [233,234,235,236] (Table 2).
-
(a)
Water
NTM are more likely to survive at surface waters than in ground waters [230]. Thus, the risk of PNTM could be potentially associated with the percentage of surface water, temperature, humidity, rain precipitation and potential of evaporation and transpiration, whereby aerolization, favoured by the mycobacterial hydrophobicity and inhalation of water droplets contribute to PNTM [230]. The hydrophobicity promotes bacterial adherence at surfaces and pellicle biofilm formation at the liquid-air interface [224].
A study by Wei et al. 2016 showed that developed and developing countries contributed 53–61% and 39–47%, respectively, to climate changes based on the emissions of carbon dioxide, methane and nitrous oxide [237]. Human activities have changed the atmosphere composition causing accumulation of greenhouse gases leading to global warming [237]. The increase of global air temperature indirectly leads to the increase of evaporation and transpiration that convert surface water, from natural (e.g. sea and river) or artificial (e.g. swimming pools and ponds) sources into atmospheric water vapour [230].
MAC has been reported as an important causative agent for hypersensitivity pneumonitis at hot tub or spa and associated with “hot tub lung”, a diffuse granulomatous lung disease [238].
-
(b)
Soil and dust particles
Activities (hobbies or job-related) associated with long-term contact with soil such as farming, and lawn and landscaping services are more likely to be exposed to NTM infection. Recovery of MAC strains is highly prevalent in the residential soil samples in Japan and the bacteria transmission is associated to duration of soil exposure for more than 2 h per week [245]. Soil properties play an important role in promoting growth and persistence of NTM in nature. The soil of countries with high-risk of infection have higher copper and sodium levels, but lower manganese levels compared to low-risk countries [230].
Trapped, dusty environments promote the aerolization of NTM and cause the development of silicosis and chronic bronchitis in South African gold miners, which may predispose to PNTM, particularly caused by M. kansasii and MAC [58]. The exposure of indoor home dust could be a source of PNTM, as M. avium ssp. hominissuis is highly found in dust from vacuum cleaner bags in Germany [246].
The presence of NTM such as M. intracellulare, M. abscessus, M. szulgai, M. fortuitum, M. avium, M. kansasii, M. simiae, M. gordonae, M. terrae complex, M. chelonae and M. malmoense has been reported in sand [30]. Dust and sand from deserts, due to storms, can travel long distances even between continents and represent a vehicle for the dispersion of microorganisms, including NTM [247,248,249]. Also, mycobacterial DNA have been found in cosmic dust [250].
-
(c)
Other sources
Dialyses centres, heater-cooler units, kitchen sinks, household refrigerator taps, home ice machines, tap water filters, retail sold fish, frozen fish, piped surfaces, soil fertilized with chicken droppings, dust from vacuum cleaners, air conditioner and cigarettes have been reported as sources for NTM colonization [335].
Some NTM (MAC) can evade the degradation within free-living Acanthamoeba polyphaga, A. castellanii and their exocysts. Nearly 88% of amoeba (Acanthamoeba, Vermamoeba, Echinamoeba, and Protacanthamoeba) from drinking water contains M. chelonae [335].
NTM can be transmitted via oral route due to the gastroesophageal reflux [335].
Another important source of transmission are the disasters. Stress, disruption of health care systems, bad hygienic-sanitary conditions, food deprivation and large migratory waves, among others, are the scenarios caused by disasters, whether natural (earthquakes, tsunamis, etc.) or man-made (wars, extreme poverty, ethnic/religious/political conflicts, etc.), which create favourable conditions for the transmission of microorganisms, turning the host susceptible, making them vulnerable to the development of multiple diseases.
War is considered a public health problem [245]. Armed conflicts are classified as one of the biggest disasters that a population can suffer, affecting health directly by the effects of weapons and indirectly breaking the structures of health systems, their supply chains and interrupting health programs as vaccination and paediatric care, among others. These factors together with the destruction of water supply networks, the houses and population overcrowding, among other factors, cause the emergence of epidemics, re-emergence of common vaccine-preventable-infectious diseases in children and diseases associated with severe malnutrition. When this scenario is impregnated with violence and insecurity, it pushes populations to move, with all the terrible consequences that are added. All these factors create optimal conditions for the proliferation of infectious diseases [251, 252].
Armed conflicts are recognized as important factors for the spread of Mtb [252]. The dissemination of NTM and the increase of the incidence of PNTM in war affected populations cannot be excluded, as the presence of the necessary conditions for this to happens is present, but the evaluation of this in the difficult conditions created by war is difficult, in fact, the possibility exists that patients in these populations (initially diagnosed as TB/MDR-TB) could be affected by PTNM.
During some natural disasters, NTM transmission may increase due to several factors. The mixture of the ocean and the fresh waters, together with the mixture of the soil with the water, could produce aerosols that contain NTM, favouring the inhalation. In addition, the free-living amoebas move from their natural niches, providing a “protected” environment for intracellular multiplication of NTM and virulence, which can potentially cause PNTM [233].
Human mobility is another source of transmission. The development of widespread transportation and communication has increased population mobility, which contribute to disease outbreaks and the spread of infections [234]. Nishiuchi et al. (2017) hypothesized that globalization of human activities, mobility and trade may increase the global transmission of MAC via fomites [235]. It is suggested that person-to-person spread is unlikely and the ability of NTM to resist desiccation and detergents might promote their survival during transportation [236] (Table 2).
Host-NTM interaction
The fragile boundary between health and disease could be broken by many associated factors, mainly related with the host and the pathogen, but the resultant of the interactions between them plays the most important role. PNTM is the undesirable resultant of these interactions. The interaction is dynamic and the relative “strength” of “weakness” of each player will depend on continuously evolving factors in each side, given at any moment a “balance”, which will determine the presence or not of the disease. The challenge of disease control lies in the measures that should be implemented to block the NTM transmission and to shift the balance towards the host resistance aimed to prevent the disease, or the recovery once it appears.
Control of PNTM
In this section, prophylactic measures and the management of the disease will be discussed.
Prophylaxis
The prevention of infectious diseases is one of the most important objectives of the health systems, which are mainly achieved through (1) pathogen-related prophylactic measures (blocking the transmission), and (2) host-related prophylactic measures (vaccines) (Table 2).
-
(a)
Pathogen-related prophylactic measures: blocking the transmission
In the case of PNTM, some measures could be implemented to decrease the transmission mainly through water and soil.
Despite better public health measures, i.e. improvement in water supply via centralized water supply systems, which help to minimize most pathogenic water-borne bacteria, unfortunately, it has promoted NTM colonization [239]. In centralized water supply systems, water travels long distances, which eventually leads to degradation of the disinfectants, i.e. decrease in chlorine concentrations due to heat inactivation [239]. Also, a study showed that despite addition of ozonation and filtration treatments, NTM can still persist in water systems [240]. NTM can be recovered in water with free and total chlorine levels of 2.5 and 2.8 mg/l, respectively, suggesting that chlorination is unable to efficiently kill mycobacteria [241]. One of the main reasons that mycobacteria can survive in this harsh environment is their ability to form a protective shield, biofilm [229]. In potable water, slow growing MAC form a highly culturable biofilm compared to RGM, suggesting that the former has better adaptation to growth in low-nutrient environments [229]. Also, M. avium is able to survive and grow in free-living amoeba, particularly Acanthamoeba lenticulata as parasites or endosymbionts, and the encystment protect M. avium from the disinfection processes [336]. Water treatment has always been focused on eliminating water-borne gastrointestinal bacteria such as Campylobacter, Salmonella, Shigella and Escherichia coli but not in the elimination of mycobacteria [337]. Considering the ability of mycobacteria to survive in these harsh environments even after treatment, a more appropriate water treatment plan is needed as the consequences of NTM infection should not be overlooked. A study by Inkinen et al. (2016) suggested that cooper pipelines might be effective in preventing NTM colonization compared to polyethylene pipelines [338].
Many studies have showed that inhalation of aerosols generated from shower-heads, therapy pools, hot-tubs and swimming pools may promote PNTM [242,243,244]. A study reported by Feazel et al. (2009) showed that among the 16 microorganism genera present in shower-heads, mycobacterial biofilms had the highest prevalence, mainly composed by M. gordonae and M. avium, which were more than 100-fold than in their background water composition [339].
Preventive measures such as cleaning shower-heads by soaking in vinegar or water, replacing shower-heads periodically, removing shower-heads completely, taking baths instead of showers, and avoiding steam rooms and hot tubs have been recommended to the public [340]. In general, the recommended preventive advices are not incorporated in our daily routine and most people still prefer showers compared to baths, partly due to convenience. Also, in line with water conservation drives and to achieve a more eco-friendly environment, a quick shower is definitely better than a bath as an average shower of 10 min uses 10 to 25 gal of water, while a bath needs at least 70 gal [341].
Inhalation of water during swimming also exposes the respiratory system to NTM. A study by Prevots et al. (2014) showed that swimming in indoor pools is associated with PNTM, which may be attributed to the formation of bacterial biofilms in closed environments which facilitates higher exposure to aerolized biofilms [41]. The lack of ventilation in closed areas facilitate inhalation of water aerosols. Also, the use of pressure washers during cleaning has indirectly increased water aerosolization and this exposure increases the risk of occupational hazard [253]. Thus, to prevent this environmental colonization, it has been suggested that such closed environments be kept dry and have proper ventilation [254].
It is recommended that workers with occupational risk should wear personal protective equipment such as masks and gloves while doing soil-related works. The World Health Organization (WHO) recommended the use of respiratory protective equipment in airborne contaminated working environments, but frequently, the workers remove the masks after wearing it for long periods of time in hot and cramped environments [255]. The use of respiratory protective equipment should be recommended also for domestic or recreational activities associated to soil and dust exposure.
-
(b)
Host-related prophylactic measures: “arming” the host by vaccination
Prophylaxis by vaccination is one of the most potent health measures to control infectious diseases, which allow the prevention of the disease, even when the measures for pathogen control are not fully effective.
Currently, there is no vaccine recommended for PNTM. Bacillus Calmette-Guerin (BCG), the only vaccine currently available for TB, has shown protective effects in extrapulmonary NTM infections, but there is no report for PNTM [43]. A review by Zimmermann et al. (2018) based on ten studies involving 12 million participants showed that BCG vaccination is protective against NTM lymphadenitis caused by MAC and Buruli ulcer caused by M. ulcerans [342]. However, the cross-reactivity between these Mycobacterium may lead to low efficacy of BCG vaccine, especially in high NTM exposure areas [343].
There are several mouse models which demonstrated the protective effects of subunit vaccines against PNTM infections. A study by Fattorini et al. (2002) using intranasal immunization of mice with M. avium GroES recombinant protein, co-administered with CpG oligodeoxynucleotides, was protective against intranasal M. avium challenge [256].
A plasmid DNA encoding M. abscessus phospholipase C formulated with copolymer 704, and administered intramuscularly in mice protected against aerosolised challenge with M. abscessus [257]. Immunization of mice with genetic constructions containing the M. abscessus MgtC gene induced protection against challenge with M. abscessus and the production of specific antibodies in mice [258].
A dewaxed whole-cell vaccine of M. ulcerans devoid of its mycolactones (cytotoxic macrolide exotoxin) and waxy cell walls, protected against M. ulcerans infection and prevented Buruli ulcer development in mice [259]. Priming with recombinant BCG expressing the Ag85A antigen of M. ulcerans, followed by a booster with recombinant M. smegmatis expressing Ag85A of M. ulcerans induced murine CD4+ T cell responses which reduced tissue damage and bacterial loads [260]. However, recombinant surface proteins MUL_2232 and MUL_3720 of M. ulcerans induced strong TH1 immune response but without protective effect [261].
An NTM vaccine commercially used in veterinary, Gudair®, containing the inactivated M. avium paratuberculosis strain 316F, is effective in decreasing the prevalence of the infection and reducing the bacteria faecal shedding in sheep [262].
The antigenic similarity and immune cross-reactivity between mycobacteria have been exploited in the development of vaccines and immunotherapy of TB [17,18,19, 27, 203,204,205,206,207,208,209,210,211, 331,332,333,334]. Proteoliposomes [263] and lipid-based preparations of the cell envelope, obtained from M. smegmatis [264], were protective against Mtb intratracheal challenge in mice, and induced cross-reactive humoral immune responses against Mtb antigens [87, 265, 266].
DAR-901, a heat-inactivated whole-cell M. obuense SRL172 strain, has been recommended as a vaccine booster for TB in adults primed with BCG as it is safe and is able to induce both cellular and humoral immune responses [267]. A murine model further demonstrated that DAR-901 conferred protection against Mtb [268].
M. habana TMC5135, a species synonymous to M. simiae serotype I, isolated from Cuba, showed protective effects against Mtb using animal models as live vaccine [269].
M. manresensis, a new species under the M. fortuitum complex, which is commonly found in drinking water, was heat-killed and used as oral vaccine to stop the progression of active TB and as adjuvant for TB treatment in a mouse model [270]. It has been developed into a galenic preparation food supplement known as Nyaditum resae® and used in clinical trials, which seemed to be effective in reducing the risk of developing active TB [271, 272].
Immunotherapy with killed M. vaccae has been used in the treatment of patients with TB, multidrug resistant TB, HIV-TB and leprosy with significant effects [273, 274].
M. indicus pranii as a booster to BCG via the aerosol route showed protective effect in a mice model of TB, inducing pro-inflammatory cytokines like IFN-γ, IL-12 and IL-17, and increased frequency of multifunctional T-cells [275]. In vitro studies showed that M. indicus pranii modulates pro-inflammatory responses via the TLR-4 pathway, activate the innate immunity [276] and induced autophagy in TB infected macrophages [277]. Clinical trials as adjunct therapy showed that it is safe with no significant side effects and able to clear the pathogens in TB relapse patients [278].
Overall, these encouraging results on NTM based vaccine candidates for the prevention of experimental NTM infections, its use as veterinary vaccines and their evaluation for the prevention and therapy of TB support the possibility to develop vaccines for the prevention of PNTM as well as immunotherapeutic tools to help in the management of these infections.
Management of the disease
When prophylactic measures fail, the disease appears in susceptible individuals; beyond this point, its control is only possible with a fast and appropriate diagnosis and therapy.
