
Abstract
The invasion of common ragweed in Slovakia was analysed based on 1635 collected records. More than 70 years have passed since the first record was sampled in 1949. Since then, it has spread in two waves, first in the lowlands, next to farmland and human settlements, and later in the mountains, on the roadsides. In the present, it is probably expanded in half of the territory of Slovakia. It is changing habitat preferences and adapting to new environments. Using eleven environmental and geographical variables, the species distribution modelling was performed to obtain probability of occurrence in high resolution (30 s) in Slovakia. Four predictions were calculated based on two models. The first prediction was based on the first model trained on ragweed records before the start of the second expansion wave in 2000. The second prediction is based on the first model and predicts the present distribution according to current climatic conditions, which have changed in twenty years. The third prediction was based on the second model, trained on all 1635 collected records from 1949 to 2022. The fourth prediction based on the second model shows the potential distribution in 2040–2061. Comparing the predictions, the climatic changes explain current expansion of common ragweed in mountains since the air temperature is the main factor shaping distribution of common ragweed in Slovakia.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
1 Introduction
Ambrosia artemisiifolia L. (common ragweed) of the Compositae family, native to the temperate regions of North America, is considered to be a dangerous invasive species and an unfavourable weed in agriculture, as well as a source of allergenic pollen (Lambdon et al. 2008; Smith et al. 2013). The introduction of the common ragweed into natural habitats in Europe began in the 19th century, however, its massive spread, mainly in lowland regions, started at the beginning of the 20th century (Rich 1994; Chauvel et al. 2006; Mang et al. 2018). The first records from Central and Eastern Europe are from 1873 in Poland (Tokarska-Guzik et al. 2011), 1883 in the Czech Republic and Austria (Pyšek et al. 2002; Essl et al. 2009), 1922 in Hungary (Csontos 2010), and 1942 in Ukraine (Song and Prots 1998). In Slovakia, the species has spread since 1949 after its introduction via contaminated Canadian crop delivery (Hrabovský et al. 2016).
Temperature is one of the most important ecological factors for common ragweed distribution as it affects seed germination and plant growth (Basset and Crompton 1975; Deen et al. 1998). The common ragweed germination process reflects the species adaptation to seasonal changes in the temperate zone (Dickerson 1968). It requires the regular daily alternating temperature of (5–) 10 to 20 (–28) °C (Li et al. 1989; Brandes and Nitzsche 2006) and seed stratification (Bazzaz 1970; Baskin and Baskin 1987; Brandes and Nitzsche 2006). Furthermore, germination is affected by other factors: photoperiod, seed burial depth, pH, water, and salt stress (Farooq et al. 2019). As an annual plant, common ragweed has a life cycle tied to one vegetation season, its life period from germination to seed production ranges from 115 days to 183 days (Li et al. 1989; Kazinczi et al. 2008). The species is not tolerant to freezing, spring or autumn frosts eliminate its growth (Essl et al. 2015). Therefore, the distribution of common ragweed is limited to the frost-free season, which correlated with its occurrence in North America up to 50° N in the first half of the 20th century, with the highest known locality at 975 m a.s.l. (Allard 1943). In European regions, the species distribution was limited mainly to very low altitudes with warm climates in the twentieth century (Mang et al. 2018).
The invasion of common ragweed into colder areas can be promoted by climate change, along with transportation and habitat disturbance (Cunze et al. 2013; Rasmussen et al. 2017; Skálová et al. 2017; Mang et al. 2018; Lemke et al. 2021; Liu et al. 2021). The territory of Slovakia extends into two European ecoregions: Pannonian mixed forests and Carpathian montane coniferous forests (Metzger et al. 2005). Until the 1990s, common ragweed occurred only in the region of the Pannonian flora, and its introduction into the area of the Carpathian flora was not expected (Jehlík 1998). However, the common ragweed expanded into mountain regions lastly (Štrba 2003; Zaliberová and Podroužková-Medvecká 2011; Hrabovský and Mičieta 2014, 2018; Eliáš 2021; Kochjarová et al. 2023).
In this study, we chronologically follow the process of the spread of common ragweed in Slovakia, changes in the means of spread, and ecological preferences. These, together with prediction models tied to changing climatic conditions, may answer the following questions: (a) Are there differences in common ragweed habitats in the past and the present? (b) Are climatic changes or ecological adaptation responsible for the expansion of ragweed into mountain regions? (c) What is the potential distribution of common ragweed in Slovakia? (d) How will the invasion proceed in the future?
2 Materials and methods
2.1 Study area
Slovakia, a European country with an area of 49,034 km², is characterized by different environmental and geographical features. It is located in the range of two climatic zones (Metzger et al. 2005). The Pannonian zone, with a warmer climate, overlaps more than one-third of the area. The mean annual air temperature of the Pannonian zone ranges from 10.8 °C (7.3 –12.2 °C) in the west to 10.5 °C (9.4–11 °C) in the east. The annual precipitation totals vary from 640 mm (534 –922 mm) to 695 mm (620 –868 mm). The colder Carpathian zone exhibits a mean annual air temperature of 7.8 °C (0.3 –10.7 °C), and annual precipitation totals 873 mm (569 –1621 mm). The given climate data was extracted from WorldClim datasets (Fick and Hijmans 2017).
The studied area was invaded by several alien plant taxa (Medvecká et al. 2012; Májeková et al. 2021), including common ragweed, a species that has unexpectedly started to spread from the warm Pannonian region to the Carpathians (Hrabovský et al. 2016; Kochjarová et al. 2023), which makes both the area and common ragweed a suitable model for the study of adaptation to changing climatic conditions.
2.2 Data collection
Recent records of ragweed in Slovakia were sampled from the entire country in the years 2017–2022. For large populations growing along roadsides, railways, or arable fields, coordinates were recorded from approximately the centre of the population. Older botanical records were collected from public and private herbaria, the State Nature Conservancy of the Slovak Republic (https://www.biomonitoring.sk), and other published sources (e.g., Jehlík 1998; Hrabovský et al. 2016; Hrabovský and Mičieta 2018; Eliáš 2022). The majority of the records gathered included habitat information. Maps and fieldwork were used to fill up the gaps left by missing data. In cases where the herbarium specimens lacked coordinates, the sites were visited, and the coordinates of the current populations were recorded. The collected point type records (Table S1) were extended by 17 additional geographic and environmental features (Table 1) using 30-second resolution raster layers in QGIS 3.22.3. The records were assigned to several types of habitats: arable fields, railways, roadsides, settlements, unpaved roads and paths, grasslands, salt marshes, river harbours and riverbanks, and sand dunes.
2.3 Data analysis
For the study of potential common ragweed habitat changes we used the cumulative numbers of records and occupied grids in different types of habitats (Skálová et al. 2017). The study area was divided into 1577 grid cells (1299 complete and 278 shared with neighbouring countries) according to modified Central European mapping with cell size 5’ × 3’ (Hrabovský 2023). An analysis of covariance (ANCOVA) with year as a covariate was performed in R (version 4. 3. 1, R Core Team 2023) to obtain regression slopes among cumulative frequency curves in the whole study area and separately in the lowlands and mountains.
