
Abstract
A species classification regarding Old World monkey adenoviruses is proposed. We determined the nucleotide sequences of PCR-amplified fragments from the genes of the IVa2, DNA-dependent DNA polymerase, penton base, and hexon proteins from every simian adenovirus (SAdV) serotype that originated from Old World monkeys for which the full genome sequence had not yet been published. We confirmed that the majority of Old Word monkey SAdVs belong to two previously established species. Interestingly, one is the most recently established human AdV species, Human mastadenovirus G, which includes a single human virus, HAdV-52, as well as SAdV-1, -2, -7, -11, -12, and -15. The other approved species, Simian mastadenovirus A includes SAdV-3, -4, -6, -9, -10, -14, and -48. Several SAdVs (SAdV-5, -8, -49, -50) together with baboon AdV-1 and rhesus monkey AdV strains A1139, A1163, A1173, A1258, A1285, A1296, A1312, A1327 and A1335 have been proposed to be classified into an additional species, Simian mastadenovirus B. Another proposed species, Simian mastadenovirus C has been described for SAdV-19, baboon AdV-2/4 and -3. Our study revealed the existence of four additional AdV lineages. The corresponding new candidate species are Simian mastadenovirus D (for SAdV-13), Simian mastadenovirus E (for SAdV-16), Simian mastadenovirus F (for SAdV-17 and -18), and Simian mastadenovirus G (for SAdV-20). Several biological and genomic properties, such as the host origin, haemagglutination profile, number of fibre genes, and G+C content of the genome, strongly support this classification. Three SAdV strains originating from the American Type Culture Collection turned out to be mixtures of at least two virus types, either of the same species (SAdV-12 and -15 types from Human mastadenovirus G) or of two different species (SAdV-5 types from Simian mastadenovirus B and Human mastadenovirus G).
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Simian adenoviruses (SAdVs) are members of the genus Mastadenovirus in the family Adenoviridae [16]. The first description of an SAdV in the literature was of a chimpanzee AdV [39], today known as SAdV-21 within the species Human mastadenovirus B. The first monkey AdVs, together with many other, mainly enteric, simian viruses from divergent families were discovered while testing poliomyelitis vaccines on kidney cell cultures made from macaques of two species [18]. Additional monkey AdV serotypes, characterised by the lack of cross-neutralisation [1], were found by screening other macaques and monkeys from two additional species: grivet and baboon [12, 19, 20, 24, 31]. Later on, when investigating chimpanzees suffering from kuru, experiments resulted in the discovery of four novel ape AdVs [38].
The first classification of monkey AdVs was based on their ability to haemagglutinate erythrocytes of different host origin, as a tool of taxon demarcation [37]. With the use of this biological assay, 16 monkey AdV strains were divided into four haemagglutination groups (HAG I–IV; Table 1). By analysing the nt sequences of the left genome ends, researchers have inferred that SAdV-16 (originally named SA7; Table 1) shares a similar organisation with HAdV-12, a member of the species Human mastadenovirus A [25]. The similarity of chimpanzee AdVs to HAdV-4 strains of the species Human mastadenovirus E was been recognised quite early [29].
A comparative sequence analysis of 25 distinct SAdV serotypes, SAdV-1 to 20, isolated from Old World monkeys (OWMs), and SAdV-21 to 25 from chimpanzees was performed by PCR amplification and sequencing of the virus-associated (VA) RNA genes [23]. All chimpanzee AdVs were proved to have two (tandem) VA RNA genes in their genomes. However, in monkey AdVs, only one VA RNA gene was detected (or none, because of the PCR imperfections). These data were applied in making the first phylogenetic tree of SAdVs [23]. The short VA RNA sequences and the first partial hexon sequences, obtained from chimpanzee AdVs in our lab, prompted us to place SAdV-21 into the species Human mastadenovirus B, and SAdV-22 to 25 into species Human mastadenovirus E [1]. These assumptions regarding the taxonomic place of the chimpanzee AdVs were confirmed later by phylogenetic analysis of other longer sequences [2, 10, 36, 40]. The first complete chimpanzee AdV genome was that of SAdV-25 [10]. It was followed by the genomes of the other four chimpanzee AdV types, SAdV-21 to 24 [40], and those of two additional chimpanzee AdVs (under the strain names of ChAd3 and ChAd6) [8, 35].
Ape AdVs have been isolated not only from chimpanzees but also from bonobos and gorillas [41]. These viruses have been proposed to be members of species Human mastadenovirus B, C and E, respectively, as they are definitely part of those genetic lineages [41]. Partial genome analysis of gorilla AdVs confirmed the theory on the mixed host origin of members of the species Human mastadenovirus B [47]. Almost all viruses in the species Human mastadenovirus D are from human sources. However, a recent study suggested that some chimpanzee AdVs might also belong to the species Human mastadenovirus D [48]. The same study also described a gorilla AdV belonging to the species Human mastadenovirus F, which previously included only human AdV serotypes (HAdV-40 and HAdV-41) [48]. The genome of a chimpanzee AdV isolate clustering in the species Human mastadenovirus A has been fully sequenced [50]. Development of vector vaccines from chimpanzee AdVs was started more than 10 years ago [49], and it has been the subject of growing interest [5]. Consequently, there is a growing number of different chimpanzee AdV isolates that have been studied. Colloca and co-workers [8] have screened more than a thousand faecal samples from chimpanzees and bonobos and isolated AdVs from around 50 % of them. The full sequences of some of these viruses indicated that they are closest to members of the species Human mastadenovirus B, C or E [8].