-
(a)
Diagnosis
The American Thoracic Society (ATS) and the Infectious Diseases Society of America (IDSA) recommend that in case of suspicion of PNTM, chest radiographic studies, three or more sputum microbiological analysis and clinical exclusion of other disorders are needed to confirm a diagnosis of PNTM should be indicated [332]. Such criteria are needed because NTM exists naturally in the environment, and isolation of NTM from non-sterile respiratory specimens does not confirm that the organism is the causative agent for lung disease [344]. High-TB burden countries are normally those of the low to middle income countries (LMIC), where resources for microbiological culture using the traditional diagnostic TB gold standard either on solid Löwenstein–Jensen medium or liquid culture [Mycobacteria Growth Indicator Tube (MGIT)], are limited. Thus, sputum smear microscopy is the most important test used for TB diagnosis [345]. With the current rise of PNTM infections, the non-specific microscopic testing, which detects all acid-fast bacilli, cannot differentiate between PNTM or TB infection. The application of chest X-ray screening to predict the presence of TB or PNTM remains subjective. As recommended by ATS/IDSA, PNTM can be diagnosed based on nodular or cavitary opacities on chest radiography [332]. Gommans et al. (2015) studied 83 PNTM patients showing that cavities were observed most frequently, while consolidations were a predictor for risk of mortality [346]. A radiographic study on 108 TB and 25 PNTM patients showed that the presence of honeycomb appearance (characteristic appearance of variably sized cysts in a background of densely scarred lung tissue) is significant in PNTM patients; consolidation, miliary nodules, cavities, atelectasis, fibrothorax and mediastinal widening are more common in TB patients; and pleural effusion, pleural lesions, and reticulonodular infiltration are observed in both TB and PNTM patients [347]. Therefore, the confirmation of PNTM in LMIC remains a diagnostic challenge for microbiologists and clinicians.
Many studies have been focused on the development of diagnostics for active, latent and drug-resistant TB [279]. Genotypic characterization tests such as Xpert MTB/RIF, loop-mediated amplification tests, and line-probe assays (LPA) which are more sensitive, easier and faster compared to phenotypic microbiological characterization for Mtb have been endorsed by WHO [279]. According to ATS/IDSA, besides Runyon classification, genotypic identification of NTM can be done using acridium ester-labelled DNA probes targeting 16S rRNA, DNA sequencing of 16S rDNA, and PCR restriction endonuclease activity targeting hsp65 [332]. Also, WHO has recommended one nucleic acid amplification test (NAAT), NTM + MDR-TB Detection Kit 2 (Nipro Co., Japan) using LPA [280]. Other NAATs commercially available to detect NTM and drug resistance are included in Table 2 [281,282,283,284,285,286,287,288,289,290]. Detecting drug resistance in the early phase of diagnosis can help determine better treatment options for patients with the possibility to eradicate the infection in shorter time (Table 3).
The current available diagnostic tests using single gene-target sequencing are unable to differentiate all the NTM species, and higher subspecies discrimination require multiple genes sequencing [344]. Species characterization is important because different species have different pathogenicity and drug susceptibility patterns [348]. The type of NTM isolated from the lungs determines the risk or likelihood of lung disease. Hence, the probability of lung disease is high if M. kansasii is detected; intermediate if MAC, M. abscessus complex, M. chelonae, M. malmoense, M. szulgai and M. xenopi are detected; low if M. simiae, M. fortuitum, and M. terrae are detected and very low if M. gordonae is detected [236, 348]. A retrospective study on PNTM patients’ survival rate and type of mycobacterium infection showed that the median survival rate due to M. xenopi infection is the shortest (7 months), followed by M. malmoense (10 months), M. kansasii (39 months), MAC (41 months) and RGM (78 months), suggesting that M. xenopi is highly virulent [346].
Advancement in high-throughput technologies has revolutionised the detection of infection causing organisms. Comparison of detection of NTM using PCR and matrix-assisted laser desorption ionization-time of flight mass spectrometry (MALDI-TOF MS) showed that the later have higher accuracy (97.4%) with faster and cost-effective performance [349]. The development of WGS has enabled identification of diverse bacteria up to the strain level [350]. For example, M. abscessus is the most drug resistant species with a wide variety of drug resistance profiles, which render treatment challenging [350]. The application of WGS helps to detect these variants for proper management of the patients [350] and it is also able to detect the potential mode of transmission either from individuals or environment [232].
-
(b)
Treatment
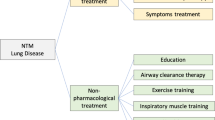
PNTM is a chronic disease which requires a long treatment (18 to 24 months) with multiple antibiotics, which is associated with serious side effects and frequent resumption of treatment due to relapses [44]. Based on the Patient-Centered Research Priorities suggested by NTMRC, experts should be consulted for prescription of antibiotics and drugs to implement the more suitable treatment [44]. The effectiveness of the treatment is assessed based on the patient’s quality of life [66]. Other adjunctive treatments such as airway clearance, exercise and probiotics which seems to ameliorate patients’ outcomes require more clinical testing [44].
Differentiation of PNTM and TB is important because the first-line antibiotics used to treat TB are less effective against NTM [351]. A study of 12 SGM and 12 RGM with 15 antimicrobial drugs showed that 24, 16 and 8 strains were resistant to isoniazid (INH), rifampicin (RIF) and streptomycin (STR), respectively [351]. The authors concluded that STR, amikacin (AMK), the fluoroquinolones (FQs), and the tetracyclines (TET) are the most effective antimicrobial agents against the 24 strains [351]. A study of 95 NTM strains with ten drugs showed that ethambutol (EMB) is the most useful agent against NTM, but its resistance rate among the tested strains was around 42% [352]. The emergence of multidrug resistant NTM (which can be resistant to five or more antimicrobial drugs) has also complicated the treatment [353]. Thus, according to the British Thoracic Society guidelines, different drugs or combination of drugs should be used to treat different species [236]. For example, treatment of MAC lung disease with RIF, EMB and macrolides [clarithromycin (CLR) or azithromycin (AZM)]; treatment of M. kansasii lung disease with RIF, EMB and macrolides; treatment of M. xenopi lung disease with RIF, EMB, macrolides and FQs/INH; and treatment M. abscessus lung disease with AMK, tigecycline (TGC), imipenem (IPM) and macrolide is recommended [236].
However, one of the most complex situations in the treatment of NTM is represented by the infections with M. abscessus complex, which comprise M. abscessus ssp. abscessus, M. abscessus ssp. massiliense and M. abscessus ssp. bolleti [354] due to the high degree of antibiotic resistance and the poor outcome of the treatment [355]. Special therapeutic problems are associated with the resistance to macrolides linked to the presence of the macrolide inducible erm(41) gene, which is active in M. abscessus ssp. abscessus and M. abscessusssp.bolletii and inactive in M. abscessus ssp. massiliense, which have a high impact in the response to the treatment [356]. Using Mtb treatment as reference, it is considered that in patients with M. abscessus ssp. abscessus infections, the results of the treatments are worse than in the case of multidrug resistant TB (MDR-TB), and equivalent to the outcome of the treatment of extensively drug resistant TB (XDR-TB); in the case of M. abscessus ssp. massiliense, the results are close to that obtained in MDR-TB [355]. M. abscessus ssp. bolletii is considered to have similar patterns of resistance to macrolides than M. abscessus ssp. abscessus [357].
Considering the different profile of antibiotic resistance and evolution of the different M. abscessus complex members, it is of great importance the strain identification to implement the more suitable therapeutic strategy [358]. “The antibiotic nightmare” [357] represented by M. abscessus complex infections, which has been also implicated in transmission associated to surgical procedures [359, 360], has stimulated the search for new therapeutic alternatives, such as the use of phage therapy, which opens a window of hope not only for M. abscessus complex but for treatment of mycobacterial diseases in general [361].
Besides the existing drugs, several approaches have been conducted using novel drugs/compounds, modified drugs, medicinal plant extracts, animal venom-derived antimicrobial products and synergistic and combination effects with other antimicrobials to combat the antibiotic resistant NTM with promising results [362]. However, the translation of these studies from in vitro to in vivo remains challenging. The drug delivery mechanism via the inhalation route has been studied to ensure that a high concentration of antibiotics can be delivered directly to the lung without cytotoxic effects to the host [362]. Screening libraries have been used to identify potential antimicrobial compounds for NTM [362].
The ability of NTM to form biofilms has enabled them to survive under environmental stress and confers protection against antibiotics causing bacterial colonization and onset of disease and invasion [228, 333]. A study by Ortíz-Pérez et al. (2011) on biofilm-producing RGM treated with antibiotics showed that biofilms are resistant to AMK, CLR and ciprofloxacin (CIP) [363]. Among these three antibiotics, CIP is the most active drug affecting the thickness of the biofilms and its combination with anti-biofilm agents such as N-acetylcysteine (NAC) and Tween 80 have resulted in higher bacterial death [364].
Besides the identification of the bacteria itself to confirm the presence of the disease, detection of the risk factors would be beneficial to reduce exacerbation of the disease [365]. Adults presenting with NTM infections should be initially screened for HIV, systemic illness and medication history which may lead to immunosuppression. If no risk factors are identified, individuals with pulmonary disease should undergo chest imaging, pulmonary function test and vitamin D level test to detect any structural lung abnormalities. If the disease is not due to pulmonary defects, then subsequent tests are needed to detect primary immunodeficiency and CF. If no risks are identified, tests to detect autoimmunity or pro-inflammatory cytokines should be taken into account to explore the presence of autoimmune diseases [365].
Patients who are diagnosed with PNTM need to be studied for the presence of underlying TB infection because the prescription of treatment only for PNTM could have risk to develop MDR-TB [366]. In addition, for elderly patients infected with PNTM who need to be treated with macrolides, rifamycin and FQs, comorbidities and associated concomitant therapies should be determined since these drugs may interact with those that interact with P-450 and disturb the metabolism of drugs [367].
Hong et al. (2015) suggested that serum carbohydrate antigen (CA) 19-9 can be a useful marker to monitor the therapeutic responses in PNTM as it is higher in PNTM than in TB, and its concentration is reduced after successful PNTM treatment but not in TB [368].
One aspect that should be considered in the treatment of PNTM is the use of immunotherapy, which could represent an important co-adjuvant of the drug treatment; in this regard, the use of M. vaccae together with antibiotic treatment was not associated with improved response in pulmonary MAC infections [291]; however, the indirect and direct evidences of the potential of vaccination in PTNM support further evaluation of this aspect. Encouraging results have been reported with the use of IFN-γ and IFN-α in the immunotherapy of PNTM [292, 293].
The growing consensus about the protective role of antibodies in mycobacterial infections [294,295,296,297,298] suggests the potential use of antibody formulations for the treatment of PNTM. Previous studies have demonstrated the role of humoral immune responses in the defence against mycobacteria in humans [299,300,301].
Monoclonal antibodies against Mtb antigens have demonstrated protective effect upon Mtb challenge after the administration by mucosal or parenteral routes to mice [302,303,304,305].
The combination of the administration of mucosal IgA monoclonal antibodies with IFN-γ demonstrated therapeutic effect in mice challenged with Mtb [302]. Commercial human gamma-globulin formulations have demonstrated prophylactic and therapeutic effect in challenge models with Mtb and BCG [306, 307].
Human IgA formulations obtained from colostrum administered by the mucosal route produced protective effects in a model of progressive TB in mice [306].
Considering these antecedents, the use of monoclonal antibodies specific of NTM antigens or human IgG or IgA formulations by the systemic or mucosal route in PNTM patients could be a new approach in combination with antibiotic treatment, or a valid alternative in cases of treatment failure [308,309,310,311,312,313,314].
-
(c)
Drug discovery
Efforts to develop new drugs for the treatment of NTM infections are continuously being made. The search for new NTM drugs is focussed on reducing the long treatment time which is accompanied by the toxicity of the drugs.
Patients with NTM infections urgently need more safe and effective treatments, preferably orally administered and capable of covering a broad spectrum of microorganisms. Different strategies have been proposed to address the drug discovery lines, i.e. de novo drug discovery and repurposing/repositioning of existing antibiotics. However, there are multiple challenges that affect the discovery and development of new antibiotics for NTM [315,316,317,318,319,320,321,322,323,324,325,326] (Table 4).
These factors have been recently summarized from different points of view. Wu et al. (2018) described them from a bacteriology and disease pathology standpoint [315], while Falkinham (2018) grouped the factors in accordance with the innate genetic defects and physiologic traits of NTM as well as the difficulties in measuring anti-NTM antibiotic activity in the laboratory [316]. In general, these challenges facilitate the survival of mycobacteria under different environments.
Conclusion
PNTM infection is multifactorial, related to the host, the microorganisms involved, the environment, the socio-economic aspects and human behaviour. Emphasis should be put on recommendations related to human activities, aimed to reduce the risk of exposure to NTM, which are being neglected. The priority to the development of new diagnostics, treatments and vaccines for TB should be expanded to PNTM, as sensitive and specific diagnostic tests, vaccines and immunotherapies for these infections are still lacking.