Species distribution modelling (SDM) was realized at higher resolution (30-second) to obtain the potential distribution of common ragweed in Slovakia for the past period (1970–2000), for the current period (2018–2021), and for the future (2040–2061) if one of the worst scenarios (model EC-Earth3-Veg, Döscher et al. 2022; scenario SSP5-8.5, WorldClim datasets, Fick and Hijmans 2017) were to come true. 17 variables as possible predictors (Table 1) were tested for multicollinearity using ‘car’ and ‘mctest’ packages in R (Fox et al. 2007; Imdadullah et al. 2016). The six correlated predictors (Table 1) with a variance inflation factor higher than 10 were excluded from SDM so that the condition number was not higher than 30 (Dormann et al. 2013). Meteorological variables (mean annual air temperature and annual precipitation) were changeable predictors; remaining nine predictors were treated as static variables, and each SDM used the same values of them. Two presence datasets were established: the first dataset contains records from 1949 to 2000, and the second dataset contains records from 1949 to 2022. The pseudo-absence dataset should be randomly created at least 1–2° from places of occurrence (Barbet-Massin et al. 2012). However, this recommendation was not possible to follow in the small study area. Therefore, pseudo-absences were generated in ten random runs for each presence dataset (Barbet-Massin et al. 2012). SDMs were calculated by ‘sdm’ function of ‘sdm’ package (Naimi, Araujo 2016) in R using 19 profile, regression-based or machine learning models: BIOCLIM, DOMAIN, Mahalanobis distance (MAHAL), generalized linear model (GLM), polynomial generalized linear model (GLMPOLY), generalized additive models (GAM), multiple adaptive regression splines (MARS), maximum likelihood (MAXLIKE), boosted regression trees (BRT), random forest (RF), classification and regression trees (CART), support vector machines (SVM), mixture discriminant analysis (MDA), flexible discriminant analysis (FDA), radial basis functions (RBF), recursive partitioning and regression trees (RPART), multi-layer perceptron (MLP), lasso and elastic net regularized generalized linear models (GLMNET), maximum entropy (MAXENT). Each model was evaluated with 10-fold cross-validation to get the area under the receiver operating characteristic curve (AUC) values and the maximum of true skill statistic (maxTSS). The first model was trained on the first presence dataset, the first pseudo-absence datasets, and meteorological variables averaged for the past period (1970–2000). The second model was trained on the second presence dataset, the second pseudo-absence datasets, and meteorological variables averaged for the current period (2018–2021). The predictions were calculated using the ‘ensemble’ function of the same ‘sdm’ package with a weighted averaging by AUC values over all predictions from all fitted models with an AUC higher than 90% and a maxTSS higher than 70% (Table S2). Based on the first model, the predictions for periods 1970–2000 and 2018–2021 were obtained. Based on the second model, the predictions for periods 2018–2021 and 2040–2061 were made. This results in four predictions (Fig. 1): (1) common ragweed distribution if the climate remained the same as before 2000; (2) common ragweed distribution trained on records from 1949 to 2000 and predicted for current period meteorological variables; (3) current common ragweed distribution trained on all records from 1949 to 2022 and predicted for current period meteorological variables; (4) future common ragweed distribution if the SSP585 global warming scenario were to come true. Probability values from 1 to 0 were distinguished by the scale: abundant (> 0.51), common (0.26–0.50), occasional (0.11–0.25), and rare occurrence (< 0.10). The correlation between predictions 2 and 3 was evaluated using the ‘cor.test’ function, method Pearson, in R to compare the first model with the second model.

Workflow diagram illustrating the data analysis
A conditional inference tree was used as a non-parametric decision tree to estimate the efficiency rates of 17 environmental factors shaping the common ragweed distribution (Table 1). Meteorological variables of the current period and occurrence data from 1949 to 2022 were used.The tree was created with the ‘ctree’ function of the ‘party’ package in R (Hothorn et al. 2015).
All collected records were divided into three groups: Pannonian populations, Carpathian populations, and transient populations at the contact zone between lowlands and mountains (up to three kilometres from the Pannonicum-Carpaticum border). Linear discriminant analysis (LDA) was performed using the ‘MASS’ package in R (Ripley et al. 2013) to find the main predictors that can explain separation between the three groups based on their ecological preferences. The LDA is a multivariate statistical method that is widely used for morphotype classification (Hastie et al. 2001), however, it is sometimes also used for ecotype classification (Sanchez and Mackenzie 2016). We split the data into test and training sets and employed 10-fold cross-validation to achieve the accuracy of the LDA classification.
3 Results
3.1 Records and habitats of common ragweed in Slovakia
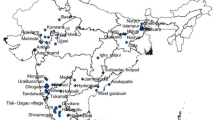
From 1949 to 2022, we collected 1635 records of common ragweed in Slovakia in 696 of 1577 grid cells (44.1% of the whole territory, Fig. 2). In Pannonian lowlands, it presents 1296 records in 527 of all 699 Pannonian grid cells (75.4% of Pannonian lowlands). In the Carpathian Mountains, 339 records in 200 of all 1066 Carpathian grid cells occur only in 18.8% of the region. From mountains, 28% of the records are located within five kilometres from lowlands, 52% of the records are no more than 20 km away, and 72% of the records are at a distance of up to 40 km from the Pannonian basin. In the period 1949–2000, 336 records were known from the lowlands and only 14 from the mountains. While the number of records in the lowlands increased not even three times in the last 22 years (2001–2022), in the mountains it increased 23 times at the same time.

Grid distribution map of common ragweed in Slovakia. A point symbolizes at least one record in a grid cell of size 5’ × 3’. The green surface represents the phytogeographical mountain area of Carpaticum
Up to 2000, human settlements and arable fields were the most frequently colonized habitats in the lowlands (60.4% of records), while in the mountains common ragweed occurred mainly along railways (57.1% of records). Roadsides are recorded at 11% in lowlands and 21.4% in mountains. From 2001 to 2022, habitat preferences changed significantly. In the lowlands, human settlements and arable fields represent only 34.2% of recorded habitats, while the proportion of roadsides increased to 47.8%. A similar increase in roadside preference also occurred in the mountains (52.8% of records).
From the point of view of the entire period (1949–2022), the difference in cumulative numbers of records or grids (Fig. 3) between roadsides, human settlements, and arable fields is not significant (Table 2), indicating the similar preference of common ragweed for these three habitats. In the lowlands, arable fields and human settlements provide the most dominant habitats for common ragweed, while in the mountains, the roadsides are a crucial vector of expansion. However, it has also begun to occur in grasslands or along forest paths.

Cumulative number of grid cells occupied by common ragweed in Slovakia until 2022, divided into nine different habitats
3.2 Potential distribution of common ragweed in Slovakia
According to the potential distribution predicted for the past period (1970–2000) (Fig. 4), common ragweed could commonly occur in lowlands. It is noticeable that 87.8% of records from 1949 to 2000 occurred in 30-second grid cells with probability values from 0.26 to 0.50 typical for common distribution, 5.7% of records belong to occasional (probability 0.11–0.25), and 4.5% of records occurred in 30-second grid cells with probability higher than 0.51, i.e., with abundant occurrence.