SAdVs have been found to be associated with several diseases of primates, including diarrhoea, pneumoenteritis, conjunctivitis, and hepatitis [4, 24, 45, 51], and some of them have been reported to induce tumours when injected into neonate rodents [21]. Short sequences from various genes of monkey AdVs have often been reported from colonies of captive macaques that were either suffering from diarrhoea [46] or not showing any clinical signs ascribed to AdVs [30, 48]. The study of monkey AdVs is much more undeveloped than that of the ape AdVs. The first full monkey AdV genome published was that of SAdV-3 (isolated from a rhesus macaque) [27]. It was proposed to be the first member of a new species, Simian mastadenovirus A. This species was approved by the International Committee on Taxonomy of Viruses and is still the only species that includes OWM AdVs exclusively [17]. The next sequenced OWM AdV genome was that of SAdV-1 (isolated from a crab-eating macaque). This virus was classified as belonging to the species Human mastadenovirus G [28], together with a HAdV type, HAdV-52 [22]. Further full-genome sequences were published from SAdV-48, -49 and -50 [41], as well as some partial sequences [4, 33]. As the interest in OWM AdVs as potential gene delivery tools increased, additional SAdV genomes were fully sequenced. These included SAdV-7 [42], followed by SAdV-6, -18 and -20 [43]. Phylogenetic analysis of AdVs that were newly isolated from rhesus macaques [43] indicated that they belonged to a common lineage with SAdV-49 and -50, which were sequenced previously [41]. For the classification of these SAdVs, the establishment of the species Simian mastadenovirus B has been proposed, whereas SAdV-48 has been described to belong to the species Simian mastadenovirus A [41, 43]. Four novel AdV strains that were found in olive baboons have been sequenced recently. One strain, baboon AdV-1 (BaAdV-1), has been proposed to be a member of the candidate species Simian mastadenovirus B, while BaAdV-2, -3 and -4 have been found to form a separate clade, representing the proposed new species Simian mastadenovirus C [7]. Recent publications have reported the discovery of AdVs in New World monkeys as well [6, 14, 15, 48].
Nowadays, the recognized diversity of SAdVs is approaching that of the HAdVs. However, while ape AdVs are well characterized and fully classified, most monkey AdVs still await classification, and only very short sequences or no sequences at all are published from their genomes. Our aim was to obtain at least partial sequences from multiple genes of every known OWM SAdV in order to explore their phylogenetic relationships and to find out how many lineages, corresponding to putative species, can be defined among them.
Materials and methods
Viruses
The prototype strains of OWM SAdV serotypes (SAdV-1 to 20), deposited in the American Type Culture Collection (ATCC), were studied initially by PCR and compared with other SAdV sequences from other laboratories (Table 1). Purified DNA from concentrated virions or cell culture supernatants were used.
PCR and DNA sequencing
Shorter or longer fragments were obtained by PCR from the genes of four well-conserved adenoviral proteins of 14 SAdVs, the full genome sequence of which has not been published previously (Table 1). The primer sequences and the estimated sizes of the expected PCR products are presented in Table 2. Fragments from the DNA-dependent DNA polymerase (pol) and the hexon genes were obtained by PCR methods published by others [26, 47]. The IVa2 gene fragment was amplified with consensus degenerate nested primers, designed in-house on the basis of highly conserved amino acid (aa) motifs taken from an alignment containing different mastadenovirus sequences. Similarly, consensus nested primers targeting the gene for the penton base were also based on an aa alignment, but containing proteins from SAdVs only (Table 2; A. Doszpoly, personal communication). To acquire the genome fragments between the PCR products, obtained from the two adjacent genes (IVa2 and pol), degenerate primers (designated as simian universal; “suniv”) were designed from nt sequences of SAdVs exclusively. For a primer-walking approach, several additional consensus suniv primers were prepared (Table 3). The PCRs were performed in a 50-μl volume with the following ingredients (final concentration): 3 mM MgCl2, 0.2 mM dNTP, 1 μM each primer, GoTaq Buffer, and 1.5 unit of GoTaq DNA polymerase enzyme (Promega Corp.). If applicable, SAdV-24 was used as a positive control. The PCR programs consisted of an initial denaturation step at 94 °C for 5 min followed by 45 cycles (94 °C, 30 s; 46 °C, 60 s; 72 °C, 60 s) and a final elongation step at 72 °C for 3 min. The program of PCRs with suniv primers was modified to fewer cycles (35), annealing at 52 °C for 30 s and elongation at 72 °C for 90 s, with a final elongation of 7 min. The size, quality and amount of PCR products were checked by loading 10 μl of the completed reaction mixtures on agarose gels. Amplified fragments were purified using a Nucleospin Extract II Kit (Macherey-Nagel) and sequenced directly on both strands using a Big Dye Terminator v3.1 Cycle Sequencing Kit (Life Technologies Inc.). Capillary electrophoresis was performed by a commercial service on a 3500 Series Genetic Analyzer (Life Technologies). If heterogeneous nt sequences were obtained, the PCR products were molecularly cloned using a CloneJET™ PCR Cloning Kit (Thermo Scientific). Chemically competent DH5α E. coli cells were transformed with the ligated vector by heat shock (90 s, 42 °C), and plasmids were purified by the alkaline lysis method. The full genome sequence was determined from the prototype strains of six SAdVs by the classical Sanger method and/or next-generation sequencing (NSG), as shown in Table 1.