References
Noviello S, Huang DB (2019) The basics and the advancements in diagnosis of bacterial lower respiratory tract infections. Diagnostics (Basel) 9. doi:https://doi.org/10.3390/diagnostics9020037
Niederman MS, Zumla A (2019) Editorial: toward improving the diagnosis, treatment and prevention of community acquired and nosocomial respiratory tract infections. Curr Opin Pulm Med 25:217–219. https://doi.org/10.1097/MCP.0000000000000577
Dawson DJ (2000) Mycobacterial terminology. J Clin Microbiol 38:3913
Mathewos B, Kebede N, Kassa T et al (2015) Characterization of mycobacterium isolates from pulmomary tuberculosis suspected cases visiting Tuberculosis Reference Laboratory at Ethiopian Health and Nutrition Research Institute, Addis Ababa Ethiopia: a cross sectional study. Asian Pac J Trop Med 8:35–40. https://doi.org/10.1016/S1995-7645(14)60184-X
Cook JL (2010) Nontuberculous mycobacteria: opportunistic environmental pathogens for predisposed hosts. Br Med Bull 96:45–59. https://doi.org/10.1093/bmb/ldq035
Donald K, Matthew EL (2017) Nontuberculous mycobacteria: pathogens of growing importance. Infectious Disease News. Source, Healiocom https://www.healio.com/infectious-disease/emerging-diseases/news/print/infectious-disease-news/%7Bc9e68741-d302-4d75-ada0-064c1db5a5e7%7D/nontuberculous-mycobacteria-pathogens-of-growing-importance
Misch EA, Saddler C, Davis JM (2018) Skin and soft tissue infections due to nontuberculous mycobacteria. Curr Infect Dis Rep 20:6. https://doi.org/10.1007/s11908-018-0611-3
Sethi S, Arora S, Gupta V et al (2014) Cutaneous Mycobacterium fortuitum infection: successfully treated with amikacin and ofloxacin combination. Indian J Dermatol 59:383–384. https://doi.org/10.4103/0019-5154.135491
NIH (2019) Mycobacterium avium complex infections. National Institute of Health, Bethesda Source: https://rarediseases.info.nih.gov/diseases/7123/mycobacterium-avium-complex-infections
Weinberger M, Berg SL, Feuerstein IM et al (1992) Disseminated infection with Mycobacterium gordonae: report of a case and critical review of the literature. Clin Infect Dis 14:1229–1239. https://doi.org/10.1093/clinids/14.6.1229
van Ingen J, Boeree MJ, de Lange WC et al (2008) Mycobacterium xenopi clinical relevance and determinants, the Netherlands. Emerg Infect Dis 14:385–389. https://doi.org/10.3201/eid1403.061393
Oh TH, Kim UJ, Kang SJ et al (2018) Disseminated invasive Mycobacterium marinum infection involving the lung of a patient with diabetes mellitus. Infect Chemother 50:59–64. https://doi.org/10.3947/ic.2018.50.1.59
Akram SM, Rawla P (2019) Mycobacterium Kansasii. StatPearls. Treasure Island (FL). Source: https://www.ncbi.nlm.nih.gov/books/NBK430906/
NIH (2019) Mycobacterium Malmoense. National Institute of Health. Source: https://rarediseases.info.nih.gov/diseases/10549/mycobacterium-malmoense
van Ingen J, Boeree MJ, de Lange WC et al (2008) Clinical relevance of Mycobacterium szulgai in The Netherlands. Clin Infect Dis 46:1200–1205. https://doi.org/10.1086/529443
Sotello D, Hata DJ, Reza M et al (2017) Disseminated Mycobacterium interjectum infection with bacteremia, hepatic and pulmonary involvement associated with a long-term catheter infection. Case Rep Infect Dis 2017:6958204. https://doi.org/10.1155/2017/6958204
Gupta RS, Lo B, Son J (2018) Phylogenomics and comparative genomic studies robustly support division of the genus Mycobacterium into an emended genus Mycobacterium and four novel genera. Front Microbiol 9. doi:https://doi.org/10.3389/fmicb.2018.00067
Bernardelli A (2007) Manual de Procedimientos: Clasificación fenotípica de las micobacterias. Dirección de Laboratorio y Control Técnico. Source: http://www.senasa.gov.ar/Archivos/File/File1443-mlab.pdf-BioSource
NCBI (Accessed on 9/10/2019) Mycobacterium genome. National Center for Biotechnology. Source: https://www.ncbi.nlm.nih.gov/genome/
Howard ST, Rhoades E, Recht J et al (2006) Spontaneous reversion of Mycobacterium abscessus from a smooth to a rough morphotype is associated with reduced expression of glycopeptidolipid and reacquisition of an invasive phenotype. Microbiology 152:1581–1590. https://doi.org/10.1099/mic.0.28625-0
Rose SJ, Bermudez LE (2016) Identification of bicarbonate as a trigger and genes involved with extracellular DNA export in mycobacterial biofilms. MBio 7:e01597–e01516. https://doi.org/10.1128/mBio.01597-16
Lopez-Marin LM, Gautier N, Laneelle MA et al (1994) Structures of the glycopeptidolipid antigens of Mycobacterium abscessus and Mycobacterium chelonae and possible chemical basis of the serological cross-reactions in the Mycobacterium fortuitum complex. Microbiology 140(Pt 5):1109–1118. https://doi.org/10.1099/13500872-140-5-1109
Aung TT, Yam JK, Lin S et al (2016) Biofilms of pathogenic nontuberculous mycobacteria targeted by new therapeutic approaches. Antimicrob Agents Chemother 60:24–35. https://doi.org/10.1128/AAC.01509-15
Belisle JT, Klaczkiewicz K, Brennan PJ et al (1993) Rough morphological variants of Mycobacterium avium. Characterization of genomic deletions resulting in the loss of glycopeptidolipid expression. J Biol Chem 268:10517–10523
Rose SJ, Babrak LM, Bermudez LE (2015) Mycobacterium avium possesses extracellular DNA that contributes to biofilm formation, structural integrity, and tolerance to antibiotics. PLoS One 10:e0128772. https://doi.org/10.1371/journal.pone.0128772
Ren H, Dover LG, Islam ST et al (2007) Identification of the lipooligosaccharide biosynthetic gene cluster from Mycobacterium marinum. Mol Microbiol 63:1345–1359. https://doi.org/10.1111/j.1365-2958.2007.05603.x
De Baere T, Moerman M, Rigouts L et al (2001) Mycobacterium interjectum as causative agent of cervical lymphadenitis. J Clin Microbiol 39:725–727. https://doi.org/10.1128/jcm.39.2.725-727.2001
Brode SK, Marchand-Austin A, Jamieson FB et al (2017) Pulmonary versus nonpulmonary nontuberculous mycobacteria, Ontario, Canada. Emerg Infect Dis 23:1898–1901. https://doi.org/10.3201/eid2311.170959
Hoefsloot W, van Ingen J, Andrejak C et al (2013) The geographic diversity of nontuberculous mycobacteria isolated from pulmonary samples: an NTM-NET collaborative study. Eur Respir J 42:1604–1613. https://doi.org/10.1183/09031936.00149212
Al-Ghafli H, Al-Hajoj S (2017) Nontuberculous mycobacteria in Saudi Arabia and Gulf Countries: a review. Can Respir J 2017:5035932. https://doi.org/10.1155/2017/5035932
Jyoti U, Dharamveer S, Amreen Z et al (2016) Prevalence and species spectrum of both pulmonary and extrapulmonary nontuberculous mycobacteria isolates at a tertiary care center. Int J Mycobacteriol 5:288–293
Lim AYH, Chotirmall SH, Fok ETK et al (2018) Profiling non-tuberculous mycobacteria in an Asian setting: characteristics and clinical outcomes of hospitalized patients in Singapore. BMC Pulm Med 18:85. https://doi.org/10.1186/s12890-018-0637-1
Ong CS, Ngeow YF, Yap SF et al (2008) Molecular identification of nontuberculous mycobacteria from clinical sources by hsp65 PRA and sequence analysis. Int J Infect Dis 12:e322–e323. https://doi.org/10.1016/j.ijid.2008.05.863
Namkoong H, Kurashima A, Morimoto K et al (2016) Epidemiology of pulmonary nontuberculous mycobacterial disease, Japan(1). Emerg Infect Dis 22:1116–1117. https://doi.org/10.3201/eid2206.151086
Lin C, Russell C, Soll B et al (2018) Increasing prevalence of nontuberculous mycobacteria in respiratory specimens from US-affiliated pacific island jurisdictions. Emerg Infect Dis 24:485–491. https://doi.org/10.3201/eid2403.171301
Chou MP, Clements AC, Thomson RM (2014) A spatial epidemiological analysis of nontuberculous mycobacterial infections in Queensland, Australia. BMC Infect Dis 14:279. https://doi.org/10.1186/1471-2334-14-279
Brode SK, Daley CL, Marras TK (2014) The epidemiologic relationship between tuberculosis and non-tuberculous mycobacterial disease: a systematic review. Int J Tuberc Lung Dis 18:1370–1377. https://doi.org/10.5588/ijtld.14.0120
Adjemian J, Daniel-Wayman S, Ricotta E et al (2018) epidemiology of nontuberculous mycobacteriosis. Semin Respir Crit Care Med 39:325–335. https://doi.org/10.1055/s-0038-1651491
Vinnard C, Longworth S, Mezochow A et al (2016) Deaths related to nontuberculous mycobacterial infections in the United States, 1999–2014. Ann Am Thorac Soc 13:1951–1955. https://doi.org/10.1513/AnnalsATS.201606-474BC
Rivero-Lezcano OM, Gonzalez-Cortes C, Mirsaeidi M (2019) The unexplained increase of nontuberculous mycobacteriosis. Int J Mycobacteriol 8:1–6. https://doi.org/10.4103/ijmy.ijmy_18_19
Prevots DR, Adjemian J, Fernandez AG et al (2014) Environmental risks for nontuberculous mycobacteria. Individual exposures and climatic factors in the cystic fibrosis population. Ann Am Thorac Soc 11:1032–1038. https://doi.org/10.1513/AnnalsATS.201404-184OC
McShane PJ, Glassroth J (2015) Pulmonary disease due to nontuberculous mycobacteria: current state and new insights. Chest 148:1517–1527. https://doi.org/10.1378/chest.15-0458
Daniel-Wayman S, Abate G, Barber DL et al (2018) Advancing translational science for pulmonary ntm infections: a roadmap for research. Am J Respir Crit Care Med. https://doi.org/10.1164/rccm.201807-1273PP
Henkle E, Aksamit T, Barker A et al (2016) Patient-Centered Research Priorities for pulmonary nontuberculous mycobacteria (NTM) infection. An NTM Research Consortium Workshop Report. Ann Am Thorac Soc 13:S379–S384. https://doi.org/10.1513/AnnalsATS.201605-387WS
Rennard S, Thomashow B, Crapo J et al (2013) Introducing the COPD Foundation guide for diagnosis and management of COPD, recommendations of the COPD Foundation. COPD 10:378–389. https://doi.org/10.3109/15412555.2013.801309
GOLD (2018) Global strategy for the diagnosis, management and prevention of chronic obstructive pulmonary disease. Global Initiative for Chronic Obstructive Lung Disease. Source: www.goldcopd.org 1–142
Balavoine C, Andrejak C, Marchand-Adam S et al (2017) Relationships between COPD and nontuberculous mycobacteria pulmonary infections. Rev Mal Respir 34:1091–1097. https://doi.org/10.1016/j.rmr.2017.09.004
Pyarali FF, Schweitzer M, Bagley V et al (2018) Increasing non-tuberculous mycobacteria infections in veterans with COPD and association with increased risk of mortality. Front Med (Lausanne) 5:311. https://doi.org/10.3389/fmed.2018.00311
Berra G, Plojoux J, Soccal PM et al (2019) Identification of non-tuberculous mycobacteria in COPD patients undergoing lung volume reduction: more frequent than expected? Respiration 98:279–280. https://doi.org/10.1159/000501697
Yeh JJ, Wang YC, Sung FC et al (2014) Nontuberculosis mycobacterium disease is a risk factor for chronic obstructive pulmonary disease: a nationwide cohort study. Lung 192:403–411. https://doi.org/10.1007/s00408-014-9574-9
Char A, Hopkinson NS, Hansell DM et al (2014) Evidence of mycobacterial disease in COPD patients with lung volume reduction surgery; the importance of histological assessment of specimens: a cohort study. BMC Pulm Med 14:124. https://doi.org/10.1186/1471-2466-14-124
Andrejak C, Nielsen R, Thomsen VO et al (2013) Chronic respiratory disease, inhaled corticosteroids and risk of non-tuberculous mycobacteriosis. Thorax 68:256–262. https://doi.org/10.1136/thoraxjnl-2012-201772
Fowler SJ, French J, Screaton NJ et al (2006) Nontuberculous mycobacteria in bronchiectasis: prevalence and patient characteristics. Eur Respir J 28:1204–1210. https://doi.org/10.1183/09031936.06.00149805
Chu H, Zhao L, Xiao H et al (2014) Prevalence of nontuberculous mycobacteria in patients with bronchiectasis: a meta-analysis. Arch Med Sci 10:661–668. https://doi.org/10.5114/aoms.2014.44857
McShane PJ, Naureckas ET, Tino G et al (2013) Non–cystic fibrosis bronchiectasis. Am J Respir Crit Care Med 188:647–656. https://doi.org/10.1164/rccm.201303-0411CI
Flume PA, Chalmers JD, Olivier KN (2018) Advances in bronchiectasis: endotyping, genetics, microbiome, and disease heterogeneity. Lancet 392:880–890. https://doi.org/10.1016/S0140-6736(18)31767-7
Xu J-F, Ji X-B, Li H-P, et al. (2013) Bronchiectasis caused by pulmonary tuberculosis: the epidemiology, clinical presentations and the differences from non-tuberculosis-caused bronchiectasis. Eur Respir J 42
Sonnenberg P, Murray J, Glynn JR et al (2000) Risk factors for pulmonary disease due to culture-positive M. tuberculosis or nontuberculous mycobacteria in South African gold miners. Eur Respir J 15:291–296
WHO (2018) Fact sheets: Tuberculosis. World Health Organization, Geneva. Source: http://www.who.int/en/news-room/fact-sheets/detail/tuberculosis
Kee SJ, Suh SP (2017) Increasing burden of nontuberculous mycobacteria in Korea. J Korean Med Sci 32:1215–1216. https://doi.org/10.3346/jkms.2017.32.8.1215
Griesenbach U, Alton EW (2015) Recent advances in understanding and managing cystic fibrosis transmembrane conductance regulator dysfunction. F1000Prime Rep 7:64. https://doi.org/10.12703/P7-64
Goetz D, Ren CL (2019) Review of cystic fibrosis. Pediatr Ann 48:e154–e161. https://doi.org/10.3928/19382359-20190327-01