Potential distribution of common ragweed in Slovakia in the past period (1970–2000); abundant (> 0.51), common (0.26–0.50), occasional (0.11–0.25), rare or absent (< 0.10)
The prediction for current climate (2018–2021) based on the first model trained on records from 1949 to 2000 differs from the previous predicted distribution in the increased probability of occasional occurrence (0.11–0.25) in mountain regions (Fig. 5). This prediction is similar to the prediction based on the second model trained on records from 1949 to 2022 (Fig. 6) in the potential area of common ragweed occurrence. There is a very strong relationship between these two predictions (r = 0.868, p < 0.001). The differences in the prediction based on the second model consist of higher probability values in lowlands and an increased probability of occasional occurrences in mountains. According to this prediction, 6.7% of common ragweed records from 1949 to 2022 occur in 30-second grid cells with probability values for common occurrence (0.26–0.50), and 91.7% of records were collected from 30-second grid cells with a probability higher than 0.51 (abundant occurrence). The fourth predicted distribution of common ragweed in Slovakia for 2040–2061 (Fig. 7) is largely identical to the third model. It predicts an ever-increasing probability of the occasional occurrence of common ragweed in the Carpathian Mountains, indicating an increase in the number of records in these areas in the future.

Potential distribution of common ragweed in Slovakia in the current period (2018–2021) based on the distribution in 1949–2000; abundant (> 0.51), common (0.26–0.50), occasional (0.11–0.25), rare or absent (< 0.10)

Potential distribution of common ragweed in Slovakia in the current period (2018–2021) based on the distribution in 1949–2022; abundant (> 0.51), common (0.26–0.50), occasional (0.11–0.25), rare or absent (< 0.10)

Potential distribution of common ragweed in Slovakia in 2040–2061; abundant (> 0.51), common (0.26–0.50), occasional (0.11–0.25), rare or absent (< 0.10)
3.3 Environmental factors influencing the spread of common ragweed
The results of the conditional inference tree (Fig. 8) indicate the several main environmental or geographical factors that connect the vast majority of all records: mean annual air temperature > 9.51 °C, altitude < 217 m above mean sea level, and early September frost probability < 3.3%. Nevertheless, common ragweed is able to survive at altitudes higher than 500 m above mean sea level (4.8% of all records) and at a temperature lower than the average temperature in the Carpathians (7.8 °C). In such cases, microclimatic conditions are important, e.g., southeast- to southwest-facing slopes exposed to the sun without shading vegetation. Anthropogenization is also important, an increase in anthropogenic elements such as roads, railways, or settlements increases the probability of ragweed occurrence, albeit casual, even at higher altitudes. The relationships between environmental variables and the estimated probability of common ragweed occurrence for the current period are shown in Supplementary Information.

Conditional inference tree presenting the efficiency rates of the main factors shaping the common ragweed distribution. The bars show the likelihood ratio of the presence (green) and absence (orange) of common ragweed under the conditions specified in ovals. The mean annual air temperature for the current period (2018–2021) is used
Out of seventeen tested variables, only early September frost probability and ruggedness index drive the separation between Pannonian and Carpathian populations in linear discriminant analysis (Fig. 9), with an estimated accuracy of 0.91.

Ordination plot of linear discriminant analysis, illustrating two different ecotypes of common ragweed and transient populations. The LD1 and LD2 axes correlate with the ruggedness index and the early September frost probability
4 Discussion
4.1 Distribution and habitat preference
Comparing the neighbouring Central European countries, we can see differences in the current distribution of common ragweed. In Austria, where mountain landscape prevails, common ragweed occupies only 22.5% of the territory (Mang et al. 2018), which is twice less than in Slovakia with 44% occurrence. Lowlands in the southeast and east of Austria, belonging to the Pannonian Basin, are the most affected by ragweed invasion (Essl et al. 2009). A similar situation can be observed in Hungary, whose territory is entirely part of the Pannonian Basin and thus has a lowland character. Hungary is invaded at level 66.4% (Mang et al. 2018), which is almost comparable to 75.4% in the Pannonian zone of Slovakia. In Central Europe, Hungary (Mang et al. 2018) and Slovakia seem to be the most affected by common ragweed invasion.
The spread of ragweed in the Pannonian basin continues also in the territory of Slovenia (Mang et al. 2018; Dolenc et al. 2021) and further on in southern and eastern European countries (Song and Prots 1998; Konstantinović et al. 2005; Galzina et al. 2010; Janjić et al. 2011; Cunze et al. 2013; Bašić et al. 2017; Afonin et al. 2018; Sirbu et al. 2022). These regions are climatically very similar to the Pannonian basin in Slovakia (Fick and Hijmans 2017), however, there is no study, which would indicate an equal level of infestation as it is in Slovakia and Hungary. The analysis of the entire Pannonian basin as a geomorphological unit would be a useful addition to analyses of ragweed distribution in individual states, as the Pannonian region can be considered a regional source of the spread of ragweed diaspores (Šikoparija et al. 2009; Karrer et al. 2012; Ferus et al. 2015).
In contrast to Slovakia, Austria, and Hungary, common ragweed is known only from 6% of the territory of the Czech Republic (Skálová et al. 2017), which is analogous to the distribution in Germany (Mang et al. 2018). The climatic conditions of the Czech Republic (annual mean temperature, annual precipitation totals) are at an intermediate level between the Carpathian Mountains and the Pannonian basin in Slovakia (Fick and Hijmans 2017). A threefold difference in the occupation of the grid cells may be caused by the absence of vast lowlands in the Czech Republic, which would create a spreading pressure on the surrounding mountain basins as it does in Slovakia, where 52–72% of the records from the Carpathian Mountains are distant within 20–40 km from Pannonian lowlands. This supports the assumption in the previous paragraph that the Carpathians are being persistently invaded due to their proximity to the Pannonian basin since the common ragweed expansion is estimated at a rate of between 6 and 20 km per/year (Makra et al. 2015).
In general, many studies have confirmed the binding of common ragweed to man-made or human-impacted habitats, such as arable land, settlements, transportation networks, etc. (e.g., Walter et al. 2005; Fumanal et al. 2008; Zelnik 2012; Ciapetta et al. 2016; Lemke et al. 2021; Zhao et al. 2022; Simard and Maheux 2022; Glaser et al. 2022) and Slovak populations are no exception. At the regional level, we observed differences in habitat preference. In lowlands with intensive land use, common ragweed is widespread in human settlements and arable fields, which is very similar to the situation in Hungary (Pinke et al. 2011). In the early years of expansion into the Slovak part of the Carpathian Mountains, railways were the typical habitat of common ragweed, but later the preference for roadsides both in mountains and lowlands increased. An identical course was observed in Austria (Essl et al. 2009) and the Czech Republic (Skálová et al. 2017), only the time interval in the change of habitat preference is different, the spread along roadsides intensified 40 years earlier in Austria than in Slovakia and the Czech Republic. It may be related to progress in infrastructure development and traffic volume (Lavoie et al. 2007) since it has a significant effect on the dispersal of common ragweed seeds (Lemke et al. 2019). A highly disturbed upper soil layer along the roadsides becomes a suitable niche for invasive species (Joly et al. 2011; Meunier and Lavoie 2012; Follak et al. 2018; Szumańska et al. 2021), which may start to prefer this habitat after a small change in environmental conditions (Deeley and Petrovskaya 2022).