Phylogenetic analysis
The identities of the sequences were checked using the BLASTX program at the website of the NCBI. Primate AdV sequence alignments were prepared with the Clustal program of the MEGA6 package [44]. For phylogeny inference, we used only partial gene sequences that were available from every studied virus. Phylogenetic calculations, based on alignments of aa sequences from IVa2 and pol, were performed using the ProtDist and PhyML algorithms provided at the Mobyle portal of the Pasteur Institute (Paris), and the ProtTest program [9]. The ProtDist analyses were run with the JTT substitution model, followed by Fitch-Margoliash analysis, applying the global rearrangements option. PhyML calculations [13] were based on a user tree obtained using ProtDist and a model determined using ProtTest (JTT with the “invariable sites” and “gamma distribution” options). Analysis of the nt sequences was performed by using PhyML on the TOPALi v2 platform [34] with the TrNef model for the penton base and the TIMef model for the hexon gene, both with invariable sites and gamma distribution options (proposed by the Model Selection module of TOPALi). Bootstrap analysis with 100 sampling replicates was applied for every tree. The trees were then visualized using the MEGA6 program [44]. Tree shrew AdV-1 (TSAdV-1) served as an outgroup.
Results
PCR and DNA sequencing
The IVa2 nested PCR yielded visible amplicons after the first round from only a few samples. Therefore, the product (253 bp without the primers) of the second round was used for additional primer design. On the other hand, the pol PCR, which was described as a nested system [47], gave sufficient products already in the first round, so we did not have to perform the second round. After removing the primer sequences, the size of this partial pol gene fragment was 952 bp, corresponding to nucleotide (nt) positions 5269 to 6220 in the SAdV-1 genome in the GenBank database (AY771780). PCRs with suniv primers (connecting the partial IVa2 and pol genes) resulted in products of 2154 to 2226 bp. After in silico translation, alignments of 287 aa from the N-terminus of the IVa2 protein and of 502 aa from the C-terminal part of pol were prepared. The length of the PCR products from the penton base gene varied between 319 and 331 bp. From the hexon gene, 253 bp of useful sequence was obtained after editing out the primers. In addition to the PCR-amplified gene fragments, the sequence of the full genome was determined for six strains. As presented in Table 1, the newly obtained sequences were submitted to GenBank and assigned the accession numbers KP853109 to KP853135 (short sequences) and KP329561 to KP329565 (full genomes).
Direct sequencing of the PCR products obtained from the penton base gene showed that three strains representing SAdV-5, -12 and -15 were not pure viruses but mixtures. Molecular cloning of the amplicons resulted in the separation of two different sequences from each mixture. The sequences from samples of SAdV-12 and SAdV-15 indicated the presence of different viruses in each “strain”, yet all of the four viruses appeared to be members of the species Human mastadenovirus G (Fig. 1c). The prototype strain of SAdV-5 was also found to contain two AdVs, but only one of them clustered with members of Human mastadenovirus G, whereas the other clustered with the members of the candidate species Simian mastadenovirus B (Fig. 1c).

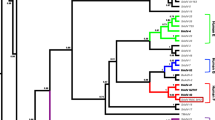
Phylogeny reconstruction based on partial aa sequences from (a) the polypeptide IVa2 and (b) the DNA-dependent DNA polymerase (pol) and (c) the partial nucleotide sequence of the penton base gene. Simian AdVs are shown by their type number only, followed by the abbreviation of the host: rh, rhesus macaque; cr, crab-eating macaque; yb, yellow baboon; gr, grivet. A black arrow indicates the hypothetical time point from which all of the fully sequenced AdVs (except SAdV-18) had two fibre genes. On the pol tree, the G+C content is shown in brackets next to the species names. Other abbreviations: HAdV, human AdV; BaAdV, baboon AdV; TSAdV, tree shrew AdV; TMAdV, titi monkey AdV
Phylogenetic analysis
In phylogeny reconstructions, the OWM SAdVs appeared in groups representing distinct lineages that were usually well separated from those of the human and ape AdVs (Figs. 1 and 2). Considering the tree topologies as well as other features of the examined viruses, the existence of at least four distinct lineages, obviously meriting the species-level demarcation, was revealed in addition to the already established or proposed HAdV and SAdV species. When preparing the hexon-based tree (Fig. 2), the corresponding sequence from several additional novel AdVs obtained from rhesus monkeys [4, 11, 33, 48] could also be included. The species Human mastadenovirus G and Simian mastadenovirus A appeared as two clear lineages on all four trees with a great number of virus types. Two OWM ATCC strains (SAdV-5 and -8) together with the previously described lineage including SAdV-49, -50 [41], nine AdVs isolated from rhesus macaques [43], and one AdV isolated from olive baboon (BaAdV-1) [7] appeared on the trees as a monophyletic clade corresponding to the recently proposed species Simian mastadenovirus B. SAdV-13 alone seemed to represent an independent lineage, candidate species Simian mastadenovirus D. A sister clade was formed by the closest virus, a recently reported novel AdV strain (23336) from rhesus macaque [32]. The distance between these two viruses does not necessarily warrant the establishment of an additional species. Similarly, SAdV-20 alone also formed an independent branch most closely diverging from the clade of the species Simian mastadenovirus A. In this case, the establishment of a novel species, Simian mastadenovirus G, seemed to be justified. SAdV-16 always appeared closest to the clade of the Simian mastadenovirus B, but as a long distinct branch on every tree, justifying the proposed species Simian mastadenovirus E. SAdV-17 and -18 were sister clades closest to members of the species Human mastadenovirus F, but always well separated from them. SAdV-19 was on the branch together with other baboon AdVs of the recently proposed species Simian mastadenovirus C [7].