Olivier KN, Weber DJ, Wallace RJ Jr et al (2003) Nontuberculous mycobacteria: multicenter prevalence study in cystic fibrosis. Am J Respir Crit Care Med 167:828–834. https://doi.org/10.1164/rccm.200207-678OC
Olivier KN (2004) The natural history of nontuberculous mycobacteria in patients with cystic fibrosis. Paediatr Respir Rev 5(Suppl A):S213–S216
Floto RA, Olivier KN, Saiman L et al (2016) US Cystic Fibrosis Foundation and European Cystic Fibrosis Society consensus recommendations for the management of non-tuberculous mycobacteria in individuals with cystic fibrosis: executive summary. Thorax 71:88–90. https://doi.org/10.1136/thoraxjnl-2015-207983
Cowman SA, Jacob J, Hansell DM et al (2018) Whole-blood gene expression in pulmonary nontuberculous mycobacterial infection. Am J Respir Cell Mol Biol 58:510–518. https://doi.org/10.1165/rcmb.2017-0230OC
Greenberger PA (2002) Allergic bronchopulmonary aspergillosis. J Allergy Clin Immunol 110:685–692. https://doi.org/10.1067/mai.2002.130179
Zander DS (2005) Allergic bronchopulmonary aspergillosis: an overview. Arch Pathol Lab Med 129:924–928. https://doi.org/10.1043/1543-2165(2005)129[924:ABAAO]2.0.CO;2
Agarwal R (2009) Allergic bronchopulmonary aspergillosis. Chest 135:805–826. https://doi.org/10.1378/chest.08-2586
Ishiguro T, Takayanagi N, Baba Y et al (2016) Pulmonary nontuberculous mycobacteriosis and chronic lower respiratory tract infections in patients with allergic bronchopulmonary mycosis without cystic fibrosis. Intern Med 55:1067–1070. https://doi.org/10.2169/internalmedicine.55.5561
Kunst H, Wickremasinghe M, Wells A et al (2006) Nontuberculous mycobacterial disease and Aspergillus-related lung disease in bronchiectasis. Eur Respir J 28:352–357. https://doi.org/10.1183/09031936.06.00139005
Naito M, Kurahara Y, Yoshida S et al (2018) Prognosis of chronic pulmonary aspergillosis in patients with pulmonary non-tuberculous mycobacterial disease. Respir Investig 56:326–331. https://doi.org/10.1016/j.resinv.2018.04.002
Redondo M, Keyt H, Dhar R et al (2016) Global impact of bronchiectasis and cystic fibrosis. Breathe (Sheff) 12:222–235. https://doi.org/10.1183/20734735.007516
Knowles MR, Daniels LA, Davis SD et al (2013) Primary ciliary dyskinesia. Recent advances in diagnostics, genetics, and characterization of clinical disease. Am J Respir Crit Care Med 188:913–922. https://doi.org/10.1164/rccm.201301-0059CI
Fowler CJ, Olivier KN, Leung JM et al (2013) Abnormal nasal nitric oxide production, ciliary beat frequency, and toll-like receptor response in pulmonary nontuberculous mycobacterial disease epithelium. Am J Respir Crit Care Med 187:1374–1381. https://doi.org/10.1164/rccm.201212-2197OC
Matsuyama M, Martins AJ, Shallom S et al (2018) Transcriptional response of respiratory epithelium to nontuberculous mycobacteria. Am J Respir Cell Mol Biol 58:241–252. https://doi.org/10.1165/rcmb.2017-0218OC
Becker KL, Arts P, Jaeger M et al (2017) MST1R mutation as a genetic cause of Lady Windermere syndrome. Eur Respir J 49. https://doi.org/10.1183/13993003.01478-2016
Namkoong H, Omae Y, Asakura T, et al. (2019) Genome-wide association study in patients with pulmonary Mycobacterium avium complex disease. Am Thorac Soc pp A1031-A1031
Szymanski EP, Leung JM, Fowler CJ et al (2015) Pulmonary nontuberculous mycobacterial infection: a multisystem, multigenic disease. Am J Respir Crit Care Med 192:618–628. https://doi.org/10.1164/rccm.201502-0387OC
Chen F, Szymanski EP, Olivier KN et al (2017) Whole-exome sequencing identifies the 6q12-q16 linkage region and a candidate gene, TTK, for pulmonary nontuberculous mycobacterial disease. Am J Respir Crit Care Med 196:1599–1604. https://doi.org/10.1164/rccm.201612-2479OC
Halstrom S, Cherry CL, Black M et al (2017) A haplotype spanning P2X7R, P2X4R and CAMKK2 may mark susceptibility to pulmonary non-tuberculous mycobacterial disease. Immunogenetics 69:287–293. https://doi.org/10.1007/s00251-017-0972-z
Takahashi M, Ishizaka A, Nakamura H et al (2000) Specific HLA in pulmonary MAC infection in a Japanese population. Am J Respir Crit Care Med 162:316–318. https://doi.org/10.1164/ajrccm.162.1.9908071
Kubo K, Yamazaki Y, Hanaoka M et al (2000) Analysis of HLA antigens in Mycobacterium avium-intracellulare pulmonary infection. Am J Respir Crit Care Med 161:1368–1371. https://doi.org/10.1164/ajrccm.161.4.9906094
Picard C, Bobby Gaspar H, Al-Herz W et al (2018) International Union of Immunological Societies: 2017 Primary Immunodeficiency Diseases Committee Report on Inborn Errors of Immunity. J Clin Immunol 38:96–128. https://doi.org/10.1007/s10875-017-0464-9
Mortaz E, Moloudizargari M, Varahram M et al (2018) what immunological defects predispose to non-tuberculosis mycobacterial infections? Iran J Allergy Asthma Immunol 17:100–109
Lake MA, Ambrose LR, Lipman MC et al (2016) “Why me, why now?” Using clinical immunology and epidemiology to explain who gets nontuberculous mycobacterial infection. BMC Med 14:54. https://doi.org/10.1186/s12916-016-0606-6
de los Angeles Garcia M, Borrero R, Marron R et al (2013) Evaluation of specific humoral immune response and cross reactivity against Mycobacterium tuberculosis antigens induced in mice immunized with liposomes composed of total lipids extracted from Mycobacterium smegmatis. BMC Immunol 14(Suppl 1):S11. https://doi.org/10.1186/1471-2172-14-S1-S11
Lee WI, Huang JL, Yeh KW et al (2011) Immune defects in active mycobacterial diseases in patients with primary immunodeficiency diseases (PIDs). J Formos Med Assoc 110:750–758. https://doi.org/10.1016/j.jfma.2011.11.004
Ottenhoff TH, Verreck FA, Hoeve MA et al (2005) Control of human host immunity to mycobacteria. Tuberculosis (Edinb) 85:53–64. https://doi.org/10.1016/j.tube.2004.09.011
Lovell JP, Zerbe CS, Olivier KN et al (2016) Mediastinal and disseminated Mycobacterium kansasii disease in GATA2 deficiency. Ann Am Thorac Soc 13:2169–2173. https://doi.org/10.1513/AnnalsATS.201603-207BC
Eguchi K, Ishimura M, Sonoda M et al (2018) Nontuberculous mycobacteria-associated hemophagocytic lymphohistiocytosis in MonoMAC syndrome. Pediatr Blood Cancer 65:e27017. https://doi.org/10.1002/pbc.27017
Mendes-de-Almeida DP, Andrade FG, Borges G et al (2019) GATA2 mutation in long stand Mycobacterium kansasii infection, myelodysplasia and MonoMAC syndrome: a case-report. BMC Med Genet 20:64. https://doi.org/10.1186/s12881-019-0799-6
Fakhri B, Cashen AF, Duncavage EJ et al (2019) Fifty shades of GATA2 mutation: a case of plasmablastic lymphoma, nontuberculous mycobacterial infection, and myelodysplastic syndrome. Clin Lymphoma Myeloma Leuk 19:e532–e535. https://doi.org/10.1016/j.clml.2019.05.015
Al-Kzayer LaFY, Yassin AK, Salih KH, et al. (2019) A Syrian refugee in Iraq diagnosed as a case of IL12RB1 deficiency in Japan using dried blood spots. Front Immunol 10. doi:https://doi.org/10.3389/fimmu.2019.00058
Nekooie-Marnany N, Deswarte C, Ostadi V et al (2018) Impaired IL-12- and IL-23-mediated immunity due to IL-12Rβ1 deficiency in Iranian patients with Mendelian Susceptibility to Mycobacterial Disease. J Clin Immunol 38:787–793. https://doi.org/10.1007/s10875-018-0548-1
Serour F, Mizrahi A, Somekh E et al (2007) Analysis of the interleukin-12/interferon-gamma pathway in children with non-tuberculous mycobacterial cervical lymphadenitis. Eur J Pediatr 166:835–841. https://doi.org/10.1007/s00431-006-0338-2
Doffinger R, Dupuis S, Picard C et al (2002) Inherited disorders of IL-12- and IFN-gamma-mediated immunity: a molecular genetics update. Mol Immunol 38:903–909. https://doi.org/10.1016/s0161-5890(02)00017-2
Krisnawati DI, Liu YC, Lee YJ et al (2019) Blockade effects of anti-interferon- (IFN-) gamma autoantibodies on IFN-gamma-regulated antimicrobial immunity. J Immunol Res 2019:1629258. https://doi.org/10.1155/2019/1629258
Liew WK, Thoon KC, Chong CY et al (2019) Juvenile-onset immunodeficiency secondary to anti-interferon-gamma autoantibodies. J Clin Immunol 39:512–518. https://doi.org/10.1007/s10875-019-00652-1
Tanaka S, Hoshino Y, Sakagami T et al (2019) Pathogenicity of Mycolicibacterium phlei, a non-pathogenic nontuberculous mycobacterium in an immunocompetent host carrying anti-interferon gamma autoantibodies: a case report. BMC Infect Dis 19:454. https://doi.org/10.1186/s12879-019-4050-z
Krisnawati DI, Liu YC, Lee YJ et al (2019) Functional neutralization of anti-IFN-gamma autoantibody in patients with nontuberculous mycobacteria infection. Sci Rep 9:5682. https://doi.org/10.1038/s41598-019-41952-1
Chetchotisakd P, Anunnatsiri S, Nanagara R et al (2018) Intravenous cyclophosphamide therapy for anti-IFN-gamma autoantibody-associated Mycobacterium abscessus infection. J Immunol Res 2018:6473629. https://doi.org/10.1155/2018/6473629
Hase I, Morimoto K, Sakagami T et al (2017) Patient ethnicity and causative species determine the manifestations of anti-interferon-gamma autoantibody-associated nontuberculous mycobacterial disease: a review. Diagn Microbiol Infect Dis 88:308–315. https://doi.org/10.1016/j.diagmicrobio.2017.05.011
Phoompoung P, Ankasekwinai N, Pithukpakorn M et al (2017) Factors associated with acquired anti IFN- gamma autoantibody in patients with nontuberculous mycobacterial infection. PLoS One 12:e0176342. https://doi.org/10.1371/journal.pone.0176342
Hase I, Morimoto K, Sakagami T et al (2015) Disseminated Mycobacterium gordonae and Mycobacterium mantenii infection with elevated anti-IFN-gamma neutralizing autoantibodies. J Infect Chemother 21:468–472. https://doi.org/10.1016/j.jiac.2015.02.003
DeLeon TT, Chung HH, Opal SM et al (2014) Mycobacterium avium complex empyema in a patient with interferon gamma autoantibodies. Hawaii J Med Public Health 73:15–17
Kampitak T, Suwanpimolkul G, Browne S et al (2011) Anti-interferon-gamma autoantibody and opportunistic infections: case series and review of the literature. Infection 39:65–71. https://doi.org/10.1007/s15010-010-0067-3
Browne SK, Holland SM (2010) Immunodeficiency secondary to anticytokine autoantibodies. Curr Opin Allergy Clin Immunol 10:534–541. https://doi.org/10.1097/ACI.0b013e3283402b41
Kotilainen H, Lokki ML, Paakkanen R et al (2014) Complement C4 deficiency: a plausible risk factor for non-tuberculous mycobacteria (NTM) infection in apparently immunocompetent patients. PLoS One 9:e91450. https://doi.org/10.1371/journal.pone.0091450
Nightingale SD, Byrd LT, Southern PM et al (1992) Incidence of Mycobacterium avium-intracellulare complex bacteremia in human immunodeficiency virus-positive patients. J Infect Dis 165:1082–1085. https://doi.org/10.1093/infdis/165.6.1082
Young LS, Inderlied CB, Berlin OG et al (1986) Mycobacterial infections in AIDS patients, with an emphasis on the Mycobacterium avium complex. Rev Infect Dis 8:1024–1033. https://doi.org/10.1093/clinids/8.6.1024
MacDonell KB, Glassroth J (1989) Mycobacterium avium complex and other nontuberculous mycobacteria in patients with HIV infection. Semin Respir Infect 4:123–132
Heidary M, Nasiri MJ, Mirsaeidi M et al (2019) Mycobacterium avium complex infection in patients with human immunodeficiency virus: a systematic review and meta-analysis. J Cell Physiol 234:9994–10001. https://doi.org/10.1002/jcp.27859
Hoover DR, Graham NM, Bacellar H et al (1995) An epidemiologic analysis of Mycobacterium avium complex disease in homosexual men infected with human immunodeficiency virus type 1. Clin Infect Dis 20:1250–1258
Zakowski P, Fligiel S, Berlin GW et al (1982) Disseminated Mycobacterium avium-intracellulare infection in homosexual men dying of acquired immunodeficiency. JAMA 248:2980–2982
Varley CD, Ku JH, Henkle E et al (2017) Disseminated nontuberculous mycobacteria in HIV-infected patients, Oregon, USA, 2007–2012. Emerg Infect Dis 23:533–535. https://doi.org/10.3201/eid2303.161708
Ristola MA, von Reyn CF, Arbeit RD et al (1999) High rates of disseminated infection due to non-tuberculous mycobacteria among AIDS patients in Finland. J Inf Secur 39:61–67. https://doi.org/10.1016/S0163-4453(99)90104-4
Karakousis PC, Moore RD, Chaisson RE (2004) Mycobacterium avium complex in patients with HIV infection in the era of highly active antiretroviral therapy. Lancet Infect Dis 4:557–565. https://doi.org/10.1016/S1473-3099(04)01130-2
Kobayashi T, Nishijima T, Teruya K et al (2016) High Mortality of disseminated non-tuberculous mycobacterial infection in HIV-infected patients in the antiretroviral therapy era. PLoS One 11:e0151682. https://doi.org/10.1371/journal.pone.0151682
Takenaka S, Ogura T, Oshima H, et al. (2019) Development and exacerbation of pulmonary nontuberculous mycobacterial infection in patients with systemic autoimmune rheumatic diseases. Mod Rheumatol: 1–6. doi:https://doi.org/10.1080/14397595.2019.1619220
Chen Y-M, Lin C-H, Chen H-H et al (2019) Risk of mycobacterial disease among cancer patients: a population-based cohort study in a TB endemic area. Cancer Epidemiol 59:64–70. https://doi.org/10.1016/j.canep.2019.01.010
Chen CY, Sheng WH, Lai CC et al (2012) Mycobacterial infections in adult patients with hematological malignancy. Eur J Clin Microbiol Infect Dis 31:1059–1066. https://doi.org/10.1007/s10096-011-1407-7