Not only roadsides are potential habitats for common ragweed in mountains or lowlands. In native area (North America), common ragweed shifted from primary habitats, which are probably the prairies, to synanthropic environments (Basset and Crompton 1975; Montagnani et al. 2017). In last 4,500 years, the prairies have been regularly disturbed by fire cycles (Brown et al. 2005), so ragweed is adapted to regular habitat disturbance. Disturbances are the keys to opening a room for common ragweed to meadow phytocoenoses at altitudes higher than 500 m above sea level with the help of strong anthropogenic pressure from arable lands, roads, and paths (Chadaeva et al. 2018; Kröel-Dulay et al. 2019). One of the vectors of spread to these habitats is seeds mixed with feed. Wildlife supplementary feeding could become a source of seeds (Strgulc Krajšek and Novak 2013; Kochjarová et al. 2023) and increase dispersal along forest paths and grasslands, thus aiding the spread of invasive species (Mortensen et al. 2009; Sărăţeanu et al. 2010). But such habitats will probably never be as frequently invaded as paved roads (Joly et al. 2011) or arable fields (Chadaeva et al. 2018), which is also indicated by the low percentage representation of our records in these habitats.
4.2 Predictions and environmental factors
The species distribution models can reveal the main environmental factors, habitats, and ways through which invasive species are spread (Coutts et al. 2011). Knowledge of the potential distribution of such species may help prevent their further spread (Szumańska et al. 2021). For that reason, the distribution modelling of common ragweed was calculated for many countries across the world (e.g., Essl et al. 2009; Pinke et al. 2011; Storkey et al. 2014; Skálová et al. 2017; Case and Stinson 2018; Wang et al. 2021; Cho et al. 2022; Chung et al. 2022; Qin et al. 2022). In our case, at the interface of two different environments, we used higher precision predictions. All our species distribution models coincide in the potential area of common ragweed occurrence in the whole Pannonian region and in the basins and warmer valleys of the Carpathian Mountains. However, predictions based on these models differ in the probability of occasional to rare occurrences of common ragweed in mountains and common to abundant occurrences in lowlands. This demonstrates the differences in models trained on records from the colonization period and on records when invasive species are adapting to new local conditions or have reached a quasi-equilibrium. Invasive species are not in equilibrium with climate (Foster et al. 2022; Formoso-Freire et al. 2023). If invasive species colonizing the potential area have not yet occupied all suitable habitats, then not all of the created pseudo-absences indicate unsuitable environmental conditions for the species, and SDM is biased (Chefaoui and Lobo 2008). In addition, individual populations of the same species may be at various stages of invasion in the study area (Gallien et al. 2012). Our second model suggests that Pannonian populations have reached a quasi-equilibrium, but Carpathian populations are only at the stage of colonization. Although, judging by current records, common ragweed does not occupy all suitable niches, there are probably many localities that have not yet been recorded. In general, according to studies in Austria, records reflect only 42–65% of the real distribution (Mang et al. 2017, 2018). Therefore, according to the prediction for the current period (Fig. 6), areas with a higher probability of occurrence are definitely more invaded, as indicated by number of our records. Under a future climate scenario, common ragweed was predicted to have a limited potential distribution expansion in mountains. This moderate trend is similar to forecasts from Europe (Storkey et al. 2014), Asia (Chung et al. 2022; Qin et al. 2022), and the eastern United States (Case and Stinson 2018).
The air temperature and increased rates of anthropogenization seem to be important factors for common ragweed distribution not only in Slovakia but also in other Central European countries (Essl et al. 2009; Skálová et al. 2017; Lemke et al. 2021). Low precipitation totals during the vegetation period can represent another limiting factor for the invasion process (Chauvel et al. 2006; Pinke et al. 2011, 2019; Wang et al. 2021; Cho et al. 2022), but in our study area, this limitation does not seem to be important. On the contrary, high precipitation totals combined with cooler weather are as much a limiting factor as low totals and a dry warm climate. Closely related to this factor are the wet soils along rivers and streams, which often contribute to the spread of ragweed (Fumanal et al. 2007; Lavoie et al. 2007; Cho et al. 2022). Although we have recorded only a few populations from these habitats, in the first years of the invasion, it was a relatively common habitat in Slovakia as well. The other factors we investigated have only a local impact on the spread of ragweed.
4.3 Climatic change impact on common ragweed expansion
The presumption that the expansion of ragweed is influenced by global warming has already been developed by many authors (e.g., Essl et al. 2009; Cunze et al. 2013; Rasmussen et al. 2017; Mang et al. 2018; Lemke et al. 2021; Liu et al. 2021). The history of the common ragweed invasion is similar in many countries around the world. Although ragweed was already introduced to various countries at the turn of the 19th and 20th centuries (Rich 1994; Pyšek et al. 2002; Chauvel et al. 2006; Csontos 2010; Tokarska-Guzik et al. 2011), and according to Essl et al. (2009) was in a lag phase, for which adaptation processes are typical (Aikio et al. 2010) and which precedes plant invasions (Larkin 2012), common ragweed in Slovakia did not need a long lag phase for the invasion. It was introduced to Slovakia around 1949 and it immediately began to spread (Jehlík 1998). The curve of cumulative records has a similar course in Slovakia (Hrabovský et al. 2016), Austria (Essl et al. 2009), the Czech Republic (Skálová et al. 2017), and even China (Liu et al. 2021). The reasons for the first common ragweed expansion may be post-war industrial development, and also climatic change. The double S shape of the curve of cumulative records in all countries indicates the second wave of invasion around the year 2000, which continues to this day and again probably reflects climatic change impact.
Based on our distribution models, climatic change seems to be responsible for common ragweed expansion. However, to a small extent, the formation of a new ecotype, which adapts to a shorter life period to avoid early autumn frosts (Fig. 9), can also have an impact on the expansion. Common ragweed is able to shorten its life cycle due to environmental factors and produce seeds of adapted genotypes (Hodgins and Rieseberg 2011; Scalone et al. 2016). An example of such adaptation can be a reduction in genome sizes, causing faster cell division and a shortening of the life cycle (Bennett 1972; Leitch and Bennett 2007). Similar adaptations can partially influence the predicted distribution model for 2040–2061 and common ragweed will more frequently occur at higher altitudes, which will have a negative effect, especially for allergy sufferers.
5 Conclusions
According to our research, common ragweed uses the most optimal conditions for its spread in different times. When highways weren’t being developed and traffic density was lower, agricultural land became the source of its spread. Later, the development of the infrastructure contributed to its expansion to other regions of Slovakia. However, the rise in temperature in mountainous areas, where it would not ordinarily be able to flourish, also contributed to its spread. The prolonged summer days and the continual introduction of seeds from the lowlands allowed populations in the mountain basins to be selected that could produce viable seeds before early autumn frosts. Today, common ragweed can be found more frequently not just in lowlands and mountain basins as a result of climate change. Future records of common ragweed are expected to become increasingly frequent in the Carpathians and it probably became abundant in many sites of the Pannonian lowlands.