Phylogenetic tree based on partial nucleotide sequences from the hexon gene. Simian AdVs are shown by their type number only, followed by the abbreviation of the host: rh, rhesus macaque; cr, crab-eating macaque; yb, yellow baboon; gr, grivet. AdVs found in rhesus macaques [4] for which only the short hexon sequence is available in GenBank are designated as “rhxx” (xx = strain number). Some types on the hexon tree are hidden as follows: rh+CHN (e): rh20, 25, 30, 34, 39, 43 and 48, CHN-39, -43, and -48; rh+CHN(f): rh2, 8, 9, 24 and 40, CHN-8; rh+CHN (g): rh51, and 59-60, CHN-30 and -51; rh+CHN (h): rh23, CHN-23 and -24. Other abbreviations: HAdV, human AdV; BaAdV, baboon AdV; TSAdV, tree shrew AdV; TMAdV, titi monkey AdV; Cynom1, cynomolgus monkey AdV-1
Discussion
We set out to complete the species allocation of OWM SAdVs. To this end, we determined full or partial DNA sequences from 14 prototype SAdVs originating from OWMs and used phylogeny inference for grouping. Subsequently, we tested the validity of every clade, i.e., putative species, using other genomic or biological characteristics. In the great majority of the cases, the results of the different comparisons were in good agreement, reinforcing the validity of the groups. The topology of the phylogenetic trees, constructed based on different genome fragments, showed some variations in the order of the branches. However, the number and content of the clusters in each analysis were comparable. The separation of the individual clades was generally supported by maximal or high bootstrap values, depending on the length and conservation status of the examined gene or protein fragment. On the trees, based on the IVa2 (Fig. 1a) and pol (Fig. 1b) aa sequences, the range of these probability values at the nodes of the proposed species was 84−100 and 89−100, respectively. The most reliable pol tree seems to show correctly also the “time” of the presumed acquisition of a second fibre gene (shown by a black arrow on Fig. 1b). From that hypothetical time point, all of the OWM AdVs and even HAdV members of the species Human mastadenovirus F and Human mastadenovirus G had two fibre genes. The only exception is SAdV-18, which probably lost one of the genes during a presumed host switch and adaptation to grivet. Furthermore, we speculate that the members of three HAdV species (Human mastadenovirus A, F and G) most probably originated from OWM AdVs.
Although the full genome sequence was available for many of the OWM AdVs, we used partial sequences to allow the inclusion of all prototype strains. PCR products were obtained from four different locations of the genome. The four genes were also different in terms of the timing of their expression during the viral life cycle. The pol gene is important because its sequence divergence is a significant criterion in the species demarcation of adenoviruses [16]. Because the amplified region of the penton base gene is highly variable, it proved to be suitable for detecting different genomic variants and even virus types in prototype SAdV strains that had earlier been supposed to be clean isolates. Molecular cloning of amplified penton base gene fragments proved that each of three ATCC strains was actually a mixture of two slightly (SAdV-12 and -15) or considerably (SAdV-5) different viruses. Consequently, the validity of their original serological comparison with other prototype strains could be questioned.
The IVa2 nested PCR was found to detect mastadenoviruses very effectively and was useful for designing additional consensus primers. The PCR, targeting the short hexon gene fragment that codes for the highly conserved N-terminal part of the capsid protein, is a widely used general AdV detection method [4, 33, 48], and therefore, additional SAdVs could be included in the hexon tree (Fig. 2). In spite of the rather limited amount of sequence data from these viruses, many of them could be safely assigned to several previously established or proposed species (Human mastadenovirus G, Simian mastadenovirus A, Simian mastadenovirus B). However, a few of them (red colobus 3 [48], rh15 and rh50 [4]) seem to stand alone, forecasting the discovery of additional genetic lineages of OWM AdVs for which further species might be needed in the future. The need to establish more novel monkey AdV species emerges also for the fully sequenced titi monkey AdV (a New World species) [6] and isolate 23336 from rhesus monkey [32].
The base composition of the genomic DNA is an additional species demarcation criterion for AdVs [16]. The G+C percentage of the partial or complete genome sequences seemed to be characteristic for the given virus groups and species. Ideally, these values should be deduced from complete genome sequences. Nonetheless, the trends seemed to be clear, even when based on partial sequences. Members of the species Simian mastadenovirus B and F have G+C-rich DNA (60.1-62.9 %). The species Simian mastadenovirus A, and E and Human mastadenovirus G include viruses with moderately high (54.4-57.9 %) G+C content, whereas the SAdVs classified into the species Simian mastadenovirus C, D, and G and Human mastadenovirus F have medium G+C content (47.8-52.6 %). Strain 23336 seems to have the lowest G+C proportion (46.7 %), as shown in Table 4.