Lai CC, Tan CK, Cheng A et al (2012) Nontuberculous mycobacterial infections in cancer patients in a medical center in Taiwan, 2005–2008. Diagn Microbiol Infect Dis 72:161–165. https://doi.org/10.1016/j.diagmicrobio.2011.10.006
Vial T, Descotes J (2003) Immunosuppressive drugs and cancer. Toxicology 185:229–240
Assfalg V, Huser N, Reim D et al (2010) Combined immunosuppressive and antibiotic therapy improves bacterial clearance and survival of polymicrobial septic peritonitis. Shock 33:155–161. https://doi.org/10.1097/SHK.0b013e3181ab9014
Bach JF (1993) Immunosuppressive therapy of autoimmune diseases. Immunol Today 14:322–326. https://doi.org/10.1016/0167-5699(93)90053-N
Fishman JA (2013) Opportunistic infections: coming to the limits of immunosuppression? Cold Spring Harb Perspect Med 3:a015669. https://doi.org/10.1101/cshperspect.a015669
Winthrop KL, Chang E, Yamashita S et al (2009) Nontuberculous mycobacteria infections and anti-tumor necrosis factor-alpha therapy. Emerg Infect Dis 15:1556–1561. https://doi.org/10.3201/eid1510.090310
Longworth SA, Vinnard C, Lee I et al (2014) Risk factors for nontuberculous mycobacterial infections in solid organ transplant recipients: a case-control study. Transpl Infect Dis 16:76–83. https://doi.org/10.1111/tid.12170
Kurosawa S, Kato M (2008) Anesthetics, immune cells, and immune responses. J Anesth 22:263–277. https://doi.org/10.1007/s00540-008-0626-2
Cardinale F, Chinellato I, Caimmi S, et al. (2011) Perioperative period: immunological modifications. Int J Immunopathol Pharmacol 24: S3–12
Angele MK, Chaudry IH (2005) Surgical trauma and immunosuppression: pathophysiology and potential immunomodulatory approaches. Langenbeck's Arch Surg 390:333–341. https://doi.org/10.1007/s00423-005-0557-4
Shakhar G, Ben-Eliyahu S (2003) Potential prophylactic measures against postoperative immunosuppression: could they reduce recurrence rates in oncological patients? Ann Surg Oncol 10:972–992. https://doi.org/10.1245/aso.2003.02.007
Doucette K, Fishman JA (2004) Nontuberculous mycobacterial infection in hematopoietic stem cell and solid organ transplant recipients. Clin Infect Dis 38:1428–1439. https://doi.org/10.1086/420746
Abd CL, Razonable RR (2016) Non-tuberculous mycobacterial infections in solid organ transplant recipients: an update. J Clin Tuberc and Other Mycobact Dis 4:1–8
Beswick J, Shin E, Michelis FV et al (2018) Incidence and risk factors for nontuberculous mycobacterial infection after allogeneic hematopoietic cell transplantation. Biol Blood Marrow Transplant 24:366–372. https://doi.org/10.1016/j.bbmt.2017.09.015
Hirama T, Brode SK, Beswick J et al (2018) Characteristics, treatment and outcomes of nontuberculous mycobacterial pulmonary disease after allogeneic haematopoietic stem cell transplant. Eur Respir J 51. https://doi.org/10.1183/13993003.02330-2017
Kang JY, Ha JH, Kang HS et al (2015) Clinical significance of nontuberculous mycobacteria from respiratory specimens in stem cell transplantation recipients. Int J Hematol 101:505–513. https://doi.org/10.1007/s12185-015-1745-9
Worldometers (2018) Current world population. Source: http://www.worldometersinfo/world-population/#ref-1
Ben-Haim MS, Kanfi Y, Mitchell SJ et al (2018) Breaking the ceiling of human maximal life span. J Gerontol A Biol Sci Med Sci 73:1465–1471. https://doi.org/10.1093/gerona/glx219
Weiskopf D, Weinberger B, Grubeck-Loebenstein B (2009) The aging of the immune system. Transpl Int 22:1041–1050. https://doi.org/10.1111/j.1432-2277.2009.00927.x
Gubbels Bupp MR, Potluri T, Fink AL et al (2018) The confluence of sex hormones and aging on immunity. Front Immunol 9:1269. https://doi.org/10.3389/fimmu.2018.01269
Brandsma C-A, de Vries M, Costa R et al (2017) Lung ageing and COPD: is there a role for ageing in abnormal tissue repair? Eur Respir Rev 26:170073. https://doi.org/10.1183/16000617.0073-2017
Barnes PJ (2019) Pulmonary diseases and ageing. Subcell Biochem 91:45–74. https://doi.org/10.1007/978-981-13-3681-2_3
Quinn TM, Hill AT (2018) Non-cystic fibrosis bronchiectasis in the elderly: current perspectives. Clin Interv Aging 13:1649–1656. https://doi.org/10.2147/CIA.S143139
Adjemian J, Olivier KN, Seitz AE et al (2012) Prevalence of nontuberculous mycobacterial lung disease in U.S. Medicare beneficiaries. Am J Respir Crit Care Med 185:881–886. https://doi.org/10.1164/rccm.201111-2016OC
Xu J, Li P, Zheng S et al (2019) Prevalence and risk factors of pulmonary nontuberculous mycobacterial infections in the Zhejiang Province of China. Epidemiol Infect 147:e269. https://doi.org/10.1017/S0950268819001626
Hu C, Huang L, Cai M et al (2019) Characterization of non-tuberculous mycobacterial pulmonary disease in Nanjing district of China. BMC Infect Dis 19:764. https://doi.org/10.1186/s12879-019-4412-6
Tan Y, Su B, Shu W et al (2018) Epidemiology of pulmonary disease due to nontuberculous mycobacteria in Southern China, 2013–2016. BMC Pulm Med 18:168. https://doi.org/10.1186/s12890-018-0728-z
Izumi K, Morimoto K, Hasegawa N et al (2019) Epidemiology of adults and children treated for nontuberculous mycobacterial pulmonary disease in Japan. Ann Am Thorac Soc 16:341–347. https://doi.org/10.1513/AnnalsATS.201806-366OC
NIH (2011) Global Health and Aging. National Institute of Aging, Source http://www.who.int/ageing/publications/global_health.pdf
Chandra RK (1981) Immunodeficiency in undernutrition and overnutrition. Nutr Rev 39:225–231. https://doi.org/10.1111/j.1753-4887.1981.tb07446.x
Honce R, Schultz-Cherry S (2019) Impact of obesity on Influenza A virus pathogenesis, immune response, and evolution. Front Immunol 10:1071. https://doi.org/10.3389/fimmu.2019.01071
Frasca D, McElhaney J (2019) Influence of obesity on pneumococcus infection risk in the elderly. Front Endocrinol (Lausanne) 10:71. https://doi.org/10.3389/fendo.2019.00071
Falagas ME, Kompoti M (2006) Obesity and infection. Lancet Infect Dis 6:438–446. https://doi.org/10.1016/S1473-3099(06)70523-0
Milner JJ, Beck MA (2012) The impact of obesity on the immune response to infection. Proc Nutr Soc 71:298–306. https://doi.org/10.1017/S0029665112000158
Marti A, Marcos A, Martinez JA (2001) Obesity and immune function relationships. Obes Rev 2:131–140
Lopez-Varela E, Garcia-Basteiro AL, Augusto OJ et al (2017) High rates of non-tuberculous mycobacteria isolation in Mozambican children with presumptive tuberculosis. PLoS One 12:e0169757. https://doi.org/10.1371/journal.pone.0169757
Ikegame S, Maki S, Wakamatsu K et al (2011) Nutritional assessment in patients with pulmonary nontuberculous mycobacteriosis. Intern Med 50:2541–2546. https://doi.org/10.2169/internalmedicine.50.5853
Wakamatsu K, Nagata N, Maki S et al (2015) Patients with MAC lung disease have a low visceral fat area and low nutrient intake. Pulm Med 2015:218253–218253. https://doi.org/10.1155/2015/218253
Hong JY, Yang GE, Ko Y et al (2016) Changes in cholesterol level correlate with the course of pulmonary nontuberculous mycobacterial disease. J Thorac Dis 8:2885–2894. https://doi.org/10.21037/jtd.2016.10.75
Jeon K, Kim SY, Jeong BH et al (2013) Severe vitamin D deficiency is associated with non-tuberculous mycobacterial lung disease: a case-control study. Respirology 18:983–988. https://doi.org/10.1111/resp.12109
Cunningham-Rundles S, McNeeley DF, Moon A (2005) Mechanisms of nutrient modulation of the immune response. J Allergy Clin Immunol 115:1119–1128; quiz 1129. https://doi.org/10.1016/j.jaci.2005.04.036
Chandra RK (2002) Nutrition and the immune system from birth to old age. Eur J Clin Nutr 56(Suppl 3):S73–S76. https://doi.org/10.1038/sj.ejcn.1601492
Ng SY, Bettany-Saltikov J, Cheung IYK et al (2018) The role of vitamin D in the pathogenesis of adolescent idiopathic scoliosis. Asian. Spine J. https://doi.org/10.31616/asj.2018.12.6.1127
Ariganjoye R (2017) Pediatric hypovitaminosis D: molecular perspectives and clinical implications. Glob Pediatr Health 4:2333794X16685504. https://doi.org/10.1177/2333794X16685504
Zosky GR, Berry LJ, Elliot JG et al (2011) Vitamin D deficiency causes deficits in lung function and alters lung structure. Am J Respir Crit Care Med 183:1336–1343. https://doi.org/10.1164/rccm.201010-1596OC
Chalmers JD, McHugh BJ, Docherty C et al (2013) Vitamin-D deficiency is associated with chronic bacterial colonisation and disease severity in bronchiectasis. Thorax 68:39–47. https://doi.org/10.1136/thoraxjnl-2012-202125
De Rui M, Toffanello ED, Veronese N et al (2014) Vitamin D deficiency and leisure time activities in the elderly: are all pastimes the same? PLoS One 9:e94805. https://doi.org/10.1371/journal.pone.0094805
Fujita K, Ito Y, Oguma T et al (2018) Association between Mycobacterium avium complex lung disease and serum vitamin D status, antimicrobial peptide levels, and bone mineral density. Medicine 97:e12463–e12463. https://doi.org/10.1097/md.0000000000012463
Gelder CM, Hart KW, Williams OM et al (2000) Vitamin D receptor gene polymorphisms and susceptibility to Mycobacterium malmoense pulmonary disease. J Infect Dis 181:2099–2102. https://doi.org/10.1086/315489
Oh J, Park HD, Kim SY et al (2019) Assessment of vitamin status in patients with nontuberculous mycobacterial pulmonary disease: potential role of vitamin A as a risk factor. Nutrients 11. https://doi.org/10.3390/nu11020343
Coleman MM, Basdeo SA, Coleman AM et al (2018) All-trans retinoic acid augments autophagy during intracellular bacterial infection. Am J Respir Cell Mol Biol 59:548–556. https://doi.org/10.1165/rcmb.2017-0382OC
Seyedrezazadeh E, Ostadrahimi A, Mahboob S et al (2008) Effect of vitamin E and selenium supplementation on oxidative stress status in pulmonary tuberculosis patients. Respirology 13:294–298. https://doi.org/10.1111/j.1440-1843.2007.01200.x
Oh J, Shin SH, Choi R et al (2019) Assessment of 7 trace elements in serum of patients with nontuberculous mycobacterial lung disease. J Trace Elem Med Biol 53:84–90. https://doi.org/10.1016/j.jtemb.2019.02.004
Friedman H, Pross S, Klein TW (2006) Addictive drugs and their relationship with infectious diseases. FEMS Immunol Med Microbiol 47:330–342. https://doi.org/10.1111/j.1574-695X.2006.00097.x
Magrone T, Russo MA, Jirillo E (2016) Cigarette smoke-mediated perturbations of the immune response: a new therapeutic approach with natural compounds. Endocr Metab Immune Disord Drug Targets 16:158–167. https://doi.org/10.2174/1872214810666160927123447
Qiu F, Liang CL, Liu H et al (2017) Impacts of cigarette smoking on immune responsiveness: up and down or upside down? Oncotarget 8:268–284. https://doi.org/10.18632/oncotarget.13613
Szabo G, Mandrekar P (2009) A recent perspective on alcohol, immunity, and host defense. Alcohol Clin Exp Res 33:220–232. https://doi.org/10.1111/j.1530-0277.2008.00842.x
Szabo G, Saha B (2015) Alcohol's effect on host defense. Alcohol Res 37:159–170
Magrone T, Jirillo E (2019) Drugs of abuse induced-subversion of the peripheral immune response and central glial activity: focus on novel therapeutic approaches. Endocr Metab Immune Disord Drug Targets 19:281–291. https://doi.org/10.2174/1871530319666181129104329
WHO (2018) Prevalence of tabacco smoking, 2016. World Health Organization, Geneva. Source: http://apps.who.int/gho/data/node.sdg.3-a-viz?lang=en
Krosnick JA, Malhotra N, Mo CH et al (2017) Perceptions of health risks of cigarette smoking: a new measure reveals widespread misunderstanding. PLoS One 12:e0182063. https://doi.org/10.1371/journal.pone.0182063
Watson NA, Clarkson JP, Donovan RJ et al (2003) Filthy or fashionable? Young people’s perceptions of smoking in the media. Health Educ Res 18:554–567
Eklund BM, Nilsson S, Hedman L et al (2012) Why do smokers diagnosed with COPD not quit smoking?—a qualitative study. Tob Induc Dis 10:17. https://doi.org/10.1186/1617-9625-10-17
Dube MF, Green CR (1982) Methods of collection of smoke for analytical purposes. Recent Adv Tob Sci 8:42–102
Cooke M (2010) The chemical components of tobacco and tobacco smoke. Chromatographia 71:977–977. https://doi.org/10.1365/s10337-010-1556-3
Fowles J, Dybing E (2003) Application of toxicological risk assessment principles to the chemical constituents of cigarette smoke. Tob Control 12:424–430. https://doi.org/10.1136/tc.12.4.424
Chong SG, Kent BD, Fitzgerald S et al (2014) Pulmonary non-tuberculous mycobacteria in a general respiratory population. Ir Med J 107:207–209
Davies BS, Roberts CH, Kaul S et al (2012) Non-tuberculous slow-growing mycobacterial pulmonary infections in non-HIV-infected patients in south London. Scand J Infect Dis 44:815–819. https://doi.org/10.3109/00365548.2012.694469
Kammerl IE, Dann A, Mossina A et al (2016) Impairment of immunoproteasome function by cigarette smoke and in chronic obstructive pulmonary disease. Am J Respir Crit Care Med 193:1230–1241. https://doi.org/10.1164/rccm.201506-1122OC
Pehote G, Bodas M, Brucia K et al (2017) Cigarette smoke exposure inhibits bacterial killing via TFEB-mediated autophagy impairment and resulting phagocytosis defect. Mediat Inflamm 2017:3028082. https://doi.org/10.1155/2017/3028082