Data availability
No datasets were generated or analysed during the current study.
References
Afonin AN, Luneva NN, Fedorova YA, Kletchkovskiy YuE, Chebanovskaya AF (2018) History of introduction and distribution of common ragweed (Ambrosia artemisiifolia L.) in the European part of the Russian Federation and in the Ukraine. EPPO Bull 48:266–273. https://doi.org/10.1111/epp.12484
Aikio S, Duncan RP, Hulme PE (2010) Lag-phases in alien plant invasions: separating the facts from the artefacts. Oikos 119:370–378. https://doi.org/10.1111/j.1600-0706.2009.17963.x
Allard HA (1943) The North American Ragweeds and their ocurrence in other parts of the World. Science 98:292–294. https://doi.org/10.1126/science.98.2544.292
Barbet-Massin M, Jiguet F, Albert CH, Thuiller W (2012) Selecting pseudo-absences for species distribution models: how, where and how many? Methods Ecol Evol 3:327–338. https://doi.org/10.1111/j.2041-210X.2011.00172.x
Bašić F, Đikić M, Gadžo D (2017) Appearance and spreading of common ragweed (Ambrosia artemisiifolia L.) in Bosnia and Herzegovina. Folia Biol Geol 58:147–155. https://doi.org/10.3986/fbg0032
Baskin JM, Baskin CC (1987) Temperature requirements for after-ripening in buried seeds of four summer annual weeds. Weed Res 27:385–389. https://doi.org/10.1111/j.1365-3180.1987.tb00776.x
Basset IJ, Crompton CW (1975) The biology of Canadian weeds. 11. Ambrosia artemisiifolia L. and A. psilostachya DC. Can J Plant Sci 55:463–476. https://doi.org/10.4141/cjps75-072
Bazzaz FA (1970) Secondary dormancy in the seeds of the common ragweed, Ambrosia artemisiifolia. Bull Torrey Bot Club 97:302–305. https://doi.org/10.2307/2483650
Bennett MD (1972) Nuclear DNA content and minimum generation time in herbaceous plants. Proc R Soc Lond B 181:109–135. https://doi.org/10.1098/rspb.1972.0042
Brandes D, Nitzsche J (2006) Biology, introduction, dispersal, and distribution of common ragweed (Ambrosia artemisiifolia L.) with special regard to Germany. Nachrichtenbl Deut Pflanzenschutzd 58:286–291
Brown KJ, Clark JS, Grimm EC, Donovan JJ, Mueller PG, Hansen BCS, Stefanova I (2005) Fire cycles in North American Interior grasslands and their relation to prairie drought. Proc Natl Acad Sci 102:8865–8870. https://doi.org/10.1073/pnas.0503621102
Case MJ, Stinson KA (2018) Climate change impacts on the distribution of the allergenic plant, common ragweed (Ambrosia artemisiifolia) in the eastern United States. PLoS ONE 13:e0205677. https://doi.org/10.1371/journal.pone.0205677
Chadaeva VA, Shhagapsoeva KA, Tsepkova NL, Shhagapsoev SH (2018) Monitoring of Ambrosia artemisiifolia L. distribution in Meadow Phytocenoses of Kabardino-Balkarian Republic (Central Caucasus). Russ J Biol Invasions 9:195–203. https://doi.org/10.1134/S2075111718020030
Chauvel B, Dessaint F, Cardinal-Legrand C, Bretagnolle F (2006) The historical spread of Ambrosia artemisiifolia L. in France from herbarium records. J Biogeogr 33:665–673. https://doi.org/10.1111/j.1365-2699.2005.01401.x
Chefaoui RM, Lobo JM (2008) Assessing the effects of pseudo-absences on predictive distribution model performance. Ecol Model 210:478–486. https://doi.org/10.1016/j.ecolmodel.2007.08.010
Cho KH, Park J-S, Kim JH, Kwon YS, Lee D-H (2022) Modeling the distribution of invasive species (Ambrosia spp.) using regression kriging and maxent. Front Ecol Evol 10:1036816. https://doi.org/10.3389/fevo.2022.1036816
Chung HI, Choi Y, Yoo Y, Engler R, Lee K, Jeon SW (2022) Integrated spatial model based evaluation methodology for optimal invasive species management: common ragweed in the Republic of Korea. Environ Res Lett 17:034047. https://doi.org/10.1088/1748-9326/ac4dc7
Ciapetta S, Ghiani A, Girardelli F, Bonini M, Citterio S, Gentili R (2016) Invasion of Ambrosia artemisiifolia in Italy: Assessment via analysis of genetic variability and herbarium data. Flora 223:106–113. https://doi.org/10.1016/j.flora.2016.05.002
Coutts SR, van Klinken RD, Yokomizo H, Buckley YM (2011) What are the key drivers of spread in invasive plants: dispersal, demography or landscape: and how can we use this knowledge to aid management? Biol Invasions 13:1649–1661. https://doi.org/10.1007/s10530-010-9922-5
Csontos P, Vitalos M, Barina Z, Kiss L (2010) Early distribution and spread of Ambrosia artemisiifolia in Central and Eastern Europe. Bot Helv 120:75–78. https://doi.org/10.1007/s00035-010-0072-2
Cunze S, Leiblein MC, Tackenberg O (2013) Range Expansion of Ambrosia artemisiifolia in Europe is promoted by Climate Change. Int Sch Res Not 610126. https://doi.org/10.1155/2013/610126
Deeley B, Petrovskaya N (2022) Propagation of invasive plant species in the presence of a road. J Theor Biol 548:111196. https://doi.org/10.1016/j.jtbi.2022.111196
Deen W, Hunt T, Swanton CJ (1998) Influence of temperature, photoperiod and irradiance on phenological development of common ragweed (Ambrosia artemisiifolia). Weed Sci 46:555–560. https://doi.org/10.1017/S0043174500091098
Dickerson CT (1968) Studies on the germination, growth, development and control of common ragweed (Ambrosia artemisiifolia L.). Dissertation, Cornell University
Dolenc A, Papež Kristanc A, Rozman S (2021) Managing the residues of invasive alien plants [in Slovenian ravnanje z ostanki invazivnih tujerodnih rastlin]. Varstvo Narave 32:5–28
Dormann CF, Elith J, Bacher S, Buchmann C, Carl G, Carré G, Marquéz JRG, Gruber B, Lafourcade B, Leitão PJ, Münkemüller T (2013) Collinearity: a review of methods to deal with it and a simulation study evaluating their performance. Ecography 36:27–46. https://doi.org/10.1111/j.1600-0587.2012.07348.x