The species with the largest number of members are Simian mastadenovirus A and B and Human mastadenovirus G [22, 27]. Recently published surveys have indicated that the viruses of species Human mastadenovirus G and Simian mastadenovirus A, are highly prevalent in macaques [4, 30]. The recently proposed species Simian mastadenovirus B includes SAdV-5, -8, -49, -50, nine other rhesus macaque isolates [41, 43], and BaAdV-1 [7]. The new finding is that SAdV-5 and -8 belong to this species, and these are the first viruses of this species that had been serotyped by virus neutralisation [37]. Previous publications have also supported this clade based on partial hexon sequences [4, 11, 33, 48].
SAdV-19 proved to be a novel member of the previously proposed species Simian mastadenovirus C [7]. This was supported by all of our trees, as well as by the G+C content and the uniqueness and uniformity of the host, the baboon.
SAdV-13 turned out to be the only representative of a separate lineage that we propose as a novel species, Simian mastadenovirus D. The exact host species (some Macaca sp.) of this AdV type is not known, unfortunately. All phylogenetic trees implied that SAdV-13 diverged from the other monkey AdVs at an early time. The G+C content of this virus is also different from that of all the other viruses (Table 4). The phylogenetically closest AdV is the rhesus monkey isolate 23336, but both the phylogenetic distance and the G+C content (46.7 vs 49.9 %) differentiate them.
SAdV-16 is closely related to members of the proposed species Simian mastadenovirus B. However, its phylogenetic distance seems to be large enough to merit the creation of a separate species. The different host species, the grivet (Chlorocebus aethiops), also supports the establishment of a new taxon for SAdV-16. However, as host switching is believed to be a relatively common phenomenon among primate AdVs [48], it cannot be excluded that this virus has a macaque origin. On the other hand, the two other grivet AdVs (SAdV-17 and -18) are phylogenetically closely related to each other but rather distantly to SAdV-16. The G+C content of the full genome of SAdV-16 is 57.9 %, which is notably different from those of the members of Simian mastadenovirus B (~62 %). Based on all data available at the moment, we propose to classify SAdV-16 as the first member of a novel species named Simian mastadenovirus E.
SAdV-17 and -18 compose the sister taxon to the species Human mastadenovirus F (Fig. 1 and 2). The phylogenetic distance and host species difference (grivet versus human) seem to be sufficient to propose a new species, Simian mastadenovirus F. The genome organisation of SAdV-18 [43] also shows very important differences compared to that of HAdV-40 and -41, the two known members of Human mastadenovirus F. SAdV-18 has one fibre gene only, while HAdV-40 and -41 have two. The Human mastadenovirus F members are unique among the primate AdVs, as the RGD motif is not present in their penton-base protein. They also lack the 12.5K gene in their E3 region, while SAdV-18 has this gene. The G+C content of HAdV-40 and -41 is 51 %, while SAdV-18 has a G+C-rich genome (61.4 %). We think that the proposal to establish a new species (Simian mastadenovirus F) for SAdV-17 and -18 is adequate.
The phylogenetic trees showed SAdV-20 to be a sister group to Simian mastadenovirus A, but it was sufficiently distant to represent a valid separate species (Simian mastadenovirus G). There was only one exception, the hexon-based tree (Fig. 2), where SAdV-20 is not separated clearly from members of Simian mastadenovirus A. Since the hexon gene is prone to be involved in homologous recombination events, it is possible that recombination occurred in this case as well. The different host species and the considerably divergent G+C content (47.8 %) of the SAdV-20 genome from that of Simian mastadenovirus A members (54.4-55.8 % in the full genome length) confirm our proposal for the new species Simian mastadenovirus G.
The penton base PCR was the best method in this study to detect different variants in three ATCC virus strains. Our conclusion is that SAdV-12 and -15 are mixtures of two types of the species Human mastadenovirus G (Fig. 1c), whilst SAdV-5 seems to be a mixture of a member of Human mastadenovirus G and a putative Simian mastadenovirus B member. Evidence of non-homogeneity was observed in cross-neutralisation experiments conducted previously [20]. SAdV-5 (a mixture of Human mastadenovirus G and Simian mastadenovirus B members) showed one-way cross-neutralisation, and SAdV-12 (seemingly a mixture of two Human mastadenovirus G members) had two-way cross-neutralisation with different putative Human mastadenovirus G types. This early experiment did not study SAdV-15, so our statement on the mixed type is based on the described penton base sequences and also on a shotgun sequencing [27] attempt that revealed the heterogeneity of SAdV-15 (unpublished).
Most of our proposals are in good agreement with the original classification of monkey AdVs according to their HAG properties [37]. For example, every member of HAG group II belongs to the species Simian mastadenovirus A (Table 4). Similarly, the uniqueness of SAdV-13 and SAdV-16 was confirmed by the fact that each of these viruses is the sole member of HAG group I and IV, respectively. SAdV-5 and -8, members of the group HAG III, belong to the species Simian mastadenovirus B. Interestingly, several other members of group HAG III clustered to the species Human mastadenovirus G. Nonetheless, this biological property remains a useful demarcation criterion [16].