Shi J, Li H, Yuan C et al (2018) Cigarette smoke-induced acquired dysfunction of cystic fibrosis transmembrane conductance regulator in the pathogenesis of chronic obstructive pulmonary disease. Oxidative Med Cell Longev 2018:6567578. https://doi.org/10.1155/2018/6567578
Ni I, Ji C, Vij N (2015) Second-hand cigarette smoke impairs bacterial phagocytosis in macrophages by modulating CFTR dependent lipid-rafts. PLoS One 10:e0121200. https://doi.org/10.1371/journal.pone.0121200
Raju SV, Jackson PL, Courville CA et al (2013) Cigarette smoke induces systemic defects in cystic fibrosis transmembrane conductance regulator function. Am J Respir Crit Care Med 188:1321–1330. https://doi.org/10.1164/rccm.201304-0733OC
Rab A, Rowe SM, Raju SV et al (2013) Cigarette smoke and CFTR: implications in the pathogenesis of COPD. Am J Phys Lung Cell Mol Phys 305:L530–L541. https://doi.org/10.1152/ajplung.00039.2013
Agusti A, Hogg JC (2019) Update on the pathogenesis of chronic obstructive pulmonary disease. N Engl J Med 381:1248–1256. https://doi.org/10.1056/NEJMra1900475
WHO (2018) Global status report on alcohol and health 2018. World Health Organization, Geneva. Source: http://apps.who.int/iris/bitstream/handle/10665/274603/9789241565639-eng.pdf?ua=1
WHO (2011) Global status report on alcohol and health. World Health Organization, Geneva. Source: http://www.who.int/substance_abuse/publications/global_alcohol_report/msbgsruprofiles.pdf
Sisson JH (2007) Alcohol and airways function in health and disease. Alcohol 41:293–307. https://doi.org/10.1016/j.alcohol.2007.06.003
Hulsebus HJ, Curtis BJ, Molina PE et al (2018) Summary of the 2017 Alcohol and Immunology Research Interest Group (AIRIG) meeting. Alcohol 69:51–56. https://doi.org/10.1016/j.alcohol.2017.10.006
Yeligar SM, Chen MM, Kovacs EJ et al (2016) Alcohol and lung injury and immunity. Alcohol 55:51–59. https://doi.org/10.1016/j.alcohol.2016.08.005
Lapinel N, Jolley SE, Ali J et al (2018) Association of substance abuse disorders in HIV patients with nontuberculous mycobacteria. Am Thorac Soc:A6103–A6103
Kim RD, Greenberg DE, Ehrmantraut ME et al (2008) Pulmonary nontuberculous mycobacterial disease: prospective study of a distinct preexisting syndrome. Am J Respir Crit Care Med 178:1066–1074. https://doi.org/10.1164/rccm.200805-686OC
Kartalija M, Ovrutsky AR, Bryan CL et al (2013) Patients with nontuberculous mycobacterial lung disease exhibit unique body and immune phenotypes. Am J Respir Crit Care Med 187:197–205. https://doi.org/10.1164/rccm.201206-1035OC
Chan ED, Iseman MD (2010) Slender, older women appear to be more susceptible to nontuberculous mycobacterial lung disease. Gend Med 7:5–18. https://doi.org/10.1016/j.genm.2010.01.005
Holt M, Kasperbauer SH, Levin A et al (2018) Nontuberculous mycobacterial pulmonary infections in men: The Lord Windermere syndrome? Am Thoracic Soc:A2606–A2606
Holt MR, Kasperbauer SH, Koelsch TL et al (2019) Similar characteristics of nontuberculous mycobacterial pulmonary disease in men and women. Eur Respir J 54. https://doi.org/10.1183/13993003.00252-2019
Fedrizzi T, Meehan CJ, Grottola A et al (2017) Genomic characterization of nontuberculous mycobacteria. Sci Rep 7:45258. https://doi.org/10.1038/srep45258
Gcebe N, Michel A, Gey van Pittius NC et al (2016) Comparative genomics and proteomic analysis of four non-tuberculous mycobacterium species and Mycobacterium tuberculosis complex: occurrence of shared immunogenic proteins. Front Microbiol 7:795. https://doi.org/10.3389/fmicb.2016.00795
van Ingen J, de Zwaan R, Dekhuijzen R et al (2009) Region of difference 1 in nontuberculous mycobacterium species adds a phylogenetic and taxonomical character. J Bacteriol 191:5865–5867. https://doi.org/10.1128/JB.00683-09
Arend SM, van Meijgaarden KE, de Boer K et al (2002) Tuberculin skin testing and in vitro T cell responses to ESAT-6 and culture filtrate protein 10 after infection with Mycobacterium marinum or M. kansasii. J Infect Dis 186:1797–1807. https://doi.org/10.1086/345760
Tran T, Bonham AJ, Chan ED et al (2019) A paucity of knowledge regarding nontuberculous mycobacterial lipids compared to the tubercle bacillus. Tuberculosis (Edinb) 115:96–107. https://doi.org/10.1016/j.tube.2019.02.008
Mukherjee R, Chatterji D (2012) Glycopeptidolipids: immuno-modulators in greasy mycobacterial cell envelope. IUBMB Life 64:215–225. https://doi.org/10.1002/iub.602
Daffe M, Crick DC, Jackson M (2014) Genetics of capsular polysaccharides and cell envelope (glyco)lipids. Microbiol Spectr 2: MGM2–0021-2013. doi:https://doi.org/10.1128/microbiolspec.MGM2-0021-2013
Sarmiento ME, Alvarez N, Chin KL et al (2019) Tuberculosis vaccine candidates based on mycobacterial cell envelope components. Tuberculosis (Edinb) 115:26–41. https://doi.org/10.1016/j.tube.2019.01.003
McNeil M, Tsang AY, Brennan PJ (1987) Structure and antigenicity of the specific oligosaccharide hapten from the glycopeptidolipid antigen of Mycobacterium avium serotype 4, the dominant Mycobacterium isolated from patients with acquired immune deficiency syndrome. J Biol Chem 262:2630–2635
Daffe M, McNeil M, Brennan PJ (1991) Novel type-specific lipooligosaccharides from Mycobacterium tuberculosis. Biochemistry 30:378–388. https://doi.org/10.1021/bi00216a011
Rombouts Y, Burguiere A, Maes E et al (2009) Mycobacterium marinum lipooligosaccharides are unique caryophyllose-containing cell wall glycolipids that inhibit tumor necrosis factor-alpha secretion in macrophages. J Biol Chem 284:20975–20988. https://doi.org/10.1074/jbc.M109.011429
van der Woude AD, Sarkar D, Bhatt A et al (2012) Unexpected link between lipooligosaccharide biosynthesis and surface protein release in Mycobacterium marinum. J Biol Chem 287:20417–20429. https://doi.org/10.1074/jbc.M111.336461
Kano H, Doi T, Fujita Y et al (2005) Serotype-specific modulation of human monocyte functions by glycopeptidolipid (GPL) isolated from Mycobacterium avium complex. Biol Pharm Bull 28:335–339. https://doi.org/10.1248/bpb.28.335
Horgen L, Barrow EL, Barrow WW et al (2000) Exposure of human peripheral blood mononuclear cells to total lipids and serovar-specific glycopeptidolipids from Mycobacterium avium serovars 4 and 8 results in inhibition of TH1-type responses. Microb Pathog 29:9–16. https://doi.org/10.1006/mpat.2000.0358
Recht J, Kolter R (2001) Glycopeptidolipid acetylation affects sliding motility and biofilm formation in Mycobacterium smegmatis. J Bacteriol 183:5718–5724. https://doi.org/10.1128/JB.183.19.5718-5724.2001
Chakraborty P, Kumar A (2019) The extracellular matrix of mycobacterial biofilms: could we shorten the treatment of mycobacterial infections? Microb Cell 6:105–122. https://doi.org/10.15698/mic2019.02.667
Carter G, Wu M, Drummond DC et al (2003) Characterization of biofilm formation by clinical isolates of Mycobacterium avium. J Med Microbiol 52:747–752. https://doi.org/10.1099/jmm.0.05224-0
Ojha A, Hatfull GF (2007) The role of iron in Mycobacterium smegmatis biofilm formation: the exochelin siderophore is essential in limiting iron conditions for biofilm formation but not for planktonic growth. Mol Microbiol 66:468–483. https://doi.org/10.1111/j.1365-2958.2007.05935.x
Hall-Stoodley L, Keevil CW, Lappin-Scott HM (1998) Mycobacterium fortuitum and Mycobacterium chelonae biofilm formation under high and low nutrient conditions. J Appl Microbiol 85(Suppl 1):60S–69S. https://doi.org/10.1111/j.1365-2672.1998.tb05284.x
Esteban J, Garcia-Coca M (2017) Mycobacterium biofilms. Front Microbiol 8:2651. https://doi.org/10.3389/fmicb.2017.02651
Williams MM, Yakrus MA, Arduino MJ et al (2009) Structural analysis of biofilm formation by rapidly and slowly growing nontuberculous mycobacteria. Appl Environ Microbiol 75:2091–2098. https://doi.org/10.1128/AEM.00166-09
Adjemian J, Olivier KN, Seitz AE et al (2012) Spatial clusters of nontuberculous mycobacterial lung disease in the United States. Am J Respir Crit Care Med 186:553–558. https://doi.org/10.1164/rccm.201205-0913OC
Ricketts WM, O'Shaughnessy TC, van Ingen J (2014) Human-to-human transmission of Mycobacterium kansasii or victims of a shared source? Eur Respir J 44:1085–1087. https://doi.org/10.1183/09031936.00066614
Bryant JM, Grogono DM, Greaves D et al (2013) Whole-genome sequencing to identify transmission of Mycobacterium abscessus between patients with cystic fibrosis: a retrospective cohort study. Lancet 381:1551–1560. https://doi.org/10.1016/S0140-6736(13)60632-7
Honda JR, Bernhard JN, Chan ED (2015) Natural disasters and nontuberculous mycobacteria: a recipe for increased disease? Chest 147:304–308. https://doi.org/10.1378/chest.14-0974
Liu JJ, Zhao K, Khan S, et al. (2014) Multi-scale population and mobility estimation with geo-tagged tweets. 1st International Workshop on Big Data Analytics for Biosecurity (BioBAD2015) arXiv:1412.0327
Nishiuchi Y, Iwamoto T, Maruyama F (2017) Infection sources of a common non-tuberculous mycobacterial pathogen, Mycobacterium avium complex. Front Med (Lausanne) 4:27. https://doi.org/10.3389/fmed.2017.00027
Haworth CS, Banks J, Capstick T et al (2017) British Thoracic Society guidelines for the management of non-tuberculous mycobacterial pulmonary disease (NTM-PD). Thorax 72:ii1–ii64. https://doi.org/10.1136/thoraxjnl-2017-210927
Wei T, Dong WJ, Yan QX et al (2016) Developed and developing world contributions to climate system change based on carbon dioxide, methane and nitrous oxide emissions. Adv Atmos Sci 33:632–643
Rickman OB, Ryu JH, Fidler ME et al (2002) Hypersensitivity pneumonitis associated with Mycobacterium avium complex and hot tub use. Mayo Clin Proc 77:1233–1237. https://doi.org/10.4065/77.11.1233
Pelletier PA, du Moulin GC, Stottmeier KD (1988) Mycobacteria in public water supplies: comparative resistance to chlorine. Microbiol Sci 5:147–148
Hilborn ED, Covert TC, Yakrus MA et al (2006) Persistence of nontuberculous mycobacteria in a drinking water system after addition of filtration treatment. Appl Environ Microbiol 72:5864–5869. https://doi.org/10.1128/AEM.00759-06
Covert TC, Rodgers MR, Reyes AL et al (1999) Occurrence of nontuberculous mycobacteria in environmental samples. Appl Environ Microbiol 65:2492–2496
Halstrom S, Price P, Thomson R (2015) Review: Environmental mycobacteria as a cause of human infection. Int J Mycobacteriol 4:81–91. https://doi.org/10.1016/j.ijmyco.2015.03.002
Glazer CS, Martyny JW, Lee B et al (2007) Nontuberculous mycobacteria in aerosol droplets and bulk water samples from therapy pools and hot tubs. J Occup Environ Hyg 4:831–840. https://doi.org/10.1080/15459620701634403
Thomson R, Tolson C, Carter R et al (2013) Isolation of nontuberculous mycobacteria (NTM) from household water and shower aerosols in patients with pulmonary disease caused by NTM. J Clin Microbiol 51:3006–3011. https://doi.org/10.1128/JCM.00899-13
Fujita K, Ito Y, Hirai T et al (2013) Genetic relatedness of Mycobacterium avium-intracellulare complex isolates from patients with pulmonary MAC disease and their residential soils. Clin Microbiol Infect 19:537–541. https://doi.org/10.1111/j.1469-0691.2012.03929.x
Lahiri A, Kneisel J, Kloster I et al (2014) Abundance of Mycobacterium avium ssp. hominissuis in soil and dust in Germany - implications for the infection route. Lett Appl Microbiol 59:65–70. https://doi.org/10.1111/lam.12243
Leski TA, Malanoski AP, Gregory MJ et al (2011) Application of a broad-range resequencing array for detection of pathogens in desert dust samples from Kuwait and Iraq. Appl Environ Microbiol 77:4285–4292. https://doi.org/10.1128/AEM.00021-11
Favet J, Lapanje A, Giongo A et al (2013) Microbial hitchhikers on intercontinental dust: catching a lift in Chad. ISME J 7:850–867. https://doi.org/10.1038/ismej.2012.152
Weil T, De Filippo C, Albanese D et al (2017) Legal immigrants: invasion of alien microbial communities during winter occurring desert dust storms. Microbiome 5:32. https://doi.org/10.1186/s40168-017-0249-7
Grebennikova TV, Syroeshkin AV, Shubralova EV et al (2018) The DNA of Bacteria of the World Ocean and the Earth in Cosmic Dust at the International Space Station. ScientificWorldJournal 2018:7360147. https://doi.org/10.1155/2018/7360147
Leaning J, Guha-Sapir D (2013) Natural disasters, armed conflict, and public health. N Engl J Med 369:1836–1842. https://doi.org/10.1056/NEJMra1109877
Eldholm V, Pettersson JHO, Brynildsrud OB et al (2016) Armed conflict and population displacement as drivers of the evolution and dispersal of Mycobacterium tuberculosis. Proc Natl Acad Sci U S A 113:13881–13886. https://doi.org/10.1073/pnas.1611283113
Fjallbrant H, Akerstrom M, Svensson E et al (2013) Hot tub lung: an occupational hazard. Eur Respir Rev 22:88–90. https://doi.org/10.1183/09059180.00002312
Nishiuchi Y, Tamura A, Kitada S et al (2009) Mycobacterium avium complex organisms predominantly colonize in the bathtub inlets of patients' bathrooms. Jpn J Infect Dis 62:182–186
WHO (1999) Hazard prevention and control in the work environment: Airborne dust. World Health Organization, Geneva. Source: http://www.who.int/occupational_health/publications/airdust/en/