Döscher R, Acosta M, Alessandri A, Anthoni P, Arsouze T, Bergman T, Bernardello R, Boussetta S, Caron L-P, Carver G, Castrillo M, Catalano F, Cvijanovic I, Davini P, Dekker E, Doblas-Reyes FJ, Docquier D, Echevarria P, Fladrich U, Fuentes-Franco R, Gröger M, von Hardenberg J, Hieronymus J, Karami MP, Keskinen J-P, Koenigk T, Makkonen R, Massonnet F, Ménégoz M, Miller PA, Moreno-Chamarro E, Nieradzik L, van Noije T, Nolan P, O’Donnell D, van den Ollinaho P, Ortega P, Prims OT, Ramos A, Reerink T, Rousset C, Ruprich-Robert Y, Le Sager P, Schmith T, Schrödner R, Serva F, Sicardi V, Sloth Madsen M, Smith B, Tian T, Tourigny E, Uotila P, Vancoppenolle M, Wang S, Wårlind D, Willén U, Wyser K, Yang S, Yepes-Arbós X, Zhang Q (2022) The EC-Earth3 Earth system model for the coupled model Intercomparison Project 6. Geosci Model Dev 15:2973–3020. https://doi.org/10.5194/gmd-15-2973-2022
Eliáš Pst (2021) K výskytu synantropných rastlín v Starých Horách a v blízkom okolí. [On the occurrence of synanthropic plants in the Staré Hory village and in its surroundings (Central Slovakia)]. Bull Slov Bot Spoločn 43:175–195
Eliáš P (2022) Zaujímavejšie floristické nálezy. [Interesting Floristic Find ] Bull Slov Bot Spoločn 44:242–256
Essl F, Dullinger S, Kleinbauer I (2009) Changes in the spatio-temporal patterns and habitat preferences of Ambrosia artemisiifolia during its invasion of Austria. Preslia 81:119–133
Essl F, Biró K, Brandes D, Broennimann O, Bullock JM, Chapman DS, Chauvel B, Dullinger S, Fumanal B, Guisan A, Karrer G, Kazinczi G, Kueffer C, Laitung B, Lavoie C, Leitner M, Mang T, Moser D, Müller-Schärer H, Petitpierre B, Richner R, Schaffner U, Smith M, Starfinter U, Vautard R, Vogl G, von der Lippe M, Follak S (2015) Biological Flora of the British Isles: Ambrosia artemisiifolia. J Ecol 103:1069–1098. https://doi.org/10.1111/1365-2745.12424
Farooq S, Onen H, Ozaslan C, Baskin CC, Gunal H (2019) Seed germination niche for common ragweed (Ambrosia artemisiifolia L.) populations naturalized in Turkey. S Afr J Bot 123:361–371. https://doi.org/10.1016/j.sajb.2019.03.031
Ferus P, Sirbu C, Eliáš P, Konôpková J, Ďurišová Ľ, Samuil C, Oprea A (2015) Reciprocal contamination by invasive plants: analysis of trade exchange between Slovakia and Romania. Biologia 70:893–904. https://doi.org/10.1515/biolog-2015-0102
Fick SE, Hijmans RJ (2017) WorldClim 2: new 1-km spatial resolution climate surfaces for global land areas. Int J Climatol 37:4302–4315. https://doi.org/10.1002/joc.5086
Follak S, Schleicher C, Schwarz M (2018) Roads support the spread of invasive Asclepias syriaca in Austria. Bodenkult: J Land Manag Food Environ 69:257–265. https://doi.org/10.2478/boku-2018-0022
Formoso-Freire V, Barbosa AM, Baselga A, Gómez-Rodríguez C (2023) Predicting the spatio-temporal pattern of range expansion under lack of equilibrium with climate. Biol Conserv 288:110361. https://doi.org/10.1016/j.biocon.2023.110361
Foster SL, Kharouba HM, Smith TW (2022) Testing the assumption of environmental equilibrium in an invasive plant species over a 130 year history. Ecography 12:e12933. https://doi.org/10.1111/ecog.06284
Fox J, Friendly GG, Graves S, Heiberger R, Monette G, Nilsson H, Ripley B, Weisberg S, Fox MJ, Suggests MASS (2007) The car package. R Foundation for Statistical computing, 1109: 1431
Fumanal B, Chauvel B, Sabatier A, Bretagnolle F (2007) Variability and cryptic heteromorphism of Ambrosia artemisiifolia seeds: what consequences for its invasion in France? Ann Bot 100:305–313. https://doi.org/10.1093/aob/mcm108
Fumanal B, Girod C, Fried G, Bretagnolle F, Chauvel B (2008) Can the large ecological amplitude of Ambrosia artemisiifolia explain its invasive success in France? Weed Res 48:349–359. https://doi.org/10.1111/j.1365-3180.2008.00627.x
Futák J (1984) Fytogeografické členenie Slovenska. In: Bertová L (ed) Flóra Slovenska IV/1. Veda, Bratislava, pp 418–420
Gallien L, Douzet R, Pratte S, Zimmermann NE, Thuiller W (2012) Invasive species distribution models – how violating the equilibrium assumption can create new insights. Glob Ecol Biogeogr 21:1126–1136. https://doi.org/10.1111/j.1466-8238.2012.00768.x
Galzina N, Barić K, Šćepanović M, Goršić M, Ostojić Z (2010) Distribution of Invasive Weed Ambrosia artemisiifolia L. in Croatia. Agric Conspec Sci 75:75–81
Glaser M, Berg C, Buldrini F, Buholzer S, Bürger J, Chiarucci A, Chytrý M, Dřevojan P, Follak S, Küzmič F, Lososová Z, Meyer S, Moser D, Pyšek P, Richner N, Šilc U, Wietzke A, Dullinger S, Essl F (2022) AgriWeedClim database: a repository of vegetation plot data from central European arable habitats over 100 years. Appl Veg Sci 25:e12675. https://doi.org/10.1111/avsc.12675
Hastie T, Tibshirani R, Friedman JH (2001) The elements of statistical learning: data mining, inference, and prediction. Springer, New York. https://doi.org/10.1007/978-0-387-21606-5
Hodgins KA, Rieseberg L (2011) Genetic differentiation in life-history traits of introduced and native common ragweed (Ambrosia artemisiifolia) populations. J Evol Biol 24:2731–2749. https://doi.org/10.1111/j.1420-9101.2011.02404.x
Hothorn T, Hornik K, Zeileis A (2015) ctree: Conditional inference trees. The comprehensive R archive network 8
Hrabovský M (2023) The simplified tools for central European vegetation mapping. Acta Bot Univ Comen 61:11–13
Hrabovský M, Mičieta K (2014) The occurrence of an invasive species Ambrosia artemisiifolia in Slovakia in the years 2008–2014. Acta Bot Univ Comen 49:9–12
Hrabovský M, Mičieta K (2018) New findings of the common ragweed (Ambrosia artemisiifolia) in Slovakia in the year 2017. Acta Bot Univ Comen 53:25–27
Hrabovský M, Ščevková J, Mičieta K, Lafférsová J, Dušička J (2016) Expansion and aerobiology of Ambrosia artemisiifolia L. in Slovakia. Ann Agric Environ Med 23:64–70. https://doi.org/10.5604/12321966.1196854
Imdadullah M, Aslam M, Altaf S (2016) Mctest: an R Package for detection of collinearity among regressors. R J 8:495–505
Janjić V, Radivojević L, Jovanović V (2011) Common ragweed (Ambrosia artemisiifolia L.): a harmful weed, ruderal and allergenic plant in the territory of Belgrade. Acta Herbologica 20:57–66
Jehlík V (1998) Cizí expanzivní Plevele České republiky a Slovenské republiky. Academia, Praha
Joly M, Bertrand P, Gbangou RY, White M-C, Dubé J, Lavoie C (2011) Paving the way for Invasive species: Road Type and the spread of common ragweed (Ambrosia artemisiifolia). Environ Manag 48:514–522. https://doi.org/10.1007/s00267-011-9711-7