The relationship between the tissue tropism of the SAdVs and their phylogenetic clustering is not as obvious as in the case of some HAdVs. For example, members of the species Human mastadenovirus F, which are typical enteric AdVs, are generally found in human stool and are common in wastewater, while some Human mastadenovirus D members are notorious for infecting the cornea and causing epidemic keratoconjunctivitis [3]. An early study has reported that SAdVs can cause epidemic conjunctivitis in macaques [45]. It is notable that in our analyses, all SAdVs implicated in this disease clustered with members of the species Simian mastadenovirus A.
The host range of some newly proposed species, as well as that of the previously approved Simian mastadenovirus A and Human mastadenovirus G is mixed (Table 4). AdVs infecting individuals of different simian genera usually also belong to separate AdV species. However, there are AdV species that include AdVs of several different monkey species. In contrast, in some cases, AdVs found in individuals of the same monkey species may belong to different viral species. It is a general feature of SAdVs and HAdVs to have mixed host origins. Thus human, chimpanzee, gorilla and bonobo AdVs have been proposed as members of Human mastadenovirus B and Human mastadenovirus C [41]. However, crossing the host barrier rarely occurs, and usually only among evolutionarily close primate species. It has been recognized most often between apes and humans, or among OWMs.
Based on phylogenetic comparisons and the biological properties of OWM AdVs, we confirmed the need to establish two previously proposed species Simian mastadenovirus B and Simian mastadenovirus C. Furthermore, we propose the establishment of four new SAdV species, Simian mastadenovirus D, E, F and G. At present, all of these newly proposed species would contain exclusively OWM AdVs. A future task would be to find and study AdVs that occur in representatives of other monkey species, especially in New World monkeys and prosimians. Such investigations would be essential for getting better insights into the evolution and host characteristics of the wider or complete spectrum of primate AdVs. We assume that the presently proposed SAdV species will be justified by the discovery and sequence and phylogenetic analyses of a large number of further monkey AdVs in the near future.
References
Benkő M, Harrach B, Russell WC (2000) Family Adenoviridae. In: Van Regenmortel MHV, Fauquet CM, Bishop DHL, Carstens EB, Estes MK, Lemon SM, Maniloff J, Mayo MA, McGeoch DJ, Pringle CR, Wickner RB (eds) Virus taxonomy, classification and nomenclature of viruses: seventh report of the international committee on taxonomy of viruses. Academic Press, San Diego, pp 227–238
Benkő M, Harrach B (2003) Molecular evolution of adenoviruses. Curr Top Microbiol Immunol 272:3–35
Benkő M (2008) Adenoviruses: Pathogenesis. In: Mahy BWJ, Van Regenmortel MHV (eds) Encyclopedia of virology, 3rd edn. Elsevier, Oxford, pp 24–29
Bányai K, Esona MD, Liu A, Wang Y, Tu X, Jiang B (2010) Molecular detection of novel adenoviruses in fecal specimens of captive monkeys with diarrhea in China. Vet Microbiol 142:416–419
Capone S, D’Alise AM, Ammendola V, Colloca S, Cortese R, Nicosia A, Folgori A (2013) Development of chimpanzee adenoviruses as vaccine vectors: challenges and successes emerging from clinical trials. Expert Rev Vaccines 12:379–393
Chen EC, Yagi S, Kelly KR, Mendoza SP, Tarara RP, Canfield DR, Maninger N, Rosenthal A, Spinner A, Bales KL, Schnurr DP, Lerche NW, Chiu CY (2011) Cross-species transmission of a novel adenovirus associated with a fulminant pneumonia outbreak in a new world monkey colony. PLoS Pathog 7:e1002155
Chiu CY, Yagi S, Lu X, Yu G, Chen EC, Liu M, Dick EJ, Carey KD, Erdman DD, Leland MM, Patterson JL (2013) A novel adenovirus species associated with an acute respiratory outbreak in a baboon colony and evidence of coincident human infection. MBio 4:e00084
Colloca S, Barnes E, Folgori A, Ammendola V, Capone S, Cirillo A, Siani L, Naddeo M, Grazioli F, Esposito ML, Ambrosio M, Sparacino A, Bartiromo M, Meola A, Smith K, Kurioka A, O’Hara GA, Ewer KJ, Anagnostou N, Bliss C, Hill AV, Traboni C, Klenerman P, Cortese R, Nicosia A (2012) Vaccine vectors derived from a large collection of simian adenoviruses induce potent cellular immunity across multiple species. Sci Transl Med 4:115ra112
Darriba D, Taboada GL, Doallo R, Posada D (2011) ProtTest 3: fast selection of best-fit models of protein evolution. Bioinformatics 27:1164–1165