Fattorini L, Creti R, Nisini R et al (2002) Recombinant GroES in combination with CpG oligodeoxynucleotides protects mice against Mycobacterium avium infection. J Med Microbiol 51:1071–1079. https://doi.org/10.1099/0022-1317-51-12-1071
Le Moigne V, Rottman M, Goulard C et al (2015) Bacterial phospholipases C as vaccine candidate antigens against cystic fibrosis respiratory pathogens: the Mycobacterium abscessus model. Vaccine 33:2118–2124. https://doi.org/10.1016/j.vaccine.2015.03.030
Le Moigne V, Belon C, Goulard C et al (2016) MgtC as a host-induced factor and vaccine candidate against Mycobacterium abscessus infection. Infect Immun 84:2895–2903. https://doi.org/10.1128/IAI.00359-16
Watanabe M, Nakamura H, Nabekura R et al (2015) Protective effect of a dewaxed whole-cell vaccine against Mycobacterium ulcerans infection in mice. Vaccine 33:2232–2239. https://doi.org/10.1016/j.vaccine.2015.03.046
Hart BE, Hale LP, Lee S (2015) Recombinant BCG expressing Mycobacterium ulcerans Ag85A imparts enhanced protection against experimental buruli ulcer. PLoS Negl Trop Dis 9:e0004046. https://doi.org/10.1371/journal.pntd.0004046
Bolz M, Benard A, Dreyer AM et al (2016) Vaccination with the surface proteins MUL_2232 and MUL_3720 of Mycobacterium ulcerans induces antibodies but fails to provide protection against buruli ulcer. PLoS Negl Trop Dis 10:e0004431. https://doi.org/10.1371/journal.pntd.0004431
Dhand NK, Eppleston J, Whittington RJ et al (2016) Changes in prevalence of ovine paratuberculosis following vaccination with Gudair(R): Results of a longitudinal study conducted over a decade. Vaccine 34:5107–5113. https://doi.org/10.1016/j.vaccine.2016.08.064
Tirado Y, Puig A, Alvarez N et al (2016) Mycobacterium smegmatis proteoliposome induce protection in a murine progressive pulmonary tuberculosis model. Tuberculosis (Edinb) 101:44–48. https://doi.org/10.1016/j.tube.2016.07.017
Garcia Mde L, Borrero R, Lanio ME et al (2014) Protective effect of a lipid-based preparation from Mycobacterium smegmatis in a murine model of progressive pulmonary tuberculosis. Biomed Res Int 2014:273129. https://doi.org/10.1155/2014/273129
Rodriguez L, Tirado Y, Reyes F et al (2011) Proteoliposomes from Mycobacterium smegmatis induce immune cross-reactivity against Mycobacterium tuberculosis antigens in mice. Vaccine 29:6236–6241. https://doi.org/10.1016/j.vaccine.2011.06.077
Borrero R, Garcia Mde L, Canet L et al (2013) Evaluation of the humoral immune response and cross reactivity against Mycobacterium tuberculosis of mice immunized with liposomes containing glycolipids of Mycobacterium smegmatis. BMC Immunol 14(Suppl 1):S13. https://doi.org/10.1186/1471-2172-14-S1-S13
von Reyn CF, Lahey T, Arbeit RD et al (2017) Safety and immunogenicity of an inactivated whole cell tuberculosis vaccine booster in adults primed with BCG: a randomized, controlled trial of DAR-901. PLoS One 12:e0175215. https://doi.org/10.1371/journal.pone.0175215
Lahey T, Laddy D, Hill K et al (2016) Immunogenicity and protective efficacy of the DAR-901 booster vaccine in a murine model of tuberculosis. PLoS One 11:e0168521. https://doi.org/10.1371/journal.pone.0168521
Valdés I, Mederos L, Echemendía M, et al. (2014) "Mycobacterium habana" as a live vaccine candidate against TB. The Art & Science of Tuberculosis Vaccine Development 2nd Edition. Source: http://tbvaccines.usm.my/finlay/?q=content/chapter-58-mycobacterium-habana-live-vaccine-candidate-against-tb
Cardona P, Marzo-Escartin E, Tapia G et al (2015) Oral administration of heat-killed Mycobacterium manresensis delays progression toward active tuberculosis in C3HeB/FeJ mice. Front Microbiol 6:1482. https://doi.org/10.3389/fmicb.2015.01482
Montane E, Barriocanal AM, Arellano AL et al (2017) Pilot, double-blind, randomized, placebo-controlled clinical trial of the supplement food Nyaditum resae(R) in adults with or without latent TB infection: Safety and immunogenicity. PLoS One 12:e0171294. https://doi.org/10.1371/journal.pone.0171294
Tukvadze N, Cardona P, Vashakidze S et al (2016) Development of the food supplement Nyaditum resae as a new tool to reduce the risk of tuberculosis development. Int J Mycobacteriol 5(Suppl 1):S101–S102. https://doi.org/10.1016/j.ijmyco.2016.09.073
Abbot NC, Beck JS, Feval F et al (2002) Immunotherapy with Mycobacterium vaccae and peripheral blood flow in long-treated leprosy patients, a randomised, placebo-controlled trial. Eur J Vasc Endovasc Surg 24:202–208
Stanford J, Stanford C, Grange J (2004) Immunotherapy with Mycobacterium vaccae in the treatment of tuberculosis. Front Biosci 9:1701–1719
Saqib M, Khatri R, Singh B et al (2016) Mycobacterium indicus pranii as a booster vaccine enhances BCG induced immunity and confers higher protection in animal models of tuberculosis. Tuberculosis (Edinb) 101:164–173. https://doi.org/10.1016/j.tube.2016.10.002
Das S, Chowdhury BP, Goswami A et al (2016) Mycobacterium indicus pranii (MIP) mediated host protective intracellular mechanisms against tuberculosis infection: involvement of TLR-4 mediated signaling. Tuberculosis (Edinb) 101:201–209. https://doi.org/10.1016/j.tube.2016.09.027
Singh B, Saqib M, Gupta A et al (2017) Autophagy induction by Mycobacterium indicus pranii promotes Mycobacterium tuberculosis clearance from RAW 264.7 macrophages. PLoS One 12:e0189606. https://doi.org/10.1371/journal.pone.0189606
Sharma SK, Katoch K, Sarin R et al (2017) Efficacy and safety of Mycobacterium indicus pranii as an adjunct therapy in category II pulmonary tuberculosis in a randomized trial. Sci Rep 7:3354. https://doi.org/10.1038/s41598-017-03514-1
Pai M, Nicol MP, Boehme CC (2016) Tuberculosis diagnostics: state of the art and future directions. Microbiol Spectr 4. doi:https://doi.org/10.1128/microbiolspec.TBTB2-0019-2016
Nathavitharana RR, Hillemann D, Schumacher SG et al (2016) multicenter noninferiority evaluation of Hain GenoType MTBDRplus version 2 and Nipro NTM+MDRTB line probe assays for detection of rifampin and isoniazid resistance. J Clin Microbiol 54:1624–1630. https://doi.org/10.1128/JCM.00251-16
Makinen J, Marjamaki M, Marttila H et al (2006) Evaluation of a novel strip test, GenoType Mycobacterium CM/AS, for species identification of mycobacterial cultures. Clin Microbiol Infect 12:481–483. https://doi.org/10.1111/j.1469-0691.2006.01380.x
Singh AK, Maurya AK, Umrao J et al (2013) Role of GenoType(R) Mycobacterium common mycobacteria/additional species assay for rapid differentiation between Mycobacterium tuberculosis complex and different species of non-tuberculous mycobacteria. J Lab Physicians 5:83–89. https://doi.org/10.4103/0974-2727.119847
Gitti Z, Neonakis I, Fanti G et al (2006) Use of the GenoType Mycobacterium CM and AS assays to analyze 76 nontuberculous mycobacterial isolates from Greece. J Clin Microbiol 44:2244–2246. https://doi.org/10.1128/JCM.02088-05
Richter E, Rusch-Gerdes S, Hillemann D (2006) Evaluation of the genotype mycobacterium assay for identification of mycobacterial species from cultures. J Clin Microbiol 44:1769–1775. https://doi.org/10.1128/JCM.44.5.1769-1775.2006
Huh HJ, Park KS, Jang MA et al (2014) Evaluation of performance of the Real-Q NTM-ID kit for rapid identification of eight nontuberculous mycobacterial species. J Clin Microbiol 52:4053–4055. https://doi.org/10.1128/JCM.01957-14
Tortoli E, Mariottini A, Mazzarelli G (2003) Evaluation of INNO-LiPA MYCOBACTERIA v2: improved reverse hybridization multiple DNA probe assay for mycobacterial identification. J Clin Microbiol 41:4418–4420
Mougari F, Loiseau J, Veziris N et al (2017) Evaluation of the new GenoType NTM-DR kit for the molecular detection of antimicrobial resistance in non-tuberculous mycobacteria. J Antimicrob Chemother 72:1669–1677. https://doi.org/10.1093/jac/dkx021
Lee MR, Cheng A, Huang YT et al (2012) Performance assessment of the DR. TBDR/NTM IVD kit for direct detection of Mycobacterium tuberculosis isolates, including rifampin-resistant isolates, and nontuberculous Mycobacteria. J Clin Microbiol 50:3398–3401. https://doi.org/10.1128/JCM.01862-12
Liu J, Yue J, Yan Z et al (2012) Performance assessment of the CapitalBio mycobacterium identification array system for identification of mycobacteria. J Clin Microbiol 50:76–80. https://doi.org/10.1128/JCM.00320-11
Wang HY, Kim H, Kim S et al (2015) Evaluation of PCR-reverse blot hybridization assay for the differentiation and identification of Mycobacterium species in liquid cultures. J Appl Microbiol 118:142–151. https://doi.org/10.1111/jam.12670
Jenkins PA, Campbell IA, Banks J et al (2008) Clarithromycin vs ciprofloxacin as adjuncts to rifampicin and ethambutol in treating opportunist mycobacterial lung diseases and an assessment of Mycobacterium vaccae immunotherapy. Thorax 63:627–634. https://doi.org/10.1136/thx.2007.087999
Bax HI, Freeman AF, Ding L et al (2013) Interferon alpha treatment of patients with impaired interferon gamma signaling. J Clin Immunol 33:991–1001. https://doi.org/10.1007/s10875-013-9882-5
Milanes-Virelles MT, Garcia-Garcia I, Santos-Herrera Y et al (2008) Adjuvant interferon gamma in patients with pulmonary atypical Mycobacteriosis: a randomized, double-blind, placebo-controlled study. BMC Infect Dis 8:17. https://doi.org/10.1186/1471-2334-8-17
Achkar JM, Chan J, Casadevall A (2015) B cells and antibodies in the defense against Mycobacterium tuberculosis infection. Immunol Rev 264:167–181. https://doi.org/10.1111/imr.12276
Glatman-Freedman A, Casadevall A (1998) Serum therapy for tuberculosis revisited: reappraisal of the role of antibody-mediated immunity against Mycobacterium tuberculosis. Clin Microbiol Rev 11:514–532
Acosta A, Norazmi MN, Sarmiento ME (2010) Antibody mediated immunity - a missed opportunity in the fight against tuberculosis? Malays J Med Sci 17:66–67
Acosta A, Yamile L, Norazmi MN et al (2012) Towards a new challenge in TB control: Development of antibody-based protection. Understanding tuberculosis-analyzing the origin of Mycobacterium tuberculosis pathogenicity. IntechOpen. https://doi.org/10.5772/31835
Acosta A, Yamile L, Norazmi MN et al (2013) The role of antibodies in the defense against tuberculosis. Tuberculosis-Current Issues in Diagnosis and Management. IntechOpen. https://doi.org/10.5772/53950
Chen T, Blanc C, Eder AZ et al (2016) Association of human antibodies to arabinomannan with enhanced mycobacterial opsonophagocytosis and intracellular growth reduction. J Infect Dis 214:300–310. https://doi.org/10.1093/infdis/jiw141
Li H, Wang X-x, Wang B et al (2017) Latently and uninfected healthcare workers exposed to TB make protective antibodies against Mycobacterium tuberculosis. Proc Natl Acad Sci U S A 114:5023–5028. https://doi.org/10.1073/pnas.1611776114
Chin ST, Ignatius J, Suraiya S et al (2015) Comparative study of IgA VH 3 gene usage in healthy TST(−) and TST(+) population exposed to tuberculosis: deep sequencing analysis. Immunology 144:302–311. https://doi.org/10.1111/imm.12372
Balu S, Reljic R, Lewis MJ et al (2011) A novel human IgA monoclonal antibody protects against tuberculosis. J Immunol 186:3113–3119. https://doi.org/10.4049/jimmunol.1003189
Hamasur B, Haile M, Pawlowski A et al (2004) A mycobacterial lipoarabinomannan specific monoclonal antibody and its F(ab') fragment prolong survival of mice infected with Mycobacterium tuberculosis. Clin Exp Immunol 138:30–38. https://doi.org/10.1111/j.1365-2249.2004.02593.x
Teitelbaum R, Glatman-Freedman A, Chen B et al (1998) A mAb recognizing a surface antigen of Mycobacterium tuberculosis enhances host survival. Proc Natl Acad Sci U S A 95:15688–15693. https://doi.org/10.1073/pnas.95.26.15688
Lopez Y, Yero D, Falero-Diaz G et al (2009) Induction of a protective response with an IgA monoclonal antibody against Mycobacterium tuberculosis 16kDa protein in a model of progressive pulmonary infection. Int J Med Microbiol 299:447–452. https://doi.org/10.1016/j.ijmm.2008.10.007
Alvarez N, Otero O, Camacho F et al (2013) Passive administration of purified secretory IgA from human colostrum induces protection against Mycobacterium tuberculosis in a murine model of progressive pulmonary infection. BMC Immunol 14(Suppl 1):S3. https://doi.org/10.1186/1471-2172-14-S1-S3
Olivares N, Puig A, Aguilar D et al (2009) Prophylactic effect of administration of human gamma globulins in a mouse model of tuberculosis. Tuberculosis (Edinb) 89:218–220. https://doi.org/10.1016/j.tube.2009.02.003
Abate G, Hamzabegovic F, Eickhoff CS et al (2019) BCG vaccination induces M. avium and M. abscessus cross-protective immunity. Front Immunol 10:234. https://doi.org/10.3389/fimmu.2019.00234
Chung SW, Choi SH, Kim TS (2004) Induction of persistent in vivo resistance to Mycobacterium avium infection in BALB/c mice injected with interleukin-18-secreting fibroblasts. Vaccine 22:398–406. https://doi.org/10.1016/j.vaccine.2003.07.002
Kim SH, Cho D, Kim TS (2001) Induction of in vivo resistance to Mycobacterium avium infection by intramuscular injection with DNA encoding interleukin-18. Immunology 102:234–241. https://doi.org/10.1046/j.1365-2567.2001.01150.x