Karrer G, Kropf M, Huppenberger A, Leitsch-Vitalos M Migration routes of Ambrosia artemisiifolia throughout Austria and neigbouring countries indicated by DNA microsatellite analyses. Neobiota 12, 7th European Conference on Biological, Invasions (2012) Pontevedra (Spain), 12–14. September 2012
Kazinczi G, Béres I, Novák R, Bíró K, Pathy Z (2008) Common ragweed (Ambrosia artemisiifolia): a review with special regards to the results in Hungary. Taxonomy, origin and distribution, morphology, live cycle and reproduction strategy. Herbologia 9:55–91
Kochjarová J, Blanár D, Jarolímek I, Slezák M (2023) Wildlife supplementary feeding facilitates spread of alien plants in forested mountainous areas: a case study from the western carpathians. Biologia 78:1381–1389. https://doi.org/10.1007/s11756-023-01339-0
Konstantinović B, Meseldžija M, Konstantinović B, Marisavljević D (2005) Determination of the occurrence and spread of the allergenic weed Ambrosia artemisiifolia in the territory of Vojvodina (Serbia). Plant protection and plant health in Europe: introduction and spread of invasive species, 9–11 June 2005, Humboldt University, Berlin
Kröel-Dulay G, Csecserits A, Szitár K, Molnár E, Szabó R, Ónodi G, Botta-Dukát Z (2019) The potential of common ragweed for further spread: invasibility of different habitats and the role of disturbances and propagule pressure. Biol Invasions 21:137–149. https://doi.org/10.1007/s10530-018-1811-3
Lambdon PW, Pyšek P, Basnou C, Hejda M, Arianoutson M, Essel F (2008) Alien flora of Europe: species diversity, temporal trends, geographical patterns and research needs. Preslia 80:101–149
Larkin DJ (2012) Lengths and correlates of lag phases in Upper-Midwest plant invasions. Biol Invasions 14:827–838. https://doi.org/10.1007/s10530-011-0119-3
Lavoie C, Jodoin Y, Goursaud de Merlis A (2007) How did common ragweed (Ambrosia artemisiifolia L.) spread in Quebec? A historical analysis using herbarium records. – J Biogeogr 34:1751–1761. https://doi.org/10.1111/j.1365-2699.2007.01730.x
Leitch IJ, Bennett MD (2007) Genome size and its uses: the impact of Flow Cytometry. In: Doležel J, Greilhuber J, Suda J (eds) Flow Cytometry with Plant cells: analysis of genes, chromosomes and genomes. Wiley-VCH, Weinheim, pp 153–176. https://doi.org/10.1002/9783527610921.ch7
Lemke A, Kowarik I, von der Lippe M (2019) How traffic facilitates population expansion of invasive species along roads: the case of common ragweed in Germany. J Appl Ecol 56:413–422. https://doi.org/10.1111/1365-2664.13287
Lemke A, Buchholz S, Kowarik I, Starfinger U, von der Lippe M (2021) Interaction of traffic intensity and habitat features shape invasion dynamics of an invasive alien species (Ambrosia artemisiifolia) in a regional road network. NeoBiota 64:155–175. https://doi.org/10.3897/neobiota.64.58775
Li S, Dongchang G, Guangqing G (1989) A study on the phenology of common ragweed and great ragweed. J Shenyang Agric Univ 20:344–350
Liu X-L, Li H-Q, Wang J-H, Sun X-P, Fu Y-Y, Xing L-G (2021) The current and future potential geographical distribution of common ragweed, Ambrosia artemisiifolia in China. Pak J Bot 53:167–172. https://doi.org/10.30848/PJB2021-1(18)
Májeková J, Jarolímek I, Zaliberová M, Medvecká J (2021) Alien (invasive) vascular plants in Slovakia – a story of successful plant immigrants. Environ Socio-econ Stud 9:23–31. https://doi.org/10.2478/environ-2021-0022
Makra L, Matyasovszky I, Hufnagel L, Tusnády G (2015) The history of ragweed in the world. Appl Ecol Environ Res 13:489–512. https://doi.org/10.15666/aeer/1302_489512
Mang T, Essl F, Moser D, Karrer G, Kleinbauer I, Dullinger S (2017) Accounting for imperfect observation and estimating true species distributions in modelling biological invasions. Ecography 40:1187–1197. https://doi.org/10.1111/ecog.02194
Mang T, Essl F, Moser D, Dullinger S (2018) Climate warming drives invasion of Ambrosia artemisiifolia in central Europe. Preslia 90:59–81. https://doi.org/10.23855/preslia.2018.059
Medvecká J, Kliment J, Májeková J, Halada Ľ, Zaliberová M, Gojdičová E, Feráková V, Jarolímek I (2012) Inventory of the alien flora of Slovakia. Preslia 84:257–309
Metzger MJ, Bunce RGH, Jongman RHG, Mücher CA, Watkins JW (2005) A climatic stratification of the environment of Europe. Glob Ecol Biogeogr 14:549–563. https://doi.org/10.1111/j.1466-822x.2005.00190.x
Meunier G, Lavoie C (2012) Roads as corridors for Invasive Plant species: New evidence from smooth bedstraw (Galium mollugo). Invasive Plant Sci Manag 5:92–100. https://doi.org/10.1614/IPSM-D-11-00049.1
Montagnani C, Gentili R, Smith M, Guarino MF, Citterio S (2017) The Worldwide Spread, Success, and impact of ragweed (Ambrosia spp). Crit Rev Plant Sci 36:139–178. https://doi.org/10.1080/07352689.2017.1360112
Mortensen D, Rauschert E, Nord A, Jones B (2009) Forest roads facilitate the spread of invasive plants. Invasive Plant Sci Manag 2:191–199. https://doi.org/10.1614/IPSM-08-125.1
Naimi B, Araújo MB (2016) sdm: a reproducible and extensible R platform for species distribution modelling Ecography 39:368–375. https://doi.org/10.1111/ecog.01881
Pinke G, Karácsony P, Czúcz B, Botta-Dukát Z (2011) Environmental and land-use variables determining the abundance of Ambrosia artemisiifolia in arable fields in Hungary. Preslia 83:219–235
Pinke G, Kolejanisz T, Vér A, Nagy K, Milics G, Schlögl G, Bede-Fazekas Á, Botta-Dukát Z, Czúcz B (2019) Drivers of Ambrosia artemisiifolia abundance in arable fields along the austrian-hungarian border. Preslia 91:369–389. https://doi.org/10.23855/preslia.2019.369
Pyšek P, Sádlo J, Mandák B (2002) Catalogue of alien plants of the Czech Republic. Preslia 74:97–186
Qin Z, Zhang JE, DiTommaso A, Diez JM, Zhao Y, Wang FG (2022) Predicting the potential distribution of three allergenic invasive Ambrosia (ragweed) species in Asia. J Environ Inf 39:49–66. https://doi.org/10.3808/jei.202000444