Farina SF, Gao GP, Xiang ZQ, Rux JJ, Burnett RM, Alvira MR, Marsh J, Ertl HC, Wilson JM (2001) Replication-defective vector based on a chimpanzee adenovirus. J Virol 75:11603–11613
Foytich KR, Deshazer G, Esona MD, Liu A, Wang Y, Tu X, Jiang B (2014) Identification of new provisional simian adenovirus species from captive monkeys, China. Emerg Infect Dis 20:1758–1759
Fuentes-Marins R, Rodriguez AR, Kalter SS, Hellman A, Crandell RA (1963) Isolation of enteroviruses from the normal baboon (Papio doguera). J Bacteriol 85:1045–1050
Guindon S, Gascuel O (2003) A simple, fast, and accurate algorithm to estimate large phylogenies by maximum likelihood. Syst Biol 52:696–704
Gál J, Hornyák Á, Mándoki M, Bakonyi T, Balka G, Szeredi L, Marosán M, Ludányi T, Forgách P, Sós E, Demeter Z, Farkas SL (2013) Novel mastadenovirus infection and clinical disease in a pygmy marmoset (Callithrix [Cebuella] pygmaea). Vet Microbiol 167:695–699
Hall NH, Archer LL, Childress AL, Wellehan JF (2012) Identification of a novel adenovirus in a cotton-top tamarin (Saguinus oedipus). J Vet Diagn Invest 24:359–363
Harrach B, Benkő M, Both GW, Brown M, Davison AJ, Echavarria M, Hess M, Jones MS, Kajon A, Lehmkuhl HD, Mautner V, Mittal SK, Wadell G (2011) Family Adenoviridae. In: King AMQ, Adams MJ, Lefkowitz E, Carstens EB (eds) Virus taxonomy, ninth report of the international committee on taxonomy of viruses. Elsevier, Oxford, pp 125–141
Harrach B (2014) Adenoviruses: general features. In: Reference module in biomedical sciences. Elsevier, Amsterdam. doi:10.1016/B978-0-12-801238-3.02523-X
Hull RN, Minner JR, Smith JW (1956) New viral agents recovered from tissue cultures of monkey kidney cells. I. Origin and properties of cytopathogenic agents S.V.1, S.V.2, S.V.4, S.V.5, S.V.6, S.V.11, S.V.12 and S.V.15. Am J Hyg 63:204–215
Hull RN, Minner JR (1957) New viral agents recovered from tissue cultures of monkey kidney cells. II. Problems of isolation and identification. Ann N Y Acad Sci 67:413–423
Hull RN, Minner JR, Mascoli CC (1958) New viral agents recovered from tissue cultures of monkey kidney cells. III. Recovery of additional agents both from cultures of monkey tissues and directly from tissues and excreta. Am J Hyg 68:31–44
Hull RN, Johnson IS, Culbertson CG, Reimer CB, Wright HF (1965) Oncogenicity of the simian adenoviruses. Science 150:1044–1046
Jones MS, Harrach B, Ganac RD, Gozum MM, Dela Cruz WP, Riedel B, Pan C, Delwart EL, Schnurr DP (2007) New adenovirus species found in a patient presenting with gastroenteritis. J Virol 81:5978–5984
Kidd AH, Garwicz D, Oberg M (1995) Human and simian adenoviruses: phylogenetic inferences from analysis of VA RNA genes. Virology 207:32–45
Kim CS, Sueltenfuss ES, Kalter SS (1967) Isolation and characterization of simian adenoviruses isolated in association with an outbreak of pneumoenteritis in vervet monkeys (Cercopithecus aethiops). J Infect Dis 117:292–300
Kimelman D, Miller JS, Porter D, Roberts BE (1985) E1a regions of the human adenoviruses and of the highly oncogenic simian adenovirus 7 are closely related. J Virol 53:399–409
Kiss I, Matiz K, Allard A, Wadell G, Benkő M (1996) Detection of homologous DNA sequences in animal adenoviruses by polymerase chain reaction. Acta Vet Hung 44:243–251
Kovács GM, Davison AJ, Zakhartchouk AN, Harrach B (2004) Analysis of the first complete genome sequence of an Old World monkey adenovirus reveals a lineage distinct from the six human adenovirus species. J Gen Virol 85:2799–2807
Kovács GM, Harrach B, Zakhartchouk AN, Davison AJ (2005) Complete genome sequence of simian adenovirus 1: an Old World monkey adenovirus with two fiber genes. J Gen Virol 86:1681–1686
Li QG, Wadell G (1988) The degree of genetic variability among adenovirus type 4 strains isolated from man and chimpanzee. Arch Virol 101:65–77
Lu J, Wang Q, Wang H, Li G, Gao G (2011) Molecular characterization of adenoviruses in fecal samples of captively bred rhesus macaques in China. Vet Microbiol 149:461–466
Malherbe H, Harwin R (1963) The cytopathic effects of vervet monkey viruses. S Afr Med J 37:407–411
Malouli D, Howell GL, Legasse AW, Kahl C, Axthelm MK, Hansen SG, Früh K (2014) Full genome sequence analysis of a novel adenovirus of rhesus macaque origin indicates a new simian adenovirus type and species. Virol Rep 3–4:18–29
Maluquer de Motes C, Hundesa A, Almeida FC, Bofill-Mas S, Girones R (2011) Isolation of a novel monkey adenovirus reveals a new phylogenetic clade in the evolutionary history of simian adenoviruses. Virol J 8:125