Buccheri S, Reljic R, Caccamo N et al (2009) Prevention of the post-chemotherapy relapse of tuberculous infection by combined immunotherapy. Tuberculosis (Edinb) 89:91–94. https://doi.org/10.1016/j.tube.2008.09.001
Reljic R, Clark SO, Williams A et al (2006) Intranasal IFN-gamma extends passive IgA antibody protection of mice against Mycobacterium tuberculosis lung infection. Clin Exp Immunol 143:467–473. https://doi.org/10.1111/j.1365-2249.2006.03012.x
Alvarez N, Borrero R, García MÁ et al (2009) Obtención y caracterización parcial de un extracto lipídico de la membrana externa de Mycobacterium smegmatis. Vaccimonitor 18:15–19
Maoz C, Shitrit D, Samra Z et al (2008) Pulmonary Mycobacterium simiae infection: comparison with pulmonary tuberculosis. Eur J Clin Microbiol Infect Dis 27:945–950. https://doi.org/10.1007/s10096-008-0522-6
Wu ML, Aziz DB, Dartois V et al (2018) NTM drug discovery: status, gaps and the way forward. Drug Discov Today 23:1502–1519. https://doi.org/10.1016/j.drudis.2018.04.001
Falkinham JO 3rd (2018) Challenges of NTM drug development. Front Microbiol 9:1613. https://doi.org/10.3389/fmicb.2018.01613
Jarlier V, Nikaido H (1994) Mycobacterial cell wall: structure and role in natural resistance to antibiotics. FEMS Microbiol Lett 123:11–18. https://doi.org/10.1111/j.1574-6968.1994.tb07194.x
van Ingen J, Boeree MJ, van Soolingen D et al (2012) Resistance mechanisms and drug susceptibility testing of nontuberculous mycobacteria. Drug Resist Updat 15:149–161. https://doi.org/10.1016/j.drup.2012.04.001
Machado D, Cannalire R, Santos Costa S et al (2015) Boosting effect of 2-phenylquinoline efflux inhibitors in combination with macrolides against Mycobacterium smegmatis and Mycobacterium avium. ACS Infect Dis 1:593–603. https://doi.org/10.1021/acsinfecdis.5b00052
Early J, Fischer K, Bermudez LE (2011) Mycobacterium avium uses apoptotic macrophages as tools for spreading. Microb Pathog 50:132–139. https://doi.org/10.1016/j.micpath.2010.12.004
Mougari F, Bouziane F, Crockett F et al (2017) Selection of resistance to clarithromycin in Mycobacterium abscessus subspecies. Antimicrob Agents Chemother 61. https://doi.org/10.1128/AAC.00943-16
Kim SW, Subhadra B, Whang J et al (2017) Clinical Mycobacterium abscessus strain inhibits autophagy flux and promotes its growth in murine macrophages. Pathog Dis 75. https://doi.org/10.1093/femspd/ftx107
Sapriel G, Konjek J, Orgeur M et al (2016) Genome-wide mosaicism within Mycobacterium abscessus: evolutionary and epidemiological implications. BMC Genomics 17:118. https://doi.org/10.1186/s12864-016-2448-1
Wu ML, Gengenbacher M, Dick T (2016) Mild nutrient starvation triggers the development of a small-cell survival morphotype in mycobacteria. Front Microbiol 7:947. https://doi.org/10.3389/fmicb.2016.00947
Aguilar-Ayala DA, Cnockaert M, Andre E et al (2017) In vitro activity of bedaquiline against rapidly growing nontuberculous mycobacteria. J Med Microbiol 66:1140–1143. https://doi.org/10.1099/jmm.0.000537
Dick T, Lee BH, Murugasu-Oei B (1998) Oxygen depletion induced dormancy in Mycobacterium smegmatis. FEMS Microbiol Lett 163:159–164. https://doi.org/10.1111/j.1574-6968.1998.tb13040.x
Tamura A, Hebisawa A, Kusaka K et al (2016) Relationship between lung cancer and Mycobacterium avium complex isolated using bronchoscopy. Open Respir Med J 10:20–28. https://doi.org/10.2174/1874306401610010020
Fujita Y, Ishii S, Hirano S et al (2011) A case of lung cancer complicated with active non-tuberculous mycobacterium (NTM) infection successfully treated with anti-cancer agents and anti-NTM agents. Nihon Kokyuki Gakkai Zasshi 49:855–860
Huang CT, Tsai YJ, Wu HD et al (2012) Impact of non-tuberculous mycobacteria on pulmonary function decline in chronic obstructive pulmonary disease. Int J Tuberc Lung Dis 16:539–545. https://doi.org/10.5588/ijtld.11.0412
Donatelli C, Mehta AC (2015) Lady Windermere syndrome: mycobacterium of sophistication. Cleve Clin J Med 82:641–643. https://doi.org/10.3949/ccjm.82a.14100
(1997) Diagnosis and treatment of disease caused by nontuberculous mycobacteria. This official statement of the American Thoracic Society was approved by the Board of Directors, March 1997. Medical Section of the American Lung Association. Am J Respir Crit Care Med 156: S1–25. doi:https://doi.org/10.1164/ajrccm.156.2.atsstatement
Griffith DE, Aksamit T, Brown-Elliott BA et al (2007) An official ATS/IDSA statement: diagnosis, treatment, and prevention of nontuberculous mycobacterial diseases. Am J Respir Crit Care Med 175:367–416. https://doi.org/10.1164/rccm.200604-571ST
Faria S, Joao I, Jordao L (2015) General overview on nontuberculous mycobacteria, biofilms, and human infection. J Pathog 2015:809014. https://doi.org/10.1155/2015/809014
Coll F, Preston M, Guerra-Assuncao JA et al (2014) PolyTB: A genomic variation map for Mycobacterium tuberculosis. Tuberculosis 94:346–354. https://doi.org/10.1016/j.tube.2014.02.005
Honda JR, Virdi R, Chan ED (2018) Global environmental nontuberculous mycobacteria and their contemporaneous man-made and natural niches. Front Microbiol 9:2029–2029. https://doi.org/10.3389/fmicb.2018.02029
Ovrutsky AR, Chan ED, Kartalija M et al (2013) Cooccurrence of free-living amoebae and nontuberculous Mycobacteria in hospital water networks, and preferential growth of Mycobacterium avium in Acanthamoeba lenticulata. Appl Environ Microbiol 79:3185–3192. https://doi.org/10.1128/AEM.03823-12
CDC (2009) A guide to drinking water treatment and sanitation for backcountry & travel use. Centers for disease control and prevention. Source: https://www.cdc.gov/healthywater/drinking/travel/backcountry_water_treatment.html
Inkinen J, Jayaprakash B, Santo Domingo JW et al (2016) Diversity of ribosomal 16S DNA- and RNA-based bacterial community in an office building drinking water system. J Appl Microbiol 120:1723–1738. https://doi.org/10.1111/jam.13144
Feazel LM, Baumgartner LK, Peterson KL et al (2009) Opportunistic pathogens enriched in showerhead biofilms. Proc Natl Acad Sci U S A 106:16393–16399. https://doi.org/10.1073/pnas.0908446106
NJH (2017) Is your shower head making you sick? National Jewish Health, Denver, CO. Source: https://www.nationaljewish.org/health-insights/health-infographics/is-your-shower-head-making-you-sick-ntm-nontuberculous-mycobacteria
Deutsch L, Bravo V (2015) How to save water on this drought-ridden Earth Day. USA Today. Source: https://www.usatoday.com/story/news/nation-now/2015/04/22/earth-day-water-conservation-tips/25957005/
Zimmermann P, Finn A, Curtis N (2018) Does BCG vaccination protect against nontuberculous mycobacterial infection? A systematic review and meta-analysis. J Infect Dis 218:679–687. https://doi.org/10.1093/infdis/jiy207
Valadas E (2004) Nontuberculous mycobacteria: clinical importance and relevance to bacille Calmette-Guerin vaccination. Clin Infect Dis 39:457–458. https://doi.org/10.1086/422326
Ryu YJ, Koh WJ, Daley CL (2016) Diagnosis and treatment of nontuberculous mycobacterial lung disease: clinicians’ perspectives. Tuberc Respir Dis (Seoul) 79:74–84. https://doi.org/10.4046/trd.2016.79.2.74
Denkinger CM, Kik SV, Pai M (2013) Robust, reliable and resilient: designing molecular tuberculosis tests for microscopy centers in developing countries. Expert Rev Mol Diagn 13:763–767. https://doi.org/10.1586/14737159.2013.850034
Gommans EP, Even P, Linssen CF et al (2015) Risk factors for mortality in patients with pulmonary infections with non-tuberculous mycobacteria: a retrospective cohort study. Respir Med 109:137–145. https://doi.org/10.1016/j.rmed.2014.10.013
Yuan MK, Lai YC, Chang CY et al (2017) Chest X-ray finding of pulmonary tuberculosis and nontuberculous mycobacterial lung diseases in patients with acid-fast bacilli smear-positive sputum. Austin Tuberculosis: Research & Treatment 2:1–5
Philley JV, DeGroote MA, Honda JR et al (2016) Treatment of non-tuberculous mycobacterial lung disease. Curr Treat Options Infect Dis 8:275–296. https://doi.org/10.1007/s40506-016-0086-4
Genc GE, Demir M, Yaman G et al (2018) Evaluation of MALDI-TOF MS for identification of nontuberculous mycobacteria isolated from clinical specimens in mycobacteria growth indicator tube medium. New Microbiol 41:214–219
Dymova M, Alkhovik O, Evdokimova L et al (2016) Whole genome-sequencing of non-tuberculous mycobacteria. Eur Respir J 48. https://doi.org/10.1183/13993003.congress-2016.PA891
Li G, Lian LL, Wan L et al (2013) Antimicrobial susceptibility of standard strains of nontuberculous mycobacteria by microplate Alamar Blue assay. PLoS One 8:e84065. https://doi.org/10.1371/journal.pone.0084065
Wang X, Li H, Jiang G et al (2014) Prevalence and drug resistance of nontuberculous mycobacteria, northern China, 2008–2011. Emerg Infect Dis 20:1252–1253. https://doi.org/10.3201/eid2007.131801
Candido PH, Nunes Lde S, Marques EA et al (2014) Multidrug-resistant nontuberculous mycobacteria isolated from cystic fibrosis patients. J Clin Microbiol 52:2990–2997. https://doi.org/10.1128/JCM.00549-14
Tortoli E, Kohl TA, Brown-Elliott BA et al (2016) Emended description of Mycobacterium abscessus, Mycobacterium abscessus subsp. abscessus and Mycobacteriumabscessus subsp. bolletii and designation of Mycobacterium abscessus subsp. massiliense comb. nov. Int J Syst Evol Microbiol 66:4471–4479. https://doi.org/10.1099/ijsem.0.001376
Pasipanodya JG, Ogbonna D, Ferro BE et al (2017) Systematic review and meta-analyses of the effect of chemotherapy on pulmonary Mycobacterium abscessus outcomes and disease recurrence. Antimicrob Agents Chemother 61. https://doi.org/10.1128/AAC.01206-17
Griffith DE (2014) Mycobacterium abscessus subsp abscessus lung disease: 'trouble ahead, trouble behind...'. F1000Prime Rep 6:107. https://doi.org/10.12703/P6-107
Nessar R, Cambau E, Reyrat JM et al (2012) Mycobacterium abscessus: a new antibiotic nightmare. J Antimicrob Chemother 67:810–818. https://doi.org/10.1093/jac/dkr578
Kwon YS, Koh WJ (2016) Diagnosis and treatment of nontuberculous mycobacterial lung disease. J Korean Med Sci 31:649–659. https://doi.org/10.3346/jkms.2016.31.5.649
Duarte RS, Lourenco MC, Fonseca Lde S et al (2009) Epidemic of postsurgical infections caused by Mycobacterium massiliense. J Clin Microbiol 47:2149–2155. https://doi.org/10.1128/JCM.00027-09
Nunes Lde S, Baethgen LF, Ribeiro MO et al (2014) Outbreaks due to Mycobacterium abscessus subsp. bolletii in southern Brazil: persistence of a single clone from 2007 to 2011. J Med Microbiol 63:1288–1293. https://doi.org/10.1099/jmm.0.074906-0
Dedrick RM, Guerrero-Bustamante CA, Garlena RA et al (2019) Engineered bacteriophages for treatment of a patient with a disseminated drug-resistant Mycobacterium abscessus. Nat Med 25:730–733. https://doi.org/10.1038/s41591-019-0437-z
Millar BC, Moore JE (2019) Antimycobacterial strategies to evade antimicrobial resistance in the nontuberculous mycobacteria. Int J Mycobacteriol 8:7–21. https://doi.org/10.4103/ijmy.ijmy_153_18
Ortiz-Perez A, Martin-de-Hijas N, Alonso-Rodriguez N et al (2011) Importance of antibiotic penetration in the antimicrobial resistance of biofilm formed by non-pigmented rapidly growing mycobacteria against amikacin, ciprofloxacin and clarithromycin. Enferm Infecc Microbiol Clin 29:79–84. https://doi.org/10.1016/j.eimc.2010.08.016
Munoz-Egea MC, Garcia-Pedrazuela M, Mahillo-Fernandez I et al (2016) Effect of antibiotics and antibiofilm agents in the ultrastructure and development of biofilms developed by nonpigmented rapidly growing mycobacteria. Microb Drug Resist 22:1–6. https://doi.org/10.1089/mdr.2015.0124
Rho J, Ahn C, Gao A et al (2018) Disparities in mortality of hispanic patients with cystic fibrosis in the United States. A national and regional cohort study. Am J Respir Crit Care Med 198:1055–1063. https://doi.org/10.1164/rccm.201711-2357OC
Morimoto K, Yoshiyama T, Okumura M et al (2012) Multidrug-resistant tuberculosis with a history of nontuberculous Mycobacteriosis: a brief report of two cases. Intern Med 51:3435–3437
Mirsaeidi M, Farshidpour M, Allen MB et al (2014) Highlight on advances in nontuberculous mycobacterial disease in North America. Biomed Res Int 2014:919474. https://doi.org/10.1155/2014/919474
Hong JY, Jang SH, Kim SY et al (2016) Elevated serum CA 19–9 levels in patients with pulmonary nontuberculous mycobacterial disease. Braz J Infect Dis 20:26–32. https://doi.org/10.1016/j.bjid.2015.09.005
Funding
This study received funding from the LRGS Grant (203.PPSK.67212002) from the Department of Higher Education, Ministry of Education, Malaysia and UMS Research Grant Scheme (SLB0172-2018) from Universiti Malaysia Sabah.
Author information
Authors and Affiliations
Corresponding authors
Ethics declarations
Conflict of interest
The authors declare that they have no conflicts of interest.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Chin, K.L., Sarmiento, M.E., Alvarez-Cabrera, N. et al. Pulmonary non-tuberculous mycobacterial infections: current state and future management. Eur J Clin Microbiol Infect Dis 39, 799–826 (2020). https://doi.org/10.1007/s10096-019-03771-0
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10096-019-03771-0