Rasmussen K, Thyrring J, Muscarella R, Borchsenius F (2017) Climate-change-induced range shifts of three allergenic ragweeds (Ambrosia L.) in Europe and their potential impact on human health. PeerJ 5:e3104. https://doi.org/10.7717/peerj.3104
Rich TCG (1994) Ragweeds (Ambrosia L.) in Britain. Grana 33:38–43. https://doi.org/10.1080/00173139409427454
Ripley B, Venables B, Bates DM, Hornik K, Gebhardt A, Firth D, Ripley MB (2013) Package ‘mass’. Cran r 538:113–120
Sanchez R, Mackenzie SA (2016) Genome-wide discriminatory information patterns of cytosine DNA methylation. Int J Mol Sci 17:938. https://doi.org/10.3390/ijms17060938
Sărăţeanu V, Moisuc A, Cotuna O (2010) Ambrosia artemisiifolia L. an invasive weed from ruderal areas to disturbed grasslands. Lucrări Ştiinţifice 53:235–238
Scalone R, Lemke A, Štefanić E, Kolseth A-K, Rašić S, Andersson L (2016) Phenological variation in Ambrosia artemisiifolia L. facilitates Near Future Establishment at Northern latitudes. PLoS ONE 11:e0166510. https://doi.org/10.1371/journal.pone.0166510
Šikoparija B, Smith M, Skjøth CA, Radišić P, Milkovska S, Šimić S, Brandt J (2009) The pannonian plain as a source of Ambrosia pollen in the Balkans. Int J Biometeorol 53:263–272. https://doi.org/10.1007/s00484-009-0212-9
Simard M-J, Maheux L (2022) Roadsides and neighboring field edges harbor different weed compositions. Front Agron 4:1005093. https://doi.org/10.3389/fagro.2022.1005093
Sirbu C, Miu IV, Gavrilidis AA, Gradinaru SR, Niculae IM, Preda C, Oprea A, Urziceanu M, Camen-Comanescu P, Nagoda E, Sirbu IM, Memedemin D, Anastasiu P (2022) Distribution and pathways of introduction of invasive alien plant species in Romania. NeoBiota 75:1–21. https://doi.org/10.3897/neobiota.75.84684
Skálová H, Guo W-Y, Wild J, Pyšek P (2017) Ambrosia artemisiifolia in the Czech Republic: history of invasion, current distribution and prediction of future spread. Preslia 89:1–16. https://doi.org/10.23855/preslia.2017.001
Smith M, Cecchi L, Skjøth CA, Karrer G, Šikoparija B (2013) Common ragweed: a threat to environmental health in Europe. Environ Int 61:115–126. https://doi.org/10.1016/j.envint.2013.08.005
Song J-S, Prots B (1998) Invasion of Ambrosia artemisiifolia l. (Compositae) in the Ukrainian carpathians mts. And the Transcarpathian Plain (Central Europe). Korean J Biol Sci 2:209–216. https://doi.org/10.1080/12265071.1998.9647409
Storkey J, Stratonovitch P, Chapman DS, Vidotto F, Semenov MA (2014) A process-based Approach to Predicting the Effect of Climate Change on the distribution of an invasive allergenic plant in Europe. PLoS ONE 9:e88156. https://doi.org/10.1371/journal.pone.0088156
Štrba P (2003) Najvyššie položená lokalita invázneho druhu Ambrosia artemisiifolia L. na Slovensku. [Maximum altitude of invasive species Ambrosia artemisiifolia L. Slovakia ] Bull Slov Bot Spoločn 25:155–156
Strgulc Krajšek S, Novak M (2013) Achenes of common ragweed (Ambrosia artemisiifolia) in packages of sunflower achenes for outdoor birds. Acta Biol Slov 56:3–9
Szumańska I, Lubińska-Mielińska S, Kamiński D, Rutkowski L, Nienartowicz A, Piernik A (2021) Invasive plant species distribution is structured by Soil and Habitat Type in the City Landscape. Plants 10:773. https://doi.org/10.3390/plants10040773
Tokarska-Guzik B, Bzdęga K, Koszela K, Żabińska I, Krzuś B, Sajan M, Sendek A (2011) Allergenic invasive plant Ambrosia artemisiifolia L. in Poland: threat and selected aspects of biology. Biodiv Res Conserv 21:39–48. https://doi.org/10.2478/v10119-011-0008-8
Walter J, Essl F, Englisch T, Kiehn M (2005) Neophytes in Austria: Habitat preferences and ecological effects. In: Nentwig W (ed): Biological Invasions – From Ecology to Control. Neobiota 6:13–25
Wang D-J, Wei H-Y, Zang X-H, Fang Y-Q, Gu W (2021) Habitat suitability modeling based on remote sensing to realize time synchronization of species and environmental variables. J Plant Ecol 14:241–256. https://doi.org/10.1093/jpe/rtaa092
Zaliberová M, Podroužková-Medvecká J (2011) Ambrosia artemisiifolia. In: Eliáš P (ed) Zaujímavejšie floristické nálezy. [Interesting floristic findings.] Bull Slov Bot Spoločn 33:103–109
Zelnik I (2012) The presence of invasive alien plant species in different habitats: case study from Slovenia. Acta Biol Slov 55:25–38
Zhao W, Liu T, Sun M, Wang H, Liu X, Su P (2022) Rapid monitoring of Ambrosia artemisiifolia in semi-arid regions based on ecological convergence and phylogenetic relationships. Front Ecol Evol 10:926990. https://doi.org/10.3389/fevo.2022.926990
Acknowledgements
The study was supported by the Operation Program of Integrated Infrastructure for the project, Advancing University Capacity and Competence in Research, Development and Innovation, ITMS2014+:313021 × 329, co-financed by the European Regional Development Fund and by the Operation Program of Integrated Infrastructure for the project, UpScale of Comenius University Capacities and Competence in Research, Development and Innovation, ITMS2014+: 313021BUZ3, co-financed by the European Regional Development Fund. We are grateful to reviewers for their inspirational ideas and recommendations.
Funding
This study was supported by Biodiversa + PAREUS.
Open access funding provided by The Ministry of Education, Science, Research and Sport of the Slovak Republic in cooperation with Centre for Scientific and Technical Information of the Slovak Republic
Author information
Authors and Affiliations
Contributions
M.H. collected and analyzed data, wrote and edited the main manuscript text and prepared all figures. S.K. edited the manuscript. R.K. supervised the data analysis. All authors reviewed the manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Hrabovský, M., Kubalová, S. & Kanka, R. The impact of changing climate on the spread of the widely expanding species Ambrosia artemisiifolia in Slovakia. Theor Appl Climatol (2024). https://doi.org/10.1007/s00704-024-05006-5
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s00704-024-05006-5