Milne I, Lindner D, Bayer M, Husmeier D, McGuire G, Marshall DF, Wright F (2009) TOPALi v2: a rich graphical interface for evolutionary analyses of multiple alignments on HPC clusters and multi-core desktops. Bioinformatics 25:126–127
Peruzzi D, Dharmapuri S, Cirillo A, Bruni BE, Nicosia A, Cortese R, Colloca S, Ciliberto G, La Monica N, Aurisicchio L (2009) A novel chimpanzee serotype-based adenoviral vector as delivery tool for cancer vaccines. Vaccine 27:1293–1300
Purkayastha A, Ditty SE, Su J, McGraw J, Hadfield TL, Tibbetts C, Seto D (2005) Genomic and bioinformatics analysis of HAdV-4, a human adenovirus causing acute respiratory disease: implications for gene therapy and vaccine vector development. J Virol 79:2559–2572
Rapoza NP (1967) A classification of simian adenoviruses based on hemagglutination. Am J Epidemiol 86:736–745
Rogers NG, Basnight M, Gibbs CJ, Gajdusek DC (1967) Latent viruses in chimpanzees with experimental kuru. Nature 216:446–449
Rowe WP, Hartley JW, Huebner RJ (1956) Additional serotypes of the APC virus group. Proc Soc Exp Biol Med 91:260–262
Roy S, Gao G, Clawson DS, Vandenberghe LH, Farina SF, Wilson JM (2004) Complete nucleotide sequences and genome organization of four chimpanzee adenoviruses. Virology 324:361–372
Roy S, Vandenberghe LH, Kryazhimskiy S, Grant R, Calcedo R, Yuan X, Keough M, Sandhu A, Wang Q, Medina-Jaszek CA, Plotkin JB, Wilson JM (2009) Isolation and characterization of adenoviruses persistently shed from the gastrointestinal tract of non-human primates. PLoS Pathog 5:e1000503
Roy S, Clawson DS, Adam VS, Medina A, Wilson JM (2011) Construction of gene transfer vectors based on simian adenovirus 7. J Gen Virol 92:1749–1753
Roy S, Sandhu A, Medina A, Clawson DS, Wilson JM (2012) Adenoviruses in fecal samples from asymptomatic rhesus macaques, United States. Emerg Infect Dis 18:1081–1088
Tamura K, Stecher G, Peterson D, Filipski A, Kumar S (2013) MEGA6: molecular evolutionary genetics analysis version 6.0. Mol Biol Evol 30:2725–2729
Vasileva VA, Ivanov MT, Rumel NB, D’Yachenko AG, Kakubava VV, Danelyan GA (1978) Isolation and biological characterization of an adenovirus of rhesus macaques. Acta Biol Med Ger 37:1281–1287
Wang Y, Tu X, Humphrey C, McClure H, Jiang X, Qin C, Glass RI, Jiang B (2007) Detection of viral agents in fecal specimens of monkeys with diarrhea. J Med Primatol 36:101–107
Wevers D, Leendertz FH, Scuda N, Boesch C, Robbins MM, Head J, Ludwig C, Kühn J, Ehlers B (2010) A novel adenovirus of Western lowland gorillas (Gorilla gorilla gorilla). Virol J 7:303
Wevers D, Metzger S, Babweteera F, Bieberbach M, Boesch C, Cameron K, Couacy-Hymann E, Cranfield M, Gray M, Harris LA, Head J, Jeffery K, Knauf S, Lankester F, Leendertz SA, Lonsdorf E, Mugisha L, Nitsche A, Reed P, Robbins M, Travis DA, Zommers Z, Leendertz FH, Ehlers B (2011) Novel adenoviruses in wild primates: a high level of genetic diversity and evidence of zoonotic transmissions. J Virol 85:10774–10784
Xiang Z, Gao G, Reyes-Sandoval A, Cohen CJ, Li Y, Bergelson JM, Wilson JM, Ertl HC (2002) Novel, chimpanzee serotype 68-based adenoviral vaccine carrier for induction of antibodies to a transgene product. J Virol 76:2667–2675
Zhou C, Tian H, Wang X, Liu W, Yang S, Shen Q, Wang Y, Ni B, Chen S, Fu X, Fei R, Zhang W (2014) The genome sequence of a novel simian adenovirus in a chimpanzee reveals a close relationship to human adenoviruses. Arch Virol 159:1765–1770
Zöller M, Mätz-Rensing K, Kaup FJ (2008) Adenoviral hepatitis in a SIV-infected rhesus monkey (Macaca mulatta). J Med Primatol 37:184–187
Acknowledgments
We would like to thank Andor Doszpoly for kindly providing the PCR primers he designed for the penton base gene of SAdVs. Thanks are also due to Alistair Kidd and Alexander Zakhartchouk for generously sharing some samples with us. The research project was supported by the Hungarian Scientific Research Fund Grant OTKA NN107632 and EU Grant Advance FP7-290002.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Additional information
L. Pantó and I. I. Podgorski contributed equally to this work.
M. Jánoska, a gifted scientist and beloved colleague passed away during the research period of this paper.
Rights and permissions
About this article

Cite this article
Pantó, L., Podgorski, I.I., Jánoska, M. et al. Taxonomy proposal for Old World monkey adenoviruses: characterisation of several non-human, non-ape primate adenovirus lineages. Arch Virol 160, 3165–3177 (2015). https://doi.org/10.1007/s00705-015-2575-z
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00705-015-2575-z